The CELL NUMBER REGULATOR SlFWL5 protein regulates aerial vegetative growth in tomato, by promoting cell expansion
Arthur Beauchet, Lucie Ehrhard, Lina Boutaleb, Valérie Rofidal, Nathalie Gonzalez, Christian Chevalier, Norbert Bollier

TL;DR
The SlFWL5 protein in tomatoes helps control leaf and stem growth by promoting cell expansion, contrary to its expected role in regulating cell number.
Contribution
SlFWL5 is shown to regulate aerial vegetative growth in tomato through cell expansion, not cell division.
Findings
SlFWL5 is localized at plasmodesmata and regulates leaf size and morphology.
SlFWL5 promotes hypocotyl growth by enhancing cell expansion.
The protein's role in cell expansion challenges its classification as a cell number regulator.
Abstract
The FW2.2-LIKE/CELL NUMBER REGULATOR (FWL/CNR) gene family comprises PLAC8 domain-containing membrane-associated proteins and was named in reference to its founding member, the FW2.2 gene, which determines fruit size in tomato via a negative regulation on cell divisions. The function of PLAC8 domain-containing FWL/CNR proteins in plants remains largely unexplored. Only recently has the molecular and cellular mechanism of FW2.2 been described as regulating plasmodesmata-mediated cell-to-cell communication. In the present study, we provided a functional analysis of SlFWL genes in tomato, aiming at investigating any direct role in the control of organ growth. Based on a combination of molecular and cellular approaches, we selected three SlFWL proteins, namely SlFWL2, -4, and -5, which are localized at the plasma membrane. Gain- and loss-of-function transgenic plants were generated to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
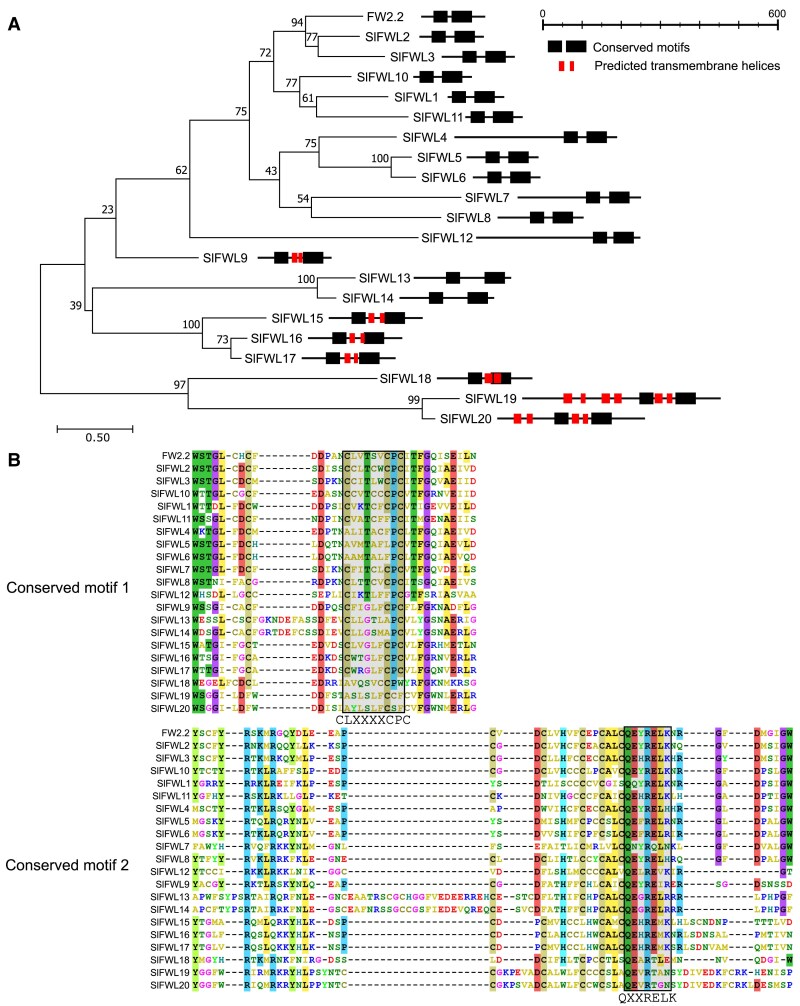 Fig. 1
Fig. 1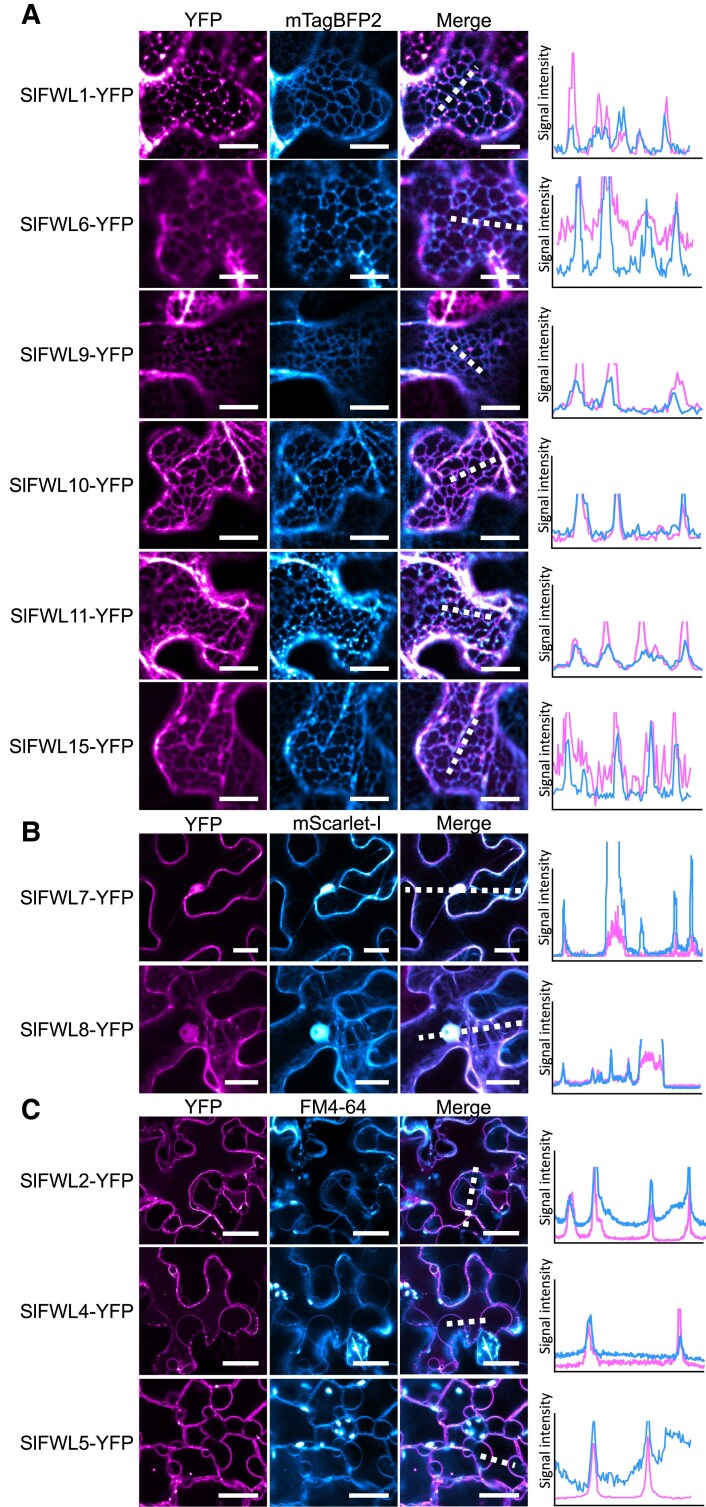 Fig. 2
Fig. 2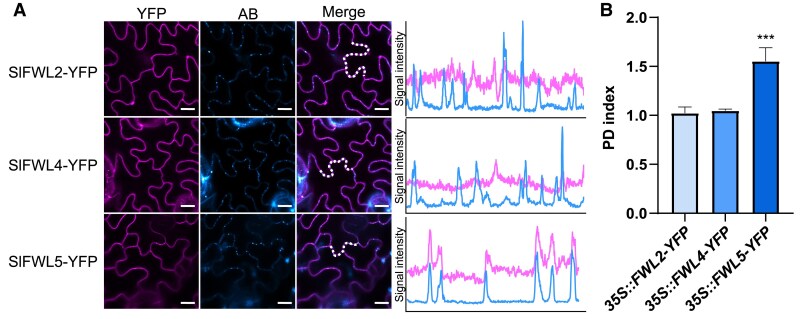 Fig. 3
Fig. 3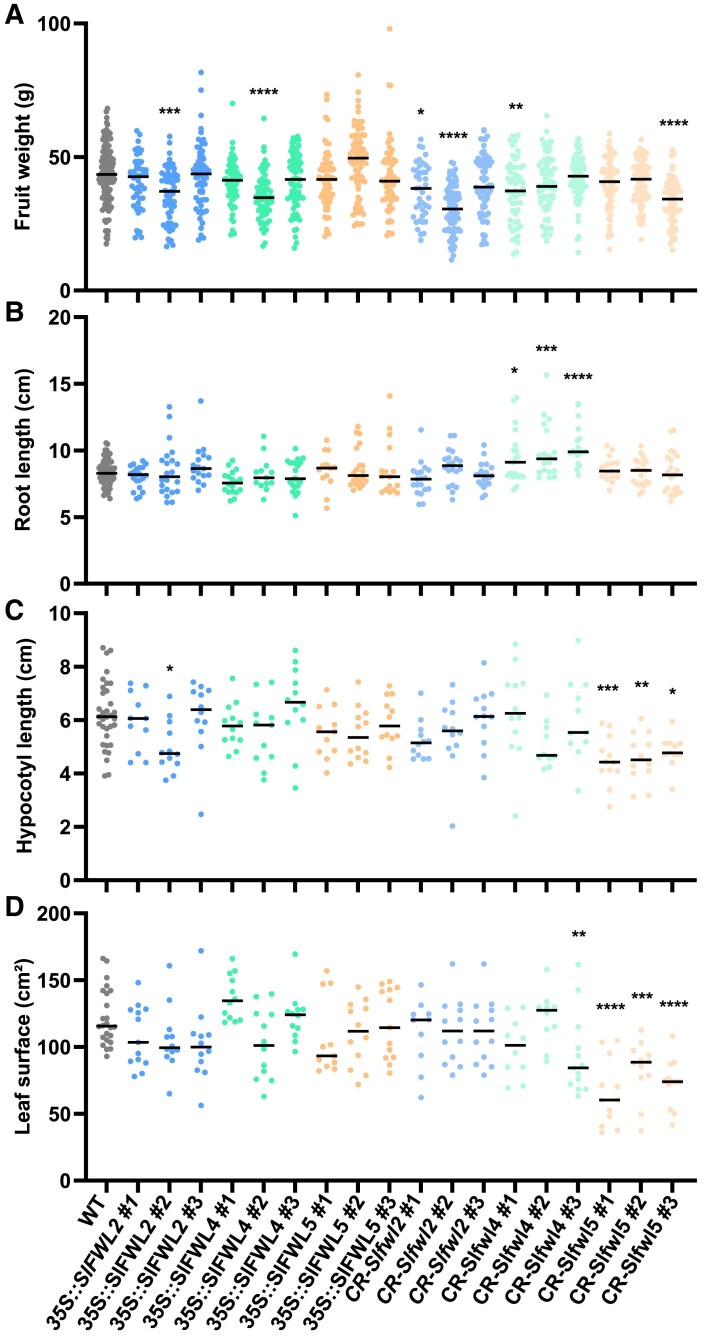 Fig. 4
Fig. 4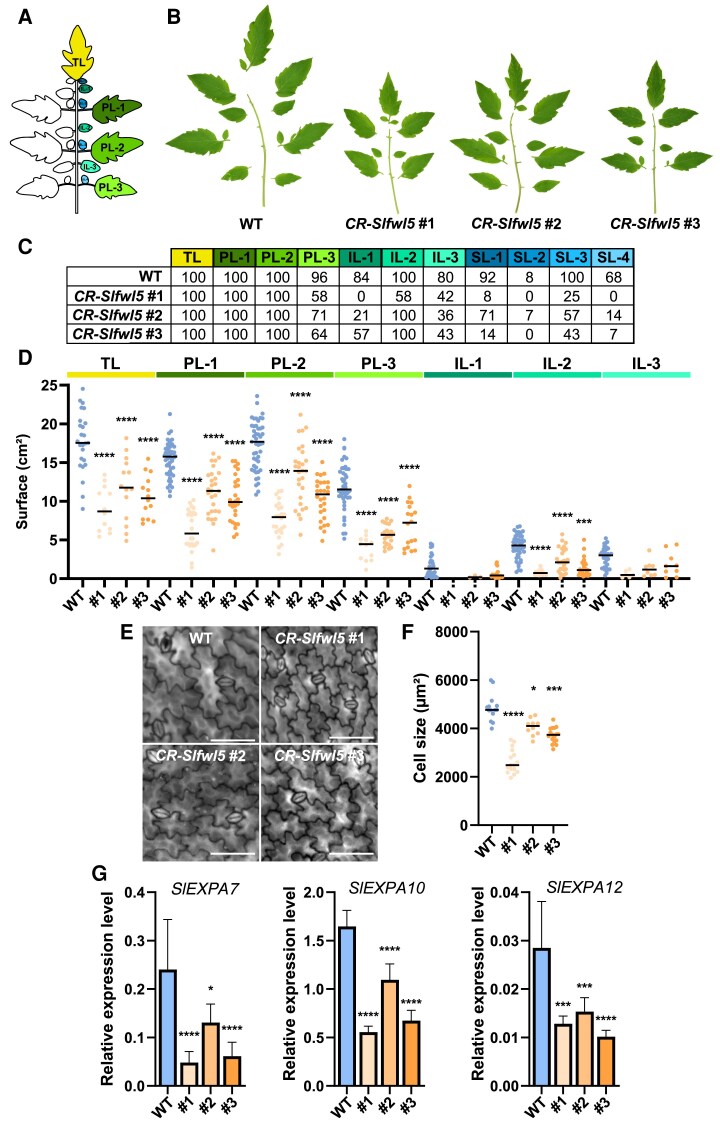 Fig. 5
Fig. 5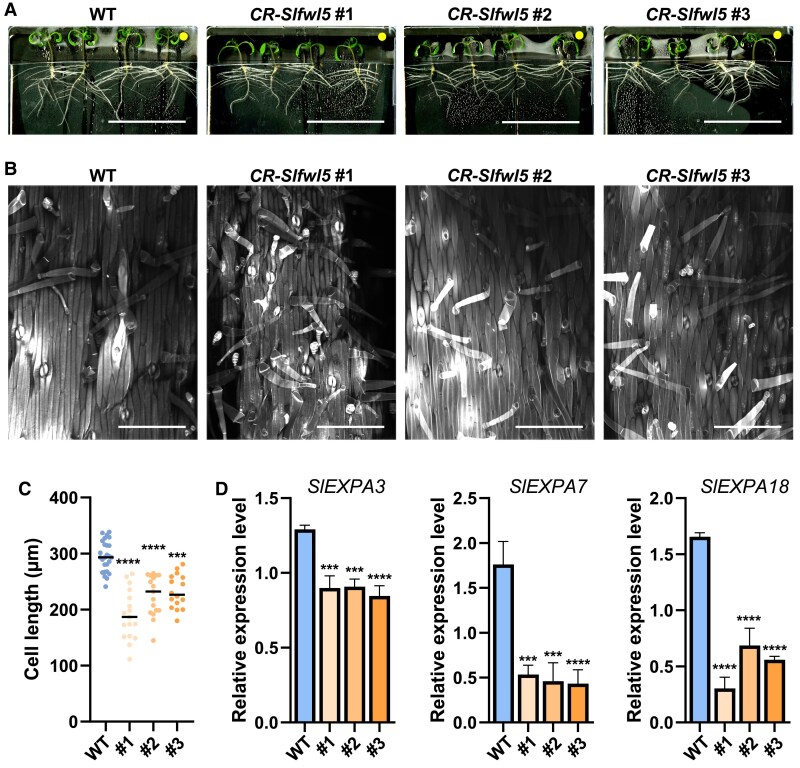 Fig. 6
Fig. 6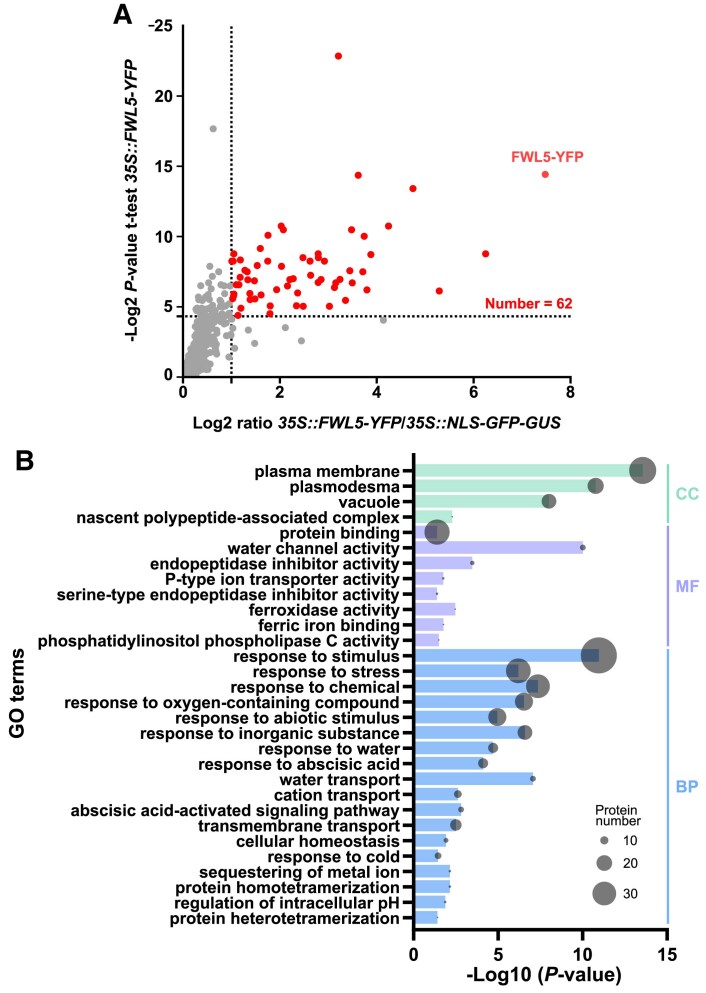 Fig. 7
Fig. 7| Gene ID | Proposed name | Name from | Name from |
|---|---|---|---|
| Solyc02g090730 |
|
|
|
| Solyc05g009620 |
|
|
|
| Solyc01g005470 |
|
|
|
| Solyc04g007900 |
|
|
|
| Solyc03g119660 |
|
|
|
| Solyc06g066590 |
|
|
|
| Solyc03g120600 |
|
|
|
| Solyc08g013910 |
|
|
|
| Solyc08g013920 |
|
|
|
| Solyc10g081410 |
|
|
|
| Solyc12g013570 |
|
|
|
| Solyc12g037950 |
|
|
|
| Solyc02g083540 |
|
| |
| Solyc10g084260 |
|
| |
| Solyc09g007490 |
|
| |
| Solyc10g018920 |
|
| |
| Solyc05g051690 |
|
| |
| Solyc03g093200 |
|
| |
| Solyc02g079390 |
|
| |
| Solyc06g048790 |
|
| |
| Solyc06g048810 |
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant nutrient uptake and metabolism · Plant Molecular Biology Research · Plant Micronutrient Interactions and Effects
Introduction
The FW2.2-LIKE/CELL NUMBER REGULATOR (FWL/CNR) gene family (Guo et al., 2010; Thibivilliers et al., 2020) was named in reference to its founding member, the FW2.2 gene (standing for Fruit Weight QTL on chromosome 2, number 2), which underlies the major quantitative trait locus (QTL) governing fruit size in tomato (Alpert et al., 1995; Frary et al., 2000). The FWL/CNR gene family encompasses hundreds of related sequences in the plant, animal, and fungal reigns (Guo et al., 2010), with a large diversity in protein size ranging from a hundred to several hundred amino acids. The FWL/CNR proteins are usually described as membrane-associated