From Biological Waste to Therapeutic Resources: A Comprehensive Review of Stem Cell Sources, Characterization, and Biomedical Potentials
Beatrice Camia, Manuela Monti

TL;DR
This paper reviews how stem cells from biological waste can be used for regenerative medicine, offering ethical and cost-effective alternatives to traditional methods.
Contribution
The paper provides a comprehensive review of waste-derived stem cell sources and their biomedical potential, emphasizing recent advances and regulatory challenges.
Findings
Stem cells from waste materials show robust proliferation and differentiation potential for regenerative medicine.
Clinical trials report up to 30–40% improvement in recovery for osteoarthritis and ischemic heart disease using waste-derived stem cells.
Regulatory agencies classify some processing methods as 'more-than-minimal manipulation,' slowing clinical adoption.
Abstract
The concept of utilizing biological waste as a resource dates back centuries, with early practices in traditional medicine repurposing discarded tissues for healing purposes. In recent decades, advances in stem cell biology have revitalized this concept by identifying multipotent stem cells within various waste materials, including urine, adipose tissue, follicular fluid, umbilical cord blood, fetal annexes, menstrual blood, and dental pulp byproducts. These sources offer a minimally invasive, ethically sound, and cost-effective alternative to conventional stem cell harvesting methods. Stem cells derived from waste materials exhibit robust proliferative abilities and multilineage differentiation potential, positioning them as valuable tools for regenerative medicine, tissue engineering, and personalized therapeutic applications. Clinical studies highlight their promise. For example,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
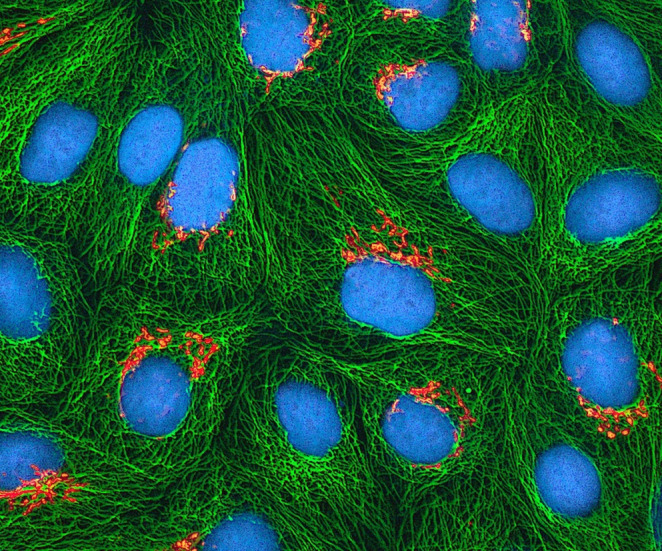 Figure 1
Figure 1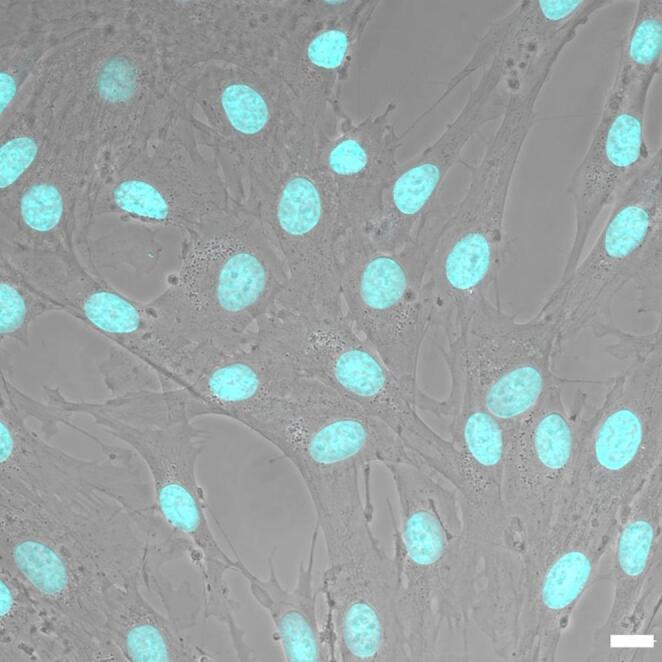 Figure 2
Figure 2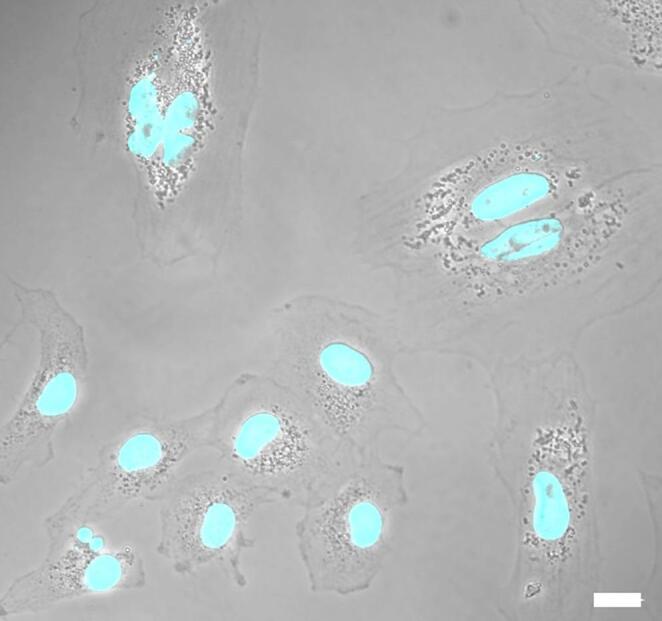 Figure 3
Figure 3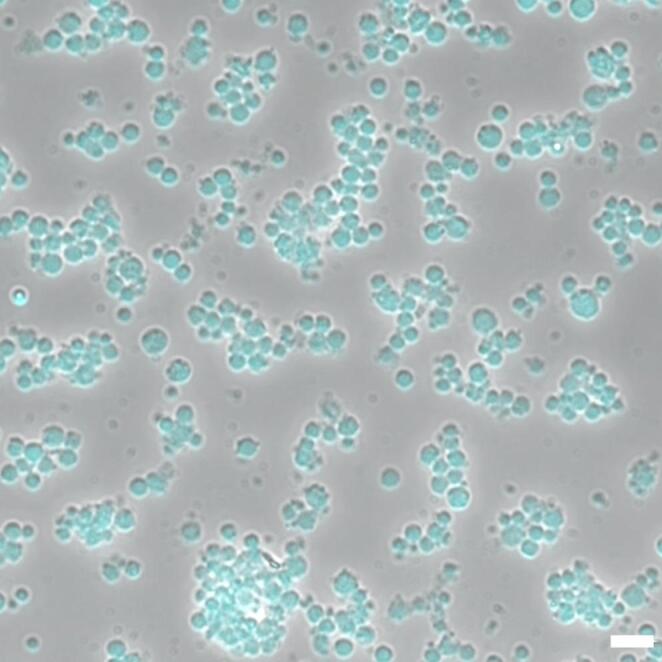 Figure 4
Figure 4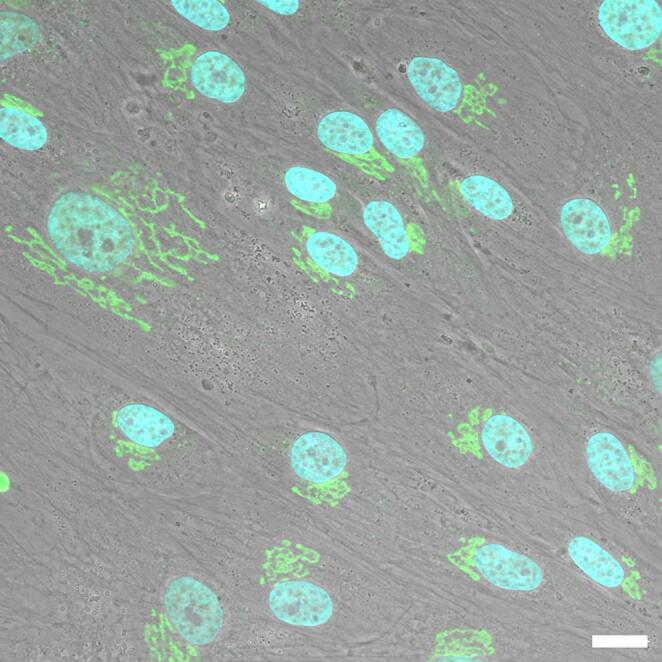 Figure 5
Figure 5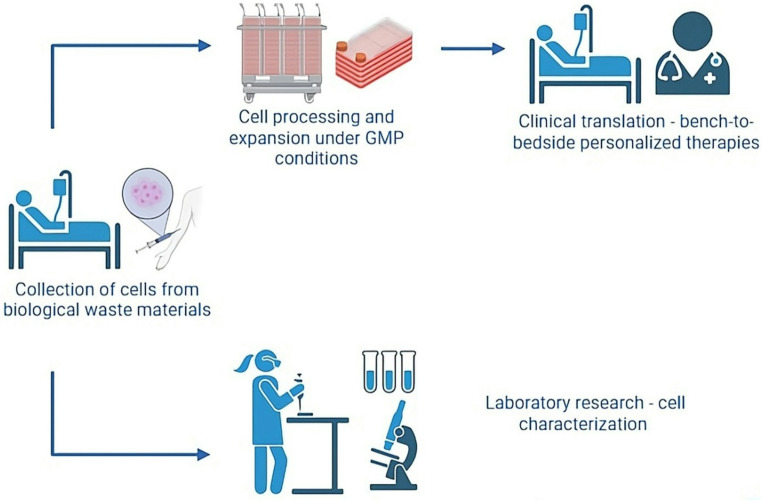 Figure 6
Figure 6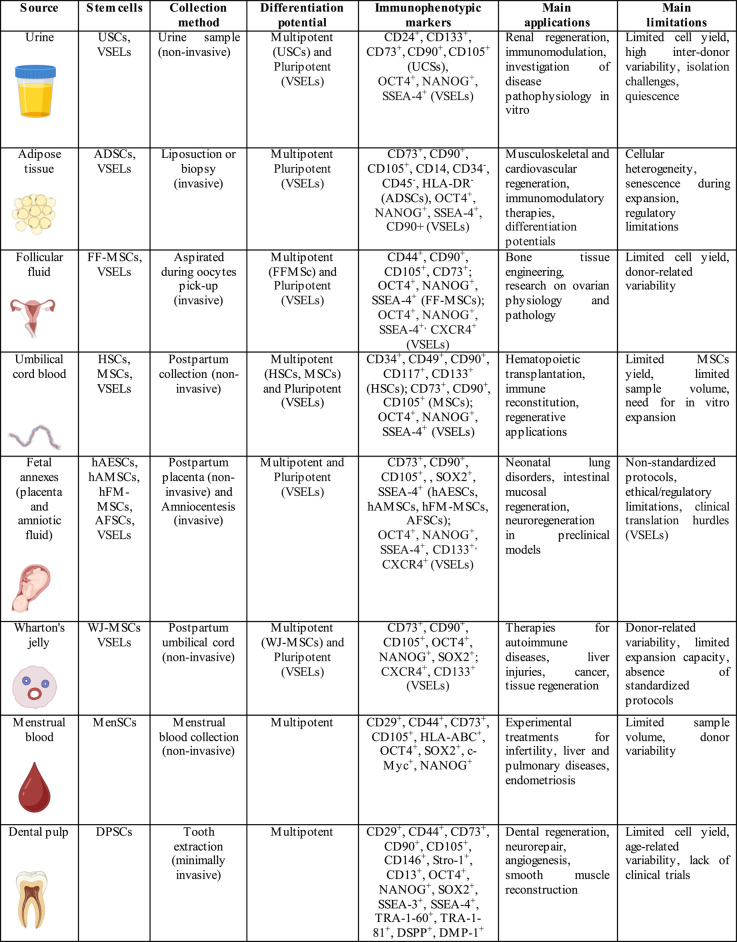 Figure 7
Figure 7- —Università degli Studi di Pavia
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPluripotent Stem Cells Research · Mesenchymal stem cell research · 3D Printing in Biomedical Research
Introduction
Historical view
"Nothing is created, nothing is destroyed, everything is transformed"
Antoine Lavoisier
The reuse of biological material—particularly substances once labeled as “waste”—represents a longstanding concept. Throughout history, biology has transformed materials once seen as useless or taboo into a valuable resource for medicine and research.
In ancient Greece and Rome, physicians and scholars such as Hippocrates, Galen, and Pliny the Elder, to cite a few [1, 2], viewed bodily fluids like blood, bile, and urine as diagnostic windows into health. They even incorporated substances such as earwax, menstrual blood, or discarded tissues into remedies [3]. Although these practices lacked a cellular understanding, they foreshadowed the therapeutic reuse of human material. By the middle Ages and Renaissance, anatomists made cadavers central to study. Pioneers such as Mondino de Liuzzi, Leonardo da Vinci, and Vesalius dissected bodies to advance knowledge of the human body [4, 5]. Later, in the 17th to 19th centuries, scientists studied discarded tissues from autopsies and surgeries under the microscope. This work laid the foundations of modern histology and pathology [6].
The discovery of the HeLa cell line in 1951, the first immortal human cell line, had a profound impact on the 20th century (Fig. 1). It also marked the beginning of a new era in biomedical research and highlighted the ethical issues surrounding informed consent [7]. Since then, researchers have identified materials once discarded—such as placenta, umbilical cord blood, amniotic fluid, and foreskin—as abundant sources of stem cells with therapeutic potential.Fig. 1. Multiphoton fluorescence image of cultured HeLa cells with a fluorescent protein targeted to the Golgi apparatus (orange), microtubules (green) and counterstained for DNA (cyan). Credit: Tom Deerinck, NIGMS, NIH. The Web site of the National Cancer Institute (https://www.cancer.gov)
