Commodity risk assessment of oak and walnut logs from the US
Antonio Vicent Civera, Paula Baptista, Anna Berlin, Elisavet Chatzivassiliou, Jaime Cubero, Nik Cunniffe, Eduardo de la Peña, Nicolas Desneux, Francesco Di Serio, Anna Filipiak, Beata Hasiów‐Jaroszewska, Hervé Jactel, Blanca B. Landa, Lara Maistrello, David Makowski

TL;DR
This paper assesses the effectiveness of a vacuum-steam-heat treatment in reducing the risk of pests in oak and walnut logs imported from the US to the EU.
Contribution
The study evaluates a new phytosanitary treatment's efficacy against specific pests using expert knowledge and systematic literature reviews.
Findings
The treatment is highly effective against Bretziella fagacearum and Geosmithia morbida in oak and walnut logs.
The treatment is less effective against pests like Arrhenodes minutus that infest deeper wood layers.
Expert knowledge elicitation provided 95% certainty on pest-free log estimates after treatment.
Abstract
The European Commission submitted to the EFSA Panel on Plant Health a Dossier from the United States proposing the use of a vacuum–steam–heat treatment as a stand‐alone phytosanitary measure to mitigate the risk of entry of Bretziella fagacearum, Geosmithia morbida and its vector Pityophthorus juglandis (thousand cankers disease complex) into the EU when trading oak (Quercus alba, Q. rubra) and walnut (Juglans nigra) logs with bark from the US. The proposed treatment consists of heating the sapwood to 56°C for 30 min at a depth of 5 cm from the cambium under vacuum and steam conditions. EFSA assessed the likelihood that logs of oak and walnut target species would be free from EU quarantine pests, basing its evaluation solely on the efficacy of the proposed treatment. In addition to B. fagacearum, G. morbida and P. juglandis, 14 other EU quarantine pests were identified as relevant…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
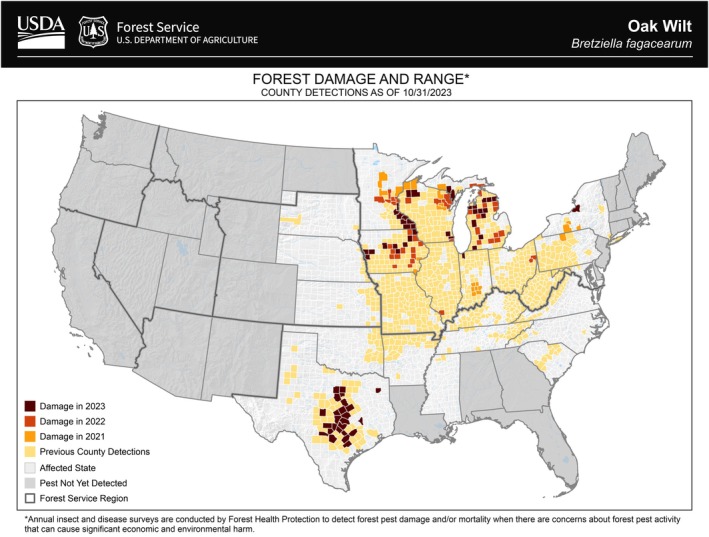 FIGURE 1
FIGURE 1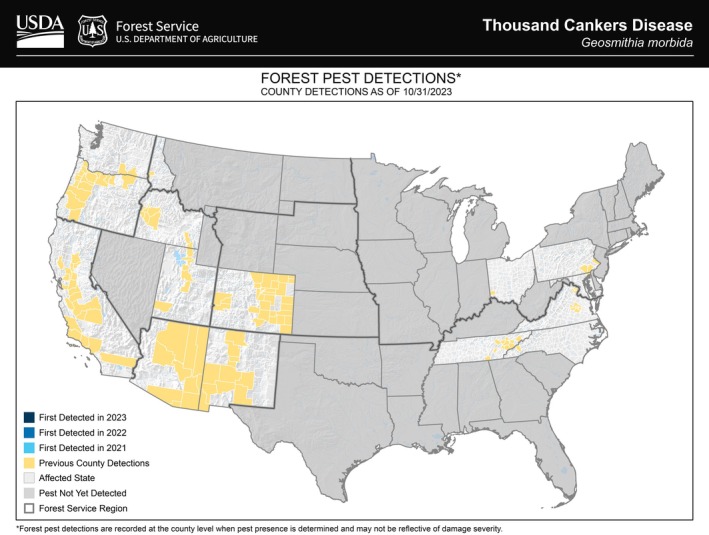 FIGURE 2
FIGURE 2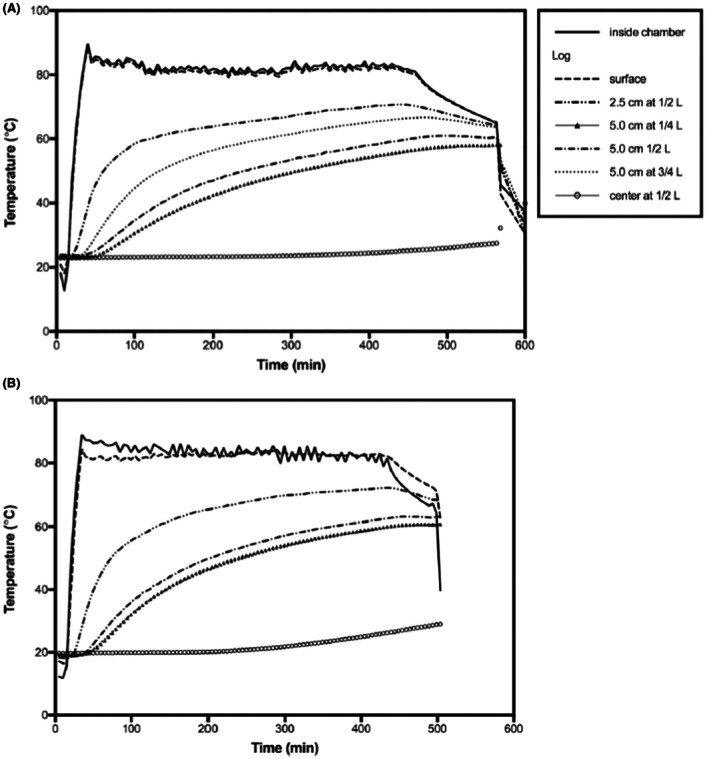 FIGURE 3
FIGURE 3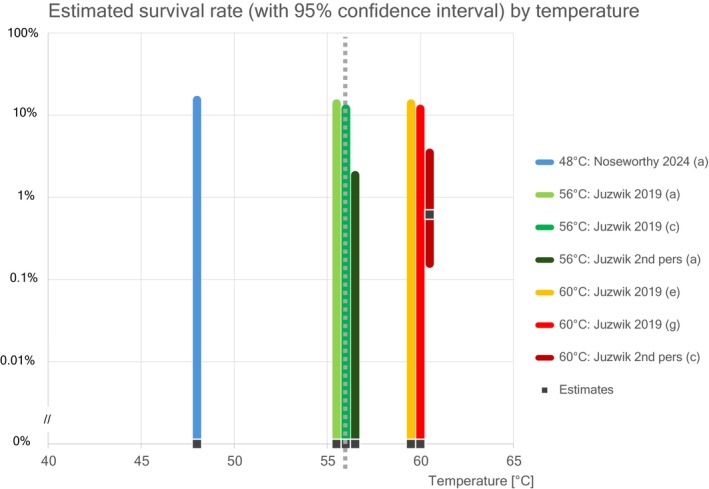 FIGURE 4
FIGURE 4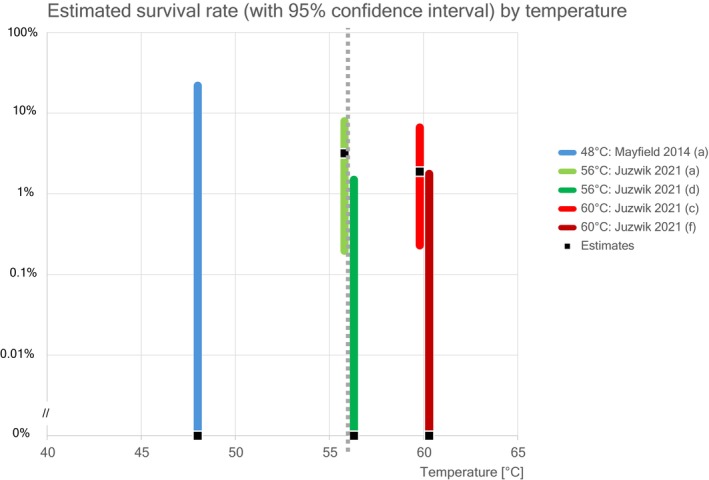 FIGURE 5
FIGURE 5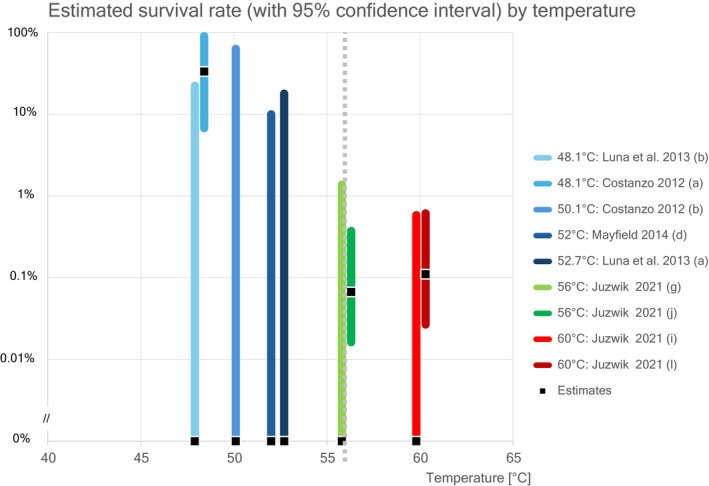 FIGURE 6
FIGURE 6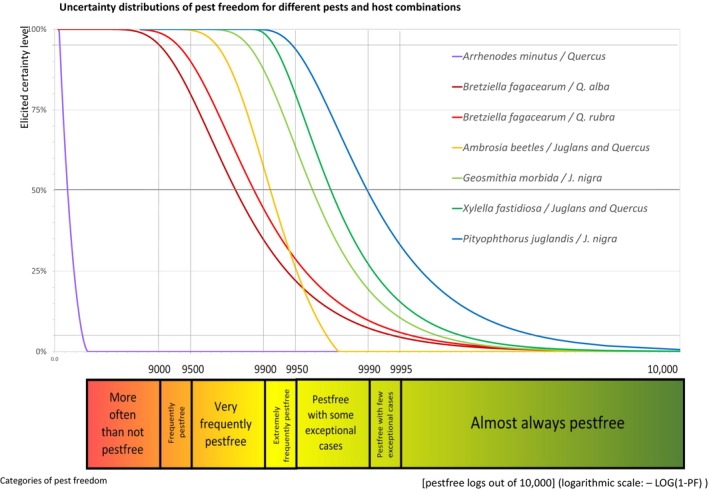 FIGURE 7
FIGURE 7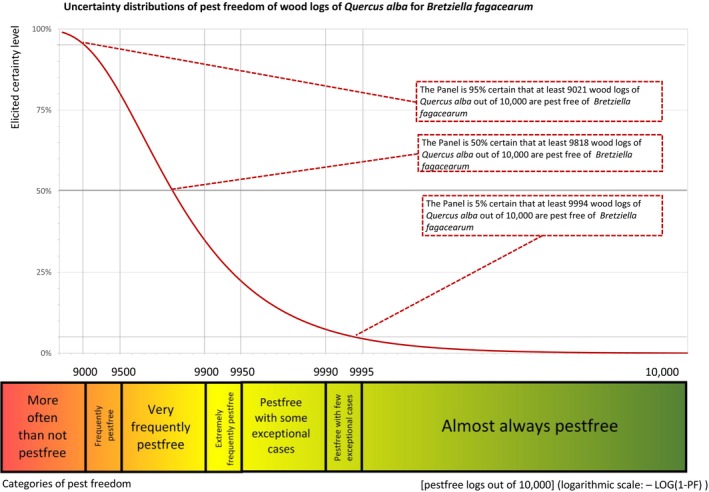 FIGURE 8
FIGURE 8| Dossier section | Overview of contents | Filename |
|---|---|---|
| 1 | Technical Dossier | Target Vacuum Steam Dossier final draft May 31 V1 |
| Oak and walnut – Annex 2.1 | ||
| Oak and walnut – Annex 2.2. | ||
| 2 | Additional information: answers to EFSA queries provided in April and May 2025 | Responses by the USDA APHIS to Questions by EFSA related to the Dossier Marked |
| Appendix 1 Society of American Foresters Forest Cover Types spreadsheet for coding list1 – Copy | ||
| Appendix 2 Second Red Oak Study_Report | ||
| Appendix 3 Second Red Oak Study_Analysis | ||
| Appendix 4 Maximum Sapwood Widths | ||
| Q1 responses for pathogens and insects | ||
| Q1 Scolytinae hosts | ||
| Q2 Copy of Species presence (003) | ||
| Q2 Silvics of N America_ | ||
| Q2 Silvics of N America_ | ||
| Q2 Silvics of N America_ |
| Database | Platform/link |
|---|---|
| Bark and Ambrosia Beetles of the Americas |
|
| CABI Crop Protection Compendium |
|
| Database of the World's Lepidopteran Hostplants |
|
| EPPO Global Database |
|
| GBIF |
|
| Nemaplex |
|
| Scalenet |
|
| Scolytinae hosts and distribution database |
|
| USDA ARS Fungal Database |
|
| Pest species/group of pests |
|
|
|
|---|---|---|---|
|
| X | X | n.a. |
|
| n.a. | n.a. | X |
|
| n.a. | n.a. | X |
| Ambrosia beetles | X | X | X |
|
| X | X | n.a. |
|
| X | X | n.a. |
| N | Name of harmful organisms (pests) | Group | Plant species | Commodity | Additional information on the commodity in the notes | Country of origin | Country of entry | Year of interception | Number of interceptions |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Acari | Mites |
| Products: wood and bark | – | US | Spain | 2012 | 1 |
| 2 |
| Insects |
| Products: wood and bark | – | US | Italy | 2019 | 1 |
| 3 | Anobiidae | Insects |
| Products: wood and bark | – | US | Spain | 2015 | 1 |
| 4 |
| 2009 | 2 | ||||||
| 5 | Arachnida | Arachnids |
| Products: wood and bark | Logs with bark | US | Italy | 2016 | 1 |
| 6 | Arctiidae – | Insects |
| Products: wood and bark | Lumber | US | Spain | 2002 | 2 |
| 7 |
| Insects |
| Products: wood and bark | – | US | France | 2005 | 1 |
| 8 | Bostrichidae | Insects |
| Products: wood and bark | – | US | Spain | 2014, 2015 | 2 |
| 9 |
| Sawn timber | Germany | 2011 | 1 | ||||
| 10 | Buprestidae | Insects |
| Products: logs | – | US | Not specified | 2022, 2023 | 16 |
| 11 | Products: wood and bark | Italy | 2019, 2020 | 2 | |||||
| 12 |
| 2017 | 1 | ||||||
| 13 |
| Insects |
| Products: wood and bark | – | US | Italy | 2020 | 1 |
| 14 | Cerambycidae | Insects |
| Products: logs | – | US | Not specified | 2021 | 1 |
| 15 |
| 2022, 2023 | 15 | ||||||
| 16 | Products: wood and bark | Spain | 2014 | 1 | |||||
| 17 | Logs | Italy | 2016, 2019 | 2 | |||||
| 18 | – | 2020 | 1 | ||||||
| 19 |
| Insects |
| Products: wood and bark | Logs | US | Italy | 2019 | 1 |
| 20 | – | 2019 | 1 | ||||||
| 21 |
| Insects |
| Products: logs | – | US | Not specified | 2022 | 5 |
| 22 |
| Products: others | 2024 | 1 | |||||
| 23 | Products: wood and bark | Italy | 2019 | 1 | |||||
| 24 |
| Logs | 2019 | 1 | |||||
| 25 |
| Insects |
| Products: wood and bark | Logs with bark | US | Germany | 2017 | 1 |
| 26 | – | Italy | 2019 | 1 | |||||
| 27 |
| Insects |
| Products: wood and bark | – | US | Italy | 2019 | 1 |
| 28 |
| Insects |
| Products: wood and bark | – | US | Italy | 2019 | 1 |
| 29 | Coccidae | Insects |
| Products: logs | – | US | Not specified | 2022 | 1 |
| 30 | Coleoptera | Insects |
| Products: others | – | US | Not specified | 2023 | 1 |
| 31 |
| Products: logs | 2022 | 1 | |||||
| 32 |
| 2022, 2023 | 12 | ||||||
| 33 | Products: others | 2022 | 2 | ||||||
| 34 | Products: sawn wood | 2024 | 1 | ||||||
| 35 |
| 2023 | 1 | ||||||
| 36 |
| Products: wood and bark | Spain | 2014 | 1 | ||||
| 37 |
| 2014 | 1 | ||||||
| 38 |
| 2014, 2016 | 2 | ||||||
| 39 | Diplopoda | Myriapoda |
| Products: wood and bark | – | US | Spain | 2016 | 1 |
| 40 | Diptera | Insects |
| Products: logs | – | US | Not specified | 2022 | 1 |
| 41 |
| Products: wood and bark | Spain | 2017 | 1 | ||||
| 42 | Entomobryidae | Collembola |
| Products: wood and bark | – | US | Spain | 2014 | 2 |
| 43 |
| Insects |
| Products: logs | – | US | Not specified | 2022 | 6 |
| 44 |
| Insects |
| Products: wood and bark | – | US | Spain | 2014 | 1 |
| 45 |
| Stumps | 2012 | 1 | |||||
| 46 | Formicidae | Insects |
| Products: wood and bark | Logs with bark | US | Italy | 2016 | 1 |
| 47 |
| Insects |
| Products: logs | – | US | Not specified | 2023 | 1 |
| 48 | Products: wood and bark | Italy | 2019 | 1 | |||||
| 49 |
| Insects |
| Products: wood and bark | – | US | Spain | 2016 | 1 |
| 50 |
| 2020 | 1 | ||||||
| 51 | Helicidae | Snails |
| Products: wood and bark | Logs with bark | US | Italy | 2016 | 1 |
| 52 | Hemiptera | Insects |
| Products: logs | – | US | Not specified | 2022 | 4 |
| 53 | Hymenoptera | Insects |
| Products: logs | – | US | Not specified | 2022 | 1 |
| 54 |
| Products: wood and bark | Spain | 2011 | 1 | ||||
| 55 | Insecta | Insects |
| Products: logs | – | US | Not specified | 2022, 2023, 2025 | 20 |
| 56 | Products: others | 2024 | 1 | ||||||
| 57 | Products: sawn wood | 2022, 2023 | 2 | ||||||
| 58 |
| 2022, 2023 | 10 | ||||||
| 59 |
| Products: wood and bark | Spain | 2014 | 4 | ||||
| 60 |
| France | 2015, 2016, 2017 | 5 | |||||
| 61 | Isoptera | Insects |
| Products: wood and bark | – | US | Spain | 2015 | 2 |
| 62 |
| 2015 | 1 | ||||||
| 63 | Lepidoptera | Insects |
| Products: logs | – | US | Not specified | 2022, 2023 | 4 |
| 64 |
