Antigenicity of key hepatitis C virus E1E2 glycoprotein neutralizing sites is genotype independent
Jessica L. Mimms, Ariadne Sinnis-Bourozikas, Nathaniel R. Felbinger, Nicole Frumento, Harry T. Paul, Arvind H. Patel, Zhenyong Keck, Steven K. H. Foung, Mansun Law, Richard A. Urbanowicz, Alexander W. Tarr, Jonathan K. Ball, Brian G. Pierce, Justin R. Bailey

TL;DR
This study shows that the antigenic properties of HCV's E1E2 glycoprotein are not determined by the virus's genotype, which is important for vaccine development.
Contribution
The study confirms that E1E2 antigenicity is genotype-independent using a diverse panel of HCV variants.
Findings
Neutralization results for gt 2–6 E1E2 variants were similar to those of a gt 1-predominant reference panel.
The antigenicity of E1E2 is not dictated by HCV genotype across genetically diverse variants.
The previously published panel of 15 HCVpp accurately represents neutralization across all HCV genotypes.
Abstract
The development of an effective prophylactic hepatitis C virus (HCV) vaccine is a priority to achieve global elimination of the virus. Accurate assessment of the neutralizing breadth of antibodies induced by vaccines and a clear understanding of the antigenic differences between viral variants included in vaccines are both critical for vaccine development. Prior studies have indicated that HCV genotypes (gts) do not dictate the sensitivity of HCV envelope glycoprotein (E1E2) variants to neutralizing antibodies. However, most of these prior studies under-sampled variants from gts 2–6. Here, we selected a genetically diverse and representative panel of gt 2–6 E1E2 variants, used them to generate HCV pseudoparticles (HCVpp), and measured neutralization of these HCVpp by neutralizing antibodies and HCV-immune plasma from persons infected with gt 1–6 viruses. We found that neutralization…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
