CCT4 promotes tunneling nanotube formation
Miyu Enomoto, Akiko Asada, Taro Saito, Kanae Ando

TL;DR
This study shows that the protein CCT4 helps form tunneling nanotubes, which are structures that connect cells and allow for communication and transport.
Contribution
The study identifies a new role for monomeric CCT4 in promoting the formation of microtubule-containing tunneling nanotubes.
Findings
Monomeric CCT4 induces the formation of nanotubes containing actin fibers and mitochondria.
CCT4 enhances microtubule dynamics and increases tubulin-containing tunneling nanotubes.
CCT4 proteins are transported intercellularly via the nanotubes they form.
Abstract
Tunneling nanotubes (TNTs) are membranous tunnel‐like structures that mediate cell‐to‐cell communication, although the molecular mechanisms of TNT formation are not fully understood. T‐complex protein 1 subunit delta (CCT4) serves as a component of the chaperonin‐containing TCP1 complex (TRiC) and also functions as a monomer. Here, we report that monomeric CCT4 promotes TNT formation in mammalian cultured cells. The expression of GFP–CCT4 proteins, which are not incorporated into the chaperonin oligomer, induces the formation of nanotubes containing actin fibers and mitochondria. CCT4 proteins are transported intercellularly via these nanotubes. The expression of monomeric CCT4 enhances microtubule dynamics and increases tubulin‐containing TNTs. Our results suggest a newly identified function of monomeric CCT4 in TNT formation. Impact statementTunneling nanotubes (TNTs) play critical…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
