Microbiome Therapeutic Lactiplantibacillus plantarum PMC105 for Systemic Carbapenem-Resistant Enterobacteriaceae Infections: Oral and Inhalation Efficacy In Vivo
Faezeh Sarafraz, Hoonhee Seo, Hanieh Tajdozian, Ali Atashi, Youjin Yoon, Sukyung Kim, Ho-Yeon Song

TL;DR
A new probiotic strain, Lactiplantibacillus plantarum PMC105, shows promise in treating deadly antibiotic-resistant bacterial infections in mice.
Contribution
Demonstrates the in vivo efficacy of PMC105 via oral and inhalation routes against systemic CRE infections.
Findings
Oral PMC105 reduced intestinal CRKP colonization and achieved 100% survival in infected mice.
Intranasal PMC105 decreased bacterial loads in multiple organs and improved survival rates.
PMC105 was confirmed safe through comprehensive safety evaluations.
Abstract
Carbapenem-resistant Enterobacteriaceae (CRE), particularly Klebsiella pneumoniae, are major causes of severe systemic infections due to their resistance to most antibiotics and the high associated mortality, representing a growing global health concern. In this study, we report the in vivo efficacy of a novel probiotic strain, Lactiplantibacillus plantarum PMC105, against systemic CRE infections. In a mouse model characterized by neutropenia and antibiotic-induced gut dysbiosis, infection with carbapenem-resistant K. pneumoniae (CRKP) resulted in 60% mortality within two weeks. However, oral administration of PMC105 significantly reduced intestinal CRKP colonization, minimized body weight loss, and resulted in 100% survival. This therapeutic effect is presumed to result from enhanced gut barrier function, driven by upregulation of the tight junction protein ZO-1 in the ileum, thereby…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
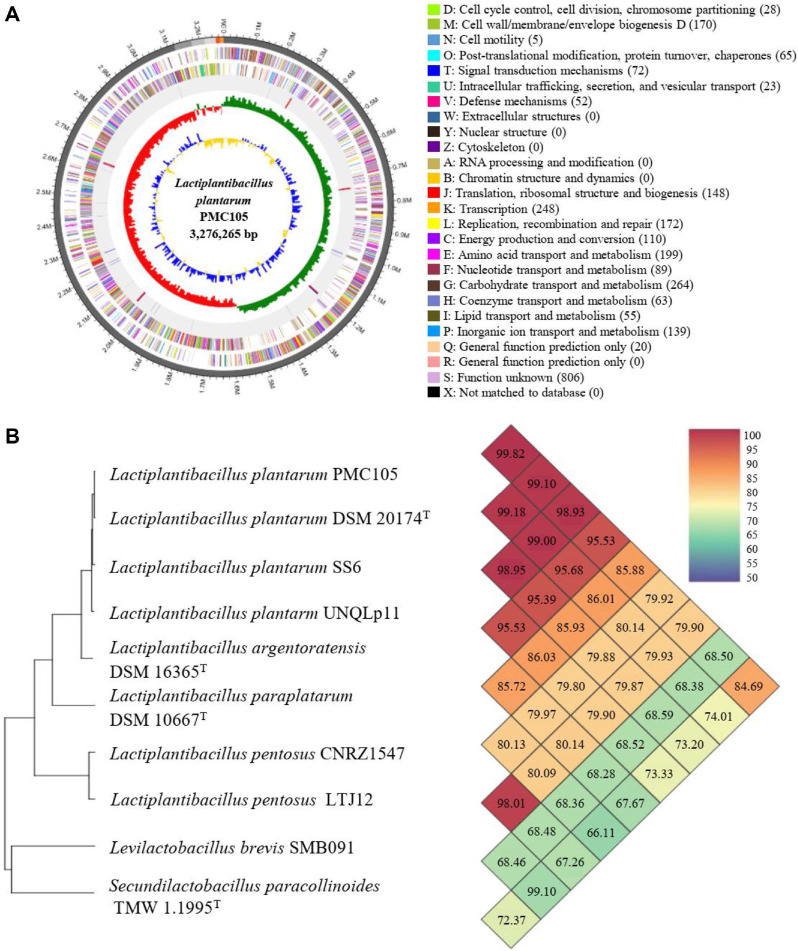 Figure 1
Figure 1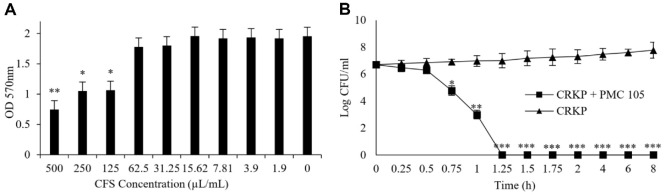 Figure 2