| Products: wood and bark | Italy | 2019 | 1 | ||||
| 65 |
| Logs with bark | 2016 | 1 | |||||
| 66 |
| – | 2017 | 1 | |||||
| 67 |
| Spain | 2011, 2013 | 2 | |||||
| 68 |
| Insects |
| Products: wood and bark | – | US | Italy | 2019 | 1 |
| 69 |
| Insects |
| Products: wood and bark | – | US | Italy | 2019 | 1 |
| 70 |
| Insects |
| Products: logs | – | US | Not specified | 2023 | 1 |
| 71 | Products: wood and bark | Germany | 2019 | 1 | |||||
| 72 | Italy | 2019 | 1 | ||||||
| 73 |
| Insects |
| Products: wood and bark | Lumber | US | Germany | 2013 | 1 |
| 74 |
| Insects |
| Products: wood and bark | – | US | Germany | 2018 | 1 |
| 75 | Orthoptera | Insects |
| Products: logs | – | US | Not specified | 2022 | 1 |
| 76 | Pentatomidae | Insects |
| Products: logs | – | US | Not specified | 2022, 2023 | 2 |
| 77 |
| Insects |
| Products: logs | – | US | Not specified | 2024 | 1 |
| 78 | Platypodidae | Insects |
| Products: wood and bark | – | US | Spain | 2014 | 1 |
| 79 |
| Insects |
| Products: wood and bark | – | US | Spain | 2020 | 1 |
| 80 |
| Insects |
| Products: wood and bark | Logs | US | Italy | 2016 | 1 |
| 81 | Scolytidae | Insects |
| Products: logs | – | US | Not specified | 2022, 2023, 2024 | 13 |
| 82 |
| Products: sawn wood | 2023 | 1 | |||||
| 83 |
| Products: wood and bark | Spain | 2015 | 1 | ||||
| 84 |
| 2014, 2015 | 2 | ||||||
| 85 |
| 2013 | 1 | ||||||
| 86 | Silvanidae | Insects |
| Products: wood and bark | – | US | Spain | 2015 | 1 |
| 87 | Siricidae | Insects |
| Products: wood and bark | – | US | Spain | 2011 | 1 |
| 88 | Tenebrionidae | Insects |
| Products: wood and bark | – | US | Spain | 2011 | 1 |
| 89 | Tephritidae (non‐European) | Insects |
| Products: wood and bark | – | US | Spain | 2011 | 3 |
| 90 | Termitidae | Insects |
| Products: logs | – | US | Not specified | 2022 | 1 |
| 91 |
| Insects |
| Products: logs | – | US | Not specified | 2022 | 1 |
| 92 |
| Insects |
| Products: wood and bark | – | US | Germany | 2019 | 1 |
| 93 |
| Insects |
| Products: logs | – | US | Not specified | 2022 | 1 |
| 94 |
| Insects |
| Products: wood and bark | – | US | Italy | 2019 | 1 |
| 95 |
| Insects |
| Products: logs | – | US | Not specified | 2022 | 2 |
| 96 |
| Insects |
| Products: logs | – | US | Not specified | 2022, 2025 | 7 |
| 97 | Products: sawn wood | 2022 | 1 | ||||||
| 98 | Products: wood and bark | Italy | 2019 | 3 |
| No. | Pest name according to EU legislation | EPPO code | Group | Pest present in the US |
|
| Pest can be associated with the stems used for log production | Pest relevant for the opinion |
|---|---|---|---|---|---|---|---|---|
| 1 |
| CROISE | Insects | Yes | Yes (EPPO, | No | No | No |
| 2 |
| 1ANSTG | Insects | Yes | No | Yes (EFSA PLH Panel, | No | No |
| 3 |
| ANOLCN | Insects | No | Yes (Dong et al., | Yes (EPPO, | Not assessed | No |
| 4 |
| ANOLGL | Insects | Yes | Yes (EPPO, | No | Yes, only for | Yes |
| 5 |
| APRIGE | Insects | No | Yes (EPPO, | Yes (EPPO, | Not assessed | No |
| 6 |
| AROMBU | Insects | No | Yes (Dong et al., | Yes (EPPO, | Not assessed | No |
| 7 |
| ARRHMI | Insects | Yes | Yes (EPPO, | No | Yes, only for | Yes |
| 8 |
| DACUDO | Insects | Yes | No | Yes (EFSA, | No | No |
| 9 |
| 1BCTRG | Insects | No | No | Yes (EPPO, | Not assessed | No |
| 10 |
| BEMITA | Insects | Yes | Yes (EFSA PLH Panel, | Yes (EFSA PLH Panel, | No | No |
| 11 |
| BEMITA | Insects | No | Yes (EFSA PLH Panel, | Yes (EFSA PLH Panel, | Not assessed | No |
| 12 |
| TETTFE | Insects | No | Yes (EPPO, | No | Not assessed | No |
| 13 |
| CERAFA | Fungi | Yes | Yes (EFSA PLH Panel, | No | Yes, only for | Yes |
| 14 |
| PHYPFR | Phytoplasmas | No | Yes (Silva‐Castaño et al., | No | Not assessed | No |
| 15 |
| CHONRO | Insects | Yes | Yes (EPPO, | No | No | No |
| 16 |
| 1CRONG | Fungi | Yes | Yes (EPPO, | No | No | No |
| 17 |
| ENDOPA | Fungi | Yes | Yes (EPPO, | No | Yes, only for | Yes |
| 18 |
| CERAVI | Fungi | Yes | Yes (Farr & Rossman, | No | Yes, only for | Yes |
| 19 |
| DIABUN | Insects | Yes | Yes (Clark et al., | Yes (Clark et al., | No | No |
| 20 |
| DIABVZ | Insects | Yes | Yes (Clark et al., | No | No | No |
| 21 |
| HYPOMA | Fungi | Yes | Yes (EPPO, | No | Yes, only for | Yes |
| 22 |
| EOTELE | Mites | Yes | Yes (EPPO, | No | No | No |
| 23 |
| EURHBR | Insects | No | No | Yes (EPPO, | Not assessed | No |
| 24 |
| XYLBFO | Insects | Yes | Yes (EPPO, | Yes (EPPO, | Yes, for both | Yes |
| 25 |
| GEOHMO | Fungi | Yes | No | Yes (EPPO, | Yes, only for | Yes |
| 26 |
| GRCPAT | Insects | Yes | Yes (EPPO, | Yes (EPPO, | No | No |
| 27 |
| GRCPVE | Insects | Yes | Yes (EFSA PLH Panel, | Yes (EFSA PLH Panel, | No | No |
| 28 |
| LASPPR | Insects | Yes | Yes (Brown, | No | No | No |
| 29 |
| HOMLTR | Insects | Yes | Yes (EPPO, | Yes (EPPO, | No | No |
| 30 |
| LEPOQU | Insects | Yes | Yes (Doering, | Yes (Doering, | No | No |
| 31 |
| LOPLJA | Insects | Yes | Yes (Batsankalashvili et al., | Yes (Batsankalashvili et al., | Yes, for both | Yes |
| 32 |
| LYCMDE | Insects | Yes | Yes (Barringer & Ciafré, | Yes (Barringer & Ciafré, | Yes, for both | Yes |
| 33 |
| MALLRA | Insects | No | Yes (EPPO, | No | Not assessed | No |
| 34 |
| 1MONCG | Insects | No | Yes (EFSA PLH Panel, | Yes (EFSA PLH Panel, | Not assessed | No |
| 35 |
| FUSAAM | Fungi | Uncertain | Uncertain | Uncertain | Not assessed | No |
| 36 |
| FUSAEW | Fungi | Yes | Yes (EPPO, | Uncertain | Yes, only for | Yes |
| 37 |
| OEMOHI | Insects | No | Yes (EPPO, | Yes (EPPO, | Not assessed | No |
| 38 |
| ONCMNI | Insects | Yes | Yes (Adlerz, | No | No | No |
| 39 |
| ONCMUN | Insects | Yes | Yes (EPPO, | Yes (EPPO, | No | No |
| 40 |
| PHMPOM | Fungi | Yes | Yes (EPPO, | Yes (EPPO, | No | No |
| 41 |
| PHYTRA | Oomycetes | Yes | Yes (EPPO, | No | Yes, only for | Yes |
| 42 |
| PITOJU | Insects | Yes | No | Yes (EPPO, | Yes, only for | Yes |
| 43 |
| POPIJA | Insects | Yes | Yes (EPPO, | Yes (EPPO, | No | No |
| 44 |
| PSDPMI | Insects | Yes | Yes (DAFNAE, | Yes (DAFNAE, | Yes, for both | Yes |
| 45 |
| PSDPPR | Insects | Yes | Yes (DAFNAE, | No | Yes, only for | Yes |
| 46 |
| 1RHAGG | Insects | Yes | No | Yes (EFSA PLH Panel, | No | No |
| 47 |
| SCITCI | Insects | Yes | Yes (EPPO, | No | No | No |
| 48 | Scolytinae (non‐European) | 1SCOLS | Insects | Yes | Yes (DAFNAE, | Yes (DAFNAE, | Yes, for both | Yes |
| 49 |
| LAPHFR | Insects | Yes | No | Yes (EPPO, | No | No |
| 50 |
| ARGPLE | Insects | No | Yes (EPPO, | Yes (EPPO, | Not assessed | No |
| 52 |
| THAUPR | Insects | No | Yes (EPPO, | Yes (EPPO, | Not assessed | No |
| 53 |
| TOXOCI | Insects | Yes | Yes (EFSA PLH Panel, | No | No | No |
| 54 |
| AELSSA | Insects | No | Yes (EPPO, | Yes (EPPO, | Not assessed | No |
| 55 |
| XIPHAA | Nematodes | Yes | Yes (Xu & Zhao, | No | No | No |
| 56 |
| XIPHRI | Nematodes | Yes | Yes (EPPO, | Yes (EPPO, | No | No |
| 57 |
| XIPHTA | Nematodes | Yes | Yes (Xu & Zhao, | No | No | No |
| 58 |
| XYLEFA | Bacteria | Yes | Yes (EFSA, | Yes (EFSA, | Yes, for both | Yes |
| Number | Current scientific name | EPPO code | Name used in the EU legislation | Taxonomic information | Group | Regulatory status | Host plant | Name of Pest datasheet |
|---|---|---|---|---|---|---|---|---|
| 1 |
| ANOLGL |
|
Coleoptera Cerambycidae | Insects | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
|
|
| 2 |
| ARRHMI |
|
Coleoptera Brentidae | Insects | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
|
|
| 3 |
| CERAFA |
|
Microascales Ceratocystidaceae | Fungi | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
|
|
| 4 |
| ENDOPA |
|
Diaporthales Cryphonectriaceae | Fungi | Protected Zone Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
|
|
| 5 |
| CERAVI |
|
Microascales Ceratocystidaceae | Fungi | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
|
|
| 6 |
| HYPOMA |
|
Xylariales Xylariaceae | Fungi | Protected Zone Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
|
|
| 7 |
| XYLBFO |
|