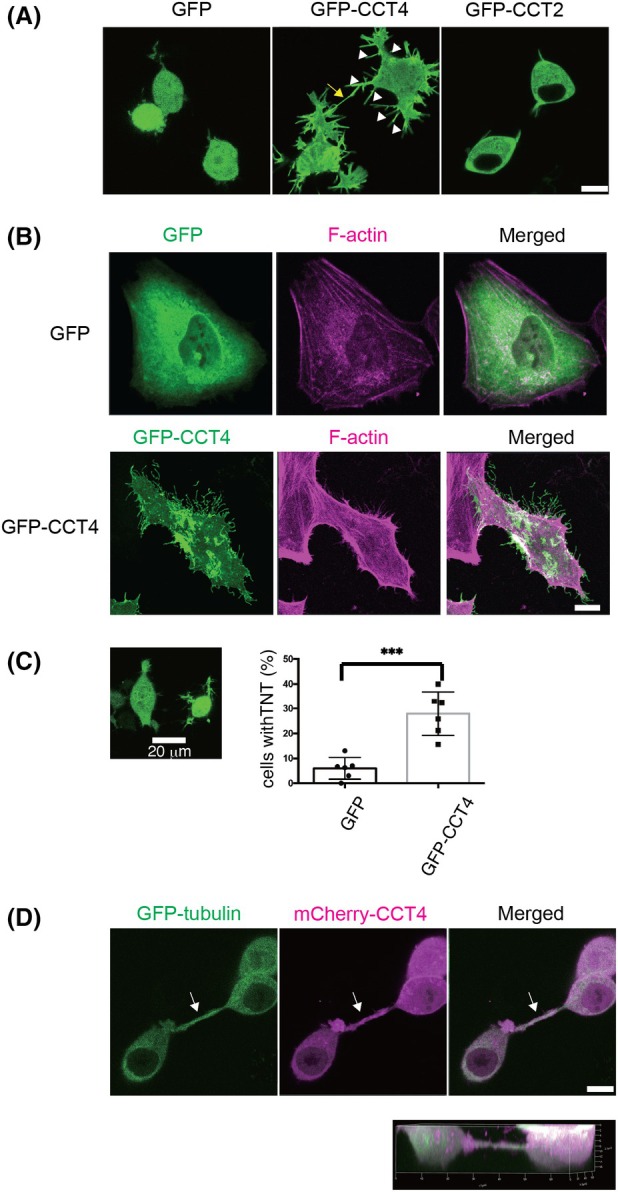 Fig. 1
Fig. 1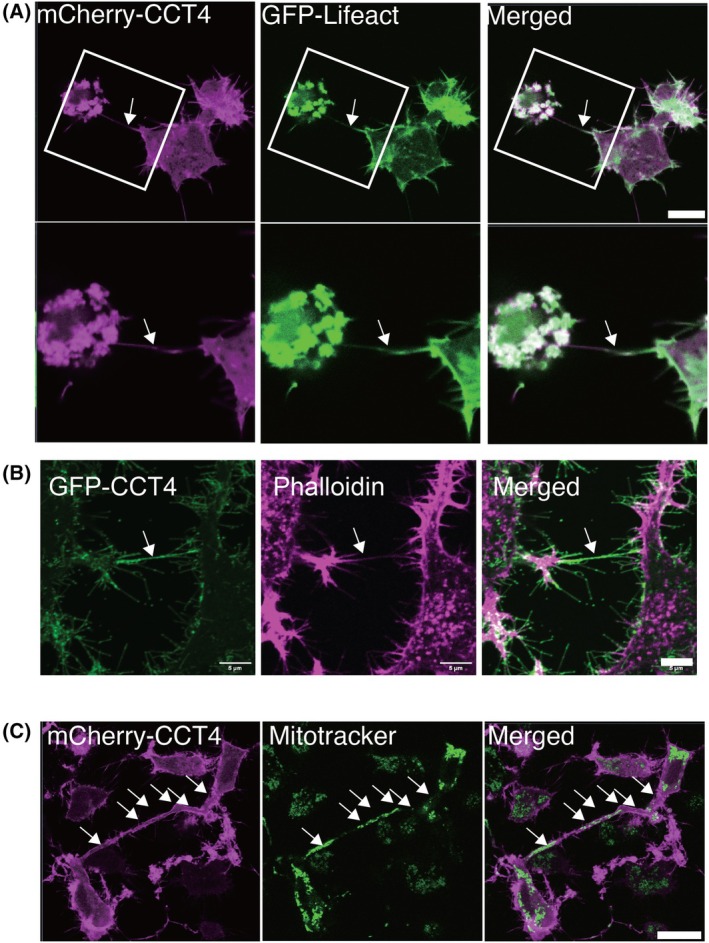 Fig. 2
Fig. 2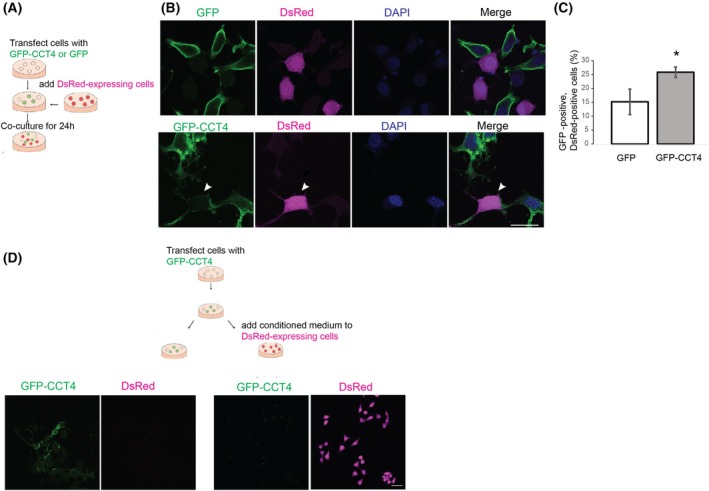 Fig. 3
Fig. 3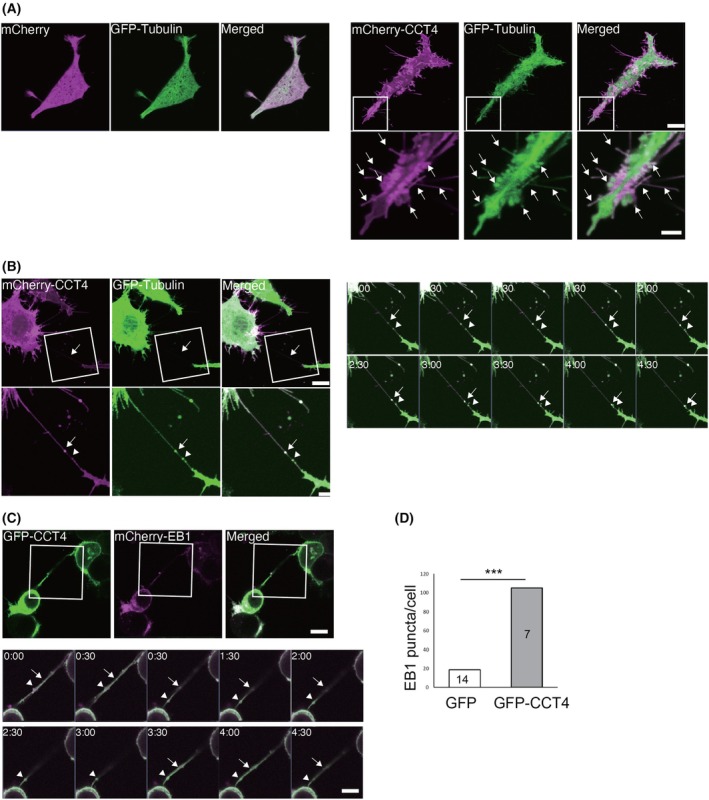 Fig. 4
Fig. 4- —Takeda Science Foundation10.13039/100007449
- —Japan Agency for Medical Research and Development10.13039/100009619
- —Japan Society for the Promotion of Science10.13039/501100001691
- —Tokyo Metropolitan University, TMU strategic research fund10.13039/501100010236
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA and protein synthesis mechanisms · ATP Synthase and ATPases Research · Protein Structure and Dynamics
Abbreviations
CCT4, chaperonin‐containing T‐complex protein 1 Subunit delta
EB1, end‐binding protein 1
F‐actin, filamentous actin
TNT, tunneling nanotube
TRiC, chaperonin‐containing TCP1 complex
Tunneling nanotubes (TNTs) are open membranous channels between connected cells, allowing direct intercellular communication [1, 2, 3]. TNTs have lengths in a broad range of 3–100 μm and widths ranging from 50 nm to 2 μm [4], contain filamentous actin (F‐actin) and sometimes microtubules, and transport small molecules such as calcium ions, macromolecules such as nucleic acids and proteins, and large cargos, including organelles such as vesicles, lysosomes, autophagosomes, and mitochondria, to mediate fast and specific responses between cells [1].