Progress in stem cell biology, regenerative medicine, and biobanks establishment have confirms the biomedical value of these resources, and enables systematically collection and use for research, diagnostics, and therapies. Against this backdrop, this narrative review examines stem cell sources obtainable from biological waste, focusing on their characteristics, differentiation capacity, and clinical applications, and encourages further exploration of “hidden reservoirs” in discarded human materials. To structure this exploration, it is essential to outline the theoretical framework that underpins the review and the key questions that guide the analysis.
Theoretical Framework and Guiding Questions
This review adopts a regenerative medicine framework that conceptualizes biological waste as a reservoir of stem cells with biomedical utility. The theoretical foundation is in stem cell biology, with a focus on self-renewal, multipotency, and paracrine signaling. It pairs these with translational perspectives that emphasize minimally invasive, ethically acceptable, and cost-effective alternatives to traditional stem cell sources. The review addresses five main questions: (1) which categories of biological waste contain stem cell populations of biomedical relevance? (2) What phenotypic markers, differentiation potential, and functional properties characterize these cells? (3) What therapeutic and research applications can they support? (4) What limitations and challenges affect their collection, standardization, and clinical translation? (5) How can exploiting these sources advance personalized and sustainable cell-based therapies?
Rationale for Source Selection
This review highlights eight biological waste materials, selected according to three principal criteria. First, they are clinically and ethically acceptable tissues or fluids, collected non-invasively (e.g., urine, menstrual blood) or as byproducts of routine medical procedures (e.g., follicular fluid, umbilical cord, fetal annexes, dental pulp). Second, these sources contain stem cell populations with multipotent properties, as demonstrated by phenotypic characterization experimental studies. Third, they show established or emerging translational potential in regenerative medicine, disease modeling, and immunomodulation.
These criteria differentiate the selected materials from other waste sources that yield insufficient or poorly characterized cell populations, present greater ethical or technical challenges, or lack robust evidence for their suitability. Focusing on these eight categories allows emphasis on stem cell reservoirs that are scientifically validated and relevant for future therapeutic development.
Literature Search and Inclusion Criteria
Searches in PubMed, Scopus, and Web of Science identified the reviewed literature. The search strategy combined the terms “stem cells,” “biological waste,” “urine,” “adipose tissue,” “follicular fluid,” “umbilical cord blood,” “placenta,” “Wharton’s jelly,” “menstrual blood,” and “dental pulp”. The review focused on references published primarily from 2000 to 2025 and incorporated earlier landmark studies when relevant to historical or conceptual framing.
Inclusion criteria were: (i) studies characterizing stem or progenitor cells isolated from the eight selected biological waste sources; (ii) reports providing phenotypic, differentiation, or functional data; and (iii) preclinical or clinical studies demonstrating translational applications. Exclusion criteria were: (i) studies lacking primary experimental data (such as opinion pieces without evidence); (ii) sources outside the defined eight waste materials; and (iii) reports with insufficient methodological detail to support reproducibility.
