A subset of type 4 secretion system effectors of Brucella spp. associates to outer membrane vesicles
Maren Ketterer, Naadir Ganief, Petra Chiquet, Adélie Lannoy, Audrey Verhaeghe, Joanne Braunweiler, Marc Dieu, Xavier De Bolle, Maxime Québatte, Christoph Dehio

TL;DR
Some Brucella proteins use outer membrane vesicles to leave the bacteria, possibly as an alternative to their main secretion system.
Contribution
Identifies a subset of Brucella T4SS effectors that associate with outer membrane vesicles for secretion.
Findings
A subset of T4SS effectors associates with outer membrane vesicles in Brucella species.
BspC and VceA use signal peptides for periplasmic export and subsequent secretion via OMVs.
OMVs may represent an alternative secretion pathway to the T4SS in Brucella.
Abstract
The establishment of a replicative niche in the hostile environment of the host presents an enormous challenge for pathogens. Intracellular pathogens such as Brucella spp., the Gram-negative causative agents of Brucellosis, must subvert diverse host functions to ensure survival and replication. One of the key adaptations to achieve this is the translocation of effector proteins into host cells via its type 4 secretion system (T4SS). But effector identification in Brucella is particularly challenging, as previously identified effectors lack a conserved translocation signal, exhibit variable requirements for translocation, and in some cases appear to be translocated in a T4SS-independent manner. Here, we demonstrate that a subset of previously described T4SS effector proteins associates with outer membrane vesicles (OMVs) in different Brucella species. Most of these effector proteins…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
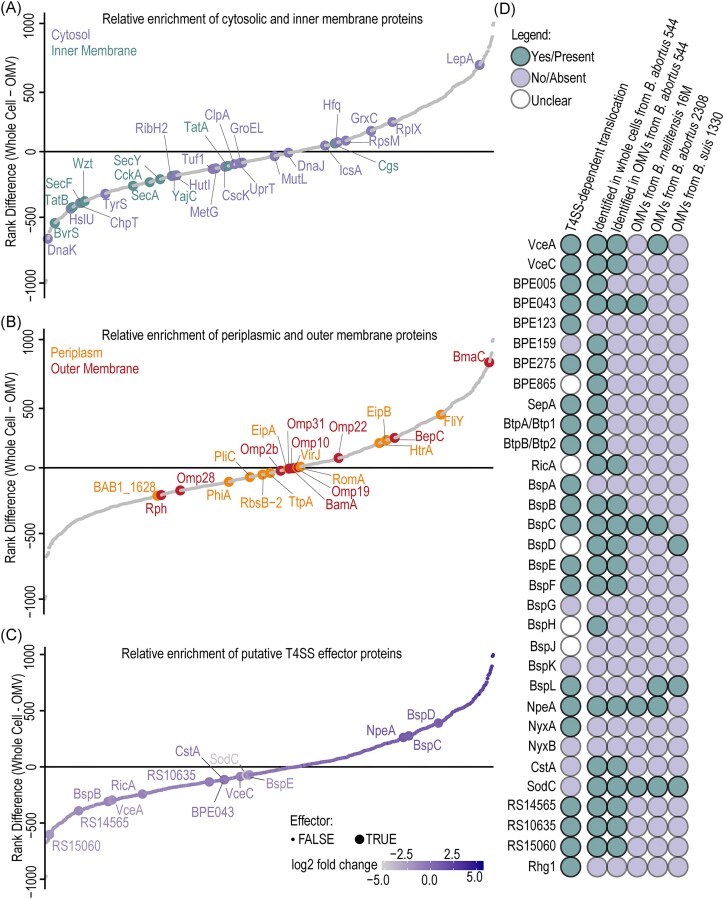 Figure 1
Figure 1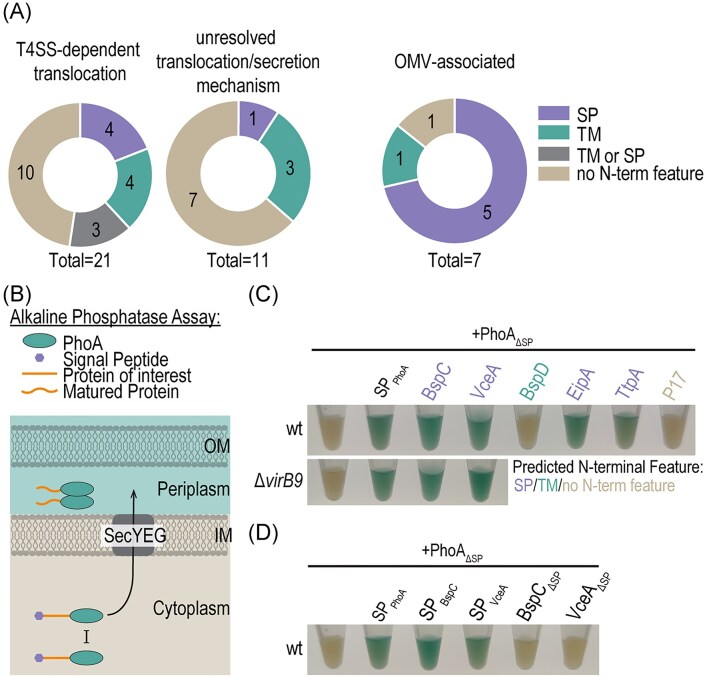 Figure 2
Figure 2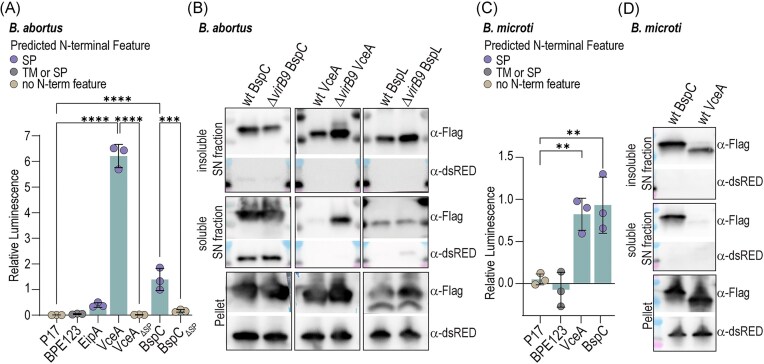 Figure 3
Figure 3- —Swiss National Science Foundation10.13039/501100001711
- —Fédération Wallonie-Bruxelles10.13039/501100002910
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBrucella: diagnosis, epidemiology, treatment · Bacterial Infections and Vaccines · Yersinia bacterium, plague, ectoparasites research
Introduction
Brucella spp. are Gram-negative, facultative intracellular bacteria and the causative agents of the zoonosis Brucellosis. The genus is divided in classical species (e.g. Brucella melitensis, Brucella abortus, and Brucella suis) and “atypical” species (e.g. Brucella microti), distinguished by their bacteriological features, host specificity, and pathogenic potential (reviewed in Occhialini et al. 2022). To ensure survival and replication Brucella manipulates host cell functions through its type 4 secretion system (T4SS), which is essential for these processes. Following uptake into host cells, Brucella initially follows the canonical endocytic pathway (Comerci et al. 2001, Celli et al. 2003, Bellaire et al. 2005, Pizarro-Cerdá et al. 1998a, b), where transient acidification of the late endosome is required for T4SS expression (Porte et al. 1999, Boschiroli et al. 2002, Altamirano-Silva et al. 2018), and effector proteins are thought to mediate conversion of the nonreplicative Brucella containing vacuole (BCV) to the replicative BCV, which is directly connected to the endoplasmic reticulum (ER) (Lestrate et al. 2000, Comerci et al. 2001, Celli et al. 2003, Bellaire et al. 2005, Starr et al. 2008, Sedzicki et al. 2018, Pizarro-Cerdá et al. 1998b).