TNTs also play important roles in stress responses and disease pathogenesis [5, 6]. TNT‐mediated material transfers regulate cancer phenotypes such as invasiveness, metabolic plasticity, and therapy resistance [5]. In neurodegenerative diseases, including Parkinson's and Alzheimer's disease, TNTs transfer protein aggregates from neurons to microglia to protect neurons [7] or spread to other neurons via TNTs to propagate [8, 9, 10, 11]. TNT‐mediated mitochondrial transfer can protect stressed cells or boost cellular fitness [7, 12]. Despite their physiological and pathological importance, the molecular mechanisms of TNT formation are not fully understood.
CCT4 is a component of the chaperonin‐containing TCP1 complex (CCT/TRiC) [13]. TRiC is formed by eight subunits and assists in the folding of various proteins involved in the cytoskeleton, translation, protein degradation, and signaling [14]. Some CCT subunits also function as monomers, working independently of TRiC [15, 16, 17]. CCT4 has been shown to influence the cytoskeletal structure and regulate F‐actin organization independently of TRiC. In cultured cells, overexpression of a monomeric form of CCT4 induces actin‐based protrusions at the cell surface [15], which are caused by CCT4 interaction with p150^Glued^ [16]. In this study, we set out to investigate whether CCT4 is also involved in TNT formation.