This structured approach ensured that the review remained focused on scientifically validated, ethically acceptable, and translationally relevant waste-derived stem cell sources.
The following section examines the fundamental biological properties of stem cells, providing a foundation for evaluating those derived from biological waste materials.
Stem Cells
Stem cells are a population of highly undifferentiated cells characterized by their remarkable abilities for self-renewal, proliferation, and differentiation into mono- or multilineage cells. Due to these unique characteristics, stem cells have become increasingly central to regenerative medicine and cutting-edge biotechnologies in recent years. Their applications extend far beyond simple cell replacement, encompassing innovative therapeutic strategies that aim to repair and regenerate damaged tissues, modulate immune responses, and restore organ function. Additionally, stem cells serve as powerful models for investigating the pathogenic mechanisms underlying complex diseases, such as degenerative and autoimmune disorders, and for advancing novel pharmacological interventions, thereby establishing them as a cornerstone of translational medicine [8, 9].
Stem cells are categorized by their potency. Embryonic stem cells (ESCs) are pluripotent, able to form all cell types, while induced pluripotent stem cells (iPSCs) are reprogrammed somatic cells with similar capabilities [10, 11]. Adult (somatic) stem cells, found in postnatal tissues, are multipotent and support tissue maintenance by renewing cells within their germ layer, but cannot generate all cell types [12]. They avoid many ethical concerns and require only minimally invasive methods for collection although traditional sources, such as bone marrow, involve invasive procedures [13].
These limitations have prompted the search for alternative, more accessible sources of adult stem cell. Some sources use non-invasive collection methods. In contrast, some clinical interventions for diagnostic or therapeutic purposes yield stem cells through invasive procedure. Both approaches demonstrate the potential to repurpose biological waste as valuable scientific resources. Promising sources of stem cells from such waste include urine, adipose tissue, ovarian follicular fluid, umbilical cord blood, amniotic fluid, placenta, Wharton’s jelly, menstrual blood, and dental pulp (Table 1) [14–18]. These materials can be collected without harming donors and offer ethical, cost-effective options for regenerative therapies.Table 1. This table summarizes the principal characteristics of stem cells derived from waste biological materials, as presented in this review, in accordance with the rationale for source selection and the pre-specified inclusion criteriaAbbreviations: USCs: urine-derived stem cells; ADSCs: adipose-derived stem cells; FF-MSCs: follicular fluid-derived mesenchymal stem cells; HSCs: hematopoietic stem cells; MSCs: mesenchymal stem cells; VSELs: very small embryonic-like stem cells; hAESCs: human amniotic epithelial stem cells; hAMSCs: human amniotic mesenchymal stem cells, hFM-MSCs: mesenchymal stromal cells from the amniochorionic membrane; AFSCs: amniotic fluid stem cells; WJ-MSCs: Wharton’s jelly mesenchymal stem cells; MenSCs: menstrual blood-derived stem cells; DPSCs: dental pulp stem cells. Icons are created in BioRender.com
Biological Waste Materials
Urine
Urine is traditionally considered a biological waste fluid—the final product of renal filtration—with the primary physiological role of eliminating metabolic waste substances such as urea, creatinine, and uric acid from the body. Additionally, urine helps maintain fluid-electrolyte balance and overall homeostasis. Recent research, however, has prompted a significant reevaluation of urine’s functions. Urine no longer carries the sole label of metabolic waste; instead, its biologically active properties interact with urinary tract tissues, particularly the urothelium. Furthermore, urine carries functional molecules and viable cells with therapeutic potential [19]. This evolving understanding opens new avenues for exploring urine-derived cells and bioactive components in regenerative medicine and disease diagnostics.
Urine is composed primarily of water, accounting for approximately 95% of its volume, while the remaining 5% consists of dissolved solutes. Beyond these basic components, urine also contains proteins, extracellular microvesicles - including exosomes- and exfoliated epithelial cells from the urinary tract. Notably, among there exfoliated populations is a specialized subgroup of cells known as urine-derived stem cells (USCs). Recent studies have highlighted the therapeutic and diagnostic potential of USCs, thereby positioning urine not merely as waste but as a valuable biological resource for precision medicine and regenerative applications [20, 21].
Zhang and collaborators first identified USCs in 2008, demonstrating their potential as a source for urological tissue reconstruction [22]. Although their precise origin remains incompletely understood, USCs are believed to originate primarily from parietal epithelial cells of the glomerulus and exhibit varying degrees of stemness and multipotency once isolated [21].
USCs are multipotent adult stem cells that can be easily harvested from the urinary sediment of midstream samples via a completely non-invasive, cost-effective (less than $70 per sample) [23], painless, and repeatable procedure.
During initial isolation, even small urine volumes provide an average of 2–7.2 cells per 100 milliliters. These cells readily expand and form cellular clones.
These clones are capable of significant in vitro expansion, reaching up to 0.5 to 1 million cells within 10–15 days [22]. The culture media for USCs expansion typically included DMEM/F12 or Keratinocyte Serum-Free Medium, supplemented with Epidermal Growth Factor, Bovine Pituitary Extract, and Fetal Bovine Serum [24, 25].
These cells maintain genetic stability, exhibit a stem cell phenotype, and demonstrate active telomerase expression over time- features crucial for cellular longevity and therapeutic viability [21, 26].
Moreover, USCs are positive for a range of markers, including CD24, CD133, CD73, CD90, and CD105, and exhibit remarkable differentiation plasticity toward multiple cell lineages, such as osteogenic, myogenic, adipogenic, neuronal, and urothelial [27, 28].
This plasticity makes them valuable in tissue regeneration, not only through direct differentiation but also via indirect therapeutic mechanisms mediated by paracrine signaling. Indeed, USCs exhibit potent paracrine activity by secreting a variety of biologically active substances, including cytokines, growth factors, and exosomes with trophic and immunomodulatory properties [21, 27]. Among these, exosomes - small extracellular vesicles (EVs) ranging from 30 to150 nm- have garnered particular attention due to their heterogeneous and bioactive molecular cargo. These vesicles modulate immune responses, reduce oxidative stress, promote angiogenesis, regulate apoptosis, and enhance tissue repair in pathological conditions such as ischemia and inflammation [29].
Additionally, USC-derived exosomes display strong antioxidant activity and exert protective effects in pathological models such as diabetic nephropathy and chronic kidney disease [28].
USCs also play critical roles in modulating immune responses and apoptosis. Notably, they can inhibit CD4 + T-cell proliferation (useful in treating inflammatory and autoimmune diseases) and help regulate the balance between pro-apoptotic and anti-apoptotic proteins [28].
Compared to other stem cell sources such as ESCs or iPSCs, USCs demonstrate a superior safety profile, exhibiting a very low risk of tumorigenicity [21, 30]. Kim and collaborators [30] conducted a study to investigate the safety and bio-distribution of Klotho (kidney-specific therapeutic factor)-enhanced human USCs in BALB/c nude mice. Their goal was to advance these cells for clinical applications in kidney diseases. After both single and repeated intravenous administration of USCs, the researchers observed that Klotho-enhanced USCs targeted injured renal tissue and restored Klotho expression in models of ischemia–reperfusion injury. This demonstrated significant therapeutic potential. Importantly, there were no signs of toxicological or immunological adverse effects in the treated animals. Long-term monitoring over twenty-six weeks found no evidence of tumorigenicity attributable to the transplanted USCs. The few tumors observed were of murine rather than human origin, consistent with spontaneous tumor formation common in this mouse strain.
From a diagnostic perspective, USCs can be used to generate patient-specific cellular models for studying renal diseases and identifying urinary biomarkers associated with chronic or degenerative kidney disorders [21, 27]. Their non-invasive collection and strong biological relevance make them suitable for predictive diagnostics and in vitro drug toxicity screening [15, 31].
The ability to isolate USCs from the same individual at different time points enables longitudinal studies and personalized medicine applications, including patient-specific disease modeling and pharmacological screening [27]. Despite these advantages, the clinical use of USCs still faces some limitations. Cell yield can vary significantly among individuals influenced by factors such as age, health status, diet, and hydration. In some cases, urine samples may contain insufficient numbers of viable cells, necessitating the collection of large or repeated volumes. Additionally, standardization of isolation and culture protocols remains an ongoing concern, as USCs may exhibit phenotypic heterogeneity based on collection and culture conditions [28].
Lastly, although they present a low tumorigenic risk, long-term safety still requires comprehensive evaluation.