Figure 2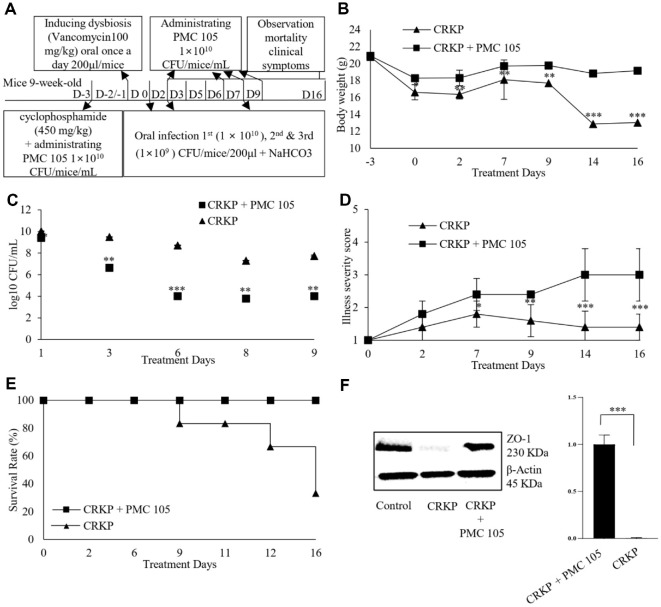 Figure 3
Figure 3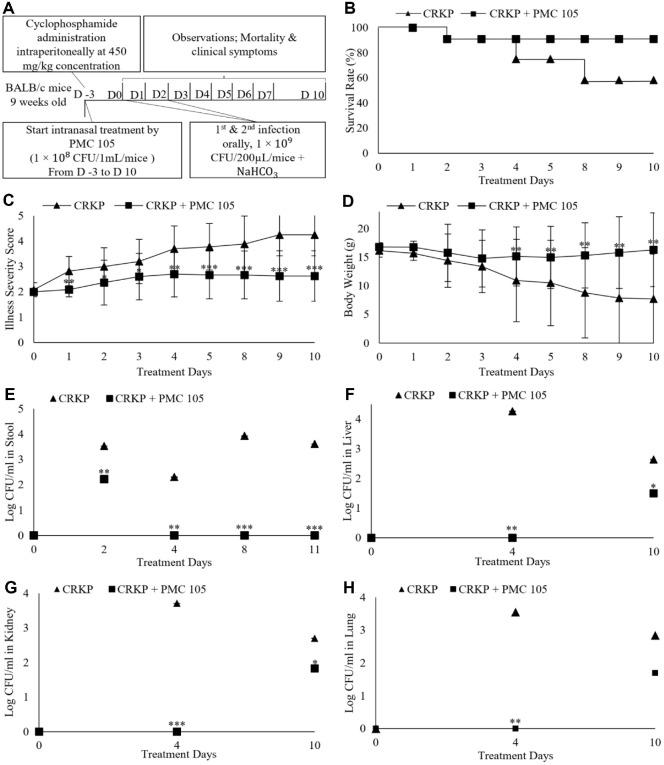 Figure 4
Figure 4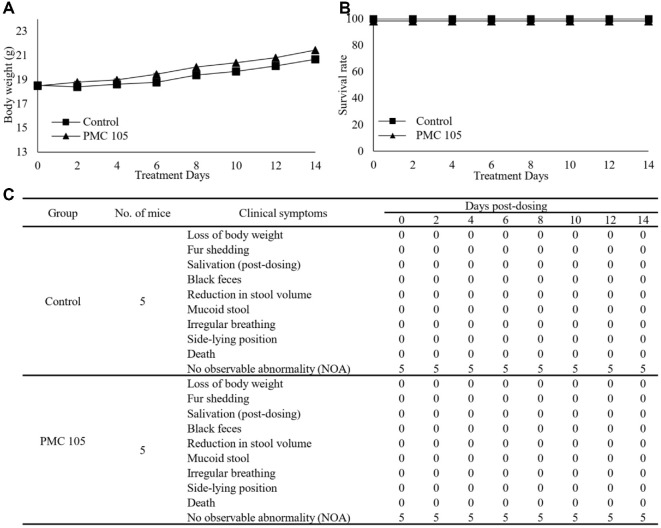 Figure 5
Figure 5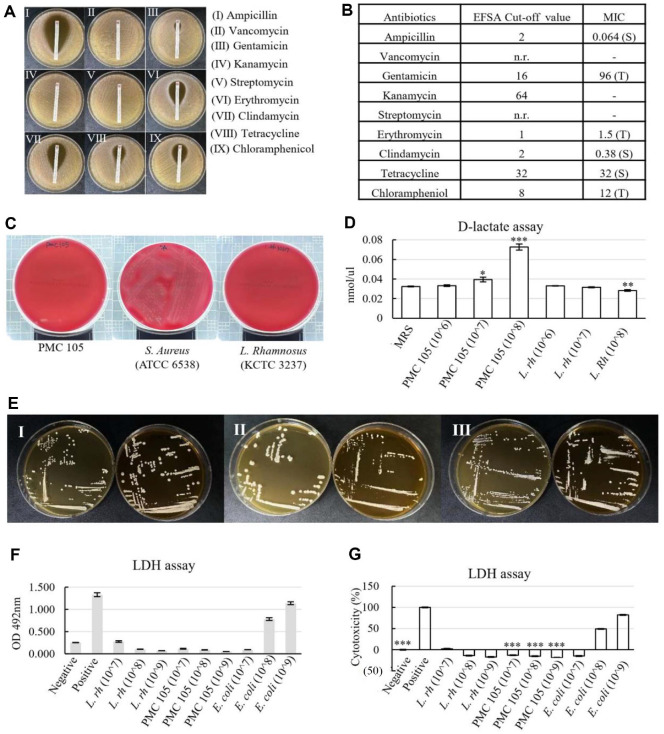 Figure 6
Figure 6 Figure 7
Figure 7- —National Research Foundation of Koreahttp://dx.doi.org/10.13039/501100003725
- —Ministry of Science and ICT, South Korea10.13039/501100014188
- —Korea Institute for Advancement of Technologyhttp://dx.doi.org/10.13039/501100003661
- —Ministry of Trade, Industry and Energy10.13039/501100003052
- —Soonchunhyang University10.13039/501100002560
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Antibiotic Resistance in Bacteria · Gut microbiota and health
Introduction
The rise in carbapenem-resistant Enterobacteriaceae (CRE) has emerged as a significant global health threat, associated with high morbidity and mortality worldwide [1]. Carbapenemases, including New Delhi metallo–β–lactamase (NDM), Klebsiella pneumoniae carbapenemase (KPC), and OXA−48, are key contributors to the antibiotic resistance mechanisms observed in CRE [2].
K. pneumoniae, a Gram-negative, rod-shaped bacterium from the Enterobacteriaceae family, is a prominent opportunistic pathogen that critically impacts the management of patients at risk of colonization or infection by such microorganisms [3]. Its ability to cause community-acquired and hospital-acquired infections, and its tendency to develop antibiotic resistance, are critical challenges worldwide [4]. At present, carbapenem-resistant Klebsiella pneumoniae (CRKP) is resistant to all available antibiotics, making it extremely difficult to treat, and leaving minimal therapeutic options [5, 6]. Addressing this urgent issue requires the development of novel treatment strategies to manage these resistant bacteria and mitigate their impact on public health [7].