Coleoptera Curculionidae Scolytinae | Insects | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
| Ambrosia beetles |
| 8 |
| GEOHMO |
|
Hypocreales Bionectriaceae | Fungi | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
|
|
| 9 |
| LOPLJA |
|
Hemiptera Diaspididae | Insects | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
|
|
| 10 |
| LYCMDE |
|
Hemiptera Fulgoridae | Insects | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
|
|
| 11 |
| FUSAEW |
|
Hypocreales Nectriaceae | Fungi | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
|
|
| 12 |
| PHYTRA |
|
Peronosporales Peronosporaceae | Oomycetes | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
|
|
| 13 |
| PITOJU |
|
Coleoptera Curculionidae Scolytinae | Insects | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
|
|
| 14 |
| PSDPMI |
|
Coleoptera Curculionidae Scolytinae | Insects | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
| Bark beetles |
| 15 |
| PSDPPR |
|
Coleoptera Curculionidae Scolytinae | Insects | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
| Bark beetles |
| 16 | Scolytinae spp. (non‐European) | 1SCOLS | Scolytinae spp. (non‐European) |
Coleoptera Curculionidae Scolytinae | Insects | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
|
1) Ambrosia beetles 2) Bark beetles |
| 17 |
| XYLEFA |
|
Lysobacterales Lysobacteraceae | Bacteria | EU Quarantine Pest according to Commission Implementing Regulation (EU) 2019/2072 |
|
|
| ISPM 15 | New Zealand (NZ MPI 2018) | Australia (DAFF‐BIOCON) |
|---|---|---|
| Wood packaging material | Sawn wood | Timber from Myrtaceae and pathogen risk species (many genera grown in NZ, US, Europe) |
|
Core temp.: 56°C Duration: 0.5 h |
Core temp.: 70, 80, 90, 100, 110, 120°C Duration: 4 h, 2 h, 1 h, 0.5 h, 20 min, 15 min |
Core temp.: 74°C Duration: 4 h (≤ 25 mm thickness) 18 h (> 200 mm thickness) Duration depends on thickness of logs (longer duration with increasing thickness) |
| Observation k out of N | Lower bound | Upper bound |
|---|---|---|
| 0 < k < N | BETA.INV(0.025, k, N‐k + 1) | BETA.INV(0.975, k + 1, N‐k) |
| k = 0 (full absence in the sample) | 0% | 1‐EXP(LN(0.05/N) ‘Upper 95% level’ |
| k = N (full presence in the sample) | EXP(LN(0.05/N) ‘Lower 95% level’ | 100% |
| Reference & study | Temperature [°C] | Duration [min] | Sample size: N | Survivors: k | Estimate of survival [%] | 95% CI lower | 95% CI upper |
|---|---|---|---|---|---|---|---|
| 48°C: Noseworthy 2024 (a) | 48 | 30 | 18 | 0 | 0% | 0% | 15% |
| 56°C: Juzwik 2019 (a) | 56 | 30 | 20 | 0 | 0% | 0% | 14% |
| 56°C: Juzwik 2019 (c) | 56 | 30 | 24 | 0 | 0% | 0% | 12% |
| 56°C: Juzwik 2nd pers (a) | 56 | 30 | 159 | 0 | 0% | 0% | 1.9% |
| 60°C: Juzwik 2019 (e) | 60 | 60 | 20 | 0 | 0% | 0% | 14% |
| 60°C: Juzwik 2019 (g) | 60 | 60 | 24 | 0 | 0% | 0% | 12% |
| 60°C: Juzwik 2nd pers (c) | 60 | 60 | 155 | 1 | 0.62% | 0.16% | 3.5% |
| Reference & study | Temperature [°C] | Duration [min] | Sample size: N | Survivors: k | Estimate of survival [%] | 95% CI lower | 95% CI upper |
|---|---|---|---|---|---|---|---|
| 48°C: Mayfield 2014 (a) | 48 | 40 | 12 | 0 | 0% | 0% | 22% |
| 56°C: Juzwik 2021 (a) | 56 | 30 | 126 | 4 | 3.2% | 0.20% | 7.9% |
| 56°C: Juzwik 2021 (d) | 56 | 30 | 199 | 0 | 0% | 0% | 1.5% |
| 60°C: Juzwik 2021 (c) | 60 | 60 | 106 | 2 | 1.9% | 0.23% | 6.6% |
| 60°C: Juzwik 2021 (f) | 60 | 60 | 168 | 0 | 0% | 0% | 1.8% |
| Reference & study | Temperature [°C] | Duration [min] | Sample size: N | Survivors: k | Estimate of survival [%] | 95% CI lower | 95% CI upper |
|---|---|---|---|---|---|---|---|
| Luna et al., 2013 (b) | 48.1 | 30 | 12 | 0 | 0% | 0% | 22% |
| Costanzo 2012 (a) | 48.1 | 0 | 3 | 1 | 33% | 6.8% | 91% |
| Costanzo 2012 (b) | 50.1 | 30 | 3 | 0 | 0% | 0% | 63% |
| Mayfield 2014 (d) | 52 | 40 | 28 | 0 | 0% | 0% | 10% |
| Luna et al., 2013 (a) | 52.7 | 30 | 15 | 0 | 0% | 0% | 18% |
| Juzwik 2021 (g) | 56 | 30 | 216 | 0 | 0% | 0% | 1.4% |
| Juzwik 2021 (j) | 56 | 30 | 1491 | 1 | 0.067% | 0.016% | 0.37% |
| Juzwik 2021 (i) | 60 | 60 | 512 | 0 | 0% | 0% | 0.58% |
| Juzwik 2021 (l) | 60 | 60 | 906 | 1 | 0.11% | 0.027% | 0.61% |
| Number | Group* | Pest species | Sometimes pest free | More often than not pest free | Frequently pest free | Very frequently pest free | Extremely frequently pest free | Pest free with some exceptional cases | Pest free with few exceptional cases | Almost always pest free |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
|
|
|
| ||||||
| 2 |
|
|
|
| ||||||
| 3 |
|
|
|
| ||||||
| 4 |
|
|
|
| ||||||
| 5 |
|
|
|
| ||||||
| 6 |
|
| ||||||||
| 7 |
|
|
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsForest Insect Ecology and Management · Plant and Fungal Interactions Research · Research on scale insects
INTRODUCTION
1
Background and Terms of Reference as provided by European Commission
1.1
Background
1.1.1
Special requirements apply to the introduction into the Union of logs of oak (Quercus L.) and walnut (Juglans L.) originating from the United States (US), in order to reduce the risk for introducing Union quarantine pests into the Union territory. Particular emphasis lies on Bretziella fagacearum, the fungus associated with oak wilt and on Geosmithia morbida, the fungus associated with thousand canker disease as well as its vector Pityophthorus juglandis. Corresponding requirements are laid down in points 83 and 90 of Annex VII to Regulation (EU) 2019/2072.1 A derogation for the introduction of oak logs with bark originating from the US has been established by Regulation (EU) 2023/1312,2 conditional to the application of a systems approach defined in that Regulation.
In June 2023, the US introduced a request for a derogation from the aforementioned requirements for logs with bark which underwent a specific vacuum and steam treatment where a minimum temperature of 56°C is reached for at least 30 min in a depth of 5 cm in the wood.
In support of the request, a report describing the treatment and summarising studies relating to efficacy and costs, was submitted.
Terms of Reference
1.1.2
EFSA is requested, pursuant to Article 29 of Regulation (EC) No 178/2002,3 to provide a scientific opinion.
In particular, EFSA is requested to assess, based on the information provided by the US, the level of certainty of freedom of oak logs from Bretziella fagacearum and of walnut logs from Geosmithia morbida and Pityophthorus juglandis when treated with vacuum and steam as proposed by the US. EFSA shall quantify the efficacy of the treatment to mitigate the risk of introduction of Union quarantine pests.
In this assessment, EFSA shall take into account the available scientific information, and in particular the scientific and technical information provided by the US, as well as existing international and regional phytosanitary standards. If necessary to complete its assessment, EFSA may ask additional technical information or clarifications from the US regarding their request to introduce logs of oak and walnut wood treated with vacuum and steam as proposed by the US into the Union.4 Following the provision of such information, EFSA shall proceed with the assessment.
Interpretation of the Terms of Reference
1.2
The applicant country clarified that only logs of Quercus alba, Q. rubra and Juglans nigra are intended for export to the EU and that the proposed vacuum steam treatment is intended as a stand‐alone measure for oak and walnut logs with bark and that no systems approach will be applied, which includes additional risk mitigation measures. In agreement with the Commission, EFSA evaluated the likelihood of pest freedom from Union quarantine pests potentially associated with Q. alba, Q. rubra and J. nigra logs from the US with emphasis on B. fagacearum, G. morbida and its vector P. juglandis and focusing only on the efficacy of the vacuum–steam–heat treatment as proposed by the applicant country. The risk of secondary infestation after vacuum–steam–heat treatment was not evaluated and hence is not covered by the current assessment.