In conclusion, urine is not only an easily accessible and non-invasive biological fluid but also a safe and versatile source of adult stem cells. UCSs hold great promise across regenerative, predictive, and diagnostic medicine, though further research is needed to optimize their clinical application.
Adipose Tissue
Adipose tissue, often discarded as surgical waste, represents a promising and accessible source of adult stem cells. Liposuction and biopsy are two methods that supply this kind of tissue. The former, while more invasive, enables the collection of larger tissue volumes, making it suitable for applications that require high cell yields. Notably, different Body Mass Index (BMI) values significantly influence the isolation of adipose-derived stem cells (ADSCs). Specifically, ADSCs derived from individuals with higher BMIs exhibit slower proliferation rates, increased inflammatory profiles, altered gene expression related to aging, and higher levels of certain inflammatory markers, such as CCL2 (C-C-motif Chemokine 2) [32]. Although the adipogenic potential of these cells can vary, suggesting potential but compromised differentiation, the overall functional differences in ADSCs from various BMI groups highlight their altered biological properties and potential impact on metabolic health and regenerative medicine applications [32].
In contrast, biopsy is a less invasive method that allows for the targeted collection of small tissue samples. This approach offers particular advantages in clinical scenarios that require minimal tissue harvesting.
Adipose tissue comprises a highly heterogeneous population of cells, including preadipocytes, pericytes, hematopoietic cells, fibroblasts, smooth muscle cells, endothelial cells, and immune cells (such as B and T lymphocytes, macrophages, and myeloid cells), along with ADSCs [14]. ADSCs reside within the stromal vascular fraction of adipose tissue, a complex microenvironment primarily composed of connective tissue and blood vessels that supports the adipocytes.
Once collected, ADSCs can be isolated using various techniques, with enzymatic digestion typically using collagenase or Liberase, or by mechanical processing methods such as microfragmentation, centrifugation, or filtration, and while both approaches target the same stromal vascular compartment, the products they generate are not equivalent [33].
Enzymatic digestion reliably produces higher yields of viable nucleated cells, enriched in clonogenic mesenchymal progenitors, and offers a single-cell suspension suitable for expansion, precise dosing, and standardized phenotypic analysis. Optimization studies confirm that careful modulation of enzyme concentration and exposure enhances recovery while maintaining stemness properties [34, 35]. Mechanical isolation, in contrast, generates a lower number of liberated cells but preserves extracellular matrix scaffolds and perivascular niches, producing microfragmented adipose tissue (MFAT) that maintains a native structural context argued to support paracrine signaling and engraftment [36]. This distinction is more than technical, as enzymatically derived ADSCs suspensions and mechanically produced MFAT differ in formulation and thus in potential mechanisms of action. Regulatory frameworks also amplify these contrasts: in Europe and the United States, authorities generally classify enzymatic digestion of adipose tissue as more-than-minimal manipulation, which requires advanced manufacturing controls, whereas they often allow mechanical methods under minimal manipulation standards, making them more accessible for point-of-care autologous therapies [37]. As defined by the US Food and Drug Administration, more-than-minimal manipulation standards refer to human cells, tissues, and cellular or tissue-based products that have undergone processing that alters their original relevant characteristics, functions, or utility, while minimal manipulation standards mean processing that does not alter the relevant biological characteristics of cells.
The similarities indicate that each technique taps into the same cellular reservoir, yet their differences—in yield, product form, regulatory classification, and clinical effect profile— underscore that researchers should consider enzymatic and mechanical isolation of human adipose stem cells complementary rather than interchangeable strategies [34].
ADSCs are mesenchymal stem cells (MSCs) with multipotent differentiation capacity and several biologically relevant characteristics. They can differentiate predominantly toward the mesodermal lineage into osteoblasts, chondrocytes, adipocytes, and myocytes, and they exhibit robust in vitro proliferation while maintaining genomic and phenotypic stability [38]. In culture, ADSCs adhere readily to plastic surfaces, assume a fibroblast-like morphology, and express an MSCs immunophenotype positive for CD73, CD90, and CD105, and negative for CD14, CD34, CD45, and HLA-DR (Fig. 2) [14, 33].Fig. 2. Phase contrast image of human ADSCs. Nuclei are stained in blue (DAPI). Magnification: 60X, bar: 10 μm. (Unpublished data)
Beyond their differentiation potential, ADSCs exert therapeutic effects via paracrine mechanisms. They secrete a variety of cytokines and growth factors, including VEGF, HGF, TGF-β, IGF-1, as well as exosomes enriched with microRNAs and bioactive proteins. The secretions contribute to modulating immune responses, enhancing angiogenesis, regulating apoptosis, and promoting tissue regeneration [39] Among all biologically derived waste materials, adipose-derived stem cells (ADSCs) are among the most extensively studied in both preclinical and clinical settings. In regenerative medicine, they have shown efficacy in bone and cartilage repair, including treatment of bone defects, osteoarthritis, and joint injuries [38, 39]. Their angiogenic, anti-apoptotic, and anti-fibrotic effects are notably in cardiovascular and ischemic diseases such as myocardial infarction and peripheral ischemia. Their immunomodulatory capabilities support applications of ADSCs in models of autoimmune and inflammatory diseases, including multiple sclerosis, lupus, Crohn’s disease, and rheumatoid arthritis [14] Furthermore, ADSCs-derived exosomes have emerged as a promising cell-free therapeutic option. These exosomes, rich in miRNAs and bioactive factors, have demonstrated regenerative potential in wound healing, musculoskeletal repair, as well as in dermatological applications [40] Overall, ADSCs are a highly versatile therapeutic tool with broad, still partially untapped, potential. However, ADSCs also present certain limitations. Donor variability and cellular heterogeneity pose challenges to treatment standardization [41]. Additionally, pathological conditions such as obesity and aging can impair cell vitality and function. Extended in vitro expansion leads to senescence, diminishing their regenerative potential over time [42]. Moreover, concerns regarding safety and manipulation persist, and regulatory constraints continue to slow clinical translation [43].
Follicular Fluid
Follicular fluid (FF) is the biological fluid found within mature ovarian follicles, the functional units of the ovary that support the development and maturation of the oocyte Granulosa cells and theca interna cells secrete the FF that contains a complex and dynamic mixture of bioactive components, including hormones, proteins and peptides, metabolites, lipids, growth factors and cytokines, antioxidants and reactive oxygen species (ROS), extracellular vesicles, and diverse cell populations [44–46]. Due to its intricate composition, FF plays a crucial role in establishing the biochemical environment required for oocyte maturation and in mediating communication between the oocyte and surrounding somatic cells During assisted reproduction technologies, particularly in vitro fertilization, FF is collected as part of the oocyte retrieval process following controlled ovarian stimulation. However, it is typically discarded- despite being a valuable source of cells and bioactive molecules Post-retrieval, the cellular component of the FF is notably heterogeneous. It includes granulosa cells, immune cells, and a population of MSC-like stem cells, as well as a minor fraction of epithelial cells likely introduced from the vaginal, cervical, or tubal epithelium during aspiration [47] Upon isolation, MSCs derived from follicular fluid (FF-MSCs) adhere to culture substrates such as plastic and glass, exhibiting progressive morphological changes. Initially rounded in shape, these cells gradually become cuboidal during early adhesion phases and eventually adopt an elongated, fibroblast-like morphology typical of MSCs (Fig. 3) [47, 48].
From an immunophenotypic perspective, FF-MSCs express surface characteristics of MSCs, including CD44, CD90, CD105, and CD73. In addition, a minority of the cell population may express pluripotency-associated markers such as OCT4, NANOG, and SSEA-4, although the functional implications of this expression remain under investigation [47] It is crucial to recognize that the composition and properties of follicular fluid are highly influenced by the physiological and pathological status of the ovary. Conditions such as polycystic ovary syndrome (PCOS), advanced maternal age, oxidative stress, and chronic inflammation can significantly alter the biochemical profile of FF, including concentrations of hormones, growth factors, cytokines, ROS, extracellular matrix proteins, and exosomes, which in turn affect the vitality and phenotypic traits of isolated FF-MSCs [49].
Metabolomics analyses have revealed that FF from patients with endometriosis, PCOS, or diminished ovarian reserve displays altered levels of key energetic metabolites and nutrients, potentially impairing oocyte quality and the functional properties of associated stem cells [49].
In particular, proteomic profiling of small antral follicles in polycystic ovaries has shown increased expression of inflammatory and immune-related pathways, along with reduced levels of proteins critical for oocyte maturation and endocrine signaling [50]. These alterations may negatively influence the differentiation potential and regenerative capabilities of FF-MSCs.