The Food and Agriculture Organization (FAO) and the World Health Organization (WHO) describe probiotics as “live microorganisms which, when administered in adequate amounts, confer a health benefit on the host” [8]. Probiotics can enhance gut health and immune response by producing antimicrobial substances that improve the host’s resistance to gastrointestinal infections [9, 10].
Among probiotics, lactobacilli, particularly Lactiplantibacillus plantarum, are recognized for their antimicrobial properties against various pathogens [11, 12]. L. plantarum, a Gram-positive bacterium, is one of the most used probiotics due to its immune-modulating and therapeutic effects [13]. Studies have demonstrated the antimicrobial impacts of lactobacilli against a wide range of pathogenic microorganisms, including carbapenem-resistant Enterobacteriaceae (CRE) [13???-17], Listeria monocytogenes [18], Salmonella typhimurium [19, 20], Escherichia coli [21, 22], Streptococcus mutans, Aggregatibacter actinomycetemcomitans [10], Staphylococcus aureus [23], Shigella species [24], Clostridium difficile [25], Pseudomonas aeruginosa [26, 27], and Salmonella species [28].
This study aims to evaluate the in vivo antimicrobial activity of L. plantarum, isolated from the traditional fermented kimchi, to assess its potential to prevent and treat CRKP intestinal infections. In addition to oral delivery, an intranasal (nebulization) route was applied in this study to explore the potential of L. plantarum PMC105 as a systemic microbiome-based therapeutic. Although oral administration is the most common route for probiotics, intranasal delivery offers unique advantages by allowing direct contact with the nasal mucosa and respiratory epithelium, where abundant mucosa-associated lymphoid tissue (NALT) can initiate systemic immune responses. Recent experimental studies have demonstrated the feasibility of intranasal probiotic delivery; for example, strains of L. rhamnosus GG [29, 30], L. johnsonii [31], and L. casei [32] administered intranasally in mice reduced pathogen burden and exhibited anti-inflammatory effects. The objective is to determine whether L. plantarum can inhibit pathogen growth and enhance immune responses in murine models, thereby offering a novel approach to managing CRKP infections.
Materials and Methods
Carbapenem-Resistant Klebsiella pneumoniae
A clinical Carbapenem-resistant Klebsiella (CRKP) isolate was obtained from the Pathogenic Resource Bank at Soonchunhyang University Hospital [33]. The strain was streaked onto MacConkey agar plates (BD Difco, USA) containing 10 mg/ml imipenem (USA) and incubated for 18 h at 37°C under aerobic conditions. The selected single colonies were then cultured in MacConkey broth (BD Difco) aerobically at 37°C for 24 h. Bacterial cultures were grown to an optical density of 1.0 at 600 nm, diluted to 2 × 10^9^ CFU/ml, preserved in sterile glycerol, and stored at −80°C.
Isolation and Preparation of Probiotics
To isolate the probiotic candidate strain, various traditional fermented foods—including kimchi, soybean products, and fermented vegetables- were used as sources. Approximately 10 g of each sample was streaked onto De Man, Rogosa, and Sharpe (MRS) agar (BD Difco) plates supplemented with vancomycin at 4 μg/ml (Sigma-Aldrich, USA) and incubated at 37°C for 18 h under a microaerophilic chamber (Daeiltech, Republic of Korea). A total of nine lactic acid bacteria isolates were initially obtained. All isolates were subjected to Gram staining and catalase testing to confirm Gram-positive, catalase-negative phenotypes. They were then screened for acid tolerance (pH 3.0), bile salt tolerance (0.3% oxgall), and antagonistic activity against CRKP using an agar well diffusion assay. Among these, three isolates produced clear inhibition zones, and one isolate demonstrated stable viability under simulated gastrointestinal conditions. This isolate showed the most potent antimicrobial activity, acid/bile tolerance, and non-hemolytic, non-cytotoxic properties. Based on these criteria, it was selected as the final probiotic candidate for further identification, characterization, and in vivo evaluation. The selected single colony was cultured in MRS Broth (BD Difco) and incubated at 37°C for 18 h. Spectrophotometry was used to determine the optical density of bacterial growth (OD = 1.0, 1 × 10^9^ CFU/ml), and the stocks were then prepared by adding 60% glycerol and stored at −80°C for further study. Later, the stock was cultured in Food-Grade Media (FGM broth) containing yeast-peptone standard type of F (MBcell, Republic of Korea), Tween 80 (Sigma-Aldrich), d–glucose (Sigma-Aldrich), and magnesium sulfate heptahydrate (Sigma-Aldrich) at 37°C for 24 h [14] for an in vivo experiment.
Identification of the Probiotic Candidate Using 16S rRNA
After isolating the probiotic candidate, the selected strain was sent to Biofact Company in South Korea for 16S ribosomal RNA gene sequencing. The two primers used for amplification were 27f (5'-AGAGTTTGATCCTGGCTCAG-3') and 1495R (5'-CTACGGCTACCTTGTTACGA-3'), as described previously [33, 34]. PCR amplification was performed with an initial denaturation at 95°C for 3 min, followed by 30 cycles of denaturation at 95°C for 20 sec, annealing at 56°C for 40 sec, and extension at 72°C for 90 sec, with a final extension at 72°C for 5 min. The PCR products were then purified using a PCR purification kit and sequenced using the BigDye Terminator v3.1 Cycle Sequencing Kit (Thermo Fisher Scientific, USA) on an ABI PRISM 3730XL DNA Analyzer (Applied Biosystems, USA). To identify the bacterial species, the NBLAST search tool (http://www.ncbi.nlm.nih.gov/blast) was used to compare the most abundant unique sequences against the NCBI GenBank microbial database. Matches were evaluated based on their e-values and bit scores, and the taxonomy of the best-matching sequence was assigned to each query.