Pro- and Cys-rich proteins, which harbor, as a principal feature, the presence of a conserved structural domain named PLAC8 (Placenta specific 8), originally identified in a protein specifically expressed in the trophoblast giant cells and spongiotrophoblast layer of the placenta from mouse (Galaviz-Hernandez et al., 2003). The PLAC8 domain is composed of one or two hydrophobic segments, predicted to form two putative transmembrane (TM) α-helices, and characterized by the presence of Cys-rich motifs of the CLXXXXCPC or CCXXXXCPC type (Song et al., 2004; Beauchet et al., 2021).
Three main functions have been assigned to PLAC8 domain-containing FWL/CNR proteins in plants: the regulation of organ size, especially in fruits; cadmium resistance; and metal ion homeostasis (Beauchet et al., 2021). Indeed, the original FW2.2 protein negatively regulates cell divisions in the pre-anthesis ovary and young developing fruit, thereby accounting for its quantitative effects on final fruit size via the modulation of cell number (Nesbitt and Tanksley, 2001; Cong et al., 2002; Liu et al., 2003). Functional analyses of FWL/CNR proteins suggest that this role in the regulation of organ growth, primarily through their effects on cell division, is a highly conserved feature in both monocotyledon and dicotyledon plants (Dahan et al., 2010; Guo et al., 2010; Li and He, 2015; Qiao et al., 2017; Ruan et al., 2020). Interestingly, and related to this function, the PLAC8 protein has also been determined to be involved in organ development and cell division control during tumorigenesis in mammals (Cabreira-Cagliari et al., 2018; Mao et al., 2021). However, the precise molecular mechanism underlying the role of FWL/CNR proteins in regulating organ growth linked to cell division/cell cycle control remains largely unexplored.
Recently, Beauchet et al. (2024) made a major breakthrough towards the identification of the molecular and cellular mode of action of FW2.2 in tomato. In this study, FW2.2 was demonstrated to be a membrane-anchored protein that locates at plasmodesmata (PDs) and regulates cell-to-cell communication by modulating PD transport capacity during fruit development, via the regulation of callose deposition at PDs (Beauchet et al., 2024). Whether this function is conserved in FWL/CNR proteins from other plant species remains to be determined. Nevertheless, several clues seem to comfirm this functionality. Indeed, the closest ortholog of FW2.2 in Arabidopsis, namely AtPCR2, is highly enriched at PDs (Brault et al., 2019). The soybean GmFWL1 and GmFWL3 proteins are localized to membrane microdomains, and interact with structural proteins of the membrane microdomains, but also with proteins involved in the metabolic process of callose deposition, such as callose synthases (Qiao et al., 2017; Cervantes-Pérez et al., 2024), like FW2.2 in tomato fruit (Beauchet et al., 2024).
An updated phylogenetic analysis performed on 13 different plant genomes released in recent years allowed the identification of 134 members in the FWL/CNR family, with a total number of 12 FWL genes including FW2.2 in tomato (Thibivilliers et al., 2020). This list has been recently extended up to 21 members (Ran et al., 2023), providing a valuable resource for the characterization of FWL genes in tomato.