Despite these influences, FF-MSCs have demonstrated multipotent differentiation capacity in vitro. Under appropriate induction conditions, they can differentiate into osteogenic, adipogenic, and chondrogenic lineages. A small subpopulation has also been reported to acquire epithelial- or neural-like morphologies during culture, suggesting morphological plasticity. However, conclusive evidence for transdifferentiation into ectodermal or endodermal lineages remains limited [47, 48, 51].
Given their properties, FF-MSCs show considerable promise in regenerative medicine. Their paracrine activity relies on the secretion of pro-regenerative, angiogenic, and immunomodulatory factors. Researchers have used these properties in models of bone tissue regeneration [52, 53].
FF-MSCs also serve as a valuable in vitro model for reproductive and toxicological research because their phenotypic and molecular characteristics partly reflect the follicular microenvironment from which they originate. FF-MSCs are useful for studying ovarian pathophysiology, assessing environmental or pharmacological exposures, and evaluating therapeutic interventions in a physiologically relevant human model, offering an ethically preferable alternative to animal testing [47, 54].
An emerging and particularly promising area of study is the analysis of FF-MSCs- derived exosomes. These EVs carry a molecular cargo rich in proteins, lipids, mRNAs, and microRNAs, with potential roles in modulating inflammation, promoting angiogenesis, and supporting tissue regeneration [55]. Nevertheless, the clinical translation of FF-MSCs faces several obstacles. The yield of viable stem cells per FF sample is typically low, and donor-dependent variability further complicates the standardization of these cells. Moreover, there remains a lack of consensus on optimal protocols for their isolation, expansion, and characterization. Until these challenges are resolved, the use of FF-MSCs will remain largely confined to the preclinical and experimental stages.
Umbilical Cord Blood
Umbilical cord blood (UCB) is the residual blood remaining in the placenta and umbilical cord following childbirth. Traditionally considered a biological waste product, UCB is now recognized as a rich and ethically accessible source of both pluripotent and multipotent stem cells [17].
Its collection is safe, non-invasive, and painless for both mother and newborn. Immediately after delivery and cord clamping, the procedure involves inserting a sterile needle into the umbilical vein. This approach yields between 60 and 100 ml of blood, with the amount depending on various maternal and perinatal factors UCB is biologically rich, containing a diverse population of hematologic and immune cells. These include T and B-lymphocytes, monocytes, macrophages, natural killer (NK) cells, granulocytes, nucleated red blood cells, and platelets. Among its most valuable components are hematopoietic stem cells (HSCs) and hematopoietic progenitor cells (HPSc), which have potent proliferative and regenerative capabilities [56, 57].
In smaller quantities, UCB also harbors MSCs, endothelial and epithelial progenitors, and regulatory immune cell subsets [58–60].
HSCs and HPCs from UCB express specific surface markers such as CD34, CD49, CD90, CD117, and CD133. Of particular interest is the CD34^+^CD38^−^ subpopulation, which represents a highly primitive stem cell phenotype. This population is significantly more abundant in UCB than in adult bone marrow, suggesting that UCB-HSCs possess enhanced capacity and distinct biological features [61].
Functionally, UCB-derived HSCs demonstrate superior hematopoietic reconstitution in vivo and greater proliferative potential in vitro. These cells have longer telomeres, contributing to enhanced replicative lifespan, and a faster transition out of the quiescent G0/G1 phase, enabling more rapid proliferation than adult-derived HSCs [58].
Such features have positioned UCB as a preferred source of stem cells for clinical transplantation, especially in the context of allogeneic hematopoietic stem cell transplantation. To date, regulatory agencies have approved UCB-HSCs to treat a broad range of hematological, genetic, and immunological diseases, including acute and chronic leukemia, lymphomas, thalassemia, sickle cell disease, and congenital immunodeficiency [62–64].
Their regenerative potential and lower risk of graft-versus-host disease (GvHD), even in partially HLA-mismatched transplants, account for the clinical success of UCB-HSCs. The immunological immaturity of neonatal immune cells contributes to this lower risk [65].
In addition to hematopoietic applications, UCB-derived CD34⁺ cells and MSCs have shown promise in regenerative medicine for their angiogenic and neurotrophic properties. Ex vivo expansion of CD34⁺ cells can enhance the secretion of vascular and neural growth factors, suggesting additional therapeutic avenues in neurodegenerative and ischemic diseases [59, 64].
Moreover, UCB cells are a versatile source of cells for advanced therapeutic applications beyond traditional hematopoietic transplantation. Researchers are currently exploring UCB-derived immune cells in a range of experimental cellular therapies, including the generation of Chimeric Antigen (CAR) T cells, regulatory T cells, virus-specific T cells, and NK cells. These approaches primarily enhance immune responses in immunocompromised patients and treat viral infections [66].
UCB-MSCs exhibit a fibroblast-like morphology and are multipotent with the ability to differentiate into osteoblasts, chondrocytes, and adipocytes. Immunophenotypically, they express CD73, CD90, and CD105 while lacking hematopoietic markers [67].
These cells exhibit enhanced biological properties compared to adult-derived MSCs, including a higher proliferative capacity, greater self-renewal potential, and maintenance of chromosomal stability after cryopreservation. They are also characterized by low immunogenicity, partly due to downregulated HLA expression, and secrete key immunomodulatory molecules such as IL-10, TGF-β, and PGE₂. Furthermore, UCB-MSCs possess robust homing capabilities, mediated by chemokine receptors and specific signals, allowing them to migrate toward sites of tissue injury and inflammation [68].
UCB-MSCs have been successfully employed in numerous clinical and preclinical settings. Notably, several studies have demonstrated their efficacy in articular cartilage regeneration and skin regeneration. In dermatology and wound healing, UCB-MSCs and their secretome—including exosomes—have demonstrated regenerative capabilities. These effects are primarily mediated through modulation of the TGF-β signaling pathway, resulting in accelerated wound closure, reduced inflammation, and improved skin architecture [69, 70].
Intra-articular injection of UCB-MSCs in patients with knee osteoarthritis has led to significant improvements in cartilage integrity, pain reduction, and overall joint function, with a strong safety profile [71, 72].
Fig. 3. Phase contrast image of human FF-MSC. Nuclei are stained in blue (DAPI). Magnification: 60X, bar: 15 μm. (Unpublished data)
In pulmonary medicine, a recent 2025 clinical trial reported that patients with pulmonary fibrosis tolerated nebulized EVs derived from UCB-MSCs well showed improvements in respiratory function and quality of life [73]. In patients with COVID-19-induced acute respiratory distress syndrome, UCB-MSCs have been used to mitigate the hyper inflammatory “cytokine storm”, resulting in reduced systemic inflammation and favorable safety outcomes in early-phase clinical trials (Supplementary Table 1) [74].
Ongoing studies are examining UCB-MSCs for the treatment of autoimmune diseases, endocrine and metabolic disorders, and neurological conditions, as well asfor applications in regenerative dermatology [75].
Finally, a rare and particularly intriguing population within UCB is that of Very Small Embryonic-Like cells (VSELs). These cells are significantly smaller than red blood cells (3–6 μm) and exhibit a high nucleus-to-cytoplasm ratio with undifferentiated chromatin (Fig. 4).Fig. 4. Phase contrast image of human UCB-derived VSEL. Nuclei are stained in blue (DAPI). Magnification: 180X, bar: 10 μm [78]
VSELs express pluripotency-associated markers such as OCT4, NANOG, and SSEA-4, and differentiate into the three germ layers under appropriate culture conditions. Evidences suggest that VSELs participate in neonatal tissue repair and fetomaternal chimerism [76–79]. However, they represent a promising but still controversial stem cell population [76–82]. Since their discovery, numerous independent studies challenge both the existence and functional significance of VSELs. Critics argue that isolation protocols may mistakenly sort cell debris, apoptotic bodies, or erythroblasts rather than living stem cells [80–82]. On the other hand, this may be due to imprecise selection methods, as VSELs are extremely small and rare (0.01%–0.03% of nucleated cells in bone marrow/UCB) [76, 83, 84].
Recent findings involving ex vivo cell expansion lay the foundations for potential applications in regenerative medicine [85]. Advancing isolation techniques and clarifying mechanisms of action, position UCB-derived VSELs as a promising pluripotent cell source for clinical therapies, particularly non-embryonic and ethically acceptable frameworks.