Whole Genome Sequencing of L. plantarum via PacBio SMRT Sequencing
Whole-genome sequencing was performed by a WGS service provider (CJBioscience, Republic of Korea) to characterize our candidate probiotic. A QIAamp DNA Mini kit (Qiagen, Germany) was used to extract genomic DNA (gDNA) from the L. plantarum strain. Sequencing was performed using PacBio technology, with the HGAP2 protocol in PacBio SMRT Analysis 2.3.0. Circlator 1.4.0 (Sanger Institute, UK) was used to process the data, generating circularized contigs from the PacBio sequencing. Protein-coding sequences (CDSs) were estimated using Prodigal 2.6.2 [35] and categorized based on orthologous groups through the EggNOG database (http://eggnogdb.embl.de). Genes encoding tRNAs were identified using tRNAscan–SE 1.3.1 [36], while other noncoding RNAs were detected through covariance model searches in the Rfam 12.0 database [37]. The OrthoANIu algorithm-based Average Nucleotide Identity (ANI) calculator was used to compute OrthoANI values for prokaryotic genome sequences [38].
Minimum Inhibitory Concentration (MIC) of L. plantarum Cell-Free Supernatant (CFS)
L. plantarum strain was cultured in MRS broth media (BD Difco) at 37°C for 24 h to reach maximal growth, and the turbidity was adjusted to 0.5 McFarland. The cultured media were centrifuged for 20 min at 4,000 rpm. 0.22 μm filters were used to filter the cell-free supernatant to obtain sterile CFS. An overnight culture of CRKP with a turbidity of 0.5 McFarland standard was diluted 1:100 into MacConkey broth (BD DifcoUSA) to reach a bacterial concentration of 1 × 10^6^ CFU/ml. The minimum inhibitory concentration (MIC) test was conducted using the microtiter method [39]. In brief, 100 μl of the diluted CRKP was added to the wells of a 96–well plate. A 2-fold serial dilution of CFS was prepared in MRS media, and 100 μl of each dilution was added to the wells containing the bacterial inoculum. The positive control contained only CRKP inoculum in fresh MRS medium, while the negative control contained a 1:1 mixture of MacConkey and MRS media without bacteria. The plate was then incubated at 37°C for 24 h. Bacterial growth was measured by reading the optical density at 570 nm using a microplate reader. The MIC was determined as the lowest concentration of CFS that completely inhibited visible bacterial growth.
Time-Kill Assay
To evaluate the bactericidal activity of L. plantarum against CRKP, a Time-kill assay was performed using its CFS, as previously described [14, 40, 41]. The L. plantarum strain was cultured in MRS broth (BD Difco) and incubated overnight at 37°C under aerobic conditions. The culture was then centrifuged, and the supernatant was filter–sterilized (0.22 μm) to obtain the CFS. For the assay, 300 μl of CRKP suspension (10^8^ CFU/ml) was inoculated into 15 ml of L. plantarum CFS and incubated at 37°C. The Control group consisted of CRKP suspended in sterile MRS broth without CFS inoculum. Aliquots were pipetted out from each tube at predetermined time intervals, and serially diluted to determine the viable CFU of CRKP on MacConkey agar (BD Difco). The experiment was conducted in triplicate, and total surviving cells were expressed as the mean log_10_ CFU/ml. Data are presented as mean ± standard deviation (SD). Statistical significance between groups was evaluated using an unpaired Student’s t-test; p < 0.05 was considered significant.
Preparation of L. plantarum Strain for In Vivo Administration
The L. plantarum strain was cultured in MRS broth (BD Difco) and incubated aerobically at 37°C for 24 h. The bacterial stock was preserved as described previously. After incubation, the culture was centrifuged at 4,000 rpm for 20 min. The supernatant was discarded, and the bacterial pellet was resuspended and adjusted to approximately 1 × 10^10^ CFU/ml for use in Model 1 and 1 × 10^8^ CFU/ml for Model 2. To determine bacterial viability, the L. plantarum suspension stored in saline was cultured on MRS agar (BD Difco) plates under aerobic conditions after one week, and colony counts were performed after 48 h of incubation. For in vivo administration, mice in the treatment group received L. plantarum via oral gavage in Model 1 (200 μl per mouse), while in Model 2, the probiotic was administered intranasally via nebulization (10 ml total per treatment for each mouse). Control group mice in both models received only an equivalent volume of sterile saline by oral gavage, as in the treatment groups.
Animal Ethics Approval and Experimental Conditions
All mouse experiments in this study were conducted per the guidelines of the Institutional Animal Ethics Committee of Soonchunhyang University (Republic of Korea), under the committee’s authorization (IACUC approval number: SCH19−0053).
Nine-week-old specific pathogen-free (SPF) female BALB/c mice (Doo Yeol Biotech, Republic of Korea) were housed in individual cages and kept under standard conditions, including a room temperature (RT) of (22−24)°C with a 12 h light/dark cycle and (30−70)% humidity in an air-conditioned animal room at (23 ± 2)°C. All mice were given a one-week adjustment period prior to the experiment, and free access to food and water ad libitum.
The study included two experimental mouse models. In Model 1, mice were randomly assigned to three groups with n = 6 per group, whereas in Model 2, they were assigned to three groups with n = 12 per group. Model 1 was designed to assess the oral preventive effect of L. plantarum and its anti-infective activity during bacterial colonization, while Model 2 evaluated its nasal efficacy following infection. All experimental procedures were performed in a biosafety level 2 (BSL−2) facility at the HM−MRC animal room of Soonchunhyang University based on World Health Organization (WHO) recommendations, and the guidelines of the Ministry of Food and Drug Safety (Registration, MFDS, No. 657).