DATA AND METHODOLOGIES
2
Data
2.1
Data provided by the applicant
2.1.1
The Panel considered all the data and information (hereafter called ‘the Dossier’) provided by the United States Department of Agriculture (USDA), Animal and Plant Health Inspection Service (APHIS) in June 2023. Additional information was provided by USDA APHIS in April and in May 2025, after EFSA's request. The Dossier was managed by EFSA.
The structure and overview of the Dossier is shown in Table 1. The number of the relevant section is indicated in the Opinion when referring to a specific part of the Dossier.
The data and supporting information provided by USDA APHIS formed the basis of the commodity risk assessment.
Literature searches performed by EFSA on the association of EU quarantine pests with oak and walnut and their presence in the US
2.1.2
The list of EU quarantine pests was retrieved from Commission Implementing Regulation 2019/2072. For each of those pests, the databases listed in Table 2 were checked for the association of the pest with Quercus and Juglans (genus and species). For the pests identified as associated with Quercus and/or Juglans, a literature search on whether they are present in the US was performed. The searches were run between January and March 2025.
Additional searches, limited to retrieve documents, were run when developing the Opinion. The available scientific information, including previous EFSA opinions on the relevant pests and the relevant literature and legislation, were taken into account.
Literature searches performed by EFSA on the temperature needed to kill pests
2.1.3
A systematic literature search was performed by EFSA in order to retrieve information on whether bark and wood colonising pests could survive the temperature reached in the vacuum–steam–heat treatment. A tailored search was conducted on the temperature extremes survived by B. fagacearum, G. morbida and its vector P. juglandis (see Appendix B). The literature retrieved on pests relevant for this opinion was screened for information on temperature limits for survival. In addition, literature on survival limits of different groups of organisms from EFSA PLH Panel (2024a) was used. The information retrieved, including studies submitted by the applicant are summarised in Section 6 and in Appendix B.
Methodologies
2.2
Identification of pests potentially associated with the commodity in addition to those listed in the terms of reference
2.2.1
To evaluate the pest risk associated with the importation of the commodity from the US, a pest list was compiled. The pest list includes all identified EU quarantine pests reported as potentially associated with Quercus and Juglans species based on information provided in the Dossier Sections 1 and 2 and on searches performed by the Panel as indicated above in Section 2.1.2. The search strategy and search syntax were adapted to each of the databases listed in Table 2, according to the options and functionalities of the different databases and CABI keyword thesaurus.
The scientific names of the EU quarantine pests were used when searching in the databases.
The compiled pest list (see Microsoft Excel® in Appendix D) includes all identified EU quarantine pests that use as host Quercus and Juglans species.
The relevance of an EU quarantine pest or a protected zone quarantine pest for this opinion was based on evidence that:
- the pest is associated with either Quercus or Juglans or both;
- the pest is present in the US;
- one or more life stages of the pest can be associated with the stems used for log production.
Pests that fulfilled all criteria were selected for further evaluation. If one of the three criteria was not fulfilled, the other criteria were not assessed.
Comparison of the information found on temperature limits of pests with the temperature achieved in the vacuum–steam–heat treatment
2.2.2
The information on temperature survival limits of relevant pests found in literature and the results of studies submitted in the Dossier were analysed considering the proposed exposure in the vacuum–steam–heat treatment (i.e. 56°C for 30 min at a sapwood depth of 5 cm). For details, see Section 6. This analysis was taken into consideration in the Expert Knowledge Elicitation (EKE).
Expert Knowledge Elicitation
2.2.3
To estimate the pest freedom of the commodity, an EKE was performed following EFSA Guidance (Annex B.8 of EFSA Scientific Committee, 2018).
The pest freedom of oak logs (Q. alba and Q. rubra) and walnut logs (J. nigra) treated as proposed by the applicant was assessed. The applicant proposed the vacuum–steam–heat treatment as a stand‐alone method without applying any other risk mitigation measures. In addition, although generally important for the estimation of pest freedom, the prevalence of pests in the country of origin of the commodity was also not considered in the EKE because the information provided by the applicant country was not sufficient (see Section 6.2).
Therefore, the EKE was focused only on the efficacy of the vacuum–steam–heat treatment of logs considering the information on temperature limits of survival of pests and the information on sapwood thickness of oak logs.
The specific EKE question was: ‘Assuming all the logs are infested by a given pest according to its expected colonisation patterns, how many logs out of 10,000 remain infested with viable pests after treatment?’
An overview of the elicitations conducted for the different combinations of pests and commodity species is provided in Table 3.
In addition to the three pests which are listed in the mandate also other pest species were identified for further evaluation. The pests were grouped according to the depth of wood they can colonise and representative species were selected for assessment, taking into consideration that 5 cm depth from the cambium is the reference for the treatment temperature and duration of treatment (56°C for 30 min).
The pests were grouped as follows:
Bark and sapwood‐associated fungi are considered to be covered by the assessment of B. fagacearum and G. morbida.
Bark and sapwood‐dwelling insects are considered to be covered by the assessment conducted for P. juglandis and ambrosia beetles.
Insects which can inhabit the heartwood are represented by A. minutus.
Insects which are dwelling on the outer bark will not be able to survive the temperatures proposed in the treatment as they will be exposed to temperatures significantly higher than 56°C (see also Section 6.2). Therefore, this group of insects was not assessed further in the EKE.
The uncertainties associated with the EKE were considered and quantified in a probability distribution, fitted to the elicited percentiles applying the semi‐formal method described in Section 3.5.2 of the EFSA Guidance on quantitative pest risk assessment (EFSA PLH Panel, 2018a). Finally, the EKE results were reported in terms of the likelihood of pest freedom, calculated by 1 minus the likelihood to be infested. The lower 5% percentile of the uncertainty distribution reflects the opinion that pest freedom is with 95% certainty above this limit.
The results of the EKE are reported in Section 7.
THE COMMODITY
3
Description
3.1
The minimum diameter of logs with bark of Q. rubra, Q. alba and J. nigra intended for export to the EU is approximately 33 cm and the maximum diameter is approximately 76 cm measured on the small end of the log (Dossier Sections 1 and 2). The minimum and maximum length of the logs would be 1.83 and 12.19 m, with the majority of logs being in the range of 2.44–4.88 m.
Production areas
3.2
All growing sites of Quercus and Juglans logs which are intended for export to the EU are located in the US. Quercus trees are grown in mixed stands with variable composition in different geographic areas. Typical species in these mixed stands include Acer saccharum, A. saccharinum, A. rubrum, Carya ovata, C. laciniosa, C. glabra, C. cordiformis, Diospyros virginiana, Fagus grandifolia, Fraxinus pennsylvanica, Fraxinus americana, Juglans nigra, Liriodendron tulipifera, Platanus occidentalis, Populus deltoides, Prunus serotina, Robinia pseudoacacia and Ulmus americana. Quercus stands are uneven aged in mixed and natural stands (Dossier Section 2).
Juglans trees are harvested in pure stands (i.e. plantations) as well as in naturally regenerated mixed stands. Juglans is not dominant in most forests but rather is generally found as scattered single trees or as small, isolated groups within hardwood stands (Dossier Section 2).
Harvesting and handling processes
3.3
Harvesting of both Quercus and Juglans is limited to the winter (October–April). Quercus trees are primarily harvested by selective cutting rather than clear‐cutting (Dossier Section 2).
Juglans trees are primarily harvested selectively, meaning only mature, high‐quality trees are chosen to be harvested while allowing younger trees to continue growing in the forest stand; this practice promotes sustainable forest management. However, they can be clear‐cut if the situation (e.g. even age plantation) warrants. The higher quality and larger trees are harvested from natural stands.
The trees are cut approximately 20–30 cm above ground during harvesting operations. Branches are cut from the main stem and often chipped on site after trees are felled (Dossier Section 2).
In most cases, the logs are removed from the forest within 7–10 days from the time of harvest. Logs are then usually transported to and stored at a concentration yard prior to treatment (Dossier Section 2).
Logs are generally stored outdoors a minimum of 2 weeks, and a maximum of 8 weeks, prior to being treated with vacuum steam (Dossier Section 2).
Trees for the production of logs intended for export are not inspected for the presence of pests in the forest before harvest or at harvest with regard to regulatory requirements. Official inspection by a federal or state phytosanitary officer is conducted post‐vacuum steam treatment when a phytosanitary certificate is requested. The inspection is conducted to fulfil the requirements of the importing country (Dossier Section 2).
Vacuum–steam–heat treatment
3.4
The applicant proposes the following vacuum–steam–heat treatment as the only phytosanitary measure (Dossier Section 2) with the following requirements (Dossier Section 1):
Heating of the sapwood region at 56°C for 30 min at 5 cm depth measured from the cambium.
In a vacuum system, the steam can penetrate even small spaces very effectively. Steam heat contains 100 times more energy than air for the same volume. Conventional hot air causes evaporative cooling, resulting in heat loss that must be maintained by continually adding more dry air. With vacuum and steam, the wood heating is much quicker and efficient compared to dry air heating (Dossier Section 1).
Dossier Section 1 reports that trials were conducted with different hardwood logs to thermal map the logs as they reached 56°C for 30 min to the core. The logs were treated individually at 200 mmHg vacuum and 110°C steam in a flexible bag directly connected to a vacuum pump and boiler. In a second testing phase, the most efficient heating regime was found to be steam with 90°C and a vacuum of 570 mmHg.
Studies with logs and different pest species were conducted where infested logs were heated in a polypropylene bag and small steel chambers to achieve the required temperature of 56°C at 5 cm below the cambium for 30 min. The results are summarised together with information from other studies in Section 6.4.
The protocol for vacuum–steam–heat treatment suggested by the applicant in Dossier Section 1 includes the following:
-
Steam shall be saturated and below 100°C.
-
Initial chamber pressure shall be 100 mmHg (Torr) or less and maintained below 760 mmHg.
-
Treatment criteria mandate that the temperature of all log bark (if present) and sapwood be heated to at least 56°C and held for 30 min.
-
Basic equipment shall include a vacuum chamber, steam generator, vacuum pump and temperature monitoring system with controls.
-
–Logs stacked in a freight container or loose outside are placed into the vacuum chamber.
-
–Split reducers like metal connector plates or S irons should be applied to the ends of logs with visible splits.
-
–The temperature of the sapwood of three large logs, one on top, one in the middle and one at the bottom of the load will be selected for temperature monitoring.
-
–At least one, minimum 20 cm deep hole is drilled into the end of each log selected on the door end of the vacuum chamber. This hole should be 6 mm underneath the sapwood/heartwood boundary line. If the sapwood cannot be easily distinguished from the heartwood, then the hole will be drilled 5 cm below the cambium. The hole should follow and remain parallel to any observed log taper. The hole should preferably be small (6 mm diameter) and sized to friction fit the temperature sensor. If the logs are to be shipped with bark on, all temperature probes in the end of the log must always be placed so that they are in an area directly under the bark. This method ensures that measured sapwood heats at a rate that is commensurate with bark attached. If there is no bark on the end circumference of the log chosen, then another log with some or all bark attached to the surface must be used. After the sensor is installed in the log end, the hole around the sensor should be plugged with a plumber's type putty (clay like sealing material) to prevent steam incursion.
-
–The temperature inside the chamber shall be monitored at the top and bottom, along with front and rear locations with dedicated atmospheric sensors. Observed temperature gradients within the chamber should be limited.
-
–The temperature shall be monitored continuously during the course of the treatment.
-
–Before closing the chamber, the interior shall be inspected for safety. The chamber is then closed and sealed and a vacuum of at least 100 mmHg shall be created.
-
–Saturated steam at less than 100°C is then immediately introduced until all zones of the chamber reach the ambient treatment temperature of at least 85°C. Temperature gradients within the chamber should not exceed +/−5°C. Chamber pressure should never exceed 760 mmHg. When all three log end temperature sensors reach 56°C, the temperature should be maintained for 30 min, then equipment can be shut down.
-
–At the end of treatment, condensate must be collected and evaporated, recycled or drained into acceptable vegetative filtration strips, and then, a final vacuum of at least 100 mmHg shall be created to dry and cool the surface of the logs.
-
–Chamber pressure will then be immediately restored to atmospheric, and door(s) opened. Care should be exercised when handling any remaining hot surfaces.
-
–Loose logs can be removed and trans‐loaded into a freight container for shipment.
-
–For logs stacked in freight containers, the container doors can be closed and sealed for shipment. Note that the entire freight container is sanitised and should be pest free.
The concept of a final vacuum and steam system for log treatment was presented in the Dossier Section 2. However, the final device and studies confirming that the required efficacy will be achieved on larger scales with big loads of logs were not presented. In the assessment, it is assumed that the final device will have the same efficacy as the devices used in the studies provided.