Interestingly, while the virB operon encoding the T4SS is indispensable for intracellular replication and essential in a transposon mutant screen, deletion of individual effector genes rarely reduces virulence, suggesting redundancy (Myeni et al. 2013, Marchesini et al. 2016, Sternon et al. 2018, Smith et al. 2020, Li et al. 2022). Moreover, the mechanisms governing effector recognition and translocation remain poorly understood. Brucella lacks the classical VirD4 coupling protein, which is essential for effector recognition in related T4SSs (O’Callaghan et al. 1999, Schulein and Dehio 2002, Schulein et al. 2005). Further, Brucella effectors do not share a consensus translocation signal designating them as T4SS substrates, as described in related systems (Huang et al. 2011, Stanger et al. 2017, Wagner et al. 2019). Instead, translocation requirements vary between effectors and may involve the N-terminus (e.g. BPE123; Marchesini et al. 2011), the C-terminus (e.g. VceC; De Jong et al. 2008), or both (e.g. BspA; Myeni et al. 2013). Other effectors (e.g. BspH, BspJ, and BPE865) appear to be translocated independently of the T4SS when N-terminally tagged, yet require the presence of the T4SS when C-terminally tagged (Myeni et al. 2013). And some effectors (e.g. BspG) are entirely T4SS-independent (Myeni et al. 2013). Collectively, these observations suggest that Brucella’s T4SS may exhibit unusual flexibility in substrate recognition and/or may operate in concert with additional, as yet undefined, secretion or translocation pathways that might depend on the T4SS-driven subcellular localization of the bacterium. However, Brucella lacks other classical secretion systems such as the type 2, 3, and 9 secretion systems, and the T4SS is regarded as the principal pathway for effector delivery.
However, several species produce outer membrane vesicles (OMVs) (Avila-Calderón et al. 2012, Pollak et al. 2012, Araiza-Villanueva et al. 2019, 2020, Socorro Ruiz-Palma et al. 2021)—nanostructures ubiquitously shed by Gram-negative bacteria, composed of outer membrane and periplasmic components. These Brucella-derived OMVs can modulate immune responses and promote bacterial internalization, but their molecular cargo and role in pathogenicity and virulence remain poorly understood (Pollak et al. 2012, Avila-Calderón et al. 2020).
Here, we investigated the protein content of OMVs from different Brucella species to determine whether described effector proteins localize to these vesicles. Using comparative proteomics we identified a subset of effectors, which are consistently present in OMVs of different species, many of which contain predicted signal peptides for periplasmic targeting. Focusing on BspC and VceA, we show that both carry functional signal peptides and are therefore exported to the periplasm. They are further secreted to culture supernatant in a T4SS-independent manner, and localize to an insoluble, OMV-containing fraction. These findings suggest that Brucella may use OMVs as an alternative seretion pathway and highlight the need to explore noncanonical secretion mechanisms in this pathogen.
Material and methods
Bacterial strains and growth conditions
Bacterial strains used in this study are listed in Table S1. Escherichia coli strains were cultivated in lysogeny broth (LB) or on Luria–Bertani agar (LA) plates at 37°C overnight supplemented with appropriate antibiotics.
Brucella abortus 2308 and B. microti CCM4915T strains were grown in tryptic soy broth (TSB, Sigma-Aldrich 22 092) or defined medium (DM) (De Barsy et al. 2011) at 37°C with agitation supplemented with appropriate antibiotics. Solid cultures were grown on LA agar or tryptic soy agar (TSA, Difco 236 950) plates with appropriate antibiotics at 37°C. Cultures were inoculated directly from frozen stocks stored at −80°C.
Cloning
All manipulations with DNA were performed following standard techniques, and plasmid integrity was verified by sequencing. Plasmids and primers are listed in Tables S2 and S3, respectively. Plasmids were conjugated into the different Brucella species following standard protocols.
Deletion of virB9 in B. microti
virB9 was deleted from B. microti in frame by double homologous recombination using the SacB allelic exchange suicide vector pNPTS138, which confers kanamycin resistance and sucrose sensitivity upon insertion. Excision of the insertion was counterselected on 5% sucrose TSA plates. For further details see Deghelt et al. (2014). Correct deletion was confirmed by colony polymerase chain reaction (PCR) using primers listed in Table S3 and via sequencing.
Alkaline phosphatase assay
Brucella microti strains expressing the different alkaline phosphatase fusions were grown for 6 h in TSB at 37°C with 170 rpm in presence of 100 ng/ml anhydrotetracycline (aTc) to induce protein expression. Volumes equal to an OD_600nm_ of 2 in 1 ml were spun down, resuspended in 100 µl fresh TSB, 200 μg/ml 5-bromo-4-chloro-3-indolyl phosphate (BCIP, Sigma-Aldrich, B6149) was added, and samples were incubated at 37°C for 1–2 h before imaging.