Experimental Design to Evaluate the Potential of L. plantarum in CRKP-Infected Mouse Models
To evaluate the therapeutic potential of the probiotic candidate against CRKP, two infection mouse models were conducted in this research: an oral preventive model and a nasal efficacy model. In both models, mice were randomly divided into three groups: a control group, an infection group, and a treatment group. To induce neutropenia, cyclophosphamide (450 mg/kg; 200 μl/mouse, Sigma-Aldrich) was administered intraperitoneally three days (d) before infection in both models. Additionally, gut dysbiosis was induced by oral administration of vancomycin (100 mg/kg; 200 μl/mouse) on days (D) (−2 and −1). Before CRKP infection, all mice were given an oral dose of 200 μl of 0.2 M sodium bicarbonate (Sigma-Aldrich) to neutralize gastric acid and facilitate bacterial colonization. After bicarbonate administration, the mice immediately received CRKP via the same oral route. In the treatment groups, mice received approximately 1 × 10^10^ CFU/ml of L. plantarum via oral gavage in the preventive model and approximately 1 × 10^8^ CFU/ml intranasally via nebulization in the efficacy model. Control groups in both models received only sterile saline orally. To assess the effect of L. plantarum on decreasing CRKP infection, body weight changes and illness severity scores were recorded throughout the experiments. For further analysis, in Model 1, fecal samples were collected from individual mice on D (0, 2, 4, 8, and 11), and intestinal tissues were dissected on D 7. In Model 2, stool samples were collected on D (0, 2, 4, 8, and 11), while tissue samples (Lung, Kidney, and Liver) were gathered on D (0, 4, and 10). CFU per gram of samples was determined by plating each sample onto MacConkey agar containing 10 μg/ml of imipenem (~100 mg diluted in 1 ml of NaCl). In addition, Western Blot analysis was performed on intestinal tissues collected from Model 1 to determine the gut integrity. All experiments were replicated at least twice under equal conditions.
Determining the Oral Preventive Impact of Probiotic Strain in the CRKP-Infected Mice Model
We investigated the preventive effect of L. plantarum against CRKP infection in the BALB/c mouse model. In this study, mice in the treatment group received L. plantarum at a concentration of 1 × 10^10^ CFU/ml in a 200 μl volume via oral gavage 3 d before the first CRKP infection (D (−3, −2, −1)), as well as on D (0, 2, 6, 7, and 9) post-infection. Vancomycin was administered orally for dysbiosis at a dose of 100 mg/kg (200 μl/mouse) on D (−2 and -1). Mice received 200 μl of CRKP orally at a concentration of 1 × 10^10^ CFU/mouse on D 0, following oral bicarbonate administration to neutralize gastric acid. Subsequent CRKP challenges were performed on D (2 and 7) using 1 × 10^9^ CFU/mouse. Fecal samples were collected individually from mice on D (1, 3, 5, 7, 8, and 9) post-infection to analyze bacterial colonization. Colony-forming units per gram of stool were determined by plating the samples on MacConkey agar with 10 μg/ml of imipenem (prepared by diluting ~100 mg in 1 ml of NaCl). The number of viable CRKP colonies was assessed, expressed as log_10_ CFU/g of stool. On D 7, mice from each group were randomly selected. The small intestines were cut open and preserved for further analysis. To assess the overall effect of L. plantarum, the mice were observed for 2 weeks following the initial infection, to monitor survival rates, illness severity scores, and body weight changes.
Investigating the Nebulization Efficacy of the Probiotic Candidate in a CRKP-infected Mouse Model
To evaluate the therapeutic potential of L. plantarum via nebulization, CRKP infection was induced in mice by orally administering 200 μl of a CRKP suspension (1 × 10^9^ CFU/mouse) along with sodium bicarbonate on D (0 and 2). During the experiment, mice in the treatment group received L. plantarum intranasally at a concentration of 1 × 10^8^ CFU/mouse, using a nebulizer for 20 min each time. To assess treatment outcomes, biological samples, including stool and tissues, were collected at designated time points. CFU per gram was quantified by plating stool, lung, liver, and kidney samples on MacConkey agar supplemented with 10 μg/mL imipenem. These analyses were performed to evaluate bacterial load and the potential of L. plantarum to reduce CRKP colonization in both intestinal and systemic organs.
Isolation of Mouse Intestinal Tissue and Protein Extraction
The integrity of the intestinal epithelium in mice was analyzed during PMC105 using a Western blot assay, following previously established protocols [42, 43]. Small intestinal tissues from mice were collected on D 7 in the preventive mouse model, flushed with phosphate-buffered saline (PBS), and cut into small fragments. Tissue pieces were homogenized in 1× RIPA lysis buffer (Abcam, UK) with a Protease Inhibitor Cocktail (Cell Signaling Technology, USA), and centrifuged at 14,000 rpm at 4 °C for 15 min. The concentration of the extracted proteins was determined using the Bicinchoninic Acid assay kit (KeyGEN BioTECH, China). After protein extraction, the samples were quantified and transferred onto polyvinylidene difluoride (PVDF) membranes. The membranes were then incubated with primary antibodies, including zonula occludens-1 (ZO−1) (Protein Tech, USA) and β–actin (Cell Signaling Technologies), followed by incubation with a horseradish peroxidase-conjugated secondary antibody (Cell Signaling Technologies). Protein bands were visualized using a ChemiDoc XRS system (Bio-Rad), and densitometric analysis was performed using ImageJ software (version 1.8.0).
Protein Identification and Western Blot Analysis
The intestinal tissues were homogenized in a RIPA lysis buffer (Abcam, UK), supplemented with a protease inhibitor cocktail (Cell Signaling Technology), followed by centrifugation at 14,000 rpm for 15 min at 4°C. The supernatant was collected, and after protein extraction for Western blot analysis, 10 μg of protein solution was mixed with sodium dodecyl sulfate (SDS) sample buffer (KeyGEN BioTECH) and denatured by heating at 100°C for 5 min in a ThermoMixer C (Germany). Proteins were separated on a (10−20) % Tris–glycine gel (BioRad), and transferred to polyvinylidene difluoride (PVDF) membranes (Immuno–Blot PVDF, BioRad) using the TransBlot system (BioRad) for 10 min. The nonspecific binding sites of membranes were blocked using 5% nonfat milk in Tris–buffered Saline with Tween 20 (TBST) at RT for 2.5 h, followed by overnight incubation at 4°C with primary antibodies: mouse β–actin (Cell Signaling Technology) as a loading control, and ZO−1 Rabbit Polyclonal antibody (Protein Tech). Subsequently, the membranes were washed three times with TBST buffer (5 min each), and incubated with the Horseradish Peroxidase (HRP) conjugated secondary antibody (Cell Signaling Technology) for 2.5 h at RT. The membranes were again washed (5 min, 3 times), and the appropriate amount of enhanced chemiluminescence reagent was added to visualize the protein bands. The Molecular Imager ChemiDoc Imaging System (Bio-Rad) was used to detect the bands, with image analysis being conducted using Image Lab and ImageJ software.