Overview of interceptions
3.5
Data on the interception of harmful organisms on Quercus and Juglans wood can provide information on some of the organisms that can be present on wood logs.
According to EUROPHYT (2025) and TRACES‐NT (2025) (accessed: 26 August 2025), there were 224 interceptions of wood and bark/logs/sawn wood of Quercus and Juglans species from the US due to the presence of harmful organisms (see Table 4) between the years 1995 and July 2025.
IDENTIFICATION OF PESTS POTENTIALLY ASSOCIATED WITH THE COMMODITY
4
The search for EU quarantine pests and protected zone quarantine pests associated with oak and walnut rendered 368 pests. Many of these pests are regulated as groups of species (e.g. Scolytinae spp. (non‐European), Cronartium spp., etc.) by the Commission Implementing Regulation (EU) 2019/2072 (see Microsoft Excel® file in Appendix D). Altogether, 58 pests including pests regulated as individual species and pests regulated as groups of species were evaluated.
Relevant EU‐quarantine pests potentially associated with the commodity
4.1
In total, 58 EU quarantine pests are reported to be associated with Quercus and/or Juglans (Table 5). Of these 58 EU quarantine pests evaluated, the following are present in the US and can be associated with the stems used for log production and hence were selected for further evaluation: Anoplophora glabripennis, Arrhenodes minutus, Bretziella fagacearum, Cryphonectria parasitica, Davidsoniella virescens, Entoleuca mammata, Euwallacea fornicatus sensu lato, Geosmithia morbida, Lopholeucaspis japonica, Lycorma delicatula, Neocosmospora euwallaceae, Phytophthora ramorum, Pityophthorus juglandis, Pseudopityophthorus minutissimus, Pseudopityophthorus pruinosus, Scolytinae spp. (non‐European) and Xylella fastidiosa.
Summary of pests selected for further evaluation
4.2
The 17 pests satisfying all the relevant criteria listed above in Section 4.1 are included in Table 6.
THE PESTS ASSESSED IN THE OPINION
5
Bretziella fagacearum
5.1
Taxonomy
5.1.1
Bretziella fagacearum is a fungus belonging to the phylum: Ascomycota; order: Microascales; and family: Ceratocystidaceae (Index Fungorum, 2025).
The synonyms are Ceratocystis fagacearum and Endoconidiophora fagacearum (Index Fungorum, 2025). Taxonomic synonyms according to Mycobank (2025) are Chalara quercina and Thielaviopsis quercina.
The English common names of B. fagacearum are oak wilt and wilt of oak (EPPO, 2025b).
Origin and distribution
5.1.2
Bretziella fagacearum was initially described in 1940 as Chalara quercina (the asexual stage) by Henry (1944) from Illinois, Iowa, Minnesota and Wisconsin. Bretz (1953) later identified and named its sexual state, Endoconidiophora fagacearum. The fungus was subsequently renamed Ceratocystis fagacearum (Hunt, 1956). However, a recent phylogenetic reclassification of Ceratocystidaceae, based on multiple genes, revealed that the oak wilt fungus did not belong to any of the established genera. Consequently, it was classified as a separate species and was renamed Bretziella fagacearum to honour Bretz, who first described its sexual stage (de Beer et al., 2017).
The pathogen is only known to be present in the US and Canada (EPPO, 2025c). In Canada, it was reported for the first time in Niagara Falls, Ontario, in June 2023 and it is under official control there (EPPO, 2023a; North American Plant Protection Organization, 2023).
The origin of B. fagacearum is unknown. According to Juzwik et al. (2008), there are two possible scenarios of the origin of the pathogen: (1) native from North America; or (2) non‐native originated from Central or South America or Mexico. However, the weight of evidence indicates that B. fagacearum is an introduced pathogen in North America.
Presence, distribution and prevalence of the pest in the US
5.1.2.1
Bretziella fagacearum has been reported in the US in the following states: Arkansas, Illinois, Indiana, Iowa, Kansas, Kentucky, Louisiana, Maryland, Michigan, Mississippi, Missouri, Nebraska, New York, North Carolina, Ohio, Oklahoma, Pennsylvania, South Carolina, South Dakota, Tennessee, Texas, Virginia, West Virginia and Wisconsin (USDA, 2025). The distribution data are available by county in the Forest Service USDA website – the most recent distribution map is from October 2023, with the previous county detections, including the damage caused from 2021 to 2023 (USDA, 2025) (Figure 1).
Distribution of Bretziella fagacearum in the US. Map updated in 2023, provided by USDA (2025).
Damage of the pathogen between 2021 and 2023 (Figure 1) was detected in Illinois, Indiana, Iowa, Michigan, Minnesota, New York, Ohio, Pennsylvania, Texas and Wisconsin (USDA, 2025).
Dossier Section 2 states that: ‘maximum prevalence in terms of number of individual diseased trees in a stand is difficult to estimate. Numbers of dying or dead Quercus rubra in a pocket partly depends on how long the disease has been present in the stand, soil type and topography. Stands with dense red oak species on course textured, sandy soils exhibit the highest rates of disease incidence due to higher rates of root graft formation among neighbouring red oak trees. Quercus rubra stands selected for harvest of high‐quality logs are on sites with heavier textured soils and lower rates of disease transmission. Maximum prevalence in Q. alba stands is generally very low due to lower rates of disease transmission among white oak species.’
Biology
5.1.3
Bretziella fagacearum is a causal agent of oak wilt. The pathogen causes a vascular wilt by colonising the sapwood of the trees, which may result in a brownish discoloration of the xylem, visible in cross sections (branches, stem) of wilted trees (EFSA PLH Panel, 2020b). The fungus develops mycelial mats under the bark of recently killed trees on which spores (i.e. conidia first and ascospores later) are produced. As the fungal mats grow, the bark is pushed away and cracks open. The fruit‐like odour emitted by the mats attracts sap beetles (Coleoptera, Nitidulidae), which may subsequently carry fungal spores of the pathogen to other oak trees (Harrington, 2013). Fresh, xylem‐penetrating wounds, leaking sap, generally less than 72 h old, are required for successful infection mediated by sap beetles (Kuntz & Drake, 1957). Wounds created by human activities (cut branch ends, fresh stump surfaces, stem wounds) or strong winds (broken branches and stems) may provide suitable infection courts, particularly in spring and early summer (Juzwik et al., 2011). Bark wounds are required for infection because sap beetles are unable to bore intact bark or woody plant tissues.
Sexual ascospores can stick to the tegument of insects and are more effectively dispersed than conidia. Bretziella fagacearum is heterothallic and can only reproduce sexually upon mating of two strains with opposite mating types. This may occur when sap beetles are visiting different mats and cross‐fertilise the fungus (Harrington, 2013).
Sporulating mats are only produced when the bark/wood interface is moist (Gibbs & French, 1980). There is a fairly narrow range of sapwood moisture content (37%–45% in spring, 44%–52% in autumn) that allows for fungal mat formation (Campbell & French, 1955). Mats are produced in a temperature range of 8–25°C with faster and larger development in warmer conditions (EFSA PLH Panel, 2020b). Bretziella fagacearum is poorly competitive as a saprophyte and is rapidly replaced by many other organisms within 1 year after the death of the tree (Gibbs & French, 1980; Sinclair & Lyon, 2005). Sporulating mats are important for the spread of the pathogen by insect vectors. Mats may also form on firewood and logs, on which the disease can be transported to new areas (Juzwik et al., 2011). Spread of oak wilt within a forest stand mostly occurs through root grafts between trees of the same oak species (Appel, 1995; Bruhn et al., 1991). Root grafting may also occur between trees of different oak species, but the importance of this type of grafting for disease transmission is unknown (Juzwik, 2008). Root graft transmission results in distinct disease foci, which can be observed in forest stands as clusters of symptomatic and killed oak trees. Bretziella fagacearum can infect many oak species, which exhibit different levels of susceptibility or resistance; see Section 5.1.5 for more details.
Ability to create resting propagules
5.1.3.1
Bretziella fagacearum produces hyphae, mycelium, conidiophores, conidia, endoconidiophores, endoconidia, ascospores, asci, perithecia and sclerotia (de Beer et al., 2017; EPPO, 2023b). There is no indication that the fungus can create resting propagules or chlamydospores.
Effect of temperature on the biology of the pest
5.1.4
Numerous studies have examined the thermal tolerances of this pathogen, revealing the specific temperature ranges that influence its growth, reproduction and persistence in various substrates, from laboratory cultures to infected oak wood.
On agar plates, the growth of B. fagacearum isolates was best between 22 and 26°C, poor at 16°C and 32°C and none above 34°C (Houston & Kuntz, 1960; Lewis, 1985). While no growth was observed, the isolates survived for 10 days at 34°C (Lewis, 1985).
In laboratory settings using Petri dishes, perithecial production occurred within a temperature range of 15°C–27°C, with an optimal temperature of 24°C. No perithecia developed at 10°C or 30°C (Cobb Jr et al., 1961).
Ascospores and endoconidia germinated between 3°C and 36°C, with an optimal germination temperature range of 21°C–32°C under laboratory conditions (Fergus, 1954; Tainter, 1986).
Both spore types survived longest under cool, dry conditions (McLaughlin & True, 1952). High temperature and humidity drastically reduced viability; at 37°C and 95% relative humidity, endoconidia survived less than 3 days and ascospores only 3 days; however, at 25%, both lasted 10 days. Overall, ascospores proved to be more resilient than endoconidia (Merek & Fergus, 1954).
According to Bretz and Morison (1953), survival of B. fagacearum in small diameter material (twigs, branches) is relatively short at temperatures of 20°C–25°C and above.
The viability of B. fagacearum in air‐dried oak lumber varied with harvest season (temperature and moisture) and oak species (Q. rubra, Q. coccinea and Q. alba). In summer‐harvested lumber, viability ceased after 14 days. However, B. fagacearum can persist longer in autumn‐harvested lumber (up to 140 days). Quercus alba generally exhibited lower recovery rates compared to Q. rubra and Q. coccinea. Air‐drying lumber to a moisture content of 20% or less resulted in a significant reduction in fungal viability (Tainter et al., 1984).
Host range and host status of Quercus and Juglans
5.1.5
Quercus, Castanea, Chrysolepis and Notholithocarpus are the only broadleaf trees known to be hosts of B. fagacearum (EPPO, 2025d). There is no information on whether B. fagacearum can also attack Juglans.
Quercus natural hosts (North American species) of B. fagacearum are Q. agrifolia, Q. alba, Q. chrysolepis, Q. coccinea, Q. dumosa, Q. ellipsoidalis, Q. engelmannii, Q. falcata, Q. fusiformis, Q. garryana, Q. imbricaria, Q. kelloggii, Q. laevis, Q. laurifolia, Q. lobata, Q. macrocarpa, Q. marilandica, Q. muehlenbergii, Q. nigra, Q. palustris, Q. phellos, Q. prinus, Q. rubra, Q. shumardii, Q. stellata, Q. texana, Q. velutina, Q. virginiana and Q. wislizenii (EFSA, 2022).
Quercus experimental hosts (European species) of B. fagacearum are Q. ilex, Q. petraea, Q. pubescens, Q. robur and Q. suber (EFSA, 2022; EFSA PLH Panel, 2018b; Pinon et al., 2003).
Quercus genera is divided into two subgenera (Quercus and Cerris) and additional sections (Hipp et al., 2020). Red oaks, section Lobatae (Q. agrifolia, Q. coccinea, Q. ellipsoidalis, Q. falcata, Q. imbricaria, Q. kelloggii, Q. laevis, Q. laurifolia, Q. marilandica, Q. nigra, Q. palustris, Q. phellos, Q. rubra, Q. shumardii, Q. texana, Q. velutina and Q. wislizenii), are highly susceptible to oak wilt (Juzwik et al., 2011) and can die within few weeks after being infected (EPPO, 2021a; Sinclair & Lyon, 2005). White oaks, section Quercus (Q. alba, Q. dumosa, Q. engelmannii, Q. garryana, Q. lobata, Q. macrocarpa, Q. muehlenbergii, Q. prinus, Q. stellata), are from highly (Q. alba) to moderately resistant to oak wilt (Juzwik et al., 2011). In Q. alba, infections by B. fagacearum may result in dieback of a few branches, but the trees can survive for many years (Juzwik et al., 2011). This is because the trees can produce new annual rings of sapwood and compartmentalise the fungus. Thus, the vascular staining associated with the fungus is observed deeper in the sapwood (EPPO, 2021a). Southern live oaks, section Virentes (Q. fusiformis, Q. virginiana), have moderate resistance to oak wilt (Juzwik et al., 2011). Intermediate oaks, section Protobalanus (Q. chrysolepis), have susceptibility between red and white oaks (Gearman & Blinnikov, 2019 citing others). European species (Q. robur, Q. petraea, Q. pubescens) were susceptible and died within 1 year following the inoculation experiment (MacDonald et al., 2001).