Axenic secretion assay
Overnight cultures were grown in 2YT in the presence of 34 μg/ml chloramphenicol until stationary phase. The secretion culture was inoculated to an OD_600nm_ of 0.4 in 10 ml DM pH 7, supplemented with 17 μg/ml chloramphenicol, as higher chloramphenicol concentrations inhibited growth in DM. Secretion cultures were incubated at 37°C at 170 rpm for 7.5 h for B. microti and 17 h for B. abortus with 100 ng/ml aTc to induce expression of fusion proteins. Samples were then processed as described below.
Detection of secreted proteins using Nano-Glo HiBiT Detection System
Secretion cultures were set up and incubated as described above. At the endpoint the OD_600nm_ was measured and volumes containing 3.89 × 10^9^ bacteria per ml (OD_600nm_ of 1) were centrifuged twice to remove bacteria. The pellets were set aside for further analysis using Nano-Glo HiBiT Lytic Detection System (Promega, N3030). 100 µl of the cleared supernatants were dispensed in a white 96-well plate in triplicate and equilibrated to RT. Sterile DM was used as negative control. Nano-Glo HiBiT Extracellular Reagent (Promega, N2420) was prepared following manufacturer’s instructions and 100 µl were dispensed to each sample. The plate was sealed (Thermo Scientific, AB-0580) and the bioluminescence was read in a Synergy H4 microplate reader (Agilent Bio Tek) after 10 min of agitation at RT. The reconstitution of the HiBiT and LgBiT fragment in the pellet was analysed using the Nano-Glo HiBiT Lytic Detection System (Promega, N3030), to show that there is general measurable bioluminescence. For this the beforementioned pellets were resuspended in phosphate buffered saline (PBS) and centrifuged to remove residual supernatant. The washed pellets were resuspended in 1 ml PBS and 100 µl was dispensed in triplicate into a white 96-well plate. 100 µl PBS were used as negative control. 50 µl of lysis buffer were added to the bacterial suspension and the samples were incubated at RT for 10 min to lyse the bacteria. The Nano-Glo HiBiT Lytic reagent was prepared following the manufacturers instructions. 50 µl of reagent were added to each well. The plate was sealed (Thermo Scientific, AB-0580) and the bioluminescence was immediately read in a Synergy H4 microplate reader (Agilent Bio Tek).
Fractionation of secretion cultures
Secretion cultures were set up and incubated as described before. After incubation a sample equivalent to OD_600nm_ of 0.6 was pelleted and put aside for western blotting. The rest of the cultures were cleared by centrifugation for 20 min at 3000 × g to pellet bacteria. The cleared supernatants were sterile filtered using Nalgene Rapid-Flow filter units with a PES membrane with 0.22 µm pore size (Nalgene, 564–0020). The filtered supernatant was tested for sterility by plating onto LB agar plates. The filtrate was then either subjected to further fractionation to concentrate OMVs or subjected directly to trichloroacetic acid (TCA)-precipitation.
TCA-precipitation of sterile-filtered supernatant
One volume of 100% TCA was added to four volumes of sterile-filtered supernatant and incubated for 1 h at 4°C. The precipitate was sedimented by centrifugation at 10 000 × g for 1 h at 4°C. The supernatant was carefully removed and discarded. The precipitate was rinsed with 0.2 ml acetone. The acetone was removed after centrifugation at 10 000 × g for 1 h at 4°C. 5 µl TRIS–HCl (pH 8.0) was added to equilibrate the pH. The precipitated supernatants were solubilized in appropriate volumes of 2x Laemmli buffer to normalize the samples to ~1.67 × 10^9^ CFU in 20 µl based on the OD_600nm_ of the original culture.
Western blots of fractionated secretion cultures
Bacteria at a density of OD_600nm_ of 0.6 (~1 × 10^9^ CFU) were pelleted, washed, and resuspended in 70 μl PBS before heat inactivation for 10 min at 95°C. Standard SDS-sample buffer (Laemmli) was added, and samples were boiled a second time.
Equal volumes were subjected to standard SDS-gel electrophoresis (BioRad, Mini-PROTEAN TGX Precast Protein Gels 4 561 094) followed by semidry western blot. Nitrocellulose membranes (Merck, GE10600001) were blocked with 5% skim milk in 0.1% Tween in PBS (PBS/T) for 1 h at room temperature (RT). Blocked membranes were incubated overnight at 4°C with primary antibody against FLAG-tag (Sigma, F1804) or dsRed (Takara, Living colors dsRED poly Ab, 632 496) in blocking buffer. Following three wash steps with PBS/T, the membranes were incubated with secondary antibody (CellSignaling, 7076S or 7074S) in blocking buffer for 1 h at RT. Before developing the membranes, they were again washed three times with PBS/T. Blots were developed using LumiGlo chemiluminescent substrate (Seracare, 5430) and imaged using the Fusion FX device (Vilber).
Fractionation of supernatant into soluble and nonsoluble fraction using ultracentrifugation
Filtered supernatants were further concentrated by ultracentrifugation to separate soluble and insoluble fraction if needed. After filtration the supernatants were subjected to ultracentrifugation in a Hitachi CS150FNX ultracentrifuge at 100 000 × g for 2 h at 4°C. The supernatant was then carefully removed and subjected to TCA-precipitation as described above, while the OMV-fraction was resuspended in 2x Laemmli normalized to initial ODs of secretion cultures (~2 × 10^8^ CFU/μl) to ensure equal loading for western blotting. Samples were boiled for 10 min at 95°C and subjected to western blotting as described above.
Prediction of putative signal peptides
Putative signal peptides and N-proximal transmembrane domains (within N-proximal 50 aa) were predicted using SingalP6.0 (https://services.healthtech.dtu.dk/services/SignalP-6.0/) (Almagro Armenteros et al. 2019), Phobius (https://phobius.sbc.su.se/) (Käll et al. 2004), and DeepTMHMM (https://services.healthtech.dtu.dk/services/DeepTMHMM-1.0/) (Hallgren et al. 2022).
Harvesting OMVs for LC–MS/MS proteomics
Brucella abortus 544 used for the preparation of OMVs for proteomics was grown as described in (Lannoy et al. 2025). In short: six 800 ml cultures were grown for 48 h to stationary phase and inactivated with 0.5% phenol. Inactivated cultures were centrifuged at 8200 × g at 4°C for 20 min. The pellet was set aside. The supernatant was further concentrated using a Pellicon tangential flow filtration system with a 10 kDa cutoff. Concentrated supernatants were centrifuged at 8200 × g at 4°C for 20 min. The supernatant was frozen at −20°C to aggregate the OMVs. After defrosting the supernatant was ultracentrifuged (47 000 × g, 4°C, 3 h) and the pellet containing the OMVs was resuspended in deionized water and dialysed at 4°C with deionized water for 3 days. Samples were stored at −80°C and then OMVs and bacterial pellets were lyophilized by freeze drying (Telstar Cryodos 50).