Materials and methods
Chemicals
Fluorescent‐labeled phalloidin St. Louis, MO, USA (Phalloidin‐Atto 647N, Sigma, catalog # 65906) and MitoTracker Green FM Waltham, MA, USA (Thermo Fisher Scientific, catalog # M7514) were purchased.
Plasmids
GFP‐CCT4 and GFP‐CCT2 [15] were provided by Dr. Julie Grantham (University of Gothenburg). mCherry‐CCT4 was constructed using NEB Builder HiFi DNA Assembly Ipswich, MA, USA (New England Biolabs, catalog # E2621S). CCT4 was amplified by PrimeSTAR Max DNA polymerase (TAKARA BIO) using primers, 5′‐CTAGCCAGCTCGTCCATGCCCGGGAAGCTACGTTCTCCGG‐3′ and 5′‐GATGTGGTAAATACTCGATAACTAGATAACTGATCATAATC‐3′ and assembled into pmCherry‐C1. EGFP‐tubulin‐6 was purchased from Addgene (catalog # 56450) (Rizzo et al., Cold Spring Harb Protoc. 2019). GFP‐Lifeact was purchased from Addgene (catalog # 58470). mCherry‐EB1 was constructed by subcloning mouse EB1 from mouse brain cDNAs into pmCherry‐N1A by NEB builder.
Cell culture and transfection
Neuro‐2a (RRID: CVCL_0470) was obtained from Dr. Masao Takeuchi at the Institute for Fermentation, Osaka, Japan. Neuro‐2a cells were cultured in minimum essential medium (MEM) supplemented with 1% nonessential amino acids, 10% fetal bovine serum (FBS), 100 U·mL^−1^ penicillin, and 0.1 mg·mL^−1^ streptomycin. HeLa (RRID:CVCL_0030) was purchased from the RIKEN BioResource Center, Japan (RCB0007). HeLa cells were cultured in D‐MEM (High Glucose) with L‐Glutamine and Phenol Red with 10% fetal bovine serum (FBS), 100 U·mL^−1^ penicillin, and 0.1 mg·mL^−1^ streptomycin. These cell lines are microplasma‐free and have been authenticated in the past three years by morphology check by microscope and growth curve analysis.
Cells were transfected with plasmids using Lipofectamine 2000 and Lipofectamine 3000 Carlsbad, California, USA (Invitrogen, catalog # L3000015 and 11 668 019) according to the manufacturer's protocol. Neuro2A cells stably expressing the fluorescent protein DsRed were established by transfecting DsRed plasmids using Lipofectamine 2000, followed by selection with medium containing 500 μg·mL^−1^ G418 (WAKO, catalog # 078–05961) for two weeks, replacing the medium every 2–3 days. Surviving clones were isolated, expanded, and screened for stable expression of DsRed using Keyence microscope BZ‐X700 Osaka, Japan (Keyence).
Imaging fixed cells
HeLa and Neuro 2a cells were plated on coverslips and transfected. Cells were fixed with 4% PFA in PBS for 20 min at 37 °C, permeabilized with 0.1% Triton X‐100 and stained with fluorescent‐labeled phalloidin or DAPI. Samples were observed using a confocal microscope Zeiss LSM710 (Zeiss), and the obtained results were analyzed by ImageJ [18].
Live cell imaging
Neuro 2a cells were plated on a noncoated glass‐bottom dish, transfected with indicated plasmids, and observed under a confocal microscope Zeiss LSM710 (Zeiss). Results were analyzed by ImageJ (NIH) [19].
Statistics
Statistical analyses were carried out with microsoft excel (Microsoft), python (Python 3), and graphpad prism (GraphPad). Differences were assessed using the Student's t‐test, chi‐squared test, or one‐way ANOVA followed by Tukey's HSD. P values < 0.05 were considered statistically significant.