Symptoms and diagnosis
5.1.6
The oak wilt symptoms are not visible during the dormant stage. Identification of oak wilt‐infected trees is more reliable for red oaks, where symptoms develop rapidly. In the white oak (Q. alba), identification of infected trees can be more difficult because of the slow development of disease symptoms. The fungus can remain undetected for many years (e.g. 20‐year infection observed in a Q. alba tree) (EFSA PLH Panel, 2020b).
Symptoms of B. fagacearum on Quercus species are crown and foliage wilting; death of foliage; water‐soaked appearance of mature leaves; bronzing and necrosis of leaf tips and margins; leaf abscission of completely green leaves; branch dieback; vascular straining in the xylem of branches and main stem; presence of mats; and death of trees (Henry, 1944; Juzwik et al., 2011).
Fungal mats are usually absent or rare on white oaks (Cones, 1967; Engelhard, 1955). Mats develop almost exclusively on red oaks, with a proportion of one‐third of infected trees actually producing mats that rupture the bark, based on a mat survey of standing dead trees (EFSA PLH Panel, 2020b).
Diagnosis of B. fagacearum is done either by culture‐based or molecular methods using wood samples from symptomatic trees (EPPO, 2023b; Yang & Juzwik, 2017). Recently, a novel non‐destructive detection method has been reported, which is based on a real‐time PCR assay using leaf petioles from fallen leaves (Chahal et al., 2025).
Pathway of entry with logs
5.1.7
Main pathways of entry for B. fagacearum are (1) wood with and without bark; (2) isolated bark; (3) plants for planting other than seeds; and (4) cut branches (EFSA PLH Panel, 2018b).
Wood from diseased oak trees presents the most likely pathway for B. fagacearum introduction (EPPO, 1997; Robinet et al., 2016), and sporulating mycelial mats on logs further amplify the risk of its spread (Juzwik et al., 2011). Wood both with and without bark is a potential carrier of the fungus (EFSA PLH Panel, 2018b), with fungal survival dependent on factors like temperature, moisture content, harvest time and oak species (Tainter et al., 1984). This fungus has shown remarkable persistence, surviving for up to 24 weeks in sawn lumber (Gibbs & French, 1980) and for up to 140 days in autumn‐harvested, air‐dried oak lumber (Tainter et al., 1984). Bretziella fagacearum can also survive at least 4 weeks in heartwood and 12 weeks in sapwood of white and black oaks (Partridge, 1961). In branches of Q. ellipsoidalis from Minnesota, the fungus in trees that died later in the summer survived sometimes even until the following spring (Gibbs, 1980).
Pityophthorus juglandis and Geosmithia morbida
5.2
Taxonomy
5.2.1
Pityophthorus juglandis is a bark beetle belonging to the order: Coleoptera and family: Curculionidae. The English common name is Walnut Twig Beetle (WTB).
Geosmithia morbida is a fungus belonging to the order: Hypocreales and family: Bionectriaceae. The English common name is Thousand Cankers Disease (TCD).
Origin and distribution
5.2.2
Pityophthorus juglandis is a bark beetle, the main vector of the pathogenic fungus Geosmithia morbida. It is endemic to Mexico and the SW states of the US (Gomez et al., 2023), but it is also present in north‐western and eastern US, where it has been introduced probably via wood trade (Newton & Fowler, 2009; Sitz et al., 2021). Outside of North America, P. juglandis was reported in Europe for the first time in 2013 in Italy (Montecchio & Faccoli, 2014), where it is currently present in Veneto, Piedmont, Lombardy, Emilia Romagna, Friuli Venezia Giulia (Bracalini et al., 2023) and Tuscany (Pennacchio et al., 2023). In 2022, the pest was also found in France (Saurat et al., 2023).
Geosmithia morbida was described by Kolařík et al. (2011) as native to Western North America (mostly California and Colorado). Its distribution in North America almost completely overlaps that of the main vector insect P. juglandis except for Mexico (Chihuahua), where only the beetle was recorded (EPPO, 2015). The pathogen is present in Italy wherever its vector is found (see above) with the sole exception of Friuli Venezia Giulia (Bracalini et al., 2023). In France (Auvergne‐Rhone Alpes), both the fungus and the vector are present (Saurat et al., 2023).
Presence, distribution and prevalence of the pest in the US
5.2.2.1
In the US, the symbiotic complex Pityophthorus juglandis/Geosmithia morbida is currently reported in the following 18 states: Arizona, California, Colorado, Idaho, Indiana, Maryland, Michigan, Missouri, Nevada, New Mexico, North Carolina, Ohio, Oregon, Pennsylvania, Tennessee, Utah, Virginia and Washington (Atkinson, 2025; Bright, 2021; DAFNAE, 2025; EPPO, 2025e). Besides, Geosmithia morbida is also present in Illinois and Minnesota without P. juglandis being found (Moore et al., 2019). In the US, the detection of P. juglandis is usually considered equivalent to that of G. morbida (Cranshaw & Tisserat, 2012), but the pathogen may also be present in the absence of the beetle or walnuts symptomatic for TCD (Moore et al., 2019). However, only in the states where both the pests are recorded infestations have been established, mostly on amenity trees in urban areas, agricultural landscape and orchards. Considering the widespread presence of several susceptible hosts, the potential alternative vectors (Section 5.2.3), the passive human‐assisted transport on infected wood despite containment measures, it is likely that the pests are also present in other states bordering those where they are currently established and maybe more widespread in the US than previously known (EPPO, 2015; Moore et al., 2019).
The distribution data are available by county in the Forest Service USDA website – the most recent distribution map is from October 2023, with the previous county detections (USDA, 2025) (Figure 2).
Distribution of Geosmithia morbida in the US. Map updated in 2023, provided by USDA (2025).
The pathogen based on Figure 2 was detected in these states: Arizona, California, Colorado, Idaho, Maryland, New Mexico, North Carolina, Ohio, Oregon, Pennsylvania, Tennessee, Utah, Virginia and Washington (USDA, 2025).
Dossier Section 2 states that: ‘the prevalence of G. morbida in natural forest stands in the eastern and southern US is considered very low or insignificant. Populations of the primary vector of G. morbida (i.e. walnut twig beetle) collapsed in Ohio, Pennsylvania, Maryland and Virginia between 2013 and 2019) and no new detections/reports in southern states between 2012 and 2024. In western states, losses are most serious in urban and community forests (e.g. CA, WA) and in timber plantations of J. nigra (outside of native range) in eastern Washington State (max. prevalence, 80%).’
Biology
5.2.3
Pityophthorus juglandis is a small bark beetle (adults 1.5–2 mm long) belonging to the group of ‘twig beetles’ typically infesting branches of trees (usually over 1.5–2 cm in diameter) as well as stems attacked by other bark beetles (EPPO, 2015; Smith & Hulcr, 2015). Pityophthorus juglandis has four life stages: egg, larva (3 instars), pupa and adult. Depending on the latitude and climatic conditions, one to three overlapping generations per year are observed in the native range (Bracalini et al., 2023; EFSA, 2020a). In North Italy, only two generations per year are observed (Faccoli et al., 2016). From spring to autumn, the swarming adults are mostly attracted by stressed trees, but healthy plants can also be infested (EPPO, 2015). Suitable hosts are found not only via host volatiles and aggregation pheromones but also via Geosmithia volatiles, confirming a close relationship between insect and fungus (Blood et al., 2018). Both young and mature trees can be attacked on stems of all sizes and branches not thinner than 1.3–2.0 cm (EFSA, 2020a). Pityophthorus juglandis is a polygamous species and the circular nuptial chamber created by the male beneath the bark can host from two to eight females (Cranshaw & Tisserat, 2012; Faccoli et al., 2016). The larvae spread in the phloem by tunnelling winding galleries that never engrave the sapwood and develop to the pupal stage in 4–8 weeks in a temperate climate (EFSA, 2020a). The emerging adults usually colonise new hosts near the natal tree, having limited flight capacity (maximum distance 3.6 km in 24 h, with 1/3 of the beetles flying < 100 m) (Kees et al., 2017). However, dispersal by air currents may also play a role, and even more the human‐assisted spread with transport of fresh wood (mostly logs and firewood) (EFSA, 2020a). Pityophthorus juglandis overwinters in the infested trees as mature larvae, pupae and adults, which emerge in late April (Cranshaw & Tisserat, 2012).
The bark beetles may be present in large numbers on infested logs (densities up to 5–6 individuals/cm^2^) (EPPO, 2015). Although adults of P. juglandis do not have mycangia, the vectoring of G. morbida is still very efficient because the elytra of emerging beetles are heavily contaminated by spores of the pathogen (Cranshaw & Tisserat, 2012; Newton & Fowler, 2009). However, P. juglandis is not the only vector of G. morbida in the US, as other 11 beetle species are known to carry propagules of the pathogen; of these, eight species are ambrosia beetles (native and non‐native), two species are native bark beetles and one species is a bark weevil (Chahal, 2018). Moreover, Moore et al. (2019) have detected G. morbida in 18 insect species using molecular methods. Even if the role of the alternative vectors needs to be further investigated, it is suspected that they may sustain the establishment of the disease and contribute to the spread of the pathogen (Chahal et al., 2019).
Geosmithia morbida is an ascomycete reproducing only asexually via conidia. Once carried beneath bark by the beetles, conidia germinate producing a hyaline/whitish mycelium that slowly grows degrading cellulose and lignin of the cell walls, hence causing tissue necrosis in the phloem (the sapwood is only superficially reached) and disrupting vascular function. Soon conidia appear on verticillate conidiophores, contaminating the body of adult beetles of the new generation which then spread the infection to new hosts (Bracalini et al., 2023; EPPO, 2015; 2020a).
Ability to create resting propagules
5.2.3.1
There is no evidence in the literature that G. morbida produces resting propagules (Bracalini et al., 2023; EFSA, 2020a; EPPO, 2015; 2020a; Kolařík et al., 2011). The absence of documented resting structures suggests that G. morbida primarily relies on its association with P. juglandis for dispersal and survival.
Effect of temperature on the biology of the pests
5.2.4
Optimal growth temperature of G. morbida is around 31°C, with a stop of growth in the lab beyond 35°C. The fungus is reported to survive up to 41°C and can maintain its viability in inoculated wheat seeds at 48°C (Bracalini et al., 2023); however, in walnut logs, G. morbida does not survive at 48°C or higher (Mayfield et al., 2014). No specific information was found about the tolerance of G. morbida to low temperatures; however, considering its strong association with the vector, it can be assumed that it is the same of the beetle. Pityophthorus juglandis has a remarkable cold tolerance, as not only adults but also larvae and pupae can be found under the bark in cold winters. The lower lethal temperatures range from −14°C to −23°C for adults (Hefty et al., 2017) and −16.9°C for larvae (Luna et al., 2013). As for high temperatures, the lethal threshold is 47.9°C for adults and 47.3°C for larvae (Luna et al., 2013), and a minimum sapwood temperature of 56°C for 40 min completely eliminates both the pathogen and the vector from walnut logs (Mayfield et al., 2014).
Host range and host status of Quercus and Juglans
5.2.5
Pityophthorus species feeding on pines in the US are very frequently associated with Geosmithia sp. (Kolařík et al., 2017). Out of conifers, however, no other broadleaf trees than Juglans and Pterocarya are known to be hosts for Pityophthorus in North America (Wood & Bright, 1992). Therefore, Quercus is not a host of P. juglandis in the US. As for G. morbida, Juglans and Pterocarya are also the sole hosts (EPPO, 2015).
Juglans hosts of both P. juglandis and G. morbida are J. ailanthifolia (syn. J. mandshurica var. sieboldiana), J. californica, J. cinerea, J. hindsii, J. major, J. mandshurica, J. microcarpa, J. mollis, J. nigra and J. regia (Atkinson, 2025; DAFNAE, 2025; EPPO, 2015; EPPO, 2025f, 2025g; Farr & Rossman, 2025). Pterocarya hosts are P. fraxinifolia, P. rohifolia and P. stenoptera for both the pests (Atkinson, 2025; DAFNAE, 2025; EPPO, 2020a, 2025f, 2025g).
Juglans hosts show increasing susceptibility from J. major to J. nigra, the latter being the most severely affected host (EFSA, 2020a; EPPO, 2015; 2020a). The other species, as well as their hybrids (J. hindsii x J. regia, J. nigra x J. regia, etc.) and the three species of Pterocarya show intermediate susceptibility levels in both the field and lab inoculation tests (EFSA, 2020a; EPPO, 2015). There is uncertainty about grafted trees, which show susceptibility considerably varying with the rootstock, and also on the susceptibility of Juglans regia, due to the wide intraspecific variations of the species (Bracalini et al., 2023; EPPO, 2015).