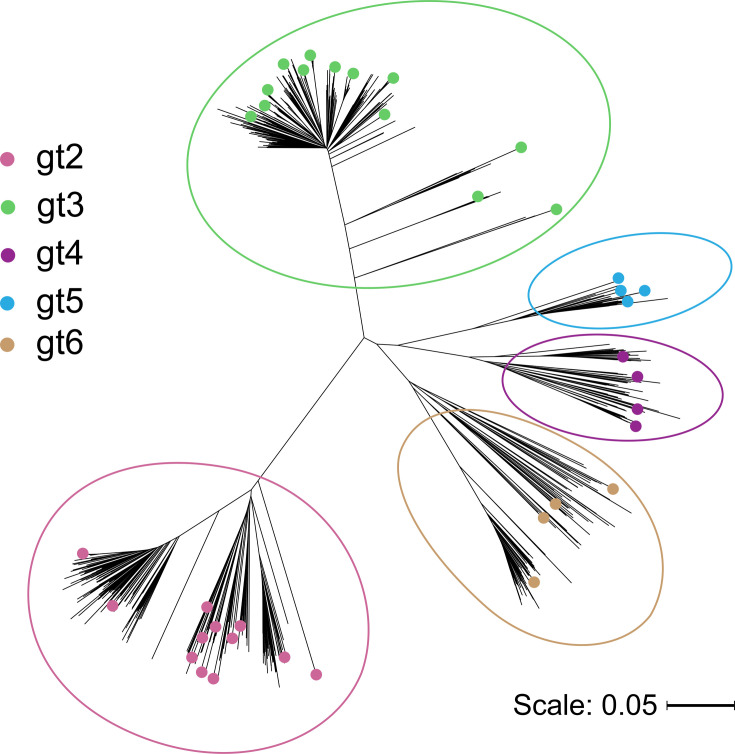 Fig. 1
Fig. 1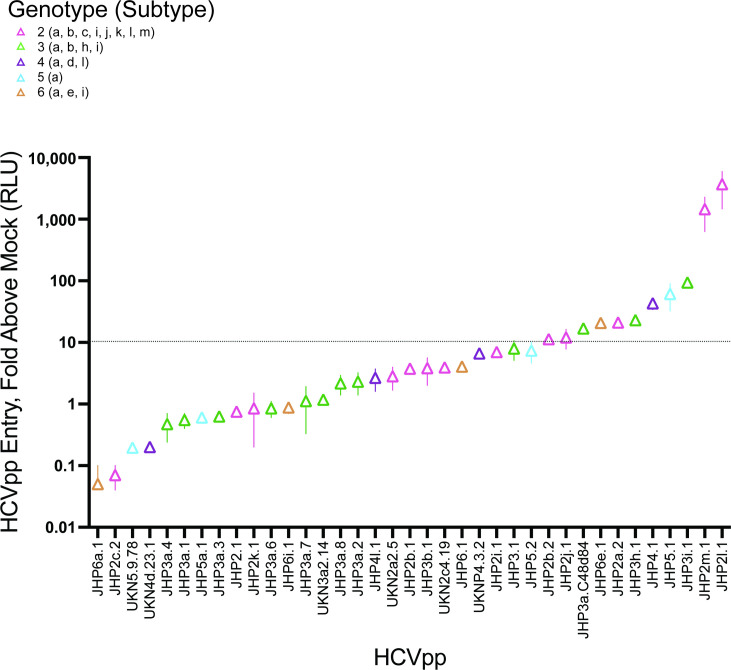 Fig. 2
Fig. 2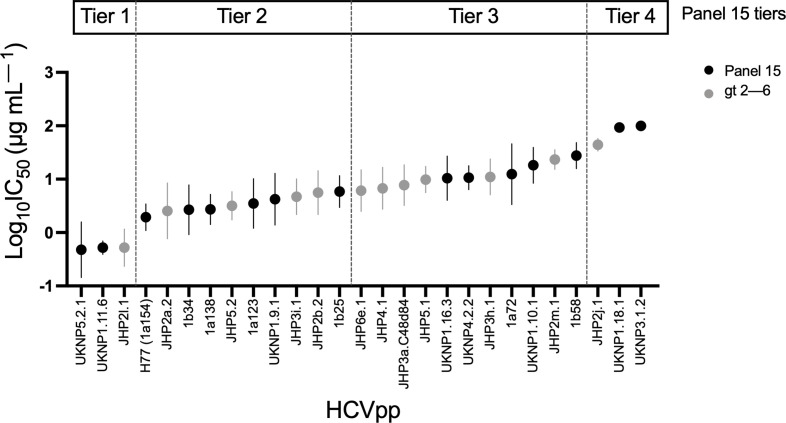 Fig. 3
Fig. 3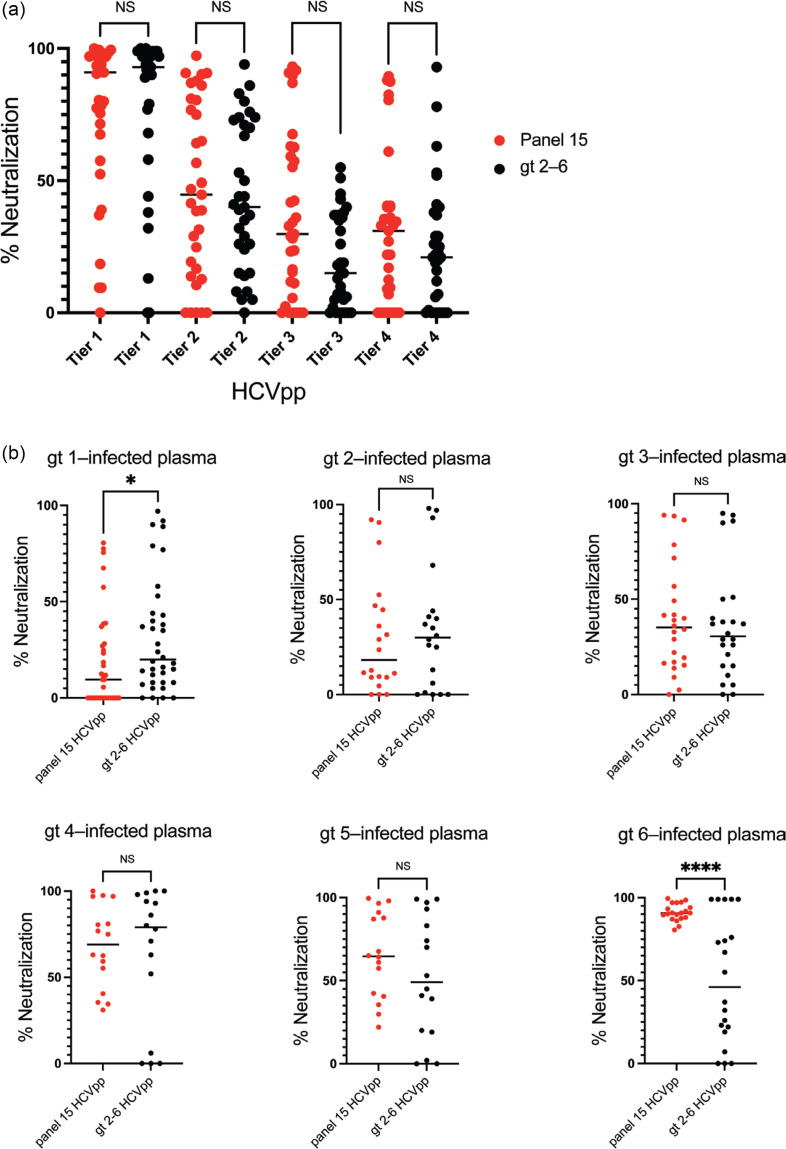 Fig. 4
Fig. 4| mAbs | HCVpp panel | HCVpp no. | Neutralizing breadth, %* | ||
|---|---|---|---|---|---|
| 100 µg ml−1 | 10 µg ml−1 | 1 µg ml−1 | |||
| hAP33 | gt 2–6 | 12 | 92 | 75 | 25 |
| Panel of 15 | 15 | 93 | 80 | 47 | |
| HCV1 | gt 2–6 | 12 | 92 | 75 | 17 |
| Panel of 15 | 15 | 87 | 87 | 40 | |
| AR4A | gt 2–6 | 12 | 92 | 83 | 42 |
| Panel of 15 | 15 | 87 | 80 | 33 | |
| HC84.26 | gt 2–6 | 12 | 100 | 92 | 67 |
| Panel of 15 | 15 | 73 | 67 | 33 | |
| AR3A | gt 2–6 | 12 | 100 | 75 | 33 |
| Panel of 15 | 15 | 60 | 53 | 13 | |
| HC33.4 | gt 2–6 | 12 | 58 | 33 | 8 |
| Panel of 15 | 15 | 73 | 40 | 27 | |
| CBH-7 | gt 2–6 | 12 | 25 | 8 | 0 |
| Panel of 15 | 15 | 33 | 13 | 13 | |
| Average | gt 2–6 | 12 | 80 | 63 | 27 |
| Average | Panel of 15 | 15 | 72 | 60 | 29 |
- —NIH
- —NIH
- —NIH
- —NIH
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHepatitis C virus research · Hepatitis B Virus Studies · Hepatitis Viruses Studies and Epidemiology
Data Availability
All data are available upon request. No new sequences were generated for this study.
Introduction
The global burden of hepatitis C virus (HCV) is staggering, with an estimated 1 million new infections annually [1]. Up to 50% of those with chronic HCV in the USA are unaware of their infection status, highlighting a critical gap in detection and treatment [2]. Left untreated, HCV can