In the present study, we provide the first functional analysis of SlFWL genes in tomato. We aimed at investigating whether SlFWLs can play a direct role in the control of organ growth. First, we re-investigated the expression profiles of SlFWL genes in different organs of tomato plants and during fruit development, and analyzed their subcellular localization. Based on this combination of molecular and cellular approaches, we selected three SlFWL genes, namely SlFWL2, -4, and -5, and generated gain- and loss-of-function transgenic plants to explore their putative role as regulators of organ growth and development in tomato. This allowed us to shed light more specifically on the critical involvement of SlFWL5 in the regulation of cell expansion during leaf development and hypocotyl growth, further underscoring the importance of some FWL/CNR family members in growth regulation.
Materials and methods
Plant materials and growth conditions
Tomato [Solanum lycopersicum cv. Ailsa Craig (AC)] and tobacco (Nicotiana benthamiana) plants were grown in soil in a greenhouse under the following conditions: 16 h day/8 h night cycle, using a set of 100 W warm white LED projectors providing an irradiance of 100 μmol m^−2^ s^−1^ at the level of the canopy. The light spectrum was constituted by equivalent levels of blue irradiation (range 430–450 nm) and red irradiation (640–660 nm). For in vitro culture, tomato seeds were sterilized for 10 min under agitation in a solution of 3.2% bleach. Seeds were then washed three times with sterile water and dried under a laminar flow hood. Seeds were sown in Murashige and Skoog medium (1/4 MS) and transferred in a growth chamber under the following conditions: 16 h day/8 h night cycle, 22 °C/20 °C day/night, using white light (Osram L36 W/77 Fluora 1400 Im) providing 80–100 μmol m^−2^ s^−1^ intensity light at the stirring plate.
Phylogenetic analysis and tools for the prediction of the CNR/FWL structure and topology
The 21 SlFWL protein sequences from tomato were mined from Thibivilliers et al. (2020) and Ran et al. (2023), and aligned using MEGA11 Muscle alignment (Tamura et al., 2021). Values for 1000 bootstrap replicates are shown on each branch.
DeepTMHMM (https://dtu.biolib.com/DeepTMHMM) (Hallgren et al., 2022, Preprint) was used to predict the presence of a TM helix in SlFWL proteins. The three-dimensional structure of the full-length SlFWL2, -3, and -5 proteins was obtained using Colabfold (Mirdita et al., 2022) The PPM 3.0 Web Server (https://opm.phar.umich.edu/ppm_server3_cgopm) (Lomize et al., 2022) was used with default parameters and plasma membrane (PM; plants) type to predict the topology and insertion of SlFWL2, -3, and -5 in the PM.
Vector constructs and plant transformation
Vectors for the overexpression of SlFWL genes in plants were generated using the Gateway^®^ cloning system (Invitrogen, Carlsbad, CA, USA), following the manufacturer’s instruction. The FWL full-length coding sequence was amplified from cDNAs prepared from various tomato tissues (cv. AC) using PrimeSTAR MAX DNA polymerase (TAKARA BIO Inc., Kusatsu, Japan) and primers including the attB sites (Supplementary Table S1). The resulting PCR products were cloned into the corresponding Gateway vectors described in Supplementary Table S2. For CRISPR/Cas9 [clustered regularly interspaced palindromic repeats (CRISPR)/CRISPR-associated protein 9] mutagenesis, constructs were assembled using the Golden Gate cloning method according to a previous method (Beauchet et al., 2024). Transgenic plants were generated by Agrobacterium tumefaciens- (strain C58C1) mediated transformation using explants of tomato cotyledons as described (Swinnen et al., 2022).