Symptoms and diagnosis
5.2.6
The Thousand Cankers Disease of walnuts may be recognised by a combination of beetle infestation and fungus infection symptoms, primarily shown by foliage yellowing/wilting of individual branches eventually leading to progressive canopy loss. However, these are nonspecific symptoms, similar to those caused by other abiotic stress factors, such as drought. Specific external symptoms on stems and branches are the entry/exit holes of adult beetles together with cankers due to the spread of the pathogen. Entry holes can be associated with sap weeping. Adults and immature stages of P. juglandis can be observed under the bark in the galleries. Although the fungus has no systemic spread within the host, several dark‐brown cankers caused by G. morbida infection appear and rapidly merge on infected tissues of phloem and superficially the outer wood, eventually leading the host to death within 3–8 years (Bracalini et al., 2023; EFSA, 2020a; EPPO, 2020a). However, the symptomatology of TCD may vary with the different susceptibility of hosts to the pathogen. In low and intermediate‐susceptible hosts, as Juglans major, J. regia, J. cinerea, J. californica, etc. and all the hybrids, scattered dieback and low progression of the disease are recorded, and the death of trees is rare. Large canker incidence and high mortality are only found in Juglans nigra (EFSA, 2020a).
Both G. morbida and its vector P. juglandis can be identified through morphological characteristics and DNA sequencing of specific genomic regions. Molecular identification of G. morbida from cultured colonies can be performed using species‐specific primers (Moore et al., 2019) or universal primers targeting the internal transcribed spacer (ITS) region (Moricca et al., 2020; Saurat et al., 2023). Identification of P. juglandis can be achieved by sequencing partial fragments of the mitochondrial cytochrome c oxidase I (COI) gene (Moricca et al., 2020). Species‐specific PCR protocols, including simplex and duplex qPCR assays, were also developed to detect G. morbida from woody tissues and bark beetles as well as P. juglandis from insect frass (Rizzo et al., 2020).
Pathway of entry with logs
5.2.7
Main pathways of entry for both P. juglandis and G. morbida are wood with bark of Juglans and untreated packaging wood of Juglans, for which the probability is considered by EPPO (2015) very high with low uncertainty.
From moderate to low probability of entry are listed the following commodities of Juglans and Pterocarya: wood without bark, plants for planting, scion wood, bark, particle wood and non‐agglomerated waste wood of deciduous trees. Finally, squared wood of Juglans and Pterocarya, wood packaging material ISPM 15 treated and agglomerated waste wood of deciduous trees fall in the group of very low probability of entry (EPPO, 2015).
Other target pests
5.3
All the information on the additional EU quarantine pests relevant for this opinion are summarised in Appendix A.
ASSESSMENT OF THE VACUUM–STEAM–HEAT TREATMENT
6
International and national standards on heat treatment
6.1
Heat treatment is a widely recognised phytosanitary method used to eliminate wood‐dwelling pests. It is a key component of international and national wood treatment standards. The temperature and duration of exposure proposed by the applicant correspond to the requirements of ISPM 15. The standard ISPM 15 mandates heating of wood packaging to a core temperature of 56°C for 30 mins, which is generally effective against most wood pests. However, some pests may survive this treatment (e.g. Ramsfield et al., 2010) prompting certain countries to adopt stricter import requirements (Allen, 2014). For example, New Zealand requires heating of sawn wood to a core temperature of 100°C for 30 min and Australia requires heating of timber from some genera to a core temperature of 74°C (see Table 7).
Pattern of temperature distribution in heat‐treated logs
6.2
Knowledge on the patterns of distribution of temperature in the cross section of heat‐treated logs is pivotal to determine whether areas of the sections are less exposed to high temperatures. By working on logs of Q. rubra (43.9–56.4 cm diameter) treated with vacuum steam, Juzwik et al. (2019) determined that the average time for probes placed 5 cm below the cambium to reach 56°C and hold for 30 min was 6.9 h. It should be noted that at this time frame, based on the temperature profiles reported in the study (Juzwik et al., 2019), the temperature at the centre of the section did not increase substantially and was similar to the ambient temperature (see Figure 3).
In another study focused on vacuum steam treated Juglans nigra logs, the cycle times for 56°C for 30 min varied from 5.0 to 7.7 h depending on the log (Juzwik et al., 2021). Based on the temperature profiles reported in the study (Juzwik et al., 2021), even after 7.7 h, the temperature at the centre of the section did not exceed 45°C.
Both for Q. rubra and J. nigra, the temperature reached at the external surface and at the cambium layer was substantially higher than that measured 5 cm below the cambium (Juzwik et al., 2019, 2021).
Temperature profiles of (A) 56°C for 30‐min treatment of Q. rubra log and (B) 60°C for 60 min at an initial vacuum of 100 mmHg. The temperatures were measured at different depths of the wood and at different distances from the end of the log (Juzwik et al., 2019).
General information on temperature limits of pests
6.3
A literature review on the temperature limits on a wide range of wood‐dwelling insects, fungi and bacteria was conducted in a previous EFSA opinion (EFSA PLH Panel, 2024a). The information retrieved suggests that a temperature of 60°C for 30 min is sufficient to eliminate all wood‐boring insects except powder post beetles where a lethal temperature of 82°C was reported. The lethal temperature of some fungal species was higher than that of insects. For example, Tansey (1971) and Schmidt (2007) reported lethal temperatures of > 82°C for short‐term exposure for some fungal species. The eradication temperature (99.99% mortality) for 11 investigated wood‐colonising fungal species (30 min exposure) ranged from 41.6°C to 69.6°C (Ramsfield et al., 2010).
Specific information on temperature limits of survival of relevant pests
6.4
Temperature limits of survival for B. fagacearum
6.4.1
The mycelium of B. fagacearum did not survive exposure to a temperature of 45°C for 4 h, but it survived 45°C when exposed for only 1 h (Lewis, 1985). The mycelium survived 46°C and died at 48°C when exposed to the heat for 30 mins (Noseworthy et al., 2024). Ascospores are more heat tolerant and survived exposure to 50°C for 2 h and were killed when exposed for 6 h to 50°C (Cole & Fergus, 1956).
Juzwik et al. (2019) investigated the survival of B. fagacearum in logs. Naturally and artificially infested red oak logs were vacuum–steam–heat treated at 56°C for 30 min and 60°C for 60 min held at a depth of 5 cm below the cambium. No living inoculum was found in the samples taken after the treatments.
However, in a second study with red oak logs (Juzwik, unpublished, information included in the Dossier), using the same methodology as described above, two samples (230 samples were positive out of 480 pretreatment samples and two samples were positive out of 480 post‐treatment samples) in the 60°C treatment tested positive for surviving B. fagacearum: one sample from the inner sapwood and one from the outer sapwood. It should be noted that these two positive samples came from two different logs out of a total of 15 treated logs. No positive samples were found in the logs treated at 56°C (253 samples were positive out of 480 pretreatment samples and no samples were positive out of 480 post‐treatment samples).
A similar study was conducted with artificially inoculated Q. alba. However, there was a very low number of positive samples found in logs before treatment (0.63% were positive out of 792 samples). Therefore, the result of zero positive samples after treatment cannot be used as strong evidence of the efficacy of the treatment.
Available information suggests that an exposure to a temperature of 56°C is generally sufficient to kill B. fagacearum mycelium. Ascospores are more heat tolerant and require 6 h of exposure to 50°C to ensure their inactivation. Ascospores are produced at the surface of the wood and hence will be exposed to higher temperatures than 56°C. Surviving B. fagacearum was found in an experiment with the suggested vacuum–steam–heat treatment of logs at a temperature of 60°C. Therefore, it seems that the proposed methodology is not always sufficient to kill all B. fagacearum.
Temperature limits of survival for Geosmithia morbida and Pityophthorus juglandis
6.4.2
The LT_99_ of larvae and adults of P. juglandis was 48.1°C and 52.7°C, respectively, for an exposure duration of 30 mins in plastic tubes (Luna et al., 2013). No larvae survived exposure in logs at temperatures of 50.1°C for 30 min at 3.8 cm depth below the cambium (Mackes et al., 2016).
No viable mycelium of G. morbida was found in J. nigra logs exposed to 48°C for 40 mins at 1 cm below cambium and no viable larvae of P. juglandis were found after exposure to 52°C for 40 min at 1 cm below cambium (Mayfield et al., 2014). Juzwik et al. (2021) reported no viable mycelium of G. morbida and no viable larvae of P. juglandis in J. nigra logs after exposure to 56°C for 30 min measured at 5 cm depth below the cambium. Some surviving G. morbida inocula were detected when the above temperatures per time exposures were applied at 3.2 cm depth below the cambium instead of 5 cm depth below the cambium.
The available information suggests that G. morbida and P. juglandis would not survive exposure to 56°C for 30 min granted that the required temperature is reached in all areas of the log where the pests can be found.
Temperature limits of survival for other relevant pests
6.4.3
Larvae of Anoplophora glabripennis did not survive exposure to a temperature of 57.8°C for 30 min, measured at the centre of heat‐treated logs (Myers & Bailey, 2011). Eggs of Lycorma delicatula withstood 50°C for 30 min; however, no eggs survived 55°C for 15 min (Zandi‐Sohani et al., 2025). For the two wood borer species, Arhopalus ferus and Hylurgus ligniperda, the lethal temperatures (LT 99.99) for a 30‐min exposure ranged from 46.9°C (larvae of H. ligniperda) to 57.4°C (eggs of A. ferus). The lethal temperatures for other life stages fell between these extremes – for example, 52°C and 53.3°C for larvae, and 51°C and 55.8°C for adults of A. ferus and H. ligniperda, respectively (Pawson et al., 2019). Concerning ambrosia beetles, Anisandrus dispar is reported not to survive temperatures exceeding 50°C for 15 min in laboratory tests (Noseworthy et al., 2023) and Xylosandrus germanus is reported not to survive 58°C for 1 min (Suh, 2014).
Further information on the temperature limits of survival of other groups of pests can be found in Appendix A. For some relevant pests (i.e. A. minutus, D. virescens, E. mammata, N. euwallaceae), no information on the temperature of survival was found. However, there is no evidence in support of a higher temperature limit of survival for these pests.
Depth of wood colonised
6.5
The depth of the wood which is colonised by the target organisms is crucial for estimating the efficacy of the proposed treatment. In case that the target organisms are found deeper than 5 cm below the cambium, they will not be exposed to the proposed temperature of 56°C for 30 min and hence may be able to survive the treatment.
According to the Dossier Section 2, B. fagacearum is not known to colonise heartwood because of the low moisture content of heartwood. Bretziella fagacearum colonises the whole circumference of sapwood and can colonise towards the sapwood–heartwood boundary in Q. rubra. Quercus alba has deeper sapwood than Q. rubra (see below) and B. fagacearum may grow deeper in the sapwood where the treatment will be less effective. In Q. alba, infections by B. fagacearum could remain asymptomatic for many years because the trees can produce new annual rings of sapwood and compartmentalise the fungus (Juzwik et al., 2011). Thus, the vascular staining associated with the fungus is observed deeper in the sapwood (EFSA, 2020b).
Data on the depth of sapwood in Q. rubra and Q. alba were submitted in the Dossier Sections 1 and 2. Measurements were conducted on 348 Q. rubra and 235 Q. alba logs. In total, 75% of the measurements resulted in a sapwood width of ≤ 2.11 cm in Q. rubra and ≤ 2.7 cm in Q. alba. However, sapwood wider than 5 cm was found in one Q. rubra and in four Q. alba logs. No data on sapwood depth were available for J. nigra.
While, in most cases, the heating of logs to a depth of 5 cm below the cambium is sufficient to cover the whole sapwood of the logs in few logs the sapwood was wider. Target organisms dwelling in wood deeper than 5 cm may not be exposed sufficiently long and not to a sufficiently high temperature to prevent their survival.
Uncertainty analysis of literature data
6.6
The uncertainty of the different studies is mainly determined by the number of individuals or amount of isolates (proxy of inoculum) of a pest exposed to the heat treatment in each experiment. To quantify the remaining uncertainties for each survival estimator the 95% confidence interval (see below Table 8) was calculated. In case of zero survival, a one‐sided interval was used; otherwise, the two‐sided interval was calculated.
In the following figures (Figures 4, 5, 6), the results are plotted against the temperature of each treatment.
** Bretziella fagacearum **
The analysis of the results of studies with B. fagacearum is presented in Table 9 and Figure 4.
95% confidence intervals for the estimate of survival of B. fagacearum for different temperatures in different studies. The interval starts at 0% (one‐sided) in case no pest survived in the experiment; otherwise, the two‐sided interval was calculated. Results of Juzwik et al. (2019) relate to measurements in inner sapwood. Letters indicate the specific study reported in the reference.
** Geosmithia morbida **
The analysis of the results of studies with G. morbida is presented in Table 10 and Figure 5.
95% confidence intervals for the estimate of survival of G. morbida for different temperatures in different studies. The interval starts at 0% (one‐sided) in case no pest survived in the experiment; otherwise, the two‐sided interval was calculated. Letters indicate the specific study reported in the reference.
** Pityophthorus juglandis **
The analysis of the results of studies with P. juglandis is presented in Table 11 and Figure 6.
95% confidence intervals for the estimate of survival of P. juglandis for different temperatures in different studies. The interval starts at 0% (one‐sided) in case no pest survived in the experiment; otherwise, the two‐sided interval was calculated. Letters indicate the specific study reported in the reference.