Preparation of OMVs for LC–MS/MS proteomics
Freeze dryed OMVs were resuspended in ultrapure water (0.4%; w/v) and treated with Triton X100 (1%; w/v) for 1 h at 37°C. The suspension was centrifuged for 3 min at 5000 × g. 10 µl of 100% TCA were added to 900 µl supernatant, vortexed, and incubated for 30 min on ice. Lysates were then centrifuged at 10 000 × g for 15 min at 4°C and washed with ice-cold acetone. The resulting protein pellet was resuspended in 100 µl 2% SDS in ddH_2_O and heated for 5 min at 95°C. The protein concentrations were measured by the Lowry test (Lowry et al. 1951) prior to protein digestion.
Protein digestion for LC–MS/MS proteomics
Protein samples were digested using the Filter Aided Sample Preparation protocol (Wiśniewski et al. 2009). Briefly, 30 KDa molecular weight cut-off filters (Millipore, Amicon Ultra), were washed using 100 µl 0.1% formic acid (FA) and centrifuged at 14 500 rpm for 15 min. Next 40 µg of protein lysate in urea buffer (8 M urea in 0.1 M Tris at pH 8.5) was added to the filter and centrifuged at 14 500 rpm for 15 min. The filter was washed three times using 200 µl urea buffer, discarding the filtrate between each centrifugation step. Proteins were reduced by adding 100 µl 8 mM DTT to each filter then mixed for 1 min at 400 rpm with a thermomixer, and then incubated at 24°C for 15 min before centrifugation at 14 500 rpm for 15 min. After discarding the filtrate, the filter was washed with 100 µl urea buffer. Proteins were then alkylated by adding 100 µl 50 mM iodoacetamide, in urea buffer, and mixing at 400 rpm on a thermomixer before incubating in the dark for 20 min at 24°C. Filters were then centrifuged at 14 500 rpm for 15 min, and washed with 100 µl urea buffer. Samples were then washed three times with 100 µl of 50 mM ammonium bicarbonate buffer (ABC, in ultrapure water). Following this, 0.8 µg of MS grade trypsin (in 80 µl ABC buffer) was added, before overnight incubation at 24°C in a high humidity environment.
Following overnight digestion, peptides were eluted by centrifugation at 14 500 rpm for 10 min, into clean LoBind centrifuge tubes, with an additional 40 µl ABC buffer added to the filter. Penultimately, 10% trifluoroacetic acid (TFA) was added to a final concentration of 0.2% TFA. Samples were subsequently dried in a SpeedVac, and up to 20 µl were transferred to an injection vial.
Proteomics data acquisition
The digest was analysed using nano-LC-ESI-MS/MS tims TOF Pro (Bruker, Billerica, MA, USA) coupled with an UHPLC nanoElute2 (Bruker).
Peptides were separated by nanoUHPLC (nanoElute2, Bruker) on a 75 μm ID, 25 cm C18 column with integrated CaptiveSpray insert (Aurora, ionopticks, Melbourne) at a flow rate of 200 nl/min, at 50°C. LC mobile phase A (0.1% FA in water) and B (acetonitrile with 0.1% FA). Samples were loaded directly on the analytical column at a constant pressure of 800 bar. The 1 µl of digest was injected, and the organic content of the mobile phase was increased linearly from 2% B to 15% in 22 min, from 15% B to 35% in 38 min, from 35% B to 85% in 3 min. Data acquisition on the tims TOF Pro was performed using Hystar 6.1 and time Control 2.0. tims TOF Pro data were acquired using 160 ms TIMS accumulation time, mobility (1/K0) range from 0.75 to 1.42 Vs/cm². Mass-spectrometric analysis were carried out using the parallel accumulation serial fragmentation (PASEF) (Meier et al. 2018) acquisition method. One MS spectra followed by six PASEF MSMS spectra per total cycle of 1.16 s. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE (Perez-Riverol et al. 2025) partner repository with the dataset identifier PXD067564.
Proteomics database search
Resulting d files were searched with FragPipe using msFragger version 4.1 and IonQuant version 1.10.27, with the default parameters for the LFQ-MBR workflow (Yu et al. 2021, 2025). Briefly, precursor and fragment mass tolerance were set to 20 ppm. The in silico peptide digestion parameters were set to use strict trypsin rules, with a maximum of 2 missed cleavages. Peptides between 7 and 50 amino acids long, and masses between 500 and 5000 daltons, peptides were generated. Methionine oxidation and N-terminal acetylation were set to variable modifications and cystein carbomidomethylation was set as a fixed modification. All results were filtered to 0.01% FDR at the precursor and protein-level. All files were searched against the UniProt (The UniProt Consortium et al. 2023) fasta database for *B. abortus (strain 2308)—*UP000002719 (downloaded on 21 July 2025). Proteins were considered “identified” if proteotypic peptides for a given protein below the FDR cut-off were identified in at least one sample. Proteins were considered “quantified” if for a given protein in at least two samples the MaxLFQ Intensity was >0. i.e, there was at least one shared peptide, between samples.
Bioinformatic analyses
OMV vs whole cell rank difference
Quantified proteins in both the OMV and whole cell fractions were ranked from highest to lowest according to their MaxLFQ intensities in each fraction. The rank difference was calculated by subtracting the OMV rank from the whole cell rank. A positive rank difference indicates relative enrichment in OMVs and a negative rank difference indicates a relative enrichment in whole cells. We also calculated the log2 Fold Change between proteins quantified in both OMVs and whole-cells.
Orthogroup mapping
Orthogroups were generated using OrthoFinder version 2.5.5 (Emms and Kelly 2019), with default settings, to match Brucella proteins between species. Which allowed us to directly compare published proteomes at the orthogroup level. The protein sequence databases for B. melitensis biotype 1 (UP000000419), B. suis biovar 1 (strain 1330) (UP000007104), and B. abortus (strain 2308) (UP000002719), were downloaded from uniprot on 21 July 2025. The orthogroup mappings can be found in Data S1.
Statistical analysis
Graphs were created with GraphPad Prism 8. Statistical analysis was performed using GraphPad Prism with ordinary one-way ANOVA. The number of independent replicates is indicated in the figure legends as n.