RNA extraction and quantitative reverse transcription–PCR analysis
Total RNA was isolated from cotyledons, hypocotyls, shoot apical meristems, leaves, roots, flowers, and pericarp tissues from fruits harvested at different developmental stages (5, 10, and 15 d post-anthesis), using TRIzol reagent (Invitrogen) in combination with the RNeasy Plant Mini Kit (Qiagen) following the manufacturer’s instructions. RNase-free DNase (Qiagen) treatment was performed on each sample. Reverse transcription was performed using the iScript™ cDNA Synthesis Kit (Bio-Rad, Hercules, CA, USA). Real-time PCR was performed using Gotaq^®^ qPCR mastermix (Promega, Madison, WI, USA) and a CFX 96 real-time system (Bio-Rad). Quantitative PCR (qPCR) primers were designed with PerlPrimer software (Marshall, 2004) to overlap two exons in order to limit genomic DNA amplification (Supplementary Table S2) and amplify a 80–200 bp long amplicon, with a Tm of 60 °C. The transcript levels of the expressed genes were normalized to that of the housekeeping genes: SlTUBULIN (Solyc04g081490) in combination with SlNUDK (Solyc01g089970) for fruit samples, or with SlEIF4α (Solyc12g095990) for other tissue samples.
Subcellular localization analysis of SlFWL proteins
The subcellular localization of SlFWLs fused to yellow fluorescent protein (YFP) was observed using confocal imaging performed on a Zeiss LSM 880 confocal laser scanning microscope equipped with fast AiryScan, using a Zeiss Plan apochromat ×10 (NA 0.45) or a C PL APO ×63 (NA 1.4) oil-immersion objective.
The top three leaves of three independent N. benthamiana plants (6–8 weeks old) were agro-infiltrated with 35S::SlFWLs-YFP constructs and plasmolyzed using 0.4 M sorbitol for 15 min before observation. Co-infiltrations with SP-mTagBFP2-HDEL as a marker of endoplasmic reticulum (ER) and mScarlet-l as a marker of both cytosol and nucleus localization were performed to ascertain the localization pattern. Staining with FM4.64 at a final concentration of 4 µM was used as a control for PM localization (Bolte et al., 2004). For FM4.64 imaging, excitation was performed at 561 nm and fluorescence emission was collected at 630–690 nm. For YFP imaging, excitation was performed at 514 nm and fluorescence emission collected at 520–580 nm. For mTagBFP2 imaging, excitation was performed at 405 nm and fluorescence emission was collected at 440–490 nm. For mScarlet-I imaging, excitation was performed at 561 nm and fluorescence emission was collected at 570–640 nm.
The localization of SlFWL2, -4, and -5 at the PM and PDs was performed in leaves of tomato 35S::SlFWL2-YFP, 35S::SlFWL4-YFP, and 35S::SlFWL5-YFP transgenic plants. Similarly, leaf epidermal cells were plasmolyzed using 0.4 M sorbitol prior to observation. Staining with aniline blue (Biosupplies, Victoria, Australia) was performed by infiltration of a 0.0125% solution; excitation was performed at 405 nm and fluorescence emission collected at 420–480 .0nm. The PD index was determined by calculating the fluorescence intensity of SlFWL2–YFP, SlFWL4–YFP, and SlFWL5–YFP at PDs, and at the PM as described previously (Grison et al., 2019).
Phenotypic characterization
Plants were cultivated randomly side-by-side with wild-type (WT) plants. Flowers were vibrated every day to ensure optimal self-pollination. Seven flowers per inflorescence were maintained to ensure proper development of fruit per inflorescence. Fruits from 4–6 plants of each genotype of two biological replicates were used to determine fruit weight at the breaker stage of fruit development. Fruits were weighted and measured using a caliper. The number of measurements ranged from n=50 to n=200 depending on the number of fruits produced by the different transgenic plants. For root and hypocotyl length measurements, seedlings were grown on 1/4 MS in 245×245×20 mm Nunc tissue culture plates placed vertically. Pictures of seedlings were digitized using a scanner every day, and root and hypocotyl length were measured using ImageJ software. For leaf surface phenotyping, pictures of full-grown leaves were taken using a Nikon D5300 and analyzed by intensity threshold filtering in ImageJ. For the analysis of leaf epidermal cell size, leaflets were cleared with 70% ethanol for several days before staining with 0.1% (w/v) Calcofluor white M2R stain for 48 h. Leaflet fragments were observed with a Zeiss Axioimager epifluorescence microscope equipped with a ×20 (NA 0.5) lens using a DAPI-BP filter and a Cool Snap HQ2 CCD Camera. For the measurements of hypocotyl epidermal cell size, freshly harvested hypocotyls were fixed in 4% (w/v) paraformaldehyde in 1× phosphate-buffered saline (PBS) under vacuum for 30 min, rinsed three times with 1× PBS, and incubated in Clearsee solution for at least 2 weeks. Samples were then stained with 0.1% (w/v) Calcofluor white M2R for 48 h in fresh Clearsee. Central epidermal cells of each hypocotyls were imaged with a Zeiss LSM 880 confocal microscope using a Plan apochromat ×10 dry objective (NA 0.45); excitation was performed at 405 nm (0.2% laser power) and fluorescence emission was collected between 430 nm and 490 nm.
Immunolabeling of callose
The level of callose deposition was quantified in leaves from 4-week-old plants, by immunofluorescence labeling using a callose-specific antibody, as described in Beauchet et al. (2024). The signal intensity was subsequently quantified as a proxy of callose deposition at PDs, by integrating the gray value of all the pixels above the same threshold. A minimum of six measurements was performed at least on five sections from at least three different leaves from different plants, and the experiment was repeated twice.
Co-immunoprecipitation and mass spectrometry analysis
Total protein extracts from 100 mg of 35S::SlFWL5-YFP leaf tissue were prepared using the following buffer: 1× PBS, cOmplete Protease Inhibitor Cocktail tablets (Roche, Mannheim, Germany), and 1% Triton X-100. Samples were incubated in the extraction buffer at 4 °C for 30 min with agitation, and then centrifuged (16 000 g, 10 min, 4 °C). Prior to co-immunoprecipitation, western blotting was used to check the presence of the expressed tagged-SlFWL5–YFP protein in the supernatant. The supernatant containing the resuspended proteins was then used for immunoprecipitation assay using anti-Green Fluorescent Protein (GFP) microbeads provided in the μMACS Epitope Tag Protein Isolation Kit according to the manufacturer’s protocol (Miltenyi Biotec, Bergisch Gladbach, Germany). Protein concentration was determined by Bradford assay according to the manufacturer’s instructions [Pierce Coomassie Plus (Bradford) Assay kit; Thermo Scientific]. Approximately, 500 μg of soluble proteins were loaded for each co-immunoprecipitation assay. The LC-MS-MS experiment was carried out as reported previously (Beauchet et al., 2024).
Gene Ontology (GO) enrichment analysis was carried out on PLAZA 5.0 (Van Bel et al., 2022) using the Plaza workbench, with a significance threshold of 0.05 and without any data filter.
The MS proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE (Perez-Riverol et al., 2022) partner repository with the dataset identifier PXD061438.
Results
Phylogenetic classification and gene expression profiles of SlFWL genes
A recent genome-wide analysis of the SlFWL family identified a total of 20 FW2.2 homologous sequences in tomato (Ran et al., 2023). These authors named the different genes according to their location on the tomato chromosomes, not taking into account the anteriority of the SlFWL gene classification (although incomplete) proposed by Thibivilliers et al. (2020). To avoid additional confusion and to conform to the principle of anteriority, we proposed to keep the latter gene annotation and to assign ascending gene names to the nine new identified SlFWL sequences according to their phylogenetic relationship to FW2.2 (Table 1).
A phylogenic tree was thus generated using the full-length sequences of FW2.2 and the 20 homologous sequences from tomato (Fig. 1A). The SlFWLs fall into two distant clades: the first clade includes FW2.2 and the 11 sequences identified by Thibivilliers et al. (2020), with the addition of SlFWL12; the second clade encompasses SlFWL13–SlFWL20. Remarkably, SlFWLs from the first clade display a highly similar protein homology, with the exception of SlFWL4, SlFWL7, SlFWL8, and SlFWL12 which harbor a long N-terminal extension upstream of the conserved PLAC8 domain. SlFWLs from the second clade differ from the first one by a longer sequence, a greater distance between the two conserved motifs of the PLAC8 domain, and/or the presence of two or more predicted TM α-helices as revealed by the use of the TM topology prediction tool DeepTMHMM (Hallgren et al., 2022, Preprint) (Fig. 1A). It is noteworthy that SlFWL9 occupies an intermediate position between the two subgroups, with a relatively shorter sequence than that of FW2.2 and the presence of two TM domains.
Phylogenetic analysis and protein structure of SlFWLs. (A) Phylogenetic tree built using the maximum likelihood method was generated using MEGA11 (Tamura et al., 2021). Values for 1000 bootstrap replicates are shown on each branch. The schematic representation of the protein structure of the SlFWL was deduced from computation analysis of protein primary sequences. Transmembrane domains were predicted using DeepTMHMM (https://dtu.biolib.com/DeepTMHMM) (Hallgren et al., 2022, Preprint). (B) Amino acid sequence comparison of the PLAC8 domain between SlFWLs and FW2.2, using Muscle alignment (MEGA11).