QUANTITATIVE ASSESSMENT OF THE PEST FREEDOM OF OAK AND WALNUT LOGS BASED ON THE EFFICACY OF THE TREATMENT
7
EKE results
7.1
An EKE was performed for pest freedom of oak logs (Q. alba and Q. rubra) and walnut logs (J. nigra) treated as proposed by the applicant for B. fagacearum, G. morbida and P. juglandis. In addition, also other pest species potentially associated with the commodity were assessed with species representing groups of organisms colonising different depths of wood. An overview of the elicitations conducted for the different combinations of pests and commodity species and a description of the methodology followed are provided in Section 2.2.3.
Bark and sapwood‐associated fungi and oomycetes (such as C. parasitica, D. virescens, E. mammata, N. euwallaceae and P. ramorum) are considered to be covered by the assessment of B. fagacearum and G. morbida and bark and sapwood‐dwelling insects (such as A. glabripennis, Pseudopityophthorus pruinosus, Pseudopityophthorus minutissimus, Euwallacea fornicatus sensu lato and Scolytinae spp. (non‐European)) are considered to be covered by P. juglandis and ambrosia beetles.
Insects which can inhabit the heartwood are represented by A. minutus.
Insects which are dwelling on the outer bark (such as L. delicatula, L. japonica) will not be able to survive the temperatures proposed in the treatment as they will be exposed to temperatures significantly higher than 56°C (see also Section 6.2). Therefore, this group of insects was not assessed further in the EKE.
The outcome of the EKE on pest freedom of oak logs (Q. alba and Q. rubra) and walnut logs (J. nigra) is presented in Table 12 and Figure 7.
Figure 8 provides an explanation of the descending distribution function describing the likelihood of pest freedom of Quercus alba logs produced in the US and treated with vacuum–steam–heat for B. fagacearum.
PANEL A
*Covers logs of all three tree species.Pest freedom categoryPest‐free plants out of 10,000Legend of marked pest freedom categoriesSometimes pest free< 5000 L Pest freedom category includes the elicited lower bound of the 90% uncertainty rangeMore often than not pest free5000–< 9000Frequently pest free9000–< 9500 M Pest freedom category includes the elicited medianVery frequently pest free9500–< 9900Extremely frequently pest free9900–< 9950 U Pest freedom category includes the elicited upper bound of the 90% uncertainty rangePest free with some exceptional cases9950–< 9990Pest free with few exceptional cases9990–< 9995Almost always pest free9995–10,000
PANEL B
Elicited certainty (y‐axis) of the number of pest‐free oak and walnut logs (x‐axis; log‐scaled) out of 10,000 designated for export to the EU from the US for all evaluated pests visualised as descending distribution function. Horizontal lines indicate the reported certainty levels (starting from the bottom 5%, 25%, 50%, 75%, 95%) Please see the reading instructions below.
Explanation of the descending distribution function describing the likelihood of pest freedom of Quercus alba logs produced in the US and treated with vacuum‐steam‐heat for Bretziella fagacearum.
Consideration of prevalence
7.2
Information on the prevalence of pests in the area of origin of the logs is important information for estimating the likelihood of pest freedom. However, this was not considered in the assessment because of the following reasons:
The applicant proposed the vacuum‐steam‐heat treatment as a stand‐alone method without applying any other risk mitigation measures.
The information provided by the applicant country on the prevalence was not sufficient.
The information on prevalence provided by the applicant in Dosser Section 2 is included below for potential use by risk managers.
The maximum prevalence of B. fagacearum is 100% on Q. rubra and 15% on Q. alba.
The prevalence of G. morbida is low to very low, with an exception for plantations in Washington State where 80% of trees were affected. In natural forest stands in the eastern and southern US, the prevalence is considered very low or insignificant. Populations of P. juglandis collapsed in Ohio, Pennsylvania, Maryland and Virginia between 2013 and 2019, and no new detections were reported in southern states between 2012 and 2024.
Pityophthorus juglandis is currently occurring in 8% of the J. nigra range.
Quercus is a major host of A. minutus, but prevalence is not available for this species. In general, it is considered widespread but always at moderate or low density.
Maximum prevalence of X. fastidiosa in urban trees of Q. rubra was 15%; no information was available for Q. alba. Juglans nigra is not reported as a host.
CONCLUSIONS
8
The efficacy of the vacuum steam treatment proposed by the US, to be used for Q. alba and Q. rubra against B. fagacearum and for J. nigra against G. morbida and P. juglandis, in mitigating the risk of introduction of Union quarantine pests was assessed. In addition to the above target pests, 14 other Union quarantine pests were identified as relevant for this opinion because they are present in the US and can potentially be associated with logs of Q. alba and/or Q. rubra and/or J. nigra. Some of these pests are regulated as groups of pests by Commission Implementing Regulation (EU) 2019/2072.
For the assessment of the treatment efficacy, the pests were grouped according to the depth of wood they can colonise. Insects which are dwelling on the outer bark (L. japonica and L. delicatula) will not be able to survive the temperatures proposed in the treatment as they will be significantly higher than 56°C. Therefore, this group of insects was not further assessed with an Expert Knowledge Elicitation (EKE).
The likelihood of pest freedom from B. fagacearum of infected and treated logs of Q. alba was estimated as ‘very frequently pest free’ with the 90% uncertainty range ranging from ‘very frequently pest free’ to ‘pest free with some exceptional cases’. For infected and treated Q. alba logs, the EKE indicated with 95% certainty that between 9021 and 10,000 logs per 10,000 will be free from B. fagacearum. The likelihood of pest freedom from B. fagacearum of infected and treated logs of Q. rubra was estimated as ‘very frequently pest free’ with the 90% uncertainty range ranging from ‘very frequently pest free’ to ‘pest free with some exceptional cases’. For infected and treated Q. rubra logs, the EKE indicated with 95% certainty that between 9347 and 10,000 logs per 10,000 will be free from B. fagacearum.
The likelihood of pest freedom from G. morbida of infected and treated logs of J. nigra was estimated as ‘very frequently pest free’ with the 90% uncertainty range ranging from ‘very frequently pest free’ to ‘pest free with some exceptional cases’. For infected and treated J. nigra logs, the EKE indicated with 95% certainty that between 9862 and 10,000 logs per 10,000 will be free from G. morbida. The likelihood of pest freedom from P. juglandis of infested and treated logs of J. nigra was estimated as ‘very frequently pest free’ with the 90% uncertainty range ranging from ‘very frequently pest free’ to ‘pest free with some exceptional cases’. For infested and treated J. nigra logs, the EKE indicated with 95% certainty that between 9948 and 10,000 will be free from P. juglandis.
The likelihood of pest freedom from ambrosia beetles of infested and treated logs of Q. alba, Q. rubra and J. nigra was estimated as ‘very frequently pest free’ with the 90% uncertainty range ranging from ‘very frequently pest free’ to ‘pest free with some exceptional cases’. For infested and treated Q. alba, Q. rubra and J. nigra logs, the EKE indicated with 95% certainty that between 9723 and 10,000 logs per 10,000 will be free from ambrosia beetles.
The likelihood of pest freedom from A. minutus of infested and treated logs of Q. alba and Q. rubra was estimated as ‘very frequently pest free’ with the 90% uncertainty range ranging from ‘very frequently pest free’ to ‘pest free with some exceptional cases’. For infested and treated Q. alba and Q. rubra logs, the EKE indicated with 95% certainty that between 1109 and 10,000 logs per 10,000 will be free from A. minutus.
The likelihood of pest freedom from X. fastidiosa of infected and treated logs of Q. alba and Q. rubra was estimated as ‘very frequently pest free’ with the 90% uncertainty range ranging from ‘very frequently pest free’ to ‘pest free with some exceptional cases’. For infected and treated Q. alba and Q. rubra logs, the EKE indicated with 95% certainty that between 9921 and 10,000 logs per 10,000 will be free from X. fastidiosa. It should be noted that wood is considered a very unlikely pathway of entry of X. fastidiosa because xylem fluid‐feeding insect vectors are not known to transfer the bacterium from detached wood to a host plant (EFSA PLH Panel, 2015).
Bark and sapwood‐associated fungi or oomycetes such as Cryphonectria parasitica, Davidsoniella virescens, Entoleuca mammata, Neocosmospora euwallacea and Phytophthora ramorum were considered to be covered by the assessment of B. fagacearum and G. morbida.
Bark and sapwood‐dwelling insects such as Anoplophora glabripennis, Euwallacea fornicates sensu lato, Pseudopityophthorus minutissimus, Pseudopityophthorus pruinosus and other Scolytinae species (non‐European) were considered to be covered by the assessment conducted for P. juglandis and ambrosia beetles.
GLOSSARY
Control (of a pest)Suppression, containment or eradication of a pest population (FAO, 2024a, 2024b).Entry (of a pest)Movement of a pest into an area where it is not yet present, or present but not widely distributed and being officially controlled (FAO, 2024b).Establishment (of a pest)Perpetuation, for the foreseeable future, of a pest within an area after entry (FAO, 2024b).Impact (of a pest)The impact of the pest on the crop output and quality and on the environment in the occupied spatial units.Introduction (of a pest)The entry of a pest resulting in its establishment (FAO, 2024b).MeasuresControl (of a pest) is defined in ISPM 5 (FAO, 2024b) as ‘Suppression, containment or eradication of a pest population’ (FAO, 2024a). Control measures are measures that have a direct effect on pest abundance. Supporting measures are organisational measures or procedures supporting the choice of appropriate risk mitigation measures that do not directly affect pest abundance.PathwayAny means that allows the entry or spread of a pest (FAO, 2024b).Phytosanitary measuresAny legislation, regulation or official procedure having the purpose to prevent the introduction or spread of quarantine pests, or to limit the economic impact of regulated non‐quarantine pests (FAO, 2024b).Quarantine pestA pest of potential economic importance to the area endangered thereby and not yet present there, or present but not widely distributed and being officially controlled (FAO, 2024b).Spread (of a pest)Expansion of the geographical distribution of a pest within an area (FAO, 2024b).
ABBREVIATIONSEKEExpert Knowledge ElicitationEPPOEuropean and Mediterranean Plant Protection OrganizationFAOFood and Agriculture OrganisationISPMInternational Standards for Phytosanitary MeasuresPLHPlant HealthTCDThousand Cankers DiseaseUSDA APHISAnimal and Plant Health Inspection Service of United States Department of AgricultureWTBWalnut Twig Beetle
REQUESTOR
European Commission
QUESTION NUMBER
EFSA‐Q‐2024‐00484
COPYRIGHT FOR NON‐EFSA CONTENT
EFSA may include images or other content for which it does not hold copyright. In such cases, EFSA indicates the copyright holder and users should seek permission to reproduce the content from the original source.
PANEL MEMBERS
Antonio Vicent Civera, Paula Baptista, Anna Berlin, Elisavet Chatzivassiliou, Jaime Cubero, Nik Cunniffe, Eduardo de la Peña, Nicolas Desneux, Francesco Di Serio, Anna Filipiak, Beata Hasiów‐Jaroszewska, Hervé Jactel, Blanca B. Landa, Lara Maistrello, David Makowski, Panagiotis Milonas, Paolo Gonthier Nikos T. Papadopoulos, Roel Potting, Hanna Susi, and Dirk Jan van Der Gaag.
Supporting information
APPENDIX D: Excel file with the EU quarantine pest list of oak and walnut species
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abdelrahman, I. (1974). The effect of extreme temperatures on California red scale, Aonidiella aurantii (mask.) (Hemiptera: Diaspididae), and its natural enemies. Australian Journal of Zoology, 22, 203–212. 10.1071/zo 9740203 · doi ↗
- 2Adlerz, W. C. (1980). Ecological observations on two leafhoppers that transmit the Pierce's disease bacterium. Proceedings of the Florida State Horticultural Society, 93, 115–120.
- 3Allen, E. (2014). ST 05: Review of heat treatment of wood and wood packaging draft. NAPPO, Science and Technology Documents, 35 pp. https://assets.ippc.int/static/media/uploads/resources/review_of_heat_treatment_of_wood_and_wood_packaging_english.pdf
- 4Almeida, R. P. P. , Pereira, E. F. , Purcell, A. H. , & Lopes, J. R. S. (2001). Multiplication and movement of a citrus strain of Xylella fastidiosa within sweet orange. Plant Disease, 85, 382–386. 10.1094/pdis.2001.85.4.382 30831970 · doi ↗ · pubmed ↗
- 5Anagnostakis, S. L. (1987). Chestnut blight: The classical problem of an introduced pathogen. Mycologia, 79(1), 23–37.
- 6Anagnostakis, S. L. , & Hillman, B. (1992). Evolution of the chestnut tree and its blight. Arnoldia, 52, 3–10.
- 7Anderson, N. A. , Ostry, M. E. , & Anderson, G. W. (1979). Insect wounds as infection sites for Hypoxylon mammatum on trembling aspen. Phytopathology, 69, 476–479. 10.1094/phyto-69-476 · doi ↗
- 8Anonymous . (1960). Index of plant diseases in the United States. Agriculture Handbook no 165, USDA‐ARS (US), 531 pp.