Acute Toxicity of Probiotic Candidate
To evaluate the oral acute toxicity of the probiotic strain, a two-week experiment was conducted using nine-week-old female BALB/c mice, following approval by the Institutional Animal Care and Use Committee (IACUC approval number: SCH25−0026). The mice were randomly divided into two groups (6 mice per group). One group received saline drinking water (control group), while the other received L. plantarum orally at a concentration of 1 × 10^10^ CFU/mouse (200 μl per mouse). Mice were housed individually under a 12 h light/dark cycle, with controlled temperature at (20−25)°C, and relative humidity of (30−70)%. The mice were monitored and recorded over a 14 d period for any signs of clinical symptoms, mortality, or changes in body weight. This experiment followed the guidelines outlined in the OECD Test Guideline 423, with minor modifications to assess the acute oral toxicity of L. plantarum in the mouse model [44].
Safety Assessment of the Probiotic Candidate According to MFDS Guidelines
The Joint FAO/WHO Working Group introduced standardized safety criteria for probiotics in 2002 [45], which were then adopted by the Korean Ministry of Food and Drug Safety (MFDS) in 2021 [46]. To assess the safety of the probiotic candidate per these guidelines, antimicrobial susceptibility (E–test), β–hemolysis activity, bile salt deconjugation, D–lactate production, and cell cytotoxicity were evaluated.
An antibiotic strip (Liofilchem, Italy) was used to assess antibiotic susceptibility using the E–test method [47]. As advised by the European Food Safety Authority (EFSA) [48], nine antibiotics were used: ampicillin, vancomycin, gentamicin, kanamycin, streptomycin, erythromycin, clindamycin, tetracycline, and chloramphenicol. After the strips were placed in agar plates and inoculated with the strains, they were incubated for 48 h at 37°C under anaerobic conditions. The measured minimum inhibitory concentration (MIC) was compared to the MIC cut-off criteria of the European Food Safety Authority (EFSA) [49].
The hemolytic activity of the probiotic strain was assessed using blood agar plates (Kisan Bio, Republic of Korea). After the strains were streaked onto agar plates and incubated for 48 h under anaerobic conditions at 37°C, hemolytic activity was assessed based on the following observations: A small area of the medium with a discolored zone ranging from green to brown indicates alpha (α) hemolysis, which shows a reduction of hemoglobin to diffuse methemoglobin. Complete lysis of red blood cells is indicated by a clear, pale–yellow zone surrounding the colonies, which is known as beta (β) hemolysis, whereas gamma (γ) hemolysis is characterized by the absence of any visible change in the medium [50].
The bile salt deconjugation activity of the strains was assessed using a plate assay with Taurodeoxycholic acid sodium salt (TDCA, Millipore, Germany) as the substrate [51]. TDCA−MRS agar (BD Difco) plates were prepared with a sodium salt concentration of 1 mM/L. Probiotic strains were streaked onto TDCA-enriched agar plates and incubated under anaerobic conditions at 37°C for 5 d. The negative and positive controls for bile salt deconjugation were Lacticaseibacillus rhamnosus KCTC 5033 and L. plantarum KCTC 3105 (KCTC, Korea), respectively.
The D–lactate assay was used to evaluate excessive D–lactate production, which, due to the potential risk of D–lactic acidosis, is a critical safety consideration [52]. D–lactate production by the candidate probiotic strains was assessed after incubation at 37°C for 24 h in MRS broth. After incubation, the culture supernatants were collected, and D–lactate concentrations were measured using an assay kit (Abcam) according to the manufacturer’s instructions. A standard D–lactate solution was prepared for reference, and absorbance was measured using a Victor Nivo microplate reader (VICTOR Nivo, PerkinElmer Instruments, USA). L. rhamnosus KCTC 5033 was used as a negative control (NC) for D–lactate production.
HT29 cells were seeded at 1 × 10^4^ cells/well in a 96-well plate in DMEM (Gibco, 10313021) containing 10% FBS (Welgene, S101−07) and 1% penicillin/streptomycin (Gibco, 15140), and cultured at 37°C, 5% CO_2_ incubator for 24 h. After washing the cultured strain with PBS, several CFU ((10^7^−10^9^) CFU) of bacteria were inoculated into 1% BSA (Sigma, A7906)-washed cells using DMEM (2% FBS and no antibiotics) for 24 h. The release of lactate dehydrogenase (LDH) into the medium was measured using the cytotoxicity detection kit (Roche, 4744926001) according to the manufacturer’s protocol. E. coli NCCP 14538 was purchased from the National Culture Collection for Pathogens (Republic of Korea) and used as the positive control, producing toxicity. The absorbance at 490 nm was measured using a microplate reader (VICTOR Nivo, PerkinElmer Instruments) and then converted to cytotoxicity (%). All safety experiments were conducted in accordance with the probiotic safety evaluation standards of the Korean Ministry of Food and Drug Safety (KMFDS) [53].
Results
16S rRNA Gene Sequence Analysis of the Probiotic Candidate
The candidate strain was identified by 16S rRNA gene sequencing, and the resulting sequences were compared with sequences in the NCBI database (Table 1). According to the analysis, the isolate’s sequence showed 99% similarity with strains of L. plantarum, including DSM 20174, NBRC 15891, CIP 103151, and NRRL B−14768. Furthermore, the sequence showed a high degree of similarity to other members of the Lactobacilli. The 16S rRNA gene sequence of the probiotic candidate generally exhibited (98−99)% sequence identity to reference sequences in the database.