Despite the expanding clinical utility of UCB-derived cells, several limitations persist. A primary constraint is the limited volume of cord blood collected at birth, which often yields an insufficient number of cells—particularly hematopoietic or mesenchymal—for adult patients without ex vivo expansion. Moreover, MSCs occur at low frequencies in UCB, necessitating prolonged and labor-intensive culture protocols to achieve therapeutic doses.
Nonetheless, ongoing advances in cellular characterization, expansion techniques, and standardized isolation methods are progressively overcoming these barriers. These developments continue to enhance the scalability, consistency, and clinical impact of UCB-derived cell therapies, positioning them at the forefront of regenerative and personalized medicine.
Fetal Annexes: Amniotic Fluid and Placenta
The placenta and amniotic fluid are vital fetal structures essential for intrauterine development and represent promising, ethically accepted sources of perinatal stem cells. The placenta is a transient organ formed from both fetal and maternal tissues. It serves as the primary interface between the mother and fetus, mediating nutritional, gaseous, hormonal, and immune exchanges. It can be collected safely and non-invasively after delivery, via either vaginal birth or cesarean section, making it a readily available and ethically favorable biological resource. In contrast, amniotic fluid surrounds the fetus during pregnancy, playing protective, antibacterial, and nutritional roles [86]. Its collection requires an invasive procedure (amniocentesis), typically performed between the 15th and 20th weeks of gestation for diagnostic purposes. Despite their differences, both sources harbor perinatal stem cells with highly favorable biological characteristics: low immunogenicity, absence of tumorigenicity, notable plasticity, and production of bioactive factors with regenerative effects.
Specifically, the placenta consists of three main populations [87–89]:
- Human Amniotic Epithelial Stem Cells (hA-ESCs), derived from the inner membrane of the placenta (the amnion), are particularly compelling due to their high plasticity, expression of pluripotency-associated markers, and low immunogenicity and tumorigenicity. These cells also release pro-regenerative factors with potential in wound healing, neuroregeneration, and epithelial repair [87, 90].
- Human Amniotic Mesenchymal Stem Cells (hA-MSCs), derived from the amniotic mesenchyme, are multipotent cells that share similarities with adult MSCs, including the ability to differentiate into osteogenic, chondrogenic, and adipogenic lineages. They secrete cytokines, exosomes, and exhibit strong immunomodulatory capacity [91].
- Mesenchymal Stromal Cells from the Amniochorionic Membrane (hFM-MSCs), originated from the fetal side of the placental wall, express key pluripotency genes, such as OCT4, NANOG, and SOX2. They are capable of mesodermal differentiation and contribute to bone regeneration and immune modulation [92].
Instead, the amniotic fluid contains Amniotic Fluid Stem Cells (AFSCs), which possess characteristics intermediate between embryonic and adult stem cells. They exhibit variable morphology (round or elongated), express embryonic markers such as SSEA-4, OCT4, and CD90, and show low tumorigenicity. Functionally, they can differentiate into cardiac, hepatic, neuronal, and pancreatic cells, and release bioactive exosomes and microRNAs with therapeutic relevance [91, 93].
Thanks to the properties described above, stem cells derived from the placenta and amniotic fluid have demonstrated considerable promise in preclinical and early clinical settings across multiple organ systems. For example, placental MSCs have shown efficacy in treating neonatal pulmonary disorders, such as bronchopulmonary dysplasia, by reducing inflammation and improving oxygenation [94]; in promoting intestinal mucosa regeneration in preclinical models of inflammatory bowel disease [95], and in stimulating neuronal regeneration following injury in rodents [96]. A 2023 review further highlights how placental cells, exosomes, and decellularized matrices can be integrated with advanced biomaterials and tissue engineering techniques (such as 3D bio printing and composite scaffolds) to regenerate skin, bone, cartilage, and the cardiovascular system, positioning the placenta as a “bio-factory” for multi-organ regenerative therapies [97]. hA-MSCs demonstrate significant regenerative potential across several medical fields. In the cardiovascular system, these cells reduce fibrosis and limit necrotic tissue following myocardial infarction [98]. In the pulmonary domain, amniotic fluid-derived mesenchymal stem cells effectively reduce inflammation and enhance alveolar permeability, improving lung function in preclinical models [99]. Moreover, in the musculoskeletal system, extracellular vesicles released by these stem cells counteract steroid-induced osteoporosis in vitro [100]. Despite this remarkable potential, the clinical use of these sources remains limited by regulatory challenges, the variability of isolation protocols, and the need for further safety and efficacy validation in human studies.
Wharton’s Jelly
Wharton’s jelly is a gelatinous connective tissue derived from the extraembryonic mesoderm, surrounding the umbilical cord vessels (two arteries and one vein). Rich in collagen, hyaluronic acid, proteoglycans, and glycoproteins, its primary role is to protect umbilical vessels from mechanical compression [101]. It has gained attention as a biological waste source of MSCs (WJ-MSCs, Wharton’s jelly MSCs), which resemble bone marrow and adipose-derived MSCs but may possess superior plasticity and proliferation capacity [102].
WJ-MSCs can be isolated via enzymatic digestion or explant culture [101]. They express typical MSCs markers (CD105, CD73, and CD90) and pluripotency markers like OCT4, NANOG, and SOX2. These multipotent cells differentiate into osteogenic, chondrogenic, adipogenic lineages, as well as mesodermal-derived cells such as neurons and pancreatic cells under appropriate induction [103].
Growing interest has also focused on the secretome of WJ-MSCs, which includes soluble factors (cytokines, chemokines, growth factors, and enzymes) as well as EVs (exosomes and microvesicles), which hold therapeutic promise in “cell-free” approaches [104] WJ-MSCs immunomodulatory properties support their use in autoimmune disease (e.g., lupus erythematosus and multiple sclerosis) and graft-versus-host disease (GVHD) by inhibiting the activation of T and B-lymphocytes, NK cells, and macrophages [104]. They promote liver regeneration by reducing fibrosis in chronic hepatitis and cirrhosis models [105], enhance cardiac repair post-myocardial infarction [106], and provide neuroprotection by secreting anti-inflammatory molecules [107]. Other promising applications include renal and lung repair, chronic wound healing and support for pancreatic function in diabetes [106].
In experimental oncology, EVs from WJ-MSCs carrying microRNAs, such as miR-125b, can inhibit tumor angiogenesis and proliferation in triple-negative breast cancer models [108].
Despite these promising perspectives, the clinical use of WJ-MSCs and their secretome is hindered by donor variability, in vitro senescence, challenges in standardizing dosing and delivery, and heterogeneity in secretome composition and isolation protocols [104].
Menstrual Blood
Menstrual blood is a complex biological fluid composed of blood, cervical mucus, shed endometrial epithelial cells, immune cells, red blood cells, platelets, occasional genital tract cells, and vaginal secretions, cyclically expelled during menstruation. It originates from the shedding of the functional layer triggered by the drop in progesterone and estrogen in the absence of fertilization. Historically considered waste, menstrual blood is now recognized as a rich, non-invasive source of adult stem cells with regenerative potential [16, 109, 110].
The most notable are menstrual blood-derived stem cells (MenSCs), multipotent adult stem cells similar to bone marrow MSCs. In culture, they initially display radial or helical growth, followed by a spindle-shaped morphology, and expand rapidly with genomic stability and no tumorigenic or immunogenic risks [111]. MenSCs express MSC markers CD29, CD44, CD73, CD105, HLA-ABC, and lack hematopoietic markers [112]. Interestingly, they also express pluripotency markers (OCT4, SOX2, c-Myc, and NANOG), although the expression of c-Kit and SSEA-4 remains uncertain [112–114]. These cells can differentiate into osteogenic, chondrogenic, and adipogenic lineages, albeit with lower osteo/adipogenic potential than bone marrow MSCs [114]. They also differentiate into various other cell types, including germ cells, endometrial, endothelial, cardiomyocyte, respiratory epithelial, neural, muscle, hepatic, and pancreatic cells [115].
MenSCs can differentiate into female reproductive cells. In vitro, they acquire ovarian-like features expressing FSHR, LHR, and oocyte-related genes, including STRA8, GDF9, SCP3, and DDX4, which suggests their applicability in reproductive regenerative medicine [116]. Animal and early clinical studies demonstrate that MenSCs can regenerate ovarian function in premature ovarian insufficiency, thereby improving endocrine and menstrual parameters [115, 117].