cause hepatocellular carcinoma, cirrhosis, end-stage liver disease and death. Although curative treatment exists, it is not widely accessible [34]. Developing a prophylactic vaccine is pivotal in reducing the incidence of new infections and the end-stage complications of HCV [56].
The genetic diversity of HCV is a barrier to vaccine development, with eight genotypes (gts) and 67 subtypes [78]. One strategy to tackle this diversity is to induce broadly neutralizing antibodies (bNAbs) that target conserved envelope glycoproteins 1 and 2 (E1E2) sites and consequently block infection by genetically diverse HCV isolates [916]. A humoral response with early bNAb induction is associated with the natural control of HCV infection in humans, and bNAbs can also prevent HCV infection in animal models [9,13, 1722].
A clear understanding of the relationship between genetic distance and antigenic differences between HCV variants is critical to facilitate the selection of variants for inclusion in panels measuring the neutralizing breadth of immune sera and variants for inclusion in candidate vaccines. In a study of HCV reinfection after spontaneous viral clearance, repeated infections with viruses carrying bNAb-sensitive E1E2 proteins were associated with induction of bNAbs [23]. Notably, greater genetic distance between re-infecting viruses was not correlated with the induction of broader neutralizing antibody responses. Similarly, prior analyses of panels of genetically diverse HCV variants have shown little association between HCV gt and sensitivity to neutralization [2426]. These findings indicate that genetic distance between HCV E1E2 variants is not a primary determinant of the antigenic differences between these variants.