Whole-Genome Analysis Results of the Candidate Strain
The assembled genome comprised a single circular chromosome of 3,276,265 base pairs, harboring 3,054 coding sequences (CDSs). Among these, 2,728 predicted protein-coding genes were categorized based on the functional classification of clusters of orthologous groups (COG) (Fig. 1A). Comparative genomic analysis using the Orthologous Average Nucleotide Identity (OrthoANI) approach revealed high genomic similarity with L. plantarum, while exhibiting lower similarity to other species within the Lactiplantibacillus genus, confirming its taxonomic identity as L. plantarum (Fig. 1B). In contrast, the L. plantarum isolate exhibited distinct genetic features compared to previously documented strains, supporting its designation as a novel strain, subsequently named PMC105 (Table 2).
Determination of the Minimum Inhibitory Concentration of the Strain’s CFS
Fig. 2A shows the results of the Minimum Inhibitory Concentration (MIC). Optical density (OD) was measured at 570 nm using a microplate reader to assess bacterial growth in each well. The MIC was determined as the lowest CFS concentration from L. plantarum that completely inhibited the growth of CRKP. It was found to be 125 μl/ml. Subsequently, the contents of the wells that showed no growth were cultured on fresh MacConkey agar plates. These results indicate the significant antimicrobial effects of L. plantarum CFS against CRKP, warranting further investigation and potential therapeutic application.
Time-kill Assay of CFS of L. plantarum Strain against CRKP Infection
The time-kill assay illustrated that the CFS of L. plantarum PMC105 showed a strong bactericidal effect against CRKP. A progressive reduction in bacterial viability was observed over time in the presence of L. plantarum CFCS, with complete inhibition of CRKP growth observed after 1.25 h of incubation, demonstrating the kill effect of L. plantarum PMC105 CFS against CRKP. On the other hand, bacterial levels in the control group (CRKP in MRS broth without CFS) were progressively increased throughout the assay. Notably, a ≥3 log_10_ CFU/ml decrease in CRKP counts was recorded for 8 h in samples treated with L. plantarum CFS, indicating significant time-dependent bactericidal activity. These findings illustrate that L. plantarum PMC105 CFS can significantly inhibit the growth and viability of CRKP under in vitro conditions (Fig. 2B).
Therapeutic Impact of L. plantarum in a Preventive Mouse Model of CRKP
The study assessed the therapeutic effects of L. plantarum on lethal CRKP infections in a mouse model. Mice were given L. plantarum at a concentration of 1 × 10^10^ CFU/ml, before oral infection with CRKP (Fig. 3A). The untreated group experienced more severe weight loss compared to the treated group, with body weights significantly lower by D (14 and 16) (p < 0.05) (Fig. 3B). From D 2 onwards, the CRKP growth rate decreased in the group treated with L. plantarum. The mice were monitored for two weeks, with CFUs measured throughout. By D 14, CRKP had reduced the viable counts in the treated group (p < 0.01). Stool samples from the pre-treated mice showed lower CRKP colony numbers compared to those from other groups until the study’s conclusion. Consequently, significant differences in CFU counts were observed between the pre-treated group (prior to L. plantarum treatment) and the untreated group (Fig. 3C). After oral infection, the untreated mice exhibited symptoms such as diarrhea and fatigue. There were considerable differences in illness severity scores between the treated and untreated groups during the study (p < 0.001) (Fig. 3D). All mice treated with L. plantarum survived 14 d post-infection, whereas 66.7% of the mice infected with CRKP in the untreated group died during the study (Fig. 3E). In addition, the Western blot assay of small intestine on D 7 illustrated a noticeable reduction in ZO−1 levels in the infection group that significantly improved gut barrier integrity by reducing intestinal permeability by PMC105 treatment rather than the treatment group, which is evidenced by the high level of tight junction protein ZO−1 in the treated mice group (Fig. 3F).
Nebulization Potential of L. plantarum on Klebsiella pneumoniae Induced in a Mouse Model
Mice were given L. plantarum intranasally at a concentration of 1 × 10^8^ CFU/ml for 20 min, before oral infection with CRKP (Fig. 4A). 60% of the infected mice in the untreated group with CRKP survived during the experiment. No mortality was observed in the nebulization treatment group of mice treated with L. plantarum (Fig. 4B). Following oral infection, untreated mice exhibited symptoms such as diarrhea and weakness. Significant differences in illness scores were observed between the treated and untreated groups from D (4 to 10) (p < 0.001) (Fig. 4C). The untreated group illustrated greater weight loss compared to the treated group, with their body weight significantly reduced on D (14 and 16) (p < 0.05) (Fig. 4D). A microbiological assessment of the normal intestinal microflora in mice was conducted by culturing fecal samples from uninfected mice on MacConkey agar plates containing 10 mg/ml imipenem. After 24 h of incubation, no bacterial growth was detected on the plates for any of the fecal samples, indicating the absence of CRE strains. The CFU of collected samples (stool, liver, kidney, and lung) was measured during the study. The treated group showed notable differences in CFU counts compared to the untreated group. CRKP reduced viable colony counts in the treated group, rather than in the untreated group, by D 10 (p < 0.01) (Fig. 4E−4H). The results demonstrated that although CRKP can colonize the intestines of mice, Nebulization with L. plantarum PMC105 reduced the bacterial burden and showed a trend toward improved survival compared with the CRKP-infected group.
Acute Oral Toxicity of the Investigated L. plantarum
An acute toxicity study of L. plantarum was conducted in a mouse model to assess potential side effects of the strain. Nine-week-old mice were administered L. plantarum at the maximum achievable dose levels (200 μl per mouse). Mortality, general appearance, and body weight were monitored for 14 d. No abnormal clinical signs or significant differences in body weight were observed between the mice treated with L. plantarum and those receiving sterile water. All mice survived throughout the experiment, and no adverse effects or mortality were observed in any group (Fig. 5).