MenSCs secrete cytokines with antifibrotic, angiogenic, anti-inflammatory, and immunoregulatory properties. In a recent murine model of liver fibrosis, MenSCs reduce fibrosis via ECM1 protein secretion acting on FoxO1 and mTOR pathways [118]; they have also shown positive therapeutic potential for the treatment of various cardiovascular diseases [119], while in respiratory diseases like acute respiratory distress syndrome, cryopreserved MenSCs restore alveolar structure, reduce inflammation, and improve gas exchange [120]. For musculoskeletal repair, MenSCs embedded in a collagen gel preserve the intervertebral disc structure after injury, as shown in a rat model [121].
Alongside cell-based applications, growing interest has emerged in the cell-free use of MenSCs, their secretome, and EVs. MenSC-derived exosomes show promising effects in reducing granulosa cell apoptosis in chemotherapy-induced ovarian damage models [122]. They have also proven effective in models of fulminant hepatic failure by improving liver function, enhancing survival, and reducing hepatocyte apoptosis [123]. Furthermore, in murine models of endometriosis, these exosomes inhibited lesion growth, angiogenesis, cellular proliferation, and invasiveness [124].
Despite their promise, MenSCs face challenges, including limited sample volume, donor variability, and a lack of standardized protocols for isolation and clinical use. Most evidence remains preclinical, with long-term efficacy and safety yet to be firmly established.
Dental Pulp
Dental pulp is a loose connective tissue that is highly vascularized and innervated. It resides in the inner central cavity of the tooth, closely associated with dentin, and together they form the dentin–pulp complex. This tissue consists mainly of two zones: the central zone, rich in blood vessels and nerves, and the peripheral zone, which contains odontoblasts that produce dentin. Unlike other connective tissues, dental pulp occupies a rigid cavity and receives limited vascular access only through the root apex [125].
The dental pulp performs multiple essential functions, including formation of dentin, nourishment of the tooth, protection from mechanical insults through the production of secondary dentin, and the transmission of sensitivity via nerve fibers responsive to thermal and mechanical stimuli [126].
Dental pulp collection involves the use of naturally exfoliated deciduous teeth or extracted permanent teeth, thereby maintaining a minimally invasive process.
This tissue has a heterogeneous composition: as mentioned, its outermost layer includes odontoblasts. Beneath them lies the acellular zone of Weil, rich in unmyelinated nerve fibers, capillaries, and fibroblastic processes. The main component is the extracellular matrix, also referred to as the “cell-rich zone” or “pulp core” [125]. Several cell types reside within dental pulp, including: fibroblasts, the most abundant cells; immune cells, present even under physiological conditions, such as granulocytes, T lymphocytes, monocytes/macrophages, dendritic cells, and to a lesser extent, NK cells, B cells, and regulatory T cells [126]. The central region, the most densely populated area, contains dental pulp stem cells (DPSCs) [18, 127]. Extraction of pulp tissue from the tooth’s internal chamber, following disinfection and longitudinal tooth opening, yields DPSCs. Immediately processing or cryopreservation preserves the tissue for later use. Enzymatic digestion with collagenase type I and dispase produces a cell suspension for standard culture conditions. Plastic adherence and fibroblast-like morphology identify stem cells, which then expand in culture for research or therapy [128].
DPSCs are mesenchymal progenitor cells mainly derived from the cranial neural crest. In addition to expressing typical mesenchymal markers, (CD29, CD44, CD73, CD90, CD105, CD146, Stro-1, CD13), DPSCs also exhibit positivity for pluripotency markers (OCT4, NANOG, SOX2, SSEA-3, SSEA-4, TRA-1–60, TRA-1–81), odontoblastic markers (Dentin sialophosphoprotein – DSPP; Dentin Matrix Protein-1 – DMP-1), and neural stem cell markers such as GFAP, Nestin, P75, HNK-1, and S-100. They are negative for hematopoietic markers including CD34, CD45, CD14, CD11b, and HLA-DR [18, 129–131].
In vitro, DPSCs show high proliferative capacity, colony formation ability, superior replicative potential, and genomic stability compared to bone marrow-derived MSCs [129, 131]. They can differentiate into odontoblast-like cells forming mineralized nodules and dentin-like structures, as well as into osteogenic, chondrogenic, adipogenic, neurogenic, myogenic, endothelial, and pancreatic lineages [128, 131, 132] (Fig. 5).
In dentistry, combinations of DPSCs and scaffolds regenerate functional, vascularized pulp-like tissues, and marksignificant progress in regenerative endodontic therapy [133–135]. Additionally, DPSCs exhibit strong osteogenic potential and have been applied in mandibular and craniofacial bone regeneration, demonstrating enhanced vascularization and bone formation compared to acellular scaffolds [136].
DPSCs have contributed to functional recovery in preclinical spinal cord injury models by expressing neuronal markers and secreting neurotrophic factors (NGF, BDNF), supporting axonal regeneration and reducing inflammation [137, 138]. They also promote angiogenesis and aid tissue regeneration in ischemic conditions.
Furthermore, DPSCs accelerate wound healing by secreting anti-inflammatory and pro-regenerative factors, thereby providing therapeutic benefits for chronic wounds and ulcers [139, 140]. They have also demonstrated potential in smooth muscle regeneration, such as bladder reconstruction, and exhibit immunomodulatory properties relevant to the treatment of autoimmune and metabolic diseases [141–143].
Fig. 5. Phase contrast image of human DPSCs. Nuclei are stained in blue (DAPI), Golgi apparatus in green. Magnification: 60X, bar: 10 μm [128]
Despite these promising preclinical and clinical applications, DPSCs face challenges for large-scale use: limited tissue volume and low cell yield per tooth, biological properties that vary with donor age, with deciduous tooth-derived DPSCs exhibiting higher proliferative capacity than those from adult teeth, and a lack of large-scale clinical trials to validate their safety and efficacy in humans (Supplementary Table 1).
Regulatory Considerations for the Use of Stem Cells Derived from Biological Waste Materials
Stem cells from biological waste materials offer a promising alternative to embryonic sources. These materials raise fewer ethical concerns. However, regulatory bodies still enforce complex oversight to address the remaining ethical issues and ensure proper governance. Such oversight protects donor rights, ensures product quality, and supports the safe and effective use of these cell products in clinical settings.
Regulatory Pathways
The extent of manipulation directly shapes manufacturing requirements. Researchers who process cells extensively must use Good Manufacturing Practice (GMP) conditions. This maintains sterility, identity, potency, and reproducibility. Laboratory interventions introduce variability and potential risks, so stringent quality controls are required. Investigators must rigorously screen waste-derived tissues for infectious agents, genetic abnormalities, and potential contaminants. Long-term safety risks, including immunogenicity, genomic instability, and tumorigenicity must also be assessed. Preclinical and clinical testing frameworks require comprehensive data. Regulators demand robust, long-term safety evidence before approving therapeutic applications (Fig. 6).Fig. 6. The regulatory pipeline encompasses patient-derived sample collection, GMP (Good Manufacturing Practice)- compliant cell processing, laboratory characterization, and ultimately, clinical translation through personalized therapies that span the bench-to-bedside continuum. Created in BioRender.com
Countries worldwide have established regulatory frameworks to oversee the development and clinical use of gene- and cell-based therapies. Safety, efficacy, and quality assurance are the core principles [144, 145].
- United States: the Food and Drug Administration (FDA) regulates these materials as Human Cells, Tissues, and Cellular and Tissue-Based Products (HCT/Ps) under 21 CFR Part 1271. Specifically, regulators primarily determine classification by the degree of manipulation. Products that undergo only “minimal manipulation” and serve homologous use may qualify for less burdensome oversight. In contrast, products that undergo extensive manipulation (e.g., expansion, differentiation, or genetic engineering) fall under the category of biological products and require premarket approval.
- Canada regulates gene and cell therapies as biologics under the Food and Drugs Act and its accompanying regulations, including the Food and Drug Regulations and the Safety of Human Cells, Tissues and Organs for Transplantation Regulations. Cell-based therapies generally involve processing a patient's cells. Gene therapies aim to modify a patient's genetic material to treat conditions. Classification depends on intended use, extent of modification, and associated risks. Health Canada established the Advanced Therapeutic Goods pathway for new products or those that do not easily fit into the existing categories.
- The European Union: The European Medicines Agency (EMA) applies the Advanced Therapy Medicinal Products (ATMP) framework regulated by the EU Directive 2001/83/EC on ATMPs, which also distinguishes between processing levels and therapeutic intent. The EMA provides also provides centralized marketing authorization across EU member states. Programs such as PRIME (PRIority Medicines) and Conditional Marketing Authorization expedite access for therapies targeting severe or rare diseases, while orphan drug incentives encourage innovation in rare disease therapeutics.