Results
Analysis of B. abortus whole-cell and OMV proteomes
Given that various Brucella species produce OMVs (Avila-Calderón et al. 2012, Pollak et al. 2012, Araiza-Villanueva et al. 2019, 2020, Socorro Ruiz-Palma et al. 2021) and that these vesicles have been shown to modulate immune responses, induce cytoskeleton rearrangements, and promote bacterial internalization in cellulo (Pollak et al. 2012, Avila-Calderón et al. 2020), we sought to determine if Brucella effector proteins localize to OMVs, which would suggest that OMVs represent a previously uncharacterized route for effector secretion.
To investigate the protein content and the relative abundance of described effector proteins in OMVs, we purified OMVs from B. abortus as described in Lannoy et al. (2025). The OMV- and whole-cell-fractions were analysed by LC–MS/MS. In total, we identified 18 016 distinct B. abortus peptide sequences corresponding to 1947 proteins at an FDR threshold of 0.01%. Among these, 1186 proteins were identified in both OMV and whole cell-fractions, and 1153 quantified in both. Here, we distinguish between identification and quantification based on the number of unique peptides detected: proteins were considered quantified only if at least one shared peptide was identified across two of three samples. 25 proteins were identified exclusively in the OMV-fractions, of which 21 were quantified, while 717 proteins were identified only in the whole cell-fraction, with 372 quantified. The large number of proteins detected exclusively in the whole cell-fraction, but absent from the OMV-fraction, argues against widespread bacterial lysis as the source of OMV-associated proteins.
Tables of all identifications and quantifications, as well as associated scores are available in Data S2 and S3, including tables of exclusively identified and quantified proteins.
Inner membrane and cytosolic proteins are depleted from the OMV fraction
To assess the quality of our OMV-preparation, we ranked all proteins quantified at least once in OMVs and whole-cell samples by abundance and calculated their log_2_ fold changes (Data S4). In this analysis, a positive rank denotes enrichment in the OMV-fraction, whereas a negative rank denotes higher abundance in whole-cells. As reference groups, we then selected 19 cytosolic, 10 inner membrane-associated, 10 outer membrane-associated, and 11 periplasmic proteins to evaluate sample purity. Inner membrane proteins were strongly depleted from the OMV-containing fractions (9 of 10 proteins with negative rank differences and all with negative log_2_ fold changes) (Fig. 1A). Cytosolic proteins were also mostly depleted from the OMV-containing fraction, with most displaying negative rank differences and log_2_ fold changes (Fig. 1A). By contrast, outer membrane and periplasmic proteins were more evenly distributed, with 6 of 10 outer membrane and 6 of 11 periplasmic proteins showing depletion (Fig. 1B). Among the outer membrane proteins, the adhesin BmaC (Bialer et al. 2021) was highly enriched in the OMV-containing fraction (log_2_ fold change = 2.56), whereas Omp2b and Omp28 were strongly depleted (log_2_ fold change of −1.91 and −1.84, respectively) (Fig. 1B and Data S4), which is consistent with Omp2b being covalentaly linked to peptidoglycan and might be true for the other depleted Omps (Godessart et al. 2021).
A subset of putative Brucella effectors associates with OMVs. (A and B) Proteins quantified at least once in OMV and whole-cell fractions of B. abortus 544 were ranked by abundance in descending order. The difference in rank (whole-cell rank—OMV rank) was plotted, with positive values indicating enrichment in OMVs and negative values enrichment in the whole-cells. To assess OMV preparation quality, the relative abundance of 19 cytosolic, 10 inner membrane-associated, 11 periplasmic proteins, and 10 outer membrane proteins was evaluated. (C) Effectors quantified at least once in OMV and whole-cell fractions were ranked by abundance in descending order. (D) In B. abortus 544, 23 described effector proteins were identified in the whole-cell fraction and 15 in the OMV-fraction. Comparison with published datasets revealed that a subset of putative Brucella effector proteins associates with OMVs in different Brucella species. OMV proteomes of B. melitensis 16 M (Avila-Calderón et al. 2020), B. abortus 2308 (Araiza-Villanueva et al. 2019), and B. suis 1330 (Socorro Ruiz-Palma et al. 2021) were analysed. T4SS-dependency was inferred from the literature.
These results indicate minor cytosolic contamination, which is expected as bacteria were grown to stationary phase and some lysis may have occurred, or alternatively that certain cytoplasmic proteins selectively associate with OMVs by an unknown mechanism. Nevertheless, the strong depletion of inner membrane proteins from the OMV-preparations, together with the exclusive identification of 717 proteins in whole-cell fractions (see above), suggests that the OMV preparation was relatively clean.
A subset of putative Brucella effectors localize to OMVs
Next, we investigated whether described effector proteins localize to OMVs and whether they are enriched in these fractions. Of the 32 described effectors, 23 were identified in the whole-cell fraction, and 15 of these were also detected in the OMV-containing fractions, indicating that a subset of effectors associates with OMVs (Fig. 1C and D). BspF was detected only once in both fractions and was therefore not quantified. Similary, BPE275 and BtpB were detected only once in the whole-cell fraction and were excluded from quantification.
To assess enrichment, proteins quantified at least once in OMV and whole-cell factions were ranked by abundance (Fig. 1C, Data S4 and S5). Log2 fold-change analysis revealed a strong correlation with rank differences (Fig S1, Data S4). In this analysis, NpeA, BspC, and BspD were enriched in OMVs releative to whole-cells, with a rank difference of 262, 276, and 390, respectively. We further compared the protein contents of B. abortus OMVs from this study with previously published datasets of OMVs from B. abortus, B. melitensis, and B. suis (Avila-Calderón et al. 2012, Araiza-Villanueva et al. 2019, Socorro Ruiz-Palma et al. 2021) (Fig. 1D, Data S6). Across at least two of the four datasets analysed, VceA, BspC, BspD, BspL, BPE043, NpeA, and SodC were consistently associated with OMVs (Data S6). Of these, five proteins (VceA, BPE043, BspC, BspL, and NpeA) have been reported as T4SS-dependent effectors, one (BspD) has an undefined translocation mechanism, and one (SodC) is T4SS-independent (De Jong et al. 2008, Marchesini et al. 2011, Myeni et al. 2013, Liu et al. 2018, Luizet et al. 2021, Giménez et al. 2024). Notably, the Brucella species have been cultured under different conditions and OMVs were enriched using different methods, suggesting that effector association with OMVs is robust and not an experimental artifact.