Results
Monomeric CCT4 promotes nanotubes that connect neighboring cells
To analyze the function of monomeric CCT4 proteins, we used CCT4 tagged with GFP in its N terminus (GFP‐CCT4), which is not incorporated into the chaperonin oligomer [15]. In Neuro2A cells and HeLa cells, GFP‐CCT4 was distributed to the plasma membrane (Fig. 1A,B), as reported with B16F1 cells previously [15]. Cells with GFP‐CCT4 expression had numerous protrusions, as previously reported [15]. In addition, those cells have nanotube‐like structures, which are thicker and longer than protrusions and connect neighboring cells (Fig. 1A,B, protrusions (arrow heads) and a tube connecting cells (arrow)). These nanotubes are often more than 1 μm in thickness and connect cells apart more than 20 μm. Formation of many protrusions and nanotube‐like structures was not observed with cells transfected with GFP alone (Fig. 1A, GFP) or those transfected with chaperonin‐containing TCP1 complex subunit 2 (CCT2) (Fig. 1A, GFP‐CCT2). We analyzed the percentage of cells with nanotubes for Neuro2A cells expressing GFP and those expressing GFP‐CCT4 and found that cells with GFP‐CCT4 expression have nanotubes significantly more often: about 5% of GFP‐expressing cells and about 30% of GFP‐CCT4‐expressing cells have nanotubes (Fig. 1C). Nanotubes thicker than 1 μm contain tubulin: 22 out of 22 nanotubes from three independent experiments were tubulin‐positive.
*GFP‐CCT4 induces tunneling nanotube formation in cultured cells. (A) Live cell imaging of Neuro2A cells transfected with GFP, GFP‐CCT4, and GFP‐CCT2. GFP‐CCT4 expression induces the formation of protrusions (arrowheads) and nanotubes connecting two cells (arrow). Scale bar, 10 μm. (B) HeLa cells transfected with GFP and GFP‐CCT4. Cells were fixed and stained with phalloidin (magenta) and DAPI (blue) to visualize actin fibers and nuclei. GFP‐CCT4 expression induces formation of protrusions. Scale bar, 10 μm. (C) GFP‐CCT4‐transfected cells develop nanotubes more often than GFP‐transfected cells. Cells with nanotubes with the neighboring cells more than 20 μm away were scored. Scale bar, 20 μm. Thirty cells in each of 6 independent wells were analyzed. Mean ± SD, **P < 0.001; Student's t‐test. (D) Nanotubes extended from CCT4‐expressing cells are floating above coverslips. Live imaging of Neuro2A cells transfected with GFP‐Tubulin (green) and mCherry‐CCT4 (magenta). A full‐focus image of Z‐stuck (top) and a 3D‐reconstructed image (bottom). The arrow indicates the floating tube. scale bar, 10 μm.
GFP‐CCT4 expression has been reported to induce retraction fibers and filopodia‐like protrusions [15], while the formation of nanotubes has not been reported previously. Retraction fibers and filopodia‐like protrusions are attached to the substrate, while TNTs are hovering over the substrate. We found that the nanotubes from GFP‐CCT4‐expressing cells are floating above coverslips (Fig. 1D), indicating that these structures are not retraction fibers or filopodia.
Nanotubes induced by GFP‐CCT4 contain F‐actin and mitochondria
TNTs contain actin fibers and microtubules, and organelles such as mitochondria are transported on microtubules in TNTs [1]. We asked whether the nanotubes of cells with GFP‐CCT4 expression have these features. Live imaging of cells expressing GFP‐LifeAct and staining of fixed cells with fluorescent‐labeled phalloidin showed that CCT4‐induced nanotubes connecting two cells contain actin fibers (Fig. 2A,B). GFP staining with a mitochondrial dye, mitotracker, revealed that these nanotubes contain mitochondria (Fig. 2C).