The primary structure of the conserved PLAC8 domains in the 20 SlFWL proteins is described in Fig. 1B. The CLXXXXCPC motif is only conserved in FW2.2, SlFWL8, and SlFWL15, while SlFWL13 and SlFWL14 harbor a CLXXXXAPC sequence, and SlFWL2, -3, and -10 harbor the CCXXXXCPC sequence. The CLXXXXCPC or CCXXXXCPC motif diverges more or less in the other SlFWL proteins, especially for the two first amino acids of the sequence. The second motif at the C-terminal part, the so-called ‘QXXRELK’ motif, appears much more conserved among the 20 SlFWL proteins.
To provide an overview of the expression pattern in tomato, the levels of transcript accumulation for SlFWL genes were monitored by quantitative reverse transcription–PCR (RT–qPCR) in various organs and fruit developmental stages from the tomato cultivar AC (Supplementary Fig. S1). This analysis showed that a large diversity of expression profiles was observed among the tested SlFWL genes, not only in terms of organs, but also in terms of the level of transcript accumulation. For instance, SlFWL3, SlFWL14, and SlFWL15 exhibited much higher expression levels, ∼100-fold more, than all the other SlFWL genes. Most of the SlFWL genes are expressed to various levels throughout fruit development. SlFWL3, SlFWL9, SlFWL10, and SlFWL15 were predominantly expressed in fruits, with SlFWL3 and SlFWL15 having the highest expression. FW2.2, SlFWL1, and SlFWL4 were highly expressed in roots and to a lesser extent in fruit. SlFWL2 was predominantly expressed in cotyledons, and to a lesser extent in hypocotyls, leaves, and flowers, and slightly in roots. SlFWL5 showed high expression in both roots and cotyledons, and to a lower extent in leaves and hypocotyls. SlFWL7 and SlFWL8 were more specifically expressed in flowers. SlFWL13, SlFWL14, SlFWL16, SlFWL17, and SlFWL18 were broadly expressed across nearly all tissues.
Subcellular localization of SlFWL proteins
The subcellular localization of SlFWL proteins was first investigated in N. benthamiana leaves, using transient expression assays. Constructs aimed at expressing the SlFWL proteins fused to YFP at the C-terminal end, under the control of the cauliflower mosaic virus (CaMV) 35S promoter, were thus generated (referred to as 35S::SlFWLX-YFP). This type of construct was chosen based on our previous work, showing that the subcellular localization of FW2.2 was independent o the position of YFP at the C- or N-terminal end of the protein (Beauchet et al., 2024). The data obtained for 11 out of 20 SlFWLs are displayed in Fig. 2. The following SlFWLs were not included in this analysis for the following reasons: (i) we could not determine the subcellular localization of SlFWL3, due to a lack of reproducibility in the results; (ii) we were unable to generate the constructs fusing the coding sequence for SlFWL12, -13, -14, -16, and -17 to YFP for cloning technical reasons; (iii) SlFWL18, -19, and -20 were excluded from the study because of their too distant phylogenetic relationship with the other SlFWLs.
Subcellular localization of SlFWLs fused to YFP in N. benthamiana leaf epidermal cells. (A) ER localization revealed by the use of the ER-specific marker construct pUBI::mTagBFP2-HDEL. Scale bar=10 µm. (B) Cytosol and nuclear localization revealed by the use of the marker construct pUBI::mScarlet-I. Scale bar=10 µm. (C) PM localization revealed by the membrane-specific dye FM4-64. Scale bar=20 µm. Intensity plots delineated by the dashed lines are shown for each co-localization pattern.
For the tested SlFWLs, three distinct patterns of subcellular localization were observed. First, SlFWL1, -6, -9, -10, -11, and -15 were distributed within the cell in a reticular pattern, consistent with a localization at the ER. This ER localization was confirmed through co-infiltration with the ER marker SP-mTagBFP2-HDEL (kindly provided by Dr J. Dragdwidge, Ghent University, Belgium) and subsequent co-localization analysis (Fig. 2A).
Second, SlFWL7 and -8 were found to localize to both the cytosol and the nucleus. The use of the co-infiltrated mScarlet-l marker, a monomeric constitutive red fluorescent protein (Bindels et al., 2017), confirmed this localization (Fig. 2B).
Third, SlFWL2, SlFWL4, and SlFWL5 were targeted to the PM, as witnessed by staining with FM4.64, a membrane-specific dye (Bolte et al., 2004), following plasmolysis induced by a 0.4 M sorbitol treatment (Fig. 2C).
We next investigated the topology of these three SlFWLs at the PM using currently available prediction tools. First, the 3D structure of these PM-localized SlFWLs was predicted using ColabFold software (Mirdita et al., 2022) based on AlphaFold 2.0 (Jumper et al., 2021) and MMseqs2 (Steinegger and Söding, 2017). Second, the insertion in the PM was modelized using the PPM 3.0 Web Server (Lomize et al., 2022) (Supplementary Fig. S2). These predictions revealed that the 3D structures of SlFWL2, SlFWL4, and SlFWL5 present similarities to that from FW2.2 (Beauchet et al., 2024), such as the absence of any TM domain and their N- and C-terminal parts predicted to be folded on the same side of the protein. These predictions suggest that they do not cross the PM, but are most probably anchored in the PM.
In planta subcellular localization of SlFWL2, SlFWL4, and SlFWL5
Owing to their localization at the PM, we performed an in planta analysis of the subcellular localization of SlFWL2, SlFWL4, and SlFWL5, aimed at investigating whether these three SlFWLs are also enriched at PDs in tomato, like FW2.2 (Beauchet et al., 2024). For this purpose, we generated stable transgenic lines expressing SlFWL2, SlFWL4, and SlFWL5 fused to YFP at the C-terminal end, under the control of the 35S promoter in the tomato cultivar AC (lines referred to as 35S::SlFWL2-YFP, 35S::SlFWL4-YFP, and 35S::SlFWL5-YFP plants).
The localization of SlFWL2–YFP, SlFWL4–YFP, and SlFWL5–YFP at the PM of tomato leaf cells was confirmed (Fig. 3A). In addition, SlFWL5–YFP was found to localize at the PM according to the pattern of punctate spots at the cell periphery, suggesting that SlFWL5–YFP was enriched at nanodomains. Staining with aniline blue to reveal callose deposition as a marker of PDs revealed that only SlFWL5–YFP co-localized with aniline blue, as shown by the overlapping signal intensity plots (Fig. 3A), thus indicating a localization at PDs. Next, we determined the PD enrichment ratio, named the ‘PD index’, which corresponds to the fluorescence intensity of SlFWL–YFP fusions at PDs versus that at the cell periphery, to quantify the enrichment of SlFWL–YFP fusions at PDs, as previously described (Beauchet et al., 2024) (Fig. 3B). While the PD index for SlFWL2–YFP and SlFWL4–YFP was equal to 1, a high PD index from 1.5 to 1.7 was measured in leaf cells of 35S::SlFWL5-YFP plants, thus demonstrating that only SlFWL5 out of the three SlFWLs tested was enriched at PDs.
*SlFWL5 is enriched at PDs. (A) Confocal microscope observations of SlFWL2, SlFWL4, and SlFWL5 localization in tomato leaf cells from 35S::SlFWL2-YFP, 35S::SlFWL4-YFP, and 35S::SlFWL5-YFP plants. Scale bar=10 μm. Intensity plots delineated by the dashed lines are shown for each co-localization pattern. AB, aniline blue. (B) PD index for SlFWL2, SlFWL4, and SlFWL5 localization in leaf cells from 35S::SlFWL2-YFP, 35S::SlFWL4-YFP, and 35S::SlFWL5-YFP plants. n>20 regions of interest from five images. Statistical analysis: Kruskal–Wallis test with post-hoc Dunn multiple comparison test. **P<0.01.
In planta functional analysis of SlFWL2, SlFWL4, and SlFWL5
To analyze the function of SlFWL2, SlFWL4, and SlFWL5 in vegetative and reproductive development, gain- and loss-of-function plants were generated in the tomato cultivar AC. SlFWL2, SlFWL4, and SlFWL5 were overexpressed constitutively and ectopically, using the 35S promoter (gain-of-function plants referred to as 35S::SlFWL2, -4, and -5 respectively). For each gene, three T_2_ lines were selected with medium to very high levels of SlFWL2, -4, and -5 overexpression in leaves, ranging from 3-fold more to 150-fold more (Supplementary Fig. S3). Alternatively, SlFWL2, SlFWL4, and SlFWL5 were knocked out using CRISPR/Cas9 technology. Two single guide RNAs (sgRNAs) per gene were designed close to the start codon, in order to create a frameshift or an early stop codon that would result in a dysfunctional protein (Supplementary Fig. S4). For each gene, we selected three independent T_0_ transgenic lines displaying homozygous mutations, either deletions or insertions, for subsequent phenotyping of vegetative and reproductive development. These loss-of-function plants are referred to as CR-Slfwl2, -4, and -5 hereafter.