Together, these results demonstrate that effector proteins of Brucella can localize to OMVs and suggest that OMVs may served as an alternative pathway for the secretion of virulence factors, a phenomenon likely overlooked in the past due to methodological limitations.
A subset of putative effector proteins encode predicted signal peptides and transmembrane domains
Next, we asked whether OMV-associated effector proteins share any structural features such as signal peptides (SPs) or transmembrane domains (TMs). Canonical SPs consist of a positively charged N-terminal region, a central hydrophobic h-region, and a polar C-terminal region containing the cleavage site for signal peptidase processing (von Heijne 1990). Proteins with classical SPs are typically secreted to the periplasm via the conserved SecYEG translocon of the inner membrane, with assistance from SRP/YidC or SecA (Walter et al. 1981, Gilmore et al. 1982).
In silico analysis with SignalP6.0 (Almagro Armenteros et al. 2019), Phobius (Käll et al. 2004), and DeepTMHMM (Hallgren et al. 2022) predicted SPs and N-proximal TMs in a subset of Brucella effector proteins (Fig. 2A , Tables S4 and S5). VceA, BspC, BspL, and SodC displayed strong SP predictions, while NpeA carried a predicted lipoprotein SP, as previously reported (Giménez et al. 2024). For BPE123, RS10635, and BspB, Phobius predicted SPs that were not revealed by SignalP6.0, whereas DeepTMHMM instead predicted N-terminal TMs for these proteins. N-proximal TMs were consistently predicted for VceC, BPE159, BPE275, BspA, BspD, BspK, and Rhg1. In total, 15 of 32 putative secreted or translocated Brucella effector proteins encoded predicted N-terminal features. Of particular note, among the seven effector proteins localized to OMVs in at least two independent datasets (Fig. 1D), five (VceA, BspC, BspL, NpeA, and SodC) carried predicted SPs, one (BspD) carried a predicted TM, and one (BPE043) lacked any predicted N-terminal feature (Fig. 2A). Importantly, all effector proteins with predicted SPs were found to localize to OMVs but periplasmic proteins were not generally enriched in the OMVs, strongly suggesting that Brucella employs OMVs as vehicles for SP-dependent secretion of virulence factors.
The OMV-associated effectors VceA and BspC encode functional SPs. (A) In silico prediction of N-terminal features of secreted and translocated Brucella proteins identified three groups: proteins with predicted N-proximal TM domains, proteins with SPs, and proteins with ambiguous predictions (TM or SP or no N-terminal feature). (B) Schematic representation of the alkaline phosphatase (PhoA) assay. PhoA dimerizes and becomes active only in nonreducing environments (periplasm or extracellular milieu), but remains inactive in the reducing cytoplasm. SPs are cleaved during export via the SecYEG translocon. OM = outer membrane, IM = inner membrane, and SecYEG = Sec translocon. (C and D) Brucella microti strains expressing the indicated PhoA-fusion proteins were grown to exponential phase in TSB. Cells corresponding to an OD600nm of 2 (per 1 ml culture) were pelleted, resuspended in 100 µl fresh TSB supplemented with 200 µg/ml BCIP, and incubated for 1–2 h at 37°C before imaging. Predicted N-terminal features are indicated by colours. Shown is one representative experiment of two independent replicates.
The putative effector proteins BspC and VceA are exported from the cytoplasm to the periplasm via the SecYEG-translocon
We further investigated the functionality of the predicted SPs. For this purpose, we selected VceA, BspC, and BspD for further validation because (i) the SP-predictions for BspC and VceA were particularly strong, while BspD has a predicted TM, and (ii) they represent different classes of putative effectors: those which can localize to OMVs (VceA), and those which are enriched in OMVs (BspC and BspD).
We used the Escherichia coli protein alkaline phosphatase (PhoA) as a reporter (Kumamoto et al. 1984, Kim and Harold 1991, Herrou et al. 2019a, b) in combination with B. microti, a Biosafety-level 2-compatible surrogate for human-pathogenic Brucella species. PhoA becomes active only when exported to the nonreducing environment of the periplasm via the SecYEG-translocon, where it catalyses the hydrolysis of organic phosphate esters (Kumamoto et al. 1984, Kim and Harold 1991). This activity can be visualized colorimetrically using substrates such as BCIP disodium salt, which is colorless but turns blue upon phosphate hydrolysis and poorly penetrates the inner membrane (Fig. 2B).
PhoA-fusions were expressed from a plasmid under an aTc-inducible promoter in Brucella. Replacement of the native PhoA SP (PhoA^M1-A21^, hereafter SP_PhoA_) with a Flag-tag (yielding Flag-PhoA^R22-K471^, hereafter PhoA_∆SP_), abolished periplasmic export while maintaining expression (Fig. 2C, Fig S2). Export was reconstituted by fusing SP_PhoA_ to the N-terminus of PhoA_∆SP_ (Fig. 2C). Control fusions confirmed the localization: the cytoplasmic protein P17 showed no BCIP conversion, whereas the periplasmic proteins EipA and TtpA resulted in blue coloration consistent with periplasmic localization (Fig. 2C). Using this assay, we found that the putative T4SS effectors VceA and BspC localized to the periplasm in a SP-dependent, and thus Sec-dependent, manner (Fig. 2C and D). In contrast, the envelope integrity protein and putative effector BspD (Myeni et al. 2013, Ketterer et al. 2024) was not exported, consistent with the absence of a predicted SP and presence of a predicted N-terminal TM (Table S5, Fig. 2C). We further tested whether export of VceA and BspC to the periplasm was T4SS-independent, as the association to OMVs suggests, and indeed, export was unaffected in a B. microti ∆virB9 mutant, which cannot assemble a functional T4SS (O’Callaghan et al. 1999).
In summary, VceA and BspC are exported from the cytoplasm to the periplasm in a SP-dependent, T4SS-independent manner via the SecYEG translocon.
VceA and BspC are secreted to the culture supernatant in a SP-dependent and T4SS-independent manner
We next tested whether the T4SS effectors VceA and BspC (De Jong et al. 2008, Myeni et al. 2013), which are exported to the periplasm via the SecYEG-translocon (Fig. 2C) and are associated to OMVs (Fig. 1C and D), could be secreted into the culture supernatant in a SP-dependent manner.