We previously selected a neutralization panel of 15 antigenically diverse gt 1a (n=8), 1b (n=4), 3a (n=1), 4a (n=1) and 5a (n=1) HCV E1E2 variants based on differences in their overall neutralization sensitivity (Tiers 1–4), as well as in patterns of relative sensitivity to seven broadly neutralizing monoclonal antibodies (mAbs) targeting diverse E1E2 epitopes (i.e. antigenic profiles) [26]. As in prior studies, we observed no association between antigenicity and HCV gt. However, a limitation of that study was that most variants analysed were from gt 1. The final set of 15 HCV pseudoparticles (HCVpp) was downselected from an original set of 56 gt 1 variants and only nine gt 2–6 variants. Thus, the number of gt 2–6 variants analysed was small and did not represent the full genetic diversity of those HCV gts, so it remained possible that some antigenic features unique to gt 2–6 variants were not represented by the final panel of 15 E1E2 variants.
To address this limitation, we selected and synthesized 37 genetically diverse and representative gt 2–6 E1E2 variants from global databases. Using the subset of 12 variants that generated highly functional HCVpp, we measured neutralization by the same panels of bNAbs and human plasma samples used to characterize the previously published panel of 15 E1E2 variants. We analysed overall neutralization sensitivity as well as patterns of relative sensitivity to different bNAbs to identify any antigenic features that were unique to the new gt 2–6 E1E2 variants relative to the existing panel.
Methods
Cell lines
Single sources of Huh7 human hepatoma [25] and Cluster of Differentiation 81-knockout (CD81-ko) Human Embryo Kidney (HEK) 293T cells [27] (Dr. Joe Grove, University of Glasgow, Glasgow, UK) were used for the production of HCVpp. Cells were grown in Dulbecco’s Modified Essential Medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% FBS and 0.1 mmol l^−1^ nonessential amino acids (Invitrogen).
Antibodies
HCV mAbs CBH-7 [11], HC84.26 [10] and HC33.4 [12] were produced by Steven Foung. mAbs AR3A [9] and AR4A [13] were produced by Mansun Law. MAb hAP33 (a chimeric mouse AP33–human crystallizable fragment (Fc) antibody) [1628] was produced by Arvind Patel, and mAb HCV1 [29] was kindly provided by Yang Wang (MassBiologics, Boston, MA, USA).
Plasma
Plasma from gt 1–3-infected donors was obtained from the Baltimore Before and After Acute Study of Hepatitis [30]. Plasma samples representing gt 4–6 HCV infections were obtained from the University of Nottingham Trent HCV Cohort study [31]. Plasma was heat inactivated at 56 °C for 30 min prior to use.
Sequence analysis
Gt 2–6 E1E2 protein sequences were obtained from the National Center for Biotechnology Information (NCBI) GenBank, Los Alamos National Laboratory (LANL) HCV database [32], and the European HCV database (euHCVdb) [33], followed by filtering to remove partial sequences and redundant sequences (exact matches), yielding a total of 824 E1E2 reference sequences. A multiple sequence alignment (MSA) of the sequences was performed using the MAFFT tool (version 7) [34], and candidate sets of panel sequences were aligned to this reference MSA using the MAFFT ‘--add’ option. Sets of candidate gt 2–6 E1E2 panel sequences were generated as before for gt 1 E1E2 panels [26], selecting clade representatives from the reference set to maximize polymorphism coverage for each gt. A phylogenetic tree of the reference sequences was generated using the neighbour-joining method in MAFFT, and the tree was visualized using iTOL [35].
HCVpp production
Human immunodeficiency virus (HIV) Group-specific antigen (Gag)-packaged HCVpp were generated by Lipofectamine-mediated transfection of HCV E1E2 plasmid, pNL4-3.Luc.R-E- plasmid containing the env-defective HIV proviral genome (NIH AIDS Reagent Program), and pAdVantage (Promega) plasmid into CD81ko-HEK293T cells as described [36]. Mock pseudoparticles (mockpps) generated without E1E2 plasmid were used as a negative control for each transfection.
HCVpp entry
Huh7 cells (15,000 per well) were plated in 96-well microplates and incubated overnight. Then, 50 µl of HCVpp was added to the Huh7 cells in triplicate, and plates were incubated at 37 °C for 5 h. HCVpp were removed and replaced with phenol-free media, and cells were incubated for 72 h at 37 °C. HCVpp entry was determined by measurement of luciferase activity of cell lysate in relative light units (RLUs).