GFP‐CCT4 expression induces the formation of nanotubes containing F‐actin and tubulin puncta. (A) Live‐imaging of Neuro2A transfected with mCherry‐CCT4 (magenta) and GFP‐Lifeact (green). Arrows indicate a nanotube connecting two cells. Scale bar, 10 μm. (B) Neuro2A was transfected with GFP‐CCT4 (green) and stained with phalloidin (magenta). Arrows indicate the nanotube connecting two cells. Scale bar, 5 μm. (C) Live imaging of Neuro2A cells transfected with mCherry‐CCT4 (magenta) was stained with Mitotracker (green). Arrows indicate mitochondria. Scale bar, 20 μm.
Nanotubes induced by GFP‐CCT4 connect neighboring cells
If nanotubes from cells expressing GFP‐CCT4 connect neighboring cells and are open‐ended, they would transfer proteins. We asked whether GFP‐CCT4 proteins are transferred intercellularly using a coculture experiment. Cells transfected with GFP‐CCT4, GFP‐CCT2, or GFP were mixed with cells that stably express DsRed. After 24 h of coculture, GFP signals in DsRed‐positive cells were analyzed (Fig. 3A). In the wells of cells transfected with GFP‐CCT4, we found some DsRed‐cells connected with GFP‐CCT4‐expressing cells show GFP signal, suggesting that GFP‐CCT4 proteins were transferred (Fig. 3B, arrowheads). DsRed cells with GFP signal were more frequently observed with the coculture of GFP‐CCT4 expressing cells than with the coculture of GFP‐expressing cells (Fig. 3C), suggesting that GFP‐CCT4 can facilitate the intracellular transfer of proteins.
*GFP‐CCT4 is transferred to neighboring cells via nanotubes. GFP‐CCT4 spreads to neighboring cells that are connected with tunneling nanotubes (TNTs). (A) Experimental scheme. (B) Neuro2A cells transfected with GFP or those transfected with GFP‐CCT4 were mixed with DsRed stable transfectants. Cells were counterstained with DAPI (blue). The arrowhead indicates a DsRed‐positive cell with GFP‐CCT4. (C) The number of cells positive for GFP, those for DsRed, and those for both GFP and DsRed were counted (top) and expressed as the percentage of GFP‐positive cells (bottom). Mean ± SE, n = 3, P < 0.05; Student's t‐test. (D) DsRed‐expressing Neuro2A cells were incubated with a conditioned medium of cells transfected with GFP‐CCT4 for 24 h. Left: Cells transfected with GFP‐CCT4. Right: DsRed‐expressing Neuro2A incubated with conditioned medium. No GFP‐positive cells were observed. scale bar, 10 μm.
Proteins can be intercellularly transported via the extracellular medium. We asked whether CCT4 is transferred to neighboring cells via the conditioned medium. Cells expressing DsRed were cultured for 24 h in the conditioned medium collected from cells transfected with GFP‐CCT4 (Fig. 3D). There were no DsRed cells that were positive for GFP (Fig. 3D). These results suggest that TNT formed in GFP‐CCT4‐expressing cells is open‐ended and transfers proteins between cells.
Monomeric CCT4 enhances tubulin polymerization in TNTs
To visualize the remodeling of the cytoskeleton in these structures, we cotransfected mCherry‐CCT4 and GFP‐tubulin. GFP‐tubulin distributes to TNTs extending from cells expressing GFP‐CCT4 (Fig. 4A, Movies S1 and S2). Interestingly, GFP‐tubulin puncta are moving from cells with GFP‐CCT4 toward the neighboring cells connected with TNTs at the speed of approximately 80 nm·min^−1^ (Fig. 4B). To observe the microtubule elongation, we co‐expressed end‐binding protein 1 (EB1) fused with mCherry, which labels the microtubule growing end [20]. Live imaging of Neuro2A cells co‐expressing GFP‐CCT4 and mCherry‐EB1 revealed that the expression of GFP‐CCT4 increased the number of EB1 puncta (Fig. 4C,D, Movie S3). EB1 puncta were also observed in TNTs, and CCT4 was observed as puncta nearby or sometimes overlapping with tubulin puncta or EB1 puncta (arrows in Fig. 4B,C, Movie S4). These results suggest that monomeric CCT4 regulates microtubule organization in the cytosol and TNTs.