The phenotypic analysis of the different transgenic plants is shown in Fig. 4. The overexpression of SlFWL2, SlFWL4, and SlFWL5 did not induce any significant effects on either reproductive or vegetative development. Only one line each out of three for the 35S::SlFWL2 and 35S::SlFWL4 constructs, namely 35S::SlFWL2#2 and 35S::SlFWL4#2, produced smaller fruits (−20% when compared with the WT) (Fig. 4A). No significant impacts could be detected either on root length (Fig. 4B) or on leaf surface (Fig. 4D), and the sole 35S::SlFWL2#2 line was negatively affected for hypocotyl length, with a reduction of an average of 20% when compared with the WT (Fig. 4C).
*Phenotypic analysis of SlFWL2, SlFWL4, and SlFWL5 gain-of-function (referred to as 35S::SlFWL2, 35S::SlFWL4, and 35S::SlFWL5) and loss-of-function (referred to as CR-Slfwl2, CR-Slfwl4, and CR-Slfwl5) plants compared with WT plants. (A) Fruit weight (at red ripe stage); n>40 fruits from four plants per line. (B) Root length (at 17 DAG); n>10 roots per line. (C) Hypocotyl length (at 17 DAG); n>10 hypocotyls per line. (D) Mature leaf surface; n>10 leaves from four plants per line. Statistical analysis: Kruskal–Wallis test with post-hoc Dunn multiple comparison test. *P<0.05; **P<0.01; ***P<0.001; ***P<0.0001.
The most noticeable effects were obtained for CRISPR knockout mutants. Two out of three SlFWL2 mutant lines (namely CR-Slfwl2#1 and CR-Slfwl2#2), and the sole CR-Slfwl5#3 line displayed a statistically significant reduction in fruit weight (–17%, –42%, and –24%, respectively, on average when compared with the WT) (Fig. 4A). All three CR-Slfwl4 knockouts lines were affected for root length, with an increase from 15% to 23% on average, when compared with WT plants, while CR-Slfwl2 and -5 lines were not affected (Fig. 4B; Supplementary Fig. S5). In plants harboring non-functional SlFWL5 alleles, the length of the hypocotyl in all three CR-Slfwl5 lines was reduced from 22% to 38% on average, when compared with the WT (Fig. 4C). More remarkably, all three CR-Slfwl5 lines displayed a marked reduction in leaf surface (from 32% to 47% o average) (Fig. 4D), which suggests that the suppression of SlFWL5 alters the overall vegetative development of aerial organs, as root development was not affected.
These results suggested that vegetative growth and development in tomato are regulated by SlFWL4 in roots and, more remarkably, by SlFWL5 in leaves and hypocotyls.
SlFWL5 participates in the control of leaf and hypocotyl growth via cell expansion
We then proceeded to a fine phenotyping of the leaf morphology in all three CR-Slfwl5 lines since the overall leaf surface was reduced compared with the WT. The fully expanded compound leaf of the cultivated tomato cultivar AC is made up of a terminal leaflet (TL) and several lateral leaflets attached to the rachis (Fig. 5A). According to the sequential order and position in the rachis, the lateral leaflets can be further categorized into primary (PL), secondary (SL), and intercalary (IL) leaflets.
*Phenotypic analysis of mature leaves from CR-Slfwl5 plants compared with those of the WT. (A) Schematic representation of a mature leaf from the WT AC background defining the terminal leaflet (TL), the primary (PL), secondary (SL), and intercalary leaflets (IL) in their sequential order. (B) Silhouettes of the sixth leaf of 8-week-old CR-Slfwl5 plants. (C) Analysis of leaf composition in CR-Slfwl5 plants; numbers indicate the percentage of leaves presenting the corresponding leaflet. The color code refers to (A). (D) Determination of the mean TL, PL, and IL surface in WT and CR-Slfwl5 plants. n>10 leaves from four plants per genotype (each dot represents one leaflet surface measurement). Statistical analysis: Kruskal–Wallis test with post-hoc Dunn multiple comparison test. ***P<0.001; ****P<0.0001. (E) Leaf epidermal cells of the WT and the three CR-Slfwl5 plants. Scale bars=100 µm. (F) Cell size determination in the leaf epidermis of the WT and the three CR-Slfwl5 plants. Statistical analysis: Kruskal–Wallis test with post-hoc Dunn multiple comparison test. *P<0.05; ***P<0.001; ***P<0.0001. n>12 image measurements from leaves 5 and 6 of five independent plants for each line. (G) Expression of SlEXPA7, SlEXPA10, and SlEXPA12 in leaves 3–4 of 6-week-old WT and CR-Slfwl5 plants.
When compared with WT leaves, the leaf morphology of all three CR-Slfwl5 lines was affected, as the production of PL-3, IL-1, IL-3, and SLs was greatly reduced (Fig. 5B). The CR-Slfwl5#1 line displayed systematically the most altered leaf composition with the lowest number of leaflets. Not only was this reduction in the leaflet number responsible for the overall reduction in leaf surface observed in all three CR-Slfwl5 lines compared with the WT (Fig. 5C), but the reduction in the surface of TL, PL, and IL also contributed to the phenotype (Fig. 5D). This reduction in leaflet surface and overall leaf surface in the three CR-Slfwl5 lines was due to a reduction in cell size ranging from 17% to 46% on average (Fig. 5E, F). The expression levels of EXPANSIN A (EXPA) genes used as cell expansion markers (Cosgrove, 2000) were further examined in leaves of 3- of 6-week-old plants of CR-Slfwl5 lines by RT–qPCR. Among the different EXPA genes expressed in tomato leaves (Lu et al., 2016) (Supplementary Fig. S6A), the expression levels of three genes, namely SlEXPA7, -A10, and *-*A12, were significantly lower in the three mutant lines compared with the WT (Fig. 5G), thus accounting for the alteration of the cell expansion process when the function of SlFWL5 is suppressed.
Since hypocotyl growth was also affected in the CR-Slfwl5 lines (Figs 4C, 6A), we next investigated whether the phenotype originated from a defect in cell growth. Clearly, the size of epidermal cells in the hypocotyl of seedlings at 10 days after germination (DAG) was significantly reduced in all CR-Slfwl5 lines compared with the WT (Fig. 6B), from 21% to 36% on average (Fig. 6C). Here too, CR-Slfwl5#1 was the most affected line. Similarly to what was observed in leaves, only three EXPA genes among those expressed in stems (Lu et al., 2016) (Supplementary Fig. S6B), namely SlEXPA3, -A7, and -A18, were significantly reduced in all CR-Slfwl5 lines (Fig. 6D). The transcript levels of SlEXPA7 and SlEXPA18 displayed the strongest down-regulation.
*Phenotypic analysis of hypocotyls from CR-Slfwl5 plants compared with those of the WT. (A) Illustration of hypocotyl length in 2-week-old WT and CR-Slfwl5 loss-of-function tomato plants. Scale bars=10 cm. (B) Hypocotyl epidermal cells in 10 DAG WT and CR-Slfwl5 seedlings. Scale bars=200 µm. (C) Cell size determination in the hypocotyl epidermis of WT and CR-Slfwl5 plants. Statistical analysis: Kruskal–Wallis test with post-hoc Dunn multiple comparison test. ***P<0.001; ***P<0.0001. n>12 images measurement from the central part of the hypocotyl of five independent plants for each line. (D) Expression of SlEXPA3, SlEXPA7, and SlEXPA18 in hypocotyls of 10 DAG WT and CR-Slfwl5 seedlings.
SlFWL5 pull-down reveals plasma membrane- and plasmodesmata-related proteins
The elucidation of the functional role of SlFWL5 in leaves and at PDs relied on a deeper characterization at the biochemical level. For this purpose, we performed an immunoprecipitation followed by tandem MS (IP-MS/MS) on 35S::SlFWL5-YFP leaves to identify interacting protein partners of SlFWL5. This proteomics approach was required to determine the most appropriate leaf developmental stage to harvest, so that the natural interacting protein partners are present in the protein extracts. Hence, the expression level of SlFWL5 was monitored in the different leaves of 4-week-old plants (Supplementary Fig. S7). Four-week-old plants produced up to eight leaves: from the newly formed leaves, such as leaves 8 and 7 emerging from the meristem, to the oldest fully developed leaves, such as. leaves 2 and 1. The highest expression of SlFWL5 was found in leaves 6, 5, and 4, when leaf growth is highly sustained by cell expansion. Protein extracts were then prepared from these three different stages and pooled prior to immunoprecipitation.