Neutralization
Neutralization assays were performed as described previously [36]. mAbs were serially fivefold diluted, starting at a concentration of 100 µg ml^−1^ (leaving the last well as PBS only), and incubated with HCVpp for 1 h at 37 °C before addition to Huh7 target cells in duplicate. HCVpp entry was measured as above. The percentage of neutralization was calculated as [1 − (RLU_mAb_/RLU_PBS_)] × 100, with the PBS RLU values averaged across two plates. Polyclonal human IgG (Thermo Fisher) was used as a negative control. Log_10_ 50% inhibitory concentrations (log_10_IC_50_) were calculated from neutralization curves fit by nonlinear regression [log(inhibitor) versus normalized response, variable slope] in Prism v10 (GraphPad Software). MAb–HCVpp tests that did not reach 50% inhibition were assigned an IC_50_ of 100 µg ml^−1^. Plasma samples were tested at a 1 : 20 dilution. Pooled plasma from ten HCV-negative donors, also at a 1 : 20 dilution, was used as a negative control. Percentage neutralization of each HCVpp was calculated as [1 − (RLU_immune plasma_/RLU_control plasma_)] × 100.
Results
Selection of representative gt 2–6 HCV E1E2 variants for functional analysis
We downloaded all available gt 2–6 full E1E2 sequences from the NCBI GenBank, LANL [32] and euHCVdb [33] databases (n=824). Using the aligned set of 824 E1E2 amino acid sequences, we used a custom script to identify amino acid polymorphisms at each position in E1E2 that were present in 2–80% of database isolates. This frequency range was chosen to focus on variable sites, while capturing polymorphisms that occur commonly enough to represent genuine circulating diversity (≥2% of isolates), excluding extremely rare substitutions that likely reflect isolated mutations or PCR errors. Based on this filtered dataset, we then identified 37 representative E1E2 variants (Table S1, available in the online Supplementary Material) expressing 97% of all common gt 2–6 polymorphisms and representative of diverse gt 2–6 phylogenetic clades (Fig. 1). We synthesized codon-optimized nucleotide sequences representing each of these amino acid sequences and cloned these into a mammalian expression vector, along with an N-terminal tissue plasminogen activator signal sequence.
Phylogenetic tree of reference and selected panel E1E2 sequences. A set of gts 2–6 E1E2 reference sequences (N=824) was processed with the MAFFT tool to generate an MSA and phylogenetic tree. The tree was visualized using iTOL. Selected panel sequences are indicated by coloured circles, and gts are circled for reference.
Wide variation in hepatoma cell entry of HCVpp gt 2–6 HCVpp
Using a standard approach, we generated HCVpp by transfecting each E1E2 expression plasmid along with a pNL4-3.Luc.R-E- expression plasmid containing the env-defective HIV proviral genome and collecting HCVpp-containing supernatants [36]. We measured the entry of HCVpp into Huh7 hepatoma cells to assess the relative function of each E1E2 glycoprotein (Fig. 2). Pseudoparticles lacking E1E2, known as mockpps, were tested in parallel to quantify nonspecific entry. Specific entries were calculated as a ratio of HCVpp entry relative to mockpp entry, with a predetermined threshold of ten-fold above mock defined as optimally functional for neutralization testing. Out of the 37 HCVpp, 11 (29.73%) exhibited specific entry exceeding the ten-fold cut-off in at least two production runs. One HCVpp (JHP5.2) was ten-fold above mock in only one production run but was carried forward to neutralization experiments. We selected these 12 HCVpp for subsequent neutralization testing; they included gts 2 (n=5), 3 (n=3), 4 (n=1), 5 (n=2) and 6 (n=1). The specific entry of HCVpp did not segregate by gt. Although some genetic diversity was lost by downselection from the original set of 37 E1E2 variants, this set of 12 highly functional HCVpp expressed 79% of all commonly observed gt 2–6 E1E2 polymorphisms.
Wide variation in hepatoma cell entry of gt 2–6 HCVpp. Hepatoma cell entry of 37 HCVpp is expressed as the fold increase in RLUs compared with the background entry of mockpp. Data represent 2–6 independent experiments for each HCVpp, performed in triplicate. Symbols indicate means, and whiskers indicate the sem. A threshold of ten-fold above mockpp entry is indicated with a dotted line.