- Japan: the Pharmaceuticals Medical Devices Act (PMD Act) regulates cell therapy products as regenerative medicine products. Japan has pioneered progressive regulation through the Act on the Safety of Regenerative Medicine and the Sakigake designation, which enable conditional, time-limited approvals coupled with rigorous post-marketing surveillance. These frameworks establish Japan as a leader in regenerative medicine.
- China: Since 2017, the National Medical Program Administration has reformed China’s approval processes. Regulations ensure GMP compliance and ethical oversight of stem cell research.
- India: The Central Drugs Standard Control Organization (CDSCO) is the national regulatory body. It ensures the safety, efficacy, and quality of drugs, cosmetics, and medical devices. Under the Drugs and Cosmetics Act, CDSCO approves new drugs and clinical trials, regulating drug imports, and setting quality standards.
- Australia: The Therapeutic Goods Administration (TGA) regulates therapies under the Biologicals Regulatory Framework, classifying products based on their risk level. Highly manipulated products face stringent pre-market evaluation. The Gene Technology Act and the National Health and Medical Research Council (NHMRC) ethical guidelines ensure the safe development of genetically modified therapies and ethical conduct in clinical trials.
- Brazil: ANVISA (Brazilian health regulatory agency) regulates ATMPs under RDC No. 260/2018. This covers somatic cell therapy, gene therapy, and tissue engineering. Oversight focuses on manipulation level, cell source, and therapeutic intent, requiring strict adherence to safety, efficacy, and quality standards prior to approval.
Overall, despite national variations, regulatory frameworks share a common foundation of ensuring safety, efficacy, and quality. They also provide accelerated pathways and incentives to encourage innovation. Japan and China emphasize conditional approvals with post-market monitoring, while the United States and Europe focus on rigorous pre-market evaluation and expedited designations. These complementary approaches collectively advance global progress in regenerative medicine.
Ethical and Legal Dimensions
Although waste-derived stem cells circumvent most ethical controversies linked to embryonic stem cell research. Some unresolved issues remain. This include commercialization, donor rights, and equitable access to therapies. In many countries, legislative restrictions on the ownership, transfer, and commercialization of biological waste hinder cross-border collaboration and delay clinical translation.
Efforts to Harmonize International Standards
As gene- and cell-based therapies gain global prominence, regulatory authorities and international organizations increasingly seek to harmonize standards for ATMPs. Harmonization aims to reduce duplication, streamline development, and ensure consistent safety, efficacy, and quality across jurisdictions. International initiatives, such as guidelines from the International Council for Harmonization of Technical Requirements for Pharmaceuticals for Human Use (ICH) and recommendations from the International Society for Stem Cell Research (ISSCR), provide frameworks for ethical conduct, preclinical testing, and clinical evaluation. Regulatory agencies are aligning definitions such as “minimal manipulation” and “homologous use,” facilitating cross-border clinical trials and technology transfer. Accelerated pathways, such as the FDA’s Breakthrough Therapy designation, the EMA’s PRIME scheme, and Japan’s Sakigake designation, expedite the approval of new therapies. These programs maintain rigorous oversight. Harmonized guidelines emphasize GMP-compliant manufacturing, robust potency and identity testing, and strict safety monitoring, including standardized adverse event reporting and post-market surveillance. Despite these advances, differences in legal frameworks, national priorities, and resource levels remain. These variations result in regulatory fragmentation. Ongoing dialogue among regulators, industry, and academic stakeholders aims to establish mutual recognition agreements, standardized terminology, and unified clinical trial protocols. This will accelerate the translation of ATMPs from research to clinical use while safeguarding patient safety and scientific integrity (https://www.ich.org/; https://www.isscr.org/).
Conclusions and Future Challenges
The use of biological waste materials in research and health sectors is central to advancing scientific progress and improving healthcare outcomes. These materials provide a foundation for developing new medical treatments and expanding our understanding of diseases. They also drive biotechnological innovation. As a result, they directly shape cost-effective research and enable breakthroughs in regenerative medicine, drug development, and diagnostic technologies [146].
Building on these promising applications, the translation of stem cells derived from biological waste into therapeutic products necessitates the establishment and maintenance of GMP-compliant facilities. These facilities present significant financial barriers. Production costs per patient dose vary according to cell type, scalability, and the extent of cell manipulation. The primary expenses stem from requirements for sterile infrastructure, rigorous quality control testing, validated processing protocols, and long-term product tracking and monitoring.
To address these challenges, artificial intelligence (AI) is emerging as a transformative tool in optimizing manufacturing protocols for advanced therapies, including stem cell–derived products and other ATMPs. Machine learning algorithms can analyze large datasets generated during cell culture, expansion, and differentiation. They can identify optimal growth conditions and predict batch outcomes. For example, deep learning models applied to imaging data can non-invasively monitor cell morphology and function, reducing reliance on destructive assays while ensuring reproducibility. Similarly, reinforcement learning and digital twin technologies enable the simulation of bioprocess parameters, including nutrient delivery, oxygenation, and shear stress, before implementation. As a result, these tools reduce the experimental burden and the likelihood of manufacturing failures.
Within this context, in GMP settings, AI can support closed-system automation by integrating with robotics and advanced sensors. These systems maintain consistent environmental conditions and minimize human error. They also ensure compliance with regulatory standards. Importantly, AI-powered predictive analytics also enhance supply chain management and cost modeling. This offers the potential to lower the cost of goods sold (COGS), which remains a key barrier to the large-scale clinical translation of ATMPs.
Nevertheless, despite its promise, the integration of AI into manufacturing protocols faces regulatory and ethical challenges. Both the EMA and FDA emphasize the need for transparency, validation, and explainability in AI-driven decision-making. This is necessary to ensure patient safety and regulatory compliance. Harmonized guidelines on AI in ATMP manufacturing are still developing. However, pilot projects suggest that AI-enabled automation can accelerate scale-up while maintaining stringent quality assurance.
In summary, adopting AI in ATMP manufacturing is critical for optimizing processes, reducing variability, and lowering costs. As regulatory frameworks adapt to digital technologies, AI will become integral to ensuring scalable and reliable cell-based therapies, establishing it as a linchpin for future advancements in the field.
The strategic use of biological waste materials is crucial for advancing health research, but it demands careful navigation of legal and ethical complexities. Establishing harmonized frameworks and ethical standards is crucial not only for maximizing benefits but also for supporting responsible and equitable progress in the field.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1 (DOCX 26.4 KB)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Elizondo-Omaña, R. E., Guzmán-López, S., & García-Rodríguez, M. L. (2005). Dissection as a teaching tool: past, present, and future. The Anatomical Record Part B: the New Anatomist, 285(1), 11-5. 10.1002/ar.b.20070. 10.1002/ar.b.2007016032753 · doi ↗ · pubmed ↗
- 2Prioreschi, A. (1991). History of medicine (pp. 182–211). F. Norman Library of Science and Medicine No. 424. Hook & Norman, The Haskell.
- 3Kemp, M. (2006). Leonardo Da vinci: The marvellous works of nature and man. Oxford University Press.10.1136/jmh.2008.00027523674544 · doi ↗ · pubmed ↗
- 4Cunningham, A. (1997). The anatomist anatomis’d: An experimental discipline in enlightenment Europe. Routledge.
- 5Skloot, R. (2010). The immortal life of Henrietta lacks. Crown Publishing Group.
- 6Takahashi, K., Tanabe, K., Ohnuki, M., Narita, M., Ichisaka, T., Tomoda, K., & Yamanaka, S. (2007). Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell, 131(5),861 − 72. 10.1016/j.cell.2007.11.019.10.1016/j.cell.2007.11.01918035408 · doi ↗ · pubmed ↗
- 7Lennon, D. P., & Caplan, A. I. (2006). Isolation of human marrow-derived mesenchymal stem cells. Experimental Hematology, 34(11),1604-5. 10.1016/j.exphem.2006.07.014.10.1016/j.exphem.2006.07.01417046583 · doi ↗ · pubmed ↗
- 8Weiss, M. L., & Troyer, D. L. (2006). Stem cells in the umbilical cord. Stem Cell Reviews and Reports, 2(2),155-62. 10.1007/s 12015-006-0022-y. 10.1007/s 12015-006-0022-y PMC 375320417237554 · doi ↗ · pubmed ↗