The IP-MS/MS experiment resulted in the identification of 62 proteins that co-immunoprecipitated with SlFWL5, which were significantly enriched in the 35S::SlFWL5-YFP sample when compared with 35S::NLS-GFP-GUS used as a control (Fig. 7A; Supplementary Dataset S1). A GO term enrichment analysis for the 62 identified proteins was performed (Fig. 7B), showing that they fall into the three types of GO domains: cellular components (CC), molecular functions (MF), and biological processes (BP). Notably, the CC terms ‘plasma membrane’ and ‘plasmodesmata’ are highly represented. Interestingly, 19 proteins out the 62 identified candidate proteins were found to belong to the FW2.2 co-immunoprecipitation proteome (Beauchet et al., 2024), and three of them also crossed with the refined PD proteome from Arabidopsis established by Brault et al. (2019), among which was Callose Synthase 10a (SlCalS10a; Solyc03g111570) (Supplementary Table S3). It is also noteworthy that among the identified candidate proteins was a subunit of the Cellulose Synthase complex (SlCesA1; Solyc08g061100), which may relate to cell wall synthesis and cell expansion.
SlFWL5 co-immunoprecipitates with several PM- and PD-localized proteins. (A) Dot plots showing enriched proteins in 35S::SlFWL5-YFP IP-MS/MS experiments in young leaves. Red dots indicate significantly enriched proteins (based on a Student’s t-test with Benjamini–Hochberg correction P<0.05 and an enrichment ratio >2). (B) Gene Ontology (GO) enrichment analysis of the identified proteins following SlFWL5 co-immunoprecipitation.
Discussion
PLAC8 domain-containing proteins in plants belong to the FW2.2-LIKE/CELL NUMBER REGULATOR (FWL/CNR) protein family. Our understanding of the function of FWL/CNR proteins still remains poorly developed, but three distinctive functions have been identified so far (Beauchet et al., 2021). These functions relate to (i) calcium uptake and signaling, (ii) metal ion transport and homeostasis, and (iii) organ size determination via the regulation of cell number. These three functions are not exclusive, as they may directly or indirectly influence each other, and examples of mutual actions on plant growth and metal ion transport (Song et al., 2015; Xiong et al., 2018; Li et al., 2019; Gao et al., 2020) or plant growth and calcium signaling (Rosa et al., 2017) have been reported. It is noteworthy that a function in improving resistance to a plant pathogen, namely Xanthomonas oryzae, has been described alongside metal ion transport (Li et al., 2019).
So far, functional studies of FWL/CNR genes are still scarce in the literature. The first reports were dedicated to the functional characterization of FW2.2 orthologs in important cereal crops such as maize and rice. In maize, the overexpression of ZmCNR1 resulted in a reduction in overall plant size and organ size (namely tassel, ear, and leaf size), thus affecting the plant biomass (Guo et al., 2010). In rice, T-DNA insertion mutants of OsFWL3 and OsFWL5 displayed an increase in the grain weight and/or plant height (Xu et al., 2013; Song et al., 2015). Similarly, OsFWL1/OsCNR1 (Ruan et al., 2020) and OsFWL4 (Gao et al., 2020) were shown to act as negative regulators of rice grain width and weight, and tiller number and plant yield, respectively. In addition, the RNAi silencing of GmFWL1 expression and the deletion of the conserved PLAC8 domain of GmFWL3 using CRISPR/Cas9 in soybean resulted in a significant reduction in nodule number in response to rhizobial infection, pointing to a critical role for GmFWL1 and GmFWL3 in nodule organogenesis (Libault et al., 2010; Cervantes-Pérez et al., 2024). In all cases, these studies confirmed a role for certain FWL/CNR genes in negatively regulating cell number, thus influencing the organ size and/or development.
Due to the scarcity of data, in the present report we have undertaken a functional analysis of some FWL/CNRs in tomato, so as to enrich our knowledge and to decipher their putative role in plant development.
The FWL/CNR protein family presents a high diversity in protein characteristics, expression pattern, and subcellular localization in tomato
The FWL/CNR gene family in tomato comprises 21 members, showing a large diversity in protein length, ranging from 98 (SlFWL1) to 505 (SlFWL19) amino acids. FWL/CNR proteins share, as a common feature, the presence of the PLAC8 domain made of two conserved motifs: the more or less divergent Cys-rich motif 1 of type CLXXXXCPC or CCXXXXCPC, and the more conserved QXXRELK motif 2 (Fig. 1). Since the CLXXXXCPC motif is present in FW2.2, ZmCNR1, OsFWL1/OsCNR1, and PfCNR1, it would be tempting to address this motif as a signature for FWL/CNR proteins involved in the regulation of organ growth. However, OsFWL4 and OsFWL5, which regulate rice grain weight, harbor the CCXXXXCPC motif. This latter motif has been described as important for conferring the cadmium resistance function (Song et al., 2004). Interestingly, OsFWL4 and OsFWL5 both also enhanced cadmium resistance in yeast cells, and OsFWL4 acts as a transporter ensuring the translocation of cadmium from roots to shoots in rice (Song et al., 2015; Xiong et al., 2018). Therefore, the nature of the PLAC8 Cys-rich motif does not indicate any function with certainty, and this is all the more so since different divergent motifs exist in the tomato FWL/CNR family members. This was particularly obvious for the three SlFWL proteins we characterized functionally in more depth, namely SlFWL2, SlFWL4 and SlFWL5, which harbor a CCXXXXCPC, ALXXXXFPC and AVXXXXLPC motif, respectively (Fig. 1).
Despite clear levels of overexpression (Supplementary Fig. S3), we did not observe any phenotypes in the 35S::SlFWL2, 35S::SlFWL4, and 35S::SlFWL5 lines (Fig. 4). This suggests that constitutive overexpression probably fails to reproduce the native spatial and temporal regulation of these FWL/CNR genes, or may trigger compensatory mechanisms. On the contrary, the inactivation of only SlFWL4 (CR-Slfwl4 plants) and SlFWL5 (CR-Slfwl5 plants) generated marked phenotypes during vegetative development: CR-Slfwl4 lines produced longer roots (Fig. 4B) and CR-Slfwl5 lines showed reduced hypocotyl length (Fig. 4C), and a spectacular reduction in leaf size (Figs 4D, 5B). Apart from SlFWL4 which seemed to be preferentially expressed in roots, these phenotypes could not be related to expression of any specific gene: SlFWL2 was found more expressed in cotyledons, and SlFWL5 was expressed in all organs, without any preferential expression in leaves (Supplementary Fig. S1).
In the present study, we investigated the subcellular localization of 11 out of 20 SlFWLs (Fig. 2). A diversity in subcellular localization was observed, as SlFWLs are located according to three distinct patterns: in both the cytosol and the nucleus for SlFWL7 and SlFWL8; at the ER for SlFWL1, -6, -9; -10, -11, and -15; and at the PM for SlFWL2, -4, and -5. Interestingly, a punctate localization in the PM was observed for SlFWL2 and -4, which may be consistent with a membrane microdomain localization, as demonstrated for GmFWL1 and GmFWL3 in soybean nodules (Qiao et al., 2017; Cervantes-Pérez et al., 2024). Therefore, this diversity in subcellular localization might be indicative of a diversity in protein function.
With the exception of SlFWL7 and SlFWL8, all the tested SlFWLs were thus found targeted to membranous compartments, most probably in connection with the presence of the hydrophobic PLAC8 domain or TM domains in the case of SlFWL15. From the original structural analysis of AtPCR1 (Song et al., 2004), it has long been accepted, and widely and systematically reported in the literature, that the PLAC8 domain in plant FWL/CNRs is composed of two hydrophobic segments (including the CLXXXXCPC or CCXXXXCPC motif), predicted to form TM domains (for a review, see Beauchet et al., 2021). However, the use of currently available tools for TM topology prediction revealed that SlFWL2, SlFWL4, and SlFWL5 may not cross the PM in the absence of TM domains, but are likely to be anchored in the outer leaflet of the PM via their PLAC8 hydrophobic domain (Supplementary Fig. S2), Such a topology would thus be very similar to that demonstrated experimentally for FW2.2 (Beauchet et al., 2024).
SlFWL5 regulates leaf size and hypocotyl growth in tomato via cell expansion
Unexpectedly, we could demonstrate that the leaf and hypocotyl growth phenotypes in CR-Slfwl5 lines are not due to impaired cell divisions, as would be expected for a member of the CELL NUMBER REGULATOR protein family, but from an impairment in cell expansion, as cell size in all three lines was greatly reduced (Figs 5E, F, 6B, C). Interestingly, this reduction in cell size was accompanied by a significant down-regulation of some leaf- and hypocotyl-expressed EXPA genes (Figs 5G, 6D). The contribution of the EXPANSIN gene expression in modulating leaf growth (Cho and Cosgrove, 2000; Goh et al., 2012) and hypocotyl growth (Caderas et al., 2000) is a long-standing observation. For leaf growth in particular, it is part of the complex molecular and hormonal network regulating leaf size (Gonzalez et al., 2012).