For this purpose, we employed a split NanoGlo (NGlo) luciferase assay, previously applied to monitor effector protein translocation in host cells by the T3SS of Salmonella and T4SS of Bartonella (Westerhausen et al. 2020, Fromm et al. 2022, 2024), as well as to quantify OMV production in culture supernatants of Bacteroides (Pardue et al. 2024). Proteins of interest were fused to the HiBiT fragment and expressed in B. abortus, while LgBiT was supplemented to cleared supernatants (to detect secreted proteins) and to lysed bacterial pellets (to measure input). Brucella abortus cultures were grown to an OD_600nm_ of 1.0–1.4 in a DM as described previously (De Barsy et al. 2011) and pelleted, and supernatants were clarified by additional centrifugation.
As expected, the cytoplasmic protein P17, which was strongly depleted from our OMV-preparation (rank difference: −184, log2 fold change: −1.43) (Data S4), was not detected in supernatants, confirming the absence of bacterial lysis (Fig. 3A and Fig S3A). Similarly, the effector protein BPE123, not associated to OMVs (Fig. 1B), was undectectable in the culture supernatant. The periplasmic protein EipA (Herrou et al. 2019a), which was depleted from our OMV-preparation (rank difference of −7, log2 fold change of −0.5) (Data S4), also did not appear in supernatants (Fig. 3A and Fig S3A), showing that overexpression of periplasmic proteins alone does not drive OMV release. By contrast, both VceA and BspC were secreted in a SP-dependent manner (Fig. 3A, Fig S3A), with VceA showing a ~5–6 fold enrichment in the supernatant relative to the pellet. Secretion of BspC and VceA was also detected in B. microti (Fig. 3C, Fig S3B), indicating that this phenomenon is conserved across Brucella species. However, while VceA was highly enriched in the OMV fraction from the B. abortus 2308 culture it was not enriched in the OMV fraction of the B. microti culture relative to pellet (Fig. 3A and C), indicating potential differences between species and strains in respect to sorting and packaging, as we did not observe enrichment of VceA in the OMV fraction from B. abortus 544 cultures. Notably, B. abortus 544 was grown to stationary phase in rich medium, while B. abortus 2308 and B. microti were grown to exponential phase in DM, which might also account for some of the observed differences.
Brucella spp. secrete VceA, BspC, and BspL to the culture supernatant. (A) Brucella abortus strains expressing the indicated HiBiT fusion proteins were grown to exponential phase in 2YT. Secretion cultures were incubated for 17 h in DM, after which bacteria were pelleted and cleared supernatants collected. Bacterial pellets were lysed, and both fractions were supplemented with LgBiT and substrate. Bioluminescence was measured using a plate reader, and relative luminescence was calculated as the ratio of cleared supernatant versus lysed pellet signals. Ordinary one-way ANOVA; n = 3. Predicted N-terminal features are indicated by colors. (B) Axenic secretion assay with B. abortus evaluating localization of BspC, VceA, and BspL in soluble (OMV-depleted) versus insoluble (OMV-containing) supernatant fractions. Sterile-filtered supernatants were fractionated by ultracentrifugation, and both fractions, alongside bacterial pellets, were analysed by SDS-PAGE and western blotting using α-Flag and α-dsRED antibodies. Shown is one exemplary blot of three independent replicates. (C) Brucella microti strains expressing the indicated HiBiT fusion proteins were analyzed as described in (A), however, due to their faster growth rate, the incubation time in DM was reduced to 8 h. n = 3. (D) Axenic secretion assay with B. microti for evaluating localization of BspC and VceA as described in (B). Shown is one exemplary blot of three independent replicates.
To confirm OMV association, sterile-filtered supernatants were fractionated by ultracentrifugation into soluble (OMV-depleted) and insoluble (OMV-containing) fractions and analysed by SDS-PAGE and western blotting (Fig. 3B). VceA as well as BspL (Luizet et al. 2021) were enriched in the insoluble, OMV-containing fraction without evidence of bacterial lysis, as assessed by absence of cytosolic dsRED, which was coexpressed from the same plasmid. In contrast, BspC overexpression resulted in partial lysis, as indicated by the presence of both dsRED and BspC in the soluble fraction (Fig. 3B).
To assess T4SS dependency, we repeated the assay in a B. abortus ΔvirB9 mutant lacking a functional T4SS (Fig. 3B). Secretion of VceA and BspC remained unaffected, confirming T4SS independence. Interestingly, VceA was additionally detected in the soluble fraction of the ΔvirB9 mutant in the absence of dsRED, though its enrichment in the insoluble fraction was preserved (Fig. 3B). Finally, BspC and VceA were again detected in OMV-containing fractions of B. microti (Fig. 3D), confirming that effector localization to OMVs is conserved across Brucella species.
Together, these data demonstrate that a subset of described T4SS effector proteins, including VceA and BspC, are secreted into the culture supernatant in a SP-dependent, T4SS-independent manner and associate with OMVs, a process conserved among Brucella species.
Discussion
One of the major challenges Brucella faces toward the establishment of its replicative niche in the ER is the manipulation of host cell processes during intracellular trafficking. Although Brucella was first described over a century ago by Sir David Bruce, the precise molecular mechanisms underlying its intracellular lifestyle remain incompletely understood. It is well established that Brucella depends on its T4SS for intracellular replication, and an increasing number of putative T4SS effectors have been identified in recent years. However, how these effector proteins are translocated into host cells and how they contribute to pathogenesis remains poorly defined. Translocation assay data are often inconsistent (Myeni et al. 2013), and the existence of T4SS-independent effector proteins suggests that additional secretion mechanisms are involved.
Several Brucella species are known to produce OMVs and OMV-resembling structures can be detected in the vicinity of bacteria during monocyte infection (Boigegrain et al. 2004, Avila-Calderón et al. 2012, 2020, Pollak et al. 2012, Araiza-Villanueva et al. 2019, Socorro Ruiz-Palma et al. 2021). Further, B. melitensis OMVs induced TNFα and IL-6 secretion and cytoskeletal rearrangements in PBMCs, but not IL-17 secretion, apoptosis, DNA damage, or proliferation; these effects were independent of LPS phenotype (Avila-Calderón et al. 2020). This demonstrates that Brucella OMVs can modulate immune responses, although the underlying factors and in vivo relevance remain unclear. We therefore hypothesized that OMVs might represent an alternative route for effector protein secretion. Indeed, our study demonstrates that several described T4SS effectors are consistently associated with OMVs produced by Brucella spp.. Strinkingly, especially effector proteins with predicted SPs were detected in OMVs, suggesting that SPs may be a prerequisite for OMV targeting. However, periplasmic proteins were not generally enriched in OMVs compared to whole-cells, implying the involvement of additional sorting mechanisms.