Distribution of neutralization sensitivity of gt 2–6 HCVpp parallels the published panel of 15 HCVpp
We measured the neutralization of this panel of 12 gt 2–6 HCVpp. The neutralization of each HCVpp was tested using serial dilutions of the same seven bNAbs used to characterize the published 15 HCVpp panel, chosen for their ability to bind to a range of neutralizing epitopes across the E2 glycoprotein or E1E2 heterodimer (Table S2) [913,16, 29]. These seven mAbs recognize five distinct antigenic sites, including Domain B/AR3 (mAb AR3A), Domain C (mAb CBH-7), Domain D (mAb HC84.26), Domain AR4 (mAb AR4A) and Domain E/AS412 (HC33.4, HCV1 and hAP33). All mAbs were isolated from humans infected with HCV, except HCV1, which was generated by immunizing a transgenic mouse expressing human antibody genes [29], and hAP33, which was generated by immunization of a WT mouse [16], and then subsequently produced as a mouse–human chimera (i.e. its variable heavy and variable light chains grafted onto a human IgG1 Fc backbone) [28]. Neutralization of the 12 gt 2–6 HCVpp was compared with the previously published neutralization results obtained with the panel of 15 HCVpp (Fig. 3). To confirm reproducibility, neutralization of four control HCVpp representing Tiers 1–4 of the published panel of 15 HCVpp [UKNP1.11.6, H77 (1a154), 1a72 and UKNP1.18.1] was re-tested in parallel with the 12 gt 2–6 HCVpp. As expected, neutralization of the four control HCVpp was consistent with previously published data for the same HCVpp (Fig. S1).
Distribution of neutralization sensitivity of gt 2–6 HCVpp parallels the published panel of 15 HCVpp. Neutralization of gt 2–6 HCVpp was measured with seven bNAbs targeting diverse epitopes across E1E2. For reference, previously published neutralization data for the standard panel of 15 HCVpp from Salas et al. [26] is included (‘panel 15’, black circles). Data represent two independent experiments for each mAb-HCVpp combination, performed in duplicate. The symbols indicate means, and the whiskers indicate the sem. HCVpp are arranged from most to least neutralization sensitive based on mean log10IC50 and divided into four tiers based on sd from the mean of the standard panel of 15 HCVpp.
We observed a wide range of neutralization sensitivity across the 12 gt 2–6 HCVpp. We ranked the HCVpp from lowest to highest mean log_10_IC_50_ across the seven bNAbs. The mAbs HCV1 and hAP33 bind very similar epitopes on E2, so we averaged the log_10_IC_50_ values for these two mAbs, giving HCV1 and hAP33 each half of the weight of the other five bNAbs in our analysis. The HCVpp were separated into four tiers based on the normal distribution of their mean log_10_IC_50_ values. The distribution of neutralization sensitivity of the 12 gt 2–6 HCVpp was similar to the distribution of the previously published panel of 15 HCVpp, with one HCVpp (8.33%) in Tier 1, four HCVpp (33.33%) in Tier 2, five HCVpp (41.67%) in Tier 3 and two HCVpp (16.67%) in Tier 4. When IC_50_ values for each antibody–HCVpp pair were averaged (with HCV1 and AP33 combined as a single entry), the mean IC_50_ was 27.42 µg ml^−1^. The average IC_50_ across all mAbs for each HCVpp ranged from 1.64 to 61.70 µg ml^−1^ (Table S2). The panel of 15 HCVpp had very similar IC_50_ values with the same mAbs (mean=35.94 µg ml^−1^; range across HCVpp: 0.63 to >100 µg ml^−1^) [26]. For each mAb, we analysed the relationship between the number of substitutions in mAb epitopes for each E1E2 variant and neutralization resistance of the HCVpp (Fig. S2). For HCV1 and AP33, we observed a trend towards greater resistance with a larger number of substitutions, although these differences were not statistically significant. For the remaining mAbs, there was no apparent relationship between the number of epitope substitutions and mAb resistance, which is consistent with prior observations that polymorphisms distant from epitopes play a dominant role in neutralization resistance [3739]. Together, these data demonstrate that the gt 2–6 HCVpp have a wide range of neutralization sensitivity, mirroring the previously published panel of 15 HCVpp.