Not all EXPA genes responded in the same way to SlFWL5 loss of function. Indeed, among the genes specifically expressed in these two organs (Lu et al., 2016), only SlEXPA7, -10, and -12, and SlEXPA3, -7, and -18, were down-regulated in leaf and hypocotyl, respectively (Figs 5G, 6D; Supplementary Fig. S6). The origin of this discrepancy is not known. It can be related to the specific function of EXPANSINS or the timing and localization of EXPA gene expression. For instance, SlEXPA2 is the major gene expressed in stems and hypocotyls, but its expression is maximal at the top of the hypocotyl, corresponding to the zone of rapid elongation, and very low in the middle part of the hypocotyl where cells have already expanded (Caderas et al., 2000). Whether this discrepancy may relate to any modification in the hormonal regulation of EXPA genes following SlFWL5 loss of function is an intriguing subject for investigation. It was shown that SlEXPA3, -7, -12, and -18 were all positively regulated by auxin, while SlEXPA10 was positively regulated by gibberellin (GA) (Lu et al., 2016), two major hormones involved in the stimulation of cell elongation during plant growth.
The inactivation of SlFWL5 not only affected leaf growth, but it also altered leaf morphology into simpler leaves, displaying a reduced number of the third PL, and of SLs,and ILs (Fig. 5C, D). This phenotype is fully relevant to hormonal control of compound leaf morphogenesis and differentiation in tomato (Bar and Ori, 2015). Indeed, GA negatively regulates leaf complexity in tomato, as increased GA levels or GA signaling accelerate leaf maturation and induce simpler leaves than in the WT (Jasinski et al., 2008; Yanai et al., 2011). In addition, GA and cytokinin (CK) present a mutual antagonistic interaction during tomato leaf development (Fleishon et al., 2011). How to relate the molecular origin of the leaf phenotype in CR-Slfwl5 plants and phytohormone signaling remains to be established, but the following step in the functional characterization of SlFWL5 may provide new lines of research.
SlFWL5 is enriched at plasmodesmata, within a protein complex composed of plasmodesmata- and plasma membrane-specific proteins
We have shown that SlFWL5 is enriched at PDs (Fig. 3), where it probably participates in a protein complex composed of PM- and PD-specific proteins (Fig. 7). Interestingly, a significant number of candidate proteins that co-immunoprecipitated with SlFWL5 were found enriched at PDs. This finding suggests that SlFWL5 may have a function in cell-to-cell communication mechanisms for the control of leaf and hypocotyl growth. SlFWL5 co-immunoprecipitates with CalS10a (Solyc03g111370), which contributes to callose homeostasis at the cell plate and at PDs, thereby regulating cytokinesis and the symplastic molecular exchanges between neighboring cells via the permeability of PDs (Saatian et al., 2018). CalS10 is required for normal plant development, as silencing of CalS10 results in retarded growth: plants display dwarfism, with a smaller stem and smaller leaves (Töller et al., 2008), a phenotype originating from either impaired cell division or cell expansion. CR-Slfwl5 loss-of-function plants displayed a similar phenotype, namely a reduced hypocotyl length and leaf surface, due to impaired cell expansion. However, our data indicate that SlFWL5 does not directly control callose deposition dynamics via regulating the CalS activity, thereby modifying the PD aperture. Indeed, according to our immunolocalization assays, the absence of a functional SlFWL5 was not associated with measurable differences in callose deposition in leaves (Supplementary Fig. S8). Therefore, the mode of action of SlFWL5 at PDs appears quite different from that of FW2.2 (Beauchet et al., 2024), and the putative interaction of SlFWL5 with CalS10 may only reflect its protein environment at PDs.
Interestingly, SlCesA1, a subunit of the Cellulose Synthase complex, was identified among the candidate proteins that co-immunoprecipitate with SlFWL5. Cellulose is the most abundant β-glucan polysaccharide of the plant cell wall, which self-assembles as microfibrils to strengthen the cell wall and contribute to the direction of cell growth during the cell expansion process (Schneider et al., 2016). Both cellulose and callose are synthesized at the PM by the large respective synthase complexes, cellulose synthases and callose synthases, and the formation of cellulose–callose networks has been demonstrated, especially as a layered cellulose–callose architecture in cell walls of epidermal leaf cells (Falter et al., 2015). Whether the SlFWL5 function in controlling leaf growth requires a regulatory role on cellulose synthases during cell expansion remains an exciting avenue for future investigation.
Unfortunately, we were unable to provide any direct experimental evidence for the physical interaction between SlFWL5 and CalS10 or SlCesA1, identified as putative interactors from our IP-MS/MS proteomics assays (Fig. 7; Supplementary Table S4). First, the very large size and multi-TM domain nature of CalS and CesA make them extremely challenging to clone and express in heterologous system. Second, both CalS and CesA proteins possess N- and C-terminal ends that face the cytosol (Verma and Hong, 2001; Kumar and Turner, 2015), whereas SlFWL5 displays both N- and C-terminal regions in the apoplast (Supplementary Fig. S2), like its counterpart FW2.2 (Beauchet et al., 2024). Therefore, this differential orientation precludes proper reconstitution of split-ubiquitin fragments or fluorescent tags, even if fused, resulting in false negatives or structural artifacts when using classical interaction methods, such as yeast two-hybrid or split-ubiquitin. Moreover, PDs and plant-specific membrane microdomains have unique lipid composition and regulatory environments that do not exist in yeast. Consequently, assays in yeast may not reflect biologically relevant interactions in plants. While in planta approaches (such as FRET-FLIM/BiFC) might appear suitable, the opposite orientation of N- and C-termini and thus this difference in protein topology prevent reliable reconstitution, and compromise the demonstration of a direct structural interaction between SlFWL5 and CalS10 or SlCesA1.
As key elements of the cell-to-cell communication machinery, PDs are implicated in processes guaranteeing the collaborative function of the cells, and developmental and patterning events (Petit et al., 2020), enabling the intercellular trafficking of different mobile signaling molecules, such as small RNAs, metabolites, hormones, and transcription factors (Wu and Gallagher, 2012; Brunkard et al., 2013). Being localized at PDs, SlFWL5 may have a regulatory role in PD activity. Hence, we can hypothesize that SlFWL5 function is to regulate the cell-to-cell movement of molecules/signals promoting leaf cell expansion and leaflet initiation. Obviously, the identification of such signaling molecules belonging to transcriptional networks involved in pathways affecting cell growth (Davière and Achard, 2013) and leaf growth (Gonzalez et al., 2012; Bar and Ori, 2014) would represent a giant leap in understanding the SlFWL5 functional role at PDs.
In conclusion, we are far from deciphering the functional complexity of the FWL/CNR protein family during plant and organ development. Unexpectedly, we have been able to demonstrate that SlFWL5 contributes to the regulation of cell expansion during organ growth, which has never been reported before for a member of the CELL NUMBER REGULATOR protein family. Therefore, this study may lead us to revise our perception of the FWL/CNR function in regulating organ size. Whatever the mechanism involved, cell division or cell expansion control, what may really matter is the potential function in cell-to-cell communication so as to regulate organ growth.
Supplementary Material
eraf444_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alpert KB, Grandillo S, Tanksley SD. 1995. Fw 2.2: a major QTL controlling fruit weight is common to both red- and green-fruited tomato species. Theoretical and Applied Genetics 91, 994–1000.24169988 10.1007/BF 00223911 · doi ↗ · pubmed ↗
- 2Bar M, Ori N. 2014. Leaf development and morphogenesis. Development 141, 4219–4230.25371359 10.1242/dev.106195 · doi ↗ · pubmed ↗
- 3Beauchet A, Bollier N, Grison M, Rofidal V, Gévaudant F, Bayer E, Gonzalez N, Chevalier C. 2024. The CELL NUMBER REGULATOR FW 2.2 protein regulates cell-to-cell communication in tomato by modulating callose deposition at plasmodesmata. Plant Physiology 196, 883–901.38588030 10.1093/plphys/kiae 198PMC 11444278 · doi ↗ · pubmed ↗
- 4Beauchet A, Gévaudant F, Gonzalez N, Chevalier C. 2021. In search of the still unknown function of FW 2.2/CELL NUMBER REGULATOR, a major regulator of fruit size in tomato. Journal of Experimental Botany 72, 5300–5311.33974684 10.1093/jxb/erab 207 · doi ↗ · pubmed ↗
- 5Bindels DS, Haarbosch L, van Weeren L, et al 2017. Mscarlet: a bright monomeric red fluorescent protein for cellular imaging. Nature Methods 14, 53–56.27869816 10.1038/nmeth.4074 · doi ↗ · pubmed ↗
- 6Bolte S, Talbot C, Boutte Y, Catrice O, Read ND, Satiat-Jeunemaitre B. 2004. FM-dyes as experimental probes for dissecting vesicle trafficking in living plant cells. Journal of Microscopy 214, 159–173.15102063 10.1111/j.0022-2720.2004.01348.x · doi ↗ · pubmed ↗
- 7Brault ML, Petit J, Immel F, et al 2019. Multiple C 2 domains and transmembrane region proteins (MCT Ps) tether membranes at plasmodesmata. EMBO Reports 20, e 47182.10.15252/embr.201847182 PMC 668013231286648 · doi ↗ · pubmed ↗
- 8Brunkard JO, Runkel AM, Zambryski AM, C P. 2013. Plasmodesmata dynamics are coordinated by intracellular signaling pathways. Current Opinion in Plant Biology 16, 614–620.23978390 10.1016/j.pbi.2013.07.007PMC 3828052 · doi ↗ · pubmed ↗