*Overexpression of CCT4 enhances microtubule dynamics. (A) Neuro2A cells transfected with mCherry (magenta) and GFP‐αtubulin (green) (Left) and mCherry‐CCT4 (magenta) and GFP‐α tubulin (Green) (Right). Blow‐up images are shown in the bottom panels. Arrows indicate co‐localization of mCherry‐CCT4 and GFP‐α tubulin. scale bars, 10 μm (top) and 5 μm (bottom). (B) Neuro2A cells were transfected with mCherry‐CCT4 (magenta) and GFP‐α tubulin (green) and imaged every 30 s for 5 min. GFP‐ α tubulin formed puncta in tunneling nanotubes (TNTs) (arrow and arrowhead), which were moving at approximately 80 nm·s−1. The arrow indicates tubulin puncta colocalized with CCT4. Scale bar, 10 μm. (C) Neuro 2A cells were transfected with GFP‐CCT4 and mCherry‐EB1. Blow‐up images of the TNT are shown at the bottom. The arrow and arrowhead indicate CCT4 puncta in TNT, and the arrow indicates CCT4 puncta overlapping with EB1. scale bars, 10 μm (top) and 5 μm (bottom). (D) GFP‐CCT4 increased the number of EB1 puncta. Three different areas containing about five cells each were imaged, and the number of EB1 puncta was counted. The numbers in the bars indicate the number of cells analyzed. **P < 0.001; Chi‐square test.
Discussion
In this study, we found that monomeric CCT4 induces TNT formation (Figs 1, 2, 3). TNTs have been reported in various conditions and cellular contexts, and they are variable in their morphology and cytoskeleton content [1]. While some TNTs contain F‐actin alone as the cytoskeleton, some contain both F‐actin and microtubules [1]. Expression of monomeric CCT4 increased the number of thick TNTs containing tubulin (Fig. 1) and CCT4‐induced TNTs contain growing microtubules (Fig. 4), suggesting that CCT4 promotes the formation of nanotubes containing microtubules.
Although TRiC is known to mediate the folding of cytoskeletal proteins, including tubulins [21, 22], the role of monomeric CCT4 in microtubule polymerization has not been reported previously. Connectome analyses revealed that CCT4 proteins interact with tubulins [18, 20, 23, 24, 25, 26] and proteins involved in microtubule remodeling such as tubulin tyrosine ligase‐like family member 3 [18, 23, 24], histone deacetylase 6 [27], histone deacetylase 11 [26], cyclin‐dependent kinase 5 [28, 29, 30], and cilia and flagella‐associated protein 52 [31]. Although these analyses do not distinguish proteins interacting with the TRiC complex from those with monomeric CCT4, some of these proteins may interact with monomeric CCT4 and regulate microtubule dynamics in TNTs. It has been reported that the expression of GFP‐CCT4 in cultured mammalian cells induces retraction fiber formation [15], and this phenotype is caused by interactions between CCT4 and p150^Glued^ [16]. Interestingly, an isoform of p150^Glued^ expressed primarily in neurons interacts with tubulin dimers and microtubules and functions as an anticatastrophic factor [32]. Morphological changes caused by GFP‐CCT4 include increased membrane curvature and lipid composition changes, such as increased phosphatidylethanolamine lipid species [33]. Such lipid composition alterations may increase membrane fluidity and might contribute to TNT formation. Further studies on proteins interacting with monomeric CCT4 in TNTs may provide insights into mechanisms underlying their morphological and functional variation.