Neutralization breadth of bNAbs is very similar when measured with either the panel of 12 gt 2–6 HCVpp or the published panel of 15 HCVpp
The neutralizing breadth (i.e. per cent of HCVpp neutralized >50%) measured for each bNAb at IC_50_ thresholds of 100, 10 or 1 µg ml^−1^ was somewhat higher for HC84.26 and AR3A mAbs when quantitated using the gt 2–6 HCVpp panel rather than the panel of 15 HCVpp, but notably, average neutralizing breadth across all mAbs was remarkably similar when measured with either HCVpp panel (Table 1). For example, at an IC_50_ threshold of 10 µg ml^−1^, an average of 63% of the variants in the gt 2–6 panel and 60% of the variants in the panel of 15 HCVpp were neutralized.
Neutralization breadth of gt 2–6-infected plasma is similar when measured with either the panel of 12 gt 2–6 HCVpp or the published panel of 15 HCVpp
Using 33 plasma samples from persons infected with HCV, we tested neutralizing breadth using the gt 2–6 HCVpp panel (Table S3). We then compared these results with previously published data, which were acquired with the panel of 15 HCVpp and the same plasma samples (Fig. 4). The plasma samples were isolated from persons infected with gt 1 (n=9), 2 (n=5), 3 (n=6), 4 (n=4), 5 (n=4) or 6 (n=5) infections. Using plasma from all six gts, there were no significant differences between neutralization of Tier 1, 2, 3 or 4 HCVpp in the panel of gt 2–6 HCVpp relative to HCVpp in the same tiers from the panel of 15 HCVpp (Figs 4a and S3). As expected, when we compared the neutralization of Tier 1–4 HCVpp by gt 2–6-infected plasma, we observed significant differences between each tier, with the exception of Tiers 3 and 4, which showed similar neutralization sensitivity (Fig. S4).
Overall sensitivity to HCV-immune plasma of Tier 1–4 variants from gt 2–6 HCVpp and the published panel of 15 HCVpp are similar. (a) Per cent neutralization of each tier of the gt 2–6 HCVpp by plasma at 1 : 20 dilution from 33 individuals infected with gt 1–6 HCV, measured in duplicate, compared to published neutralization values for the panel of 15 HCVpp measured with the same plasma samples (Salas et al. [26]). Each point indicates the mean neutralization of all HCVpp in the indicated tier by a single plasma sample. Neutralization values <0 were normalized to 0. Horizontal lines indicate medians. Groups were compared by the Kruskal–Wallis test, with adjustment for multiple comparisons. (b) Neutralization of each HCVpp panel by HCV-immune plasma, with results segregated by the infecting gt of each participant. Each point indicates the mean neutralization of all HCVpp in each tier by a single plasma sample. Horizontal lines indicate medians. Non-normally distributed groups (gt 1–4 and gt 6) were compared by Kolmogorov–Smirnov test. The normally distributed group (gt 5) was compared by an unpaired t-test. ns, not significant; ∗P<0.05, ∗∗∗∗P<0.0001.
We then compared neutralization of the panel of gt 2–6 HCVpp and the panel of 15 HCVpp with plasma segregated by the HCV gt infecting each participant. There were no differences between panels in neutralization by gt 2-, 3-, 4- or 5-infected plasma. Notably, however, relative to the panel of 15 HCVpp, the panel of gt 2–6 HCVpp was slightly more sensitive to neutralization by gt 1-infected plasma and significantly more resistant to neutralization by gt 6-infected plasma (Figs 4b and S5). Overall, very similar neutralization results were obtained when testing HCV-immune plasma with either the panel of gt 2–6 HCVpp or the published panel of 15 HCVpp.
Discussion
Producing a vaccine for HCV demands an accurate assessment of the neutralizing breadth of antibodies elicited by the vaccine, and an understanding of the antigenic differences between potential vaccine variants. We previously identified a standardized panel of 15 antigenically diverse gt 1a, 1b, 3a, 4a and 5a HCV E1E2 variants for use in neutralization assays [26], but some concern remained that this panel did not fully represent the antigenic diversity of all gts. Therefore, we focused in this study on identifying genetically diverse and representative gt 2–6 E1E2 variants, and we compared the antigenic characteristics of these variants with those described for the published panel of 15 HCVpp. We observed remarkable similarity between the new gt 2–6 HCVpp panel and the existing 15 HCVpp panel in the range and magnitude of neutralization sensitivity to bNAbs or HCV-immune plasma. Neutralization breadth for each of the reference bNAbs was very similar when measured with either the published panel of 15 HCVpp or the new gt 2–6 panel. The only notable difference between the panels was in sensitivity to neutralization by plasma from gt 6-infected persons, with the new gt 2–6 panel displaying greater resistance to neutralization relative to the published panel of 15 HCVpp. The underlying mechanism is unclear, but we would speculate that this may indicate that the NAbs induced by gt 6 infection may target some unique epitopes relative to the NAbs induced by gt 1–5 infections. Further work to understand this phenomenon is warranted, and inclusion of these gt 2–6 variants should be considered for studies of neutralization in gt 6-infected individuals.