In our analysis, 15 of 32 described effector proteins were detected in B. abortus OMVs grown to stationary phase in rich medium. Comparative proteomics with previously published datasets from B. melitensis, B. abortus, and B. suis revealed seven effectors (VceA, BspC, BspD, BspL, BPE043, NpeA, and SodC) consistently present in at least two of four datasets, suggesting robust OMV association. NpeA, BspC, and BspD were enriched in OMVs relative to whole-cells in our dataset. Importantly, these findings were made under axenic growth conditions, and effector expression is likely to differ during host infection, potentially increasing the repertoire of OMV-associated proteins. It will therefore be important to study the influence of infection-mimicking conditions (such as acidic pH and nutrient limitation) on OMV-cargo composition in the future. The enrichment of certain proteins in the OMVs compared to the whole-cell and the association of several effector proteins with OMVs of different Brucella species may favor a model of controlled vesicle biogenesis rather than passive blebbing, suggesting that OMVs could serve as specific vehicles for virulence factor secretion. It should thus be explored in the future if the OMVs of Brucella spp. mediate direct translocation of effectors into host cells by adapting and utilizing reporter assays such as the TEM1 β-lactamase assay. Brucella, as a facultative intracellular pathogen, may employ OMVs at defined infection stages. OMVs could be produced within a host membrane-bound compartment as described for Salmonella enterica serovar Typhimurium, and released to the host cell cytosol (Yoon et al. 2011, Guidi et al. 2013). Alternatively, OMV release could occur upon T4SS-dependent damage of the Brucella-containing vacuole (BCV), observed as early as 6 h postinfection (Costa Franco et al. 2018, Tana et al. 2021, Hiyoshi et al. 2022). Such BCV damage could release luminal components, including OMVs, or even bacteria into the cytoplasm. Whether Brucella produces OMVs in cellulo or in vivo remains an open question.
For other pathogens, the role of OMVs in virulence is well documented. Helicobacter pylori OMVs deliver VacA toxin, detectable in gastric biopsies (Fiocca et al. 1999). Salmonella enterica serovar Typhimurium produces toxin-containing OMVs inside host cells, which are secreted and taken up by neighboring cells (Guidi et al. 2013). Coxiella burnetii appears to release periplasmic proteins into host cell cytosol, and OMVs have been proposed as transport vehicles (Stead et al. 2013). Legionella pneumophila OMVs inhibit phagosome–lysosome fusion (Fernandez-Moreira et al. 2006) and promote replication when host cells are pretreated with OMVs (Jung et al. 2016). Interestingly, Legionella T4SS mutants remain viable but do not follow normal intracellular trafficking, raising the possibility that OMVs partially compensate for T4SS function (Fernandez-Moreira et al. 2006). Similarly, Brucella T4SS mutants remain viable but fail to replicate or follow canonical intracellular trafficking (Comerci et al. 2001, Celli et al. 2003). This raises the possibility that additional secretion mechanisms, including OMV production inside host cells, contribute to intracellular survival.
In conclusion, although we do not demonstrate effector secretion into host cells, our data show that multiple Brucella effector proteins are reproducibly associated with OMVs, indicating that OMVs may represent a potential pathway for virulence factor export. The selective enrichment of specific effectors in OMVs supports a model of regulated vesicle biogenesis rather than passive release. However, functional validation of OMV-mediated effector delivery and analyses under infection-mimicking or in cellulo conditions will be essential to determine the biological relevance of this pathway. Such studies will be critical to establish whether OMVs contribute directly to effector translocation and Brucella pathogenesis during host infection.
Supplementary Material
uqaf046_Supplemental_Files
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Almagro Armenteros JJ, Tsirigos KD, Sønderby CK et al. Signal P 5.0 improves signal peptide predictions using deep neural networks. Nat Biotechnol. 2019;37:420–3. 10.1038/s 41587-019-0036-z.30778233 · doi ↗ · pubmed ↗
- 2Altamirano-Silva P, Meza-Torres J, Castillo-Zeledon A et al. Brucella abortus senses the intracellular environment through the Bvr R/Bvr S two-component system, which allows B. abortus to adapt to its replicative niche. Infect Immun. 2018;86:1–15. 10.1128/IAI.00713-17.PMC 586502829378792 · doi ↗ · pubmed ↗
- 3Araiza-Villanueva M, Avila-Calderón ED, Flores-Romo L et al. Proteomic analysis of membrane blebs of Brucella abortus 2308 and RB 51 and their evaluation as an acellular vaccine. Front Microbiol. 2019;10. 10.3389/fmicb.2019.02714.PMC 689501231849872 · doi ↗ · pubmed ↗
- 4Avila-Calderón ED, Lopez-Merino A, Jain N et al. Characterization of outer membrane vesicles from Brucella melitensis and protection induced in mice. Clin Dev Immunol. 2012;2012:352493.22242036 10.1155/2012/352493 PMC 3254011 · doi ↗ · pubmed ↗
- 5Avila-Calderón ED, Medina-Chávez O, Flores-Romo L et al. Outer membrane vesicles from Brucella melitensis modulate immune response and induce cytoskeleton rearrangement in peripheral blood mononuclear cells. Front Microbiol. 2020;11.10.3389/fmicb.2020.556795 PMC 760430333193138 · doi ↗ · pubmed ↗
- 6The Uni Prot Consortium, Bateman A, Martin M-J et al. Uni Prot: the Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023;51:D 523–31.36408920 10.1093/nar/gkac 1052 PMC 9825514 · doi ↗ · pubmed ↗
- 7Bellaire BH, Roop RM, Cardelli JA. Opsonized virulent Brucella abortus replicates within nonacidic, endoplasmic reticulum-negative, LAMP-1-positive phagosomes in human monocytes. Infect Immun. 2005;73:3702–13. 10.1128/IAI.73.6.3702-3713.2005.15908400 PMC 1111828 · doi ↗ · pubmed ↗
- 8Bialer MG, Ferrero MC, Delpino MV et al. Adhesive functions or pseudogenization of type va autotransporters in Brucella species. Front Cell Infect Microbiol. 2021;11:607610. 10.3389/fcimb.2021.607610.33987105 PMC 8111173 · doi ↗ · pubmed ↗