CCT4 has also been linked to neurological diseases: a mutation in CCT4/5 subunits causes sensory neuropathy [34], and CCT4 expression is upregulated in AD [35]. Functions of CCT4 in TNT formations may also contribute to understanding the pathogenesis of these disorders.
Conflicts of interest
The authors declare no conflict of interest.
Author contributions
Conceptualization: AA, ME, and KA, Methodology: AA, ME, and KA, Formal analysis: AA, ME, and KA, Investigation: AA, ME, and KA, Writing – original draft preparation: ME and KA, Writing – review and editing: AA, TS, and KA, Supervision: AA, TS, and KA, Project administration: KA, Funding acquisition: KA.
Supporting information
Movie S1. GFP‐ αtubulin (green) in Neuro2A cells expressing mCherry.
Movie S2. GFP‐ αtubulin (green) in Neuro2A cells expressing mCherry‐CCT4.
Movie S3. GFP‐ αtubulin (green) and mCherry‐CCT4 (magenta) in the nanotube (Fig. 4B).
Movie S4. mCherry‐EB1 in Neuro 2A cells expressing GFP‐CCT4 (Fig. 4C).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cordero Cervantes D and Zurzolo C (2021) Peering into tunneling nanotubes‐the path forward. EMBO J 40, e 105789.33646572 10.15252/embj.2020105789 PMC 8047439 · doi ↗ · pubmed ↗
- 2Rustom A , Saffrich R , Markovic I , Walther P and Gerdes HH (2004) Nanotubular highways for intercellular organelle transport. Science 303, 1007–1010.14963329 10.1126/science.1093133 · doi ↗ · pubmed ↗
- 3Sartori‐Rupp A , Cordero Cervantes D , Pepe A , Gousset K , Delage E , Corroyer‐Dulmont S , Schmitt C , Krijnse‐Locker J and Zurzolo C (2019) Correlative cryo‐electron microscopy reveals the structure of TN Ts in neuronal cells. Nat Commun 10, 342.30664666 10.1038/s 41467-018-08178-7PMC 6341166 · doi ↗ · pubmed ↗
- 4Saha T , Dash C , Jayabalan R , Khiste S , Kulkarni A , Kurmi K , Mondal J , Majumder PK , Bardia A , Jang HL et al. (2022) Intercellular nanotubes mediate mitochondrial trafficking between cancer and immune cells. Nat Nanotechnol 17, 98–106.34795441 10.1038/s 41565-021-01000-4PMC 10071558 · doi ↗ · pubmed ↗
- 5Pinto G , Brou C and Zurzolo C (2020) Tunneling nanotubes: the fuel of tumor progression? Trends Cancer 6, 874–888.32471688 10.1016/j.trecan.2020.04.012 · doi ↗ · pubmed ↗
- 6Chastagner P , Loria F , Vargas JY , Tois J , Diamond IM , Okafo G , Brou C and Zurzolo C (2020) Fate and propagation of endogenously formed tau aggregates in neuronal cells. EMBO Mol Med 12, e 12025.33179866 10.15252/emmm.202012025 PMC 7721367 · doi ↗ · pubmed ↗
- 7Scheiblich H , Eikens F , Wischhof L , Opitz S , Jungling K , Cserep C , Schmidt SV , Lambertz J , Bellande T , Posfai B et al. (2024) Microglia rescue neurons from aggregate‐induced neuronal dysfunction and death through tunneling nanotubes. Neuron 112, 3106–3125.39059388 10.1016/j.neuron.2024.06.029 · doi ↗ · pubmed ↗
- 8Abounit S , Bousset L , Loria F , Zhu S , de Chaumont F , Pieri L , Olivo‐Marin JC , Melki R and Zurzolo C (2016) Tunneling nanotubes spread fibrillar alpha‐synuclein by intercellular trafficking of lysosomes. EMBO J 35, 2120–2138.27550960 10.15252/embj.201593411 PMC 5048354 · doi ↗ · pubmed ↗