Neutralization results with the two panels are similar, even though the original panel of 15 HCVpp is dominated by gt 1 variants, while the new panel is focused exclusively on gt 2–6 variants. The original panel of 15 E1E2 variants expresses only 60% of common gt 2–6 polymorphisms, while the gt 2–6 panel expresses 79% of those polymorphisms. The similarity in neutralization results between panels supports prior studies [1724384041] indicating that the majority of E1E2 amino acid differences across gts are irrelevant to heterologous NAb sensitivity. Given the antigenic similarity between these panels, and the extensive data already generated across multiple laboratories using the original panel of 15 HCVpps, we would propose that there is no need to add these new gt 2–6 variants to the existing 15 HCVpp panel for standardized neutralization testing. These data provide further confirmation that neutralization data obtained with the panel of 15 HCVpp are generally representative of neutralization of all HCV variants from gts 1–6.
There are some limitations to this study. First, relatively few gt 2–6 E1E2 sequences are available in global databases. Even though all available sequences were analysed for this study, there may yet be variants in nature from gts 2–6, or other gts, with unique antigenic characteristics. Work should continue to sequence new variants from these still under-sampled gts. While we were careful to use bNAbs that target a variety of neutralizing epitopes, and we also tested neutralization with polyclonal sera that contain a variety of biologically relevant NAbs, it is possible that bNAbs isolated in the future might target new epitopes, which could result in different results between the two HCVpp panels.
A vaccine for HCV is urgently needed, and development of this vaccine will require accurate measurement of antibody neutralization breadth and understanding of the antigenic differences between E1E2 variants. We have characterized a genetically diverse and representative set of gt 2–6 E1E2 variants and found that neutralization results obtained with these variants very closely mirror results obtained with our previously published gt 1-dominant panel of 15 HCVpp. These data confirm that neutralization results obtained with the standardized panel of 15 HCVpp are generally representative of neutralization of all HCV gts.
Supplementary material
10.1099/jgv.0.002201Uncited Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1World Health Organization Global Hepatitis Report 2024
- 2Terrault NA Hepatitis C elimination: challenges with under-diagnosis and under-treatment F 1000 Res 201985410.12688/f 1000 research.15892.1PMC 633824430687501 · doi ↗ · pubmed ↗
- 3Kanwal F Kramer J Asch SM Chayanupatkul M Cao Y et al Risk of Hepatocellular cancer in HCV patients treated with direct-acting antiviral agents Gastroenterology 2017153996100510.1053/j.gastro.2017.06.01228642197 · doi ↗ · pubmed ↗
- 4Hamdane N Jühling F Crouchet E El Saghire H Thumann C et al HCV-induced epigenetic changes associated with liver cancer risk persist after sustained virologic response Gastroenterology 20191562313232910.1053/j.gastro.2019.02.03830836093 PMC 8756817 · doi ↗ · pubmed ↗
- 5Bailey JR Barnes E Cox AL Approaches, progress, and challenges to hepatitis C vaccine development Gastroenterology 201915641843010.1053/j.gastro.2018.08.06030268785 PMC 6340767 · doi ↗ · pubmed ↗
- 6Bartenschlager R Baumert TF Bukh J Houghton M Lemon SM et al Critical challenges and emerging opportunities in hepatitis C virus research in an era of potent antiviral therapy: considerations for scientists and funding agencies Virus Res 2018248536210.1016/j.virusres.2018.02.01629477639 · doi ↗ · pubmed ↗
- 7Simmonds P Becher P Bukh J Gould EA Meyers G et al ICTV virus taxonomy profile: flaviviridae J Gen Virol 2017982310.1099/jgv.0.00067228218572 PMC 5370391 · doi ↗ · pubmed ↗
- 8Smith DB Bukh J Kuiken C Muerhoff AS Rice CM et al Expanded classification of hepatitis C virus into 7 genotypes and 67 subtypes: updated criteria and genotype assignment web resource Hepatology 20145931832710.1002/hep.2674424115039 PMC 4063340 · doi ↗ · pubmed ↗