Comprehensive Safety Assessment of the Probiotic Candidate
Fig. 6A and 6B demonstrate the profiles of antibiotic susceptibility of the probiotic strain. The L. plantarum PMC105 was susceptible to clindamycin, tetracycline, and ampicillin, but tolerant to gentamicin, kanamycin, erythromycin, and chloramphenicol.
Bacterial infections, including invasive infections, can often lead to hemolytic symptoms, such as anemia, fever, and skin rashes [54]. Thus, investigating the hemolytic activity of probiotics is crucial to confirm their safety. The hemolytic activity of L. plantarum PMC105 was assessed using blood agar plates by monitoring the presence or absence of clear (β–hemolysis), greenish (α–hemolysis), or no zones (γ–hemolysis) around the bacterial colonies after incubation. In the current study, the probiotic strain did not produce any clear or greenish zones around its colonies, indicating γ–hemolytic activity (Fig. 6C). This demonstrates L. plantarum PMC105 did not lyse red blood cells (RBCs) in the examination, confirming its non-hemolytic nature.
Fig. 6D illustrates the D–lactate production results, with concentrations expressed in mmol/μl. L. rhamnosus KCTC3237 and L. plantarum PMC105 were evaluated using the D–lactate kit. When the CFU of L. plantarum PMC105 gradually increased, the level of D–lactate production increased. This observation is notable, as elevated D–lactate levels can cause D–lactic acidosis, posing potential safety concerns in probiotic applications. Conversely, the minimal or undetectable D–lactate production in the other CFUs indicates their suitability for probiotic use, minimizing the risk of D–lactic acidosis in clinical conditions. This characteristic is essential to ensure a favorable safety profile during drug development.
In this study, we investigated bile salt deconjugation activity using the standard plate assay method. In agar plates containing taurodeoxycholic acid (TDCA), one of the major conjugated bile salts in the human gastrointestinal tract, overall deconjugation was observed by strains PMC 105, as well as control strains L. rhamnosus KCTC3237 and L. plantarum KCTC3105 (Fig. 6E). The bile salt deconjugation activity is an important property of probiotics, as it promotes their survival in the gastrointestinal tract by improving bile salt tolerance, an essential factor to maintain their viability and functionality in the challenging intestinal environment.
Fig. 6F and 6G show the cytotoxicity of L. plantarum PMC105, as measured by lactate dehydrogenase (LDH). The results showed that L. plantarum PMC105 has no cytotoxic effect on the HT29 cell line.
Discussion
The opportunistic pathogen CRKP, which is resistant to a wide range of antibiotics, poses a significant threat. The World Health Organization lists CRKP as a critical priority antibiotic-resistant pathogen for which new treatments are urgently needed [55]. Previous research has shown that by modulating the immune system and maintaining intestinal cell homeostasis, metabolites and functional factors from the gut microbiota strongly impact the host gastrointestinal tract [56]. Probiotics are promising candidates for treating multidrug-resistant infections.
L. plantarum is known as a potential antimicrobial probiotic against a broad range of gastrointestinal pathogens, such as Es. coli and S. aureus, with different mechanisms that include the production of bacteriocins, immune system modulation, production of short fatty acids, and maintenance of the intestinal barrier integrity [57]. In contrast, Listeria monocytogenes is known for its antibacterial metabolites [58]. In this study, L. plantarum PMC105, isolated from a traditional Korean fermented kimchi, was evaluated for its potential to prevent or suppress CRKP infection, focusing on its safety profile, effects on intestinal barrier function, and clinical outcomes in mouse models.
Several studies have proven the tolerance of L. plantarum in gastrointestinal conditions. L. plantarum is resistant to low pH conditions and can ferment lactic and deconjugate bile salts. Lactic acid fermentation helps L. plantarum survive in a gut-acidic environment while reducing the risk of metabolic acidosis, thereby improving gut health [59].
In this study, the oral preventive and intranasal nebulization models were utilized to evaluate the therapeutic efficacy of L. plantarum PMC105 against CRKP infection in immunocompromised mice. The preventive model, which involved oral administration of L. plantarum prior to CRKP infection, aimed to inhibit initial colonization and improve gut barrier function. In contrast, the nebulization model assessed the strain’s therapeutic potential post-infection, targeting reductions in systemic bacterial burden and illness severity. The preventive model showed strong protective potential, with treated mice showing significantly reduced illness severity, decreased CRKP colonization in the gut, improved maintenance of body weight, and 100% survival, compared to a 60%mortality rate in the untreated group. In addition, PMC105 maintained intestinal barrier integrity, as indicated by preserved ZO−1 protein expression levels in Western blot assays. This indicates that L. plantarum improves the intestinal barrier function and supports the local immune response prior to infection. Although the nebulization model was applied after CRKP infection, it still demonstrated promising outcomes. The intranasal delivery of L. plantarum resulted in reduced bacterial loads in stool and organs, enhanced clinical condition, and improved survival rate. These findings indicate that even after pathogen colonization, L. plantarum may provide systemic protective effects, potentially by modulating mucosal immune responses or inhibiting bacterial spread. Comparison between the two models showed that oral treatment achieved a higher survival rate, suggesting a more effective role in blocking CRKP colonization and infection progression [60, 61].
Statistical analysis of the oral preventive mouse model showed that L. plantarum decreased the average illness severity score by ~66.7%, and increased body weight by approximately 230% by D 16, compared to the untreated group. All treated mice survived in the preventive model study; however, the nebulization-treated group demonstrated 91.66% survival compared with untreated mice by 58.33% of survival rate. Furthermore, to compare with a previous study using Bacillus velezensis, which reported that 80% of mice survived, L. plantarum PMC105 demonstrated greater preventive efficacy against CRKP infection [33].
Our findings also support the anti-colonization potential of L. plantarum. While L. fermentum has been shown to reduce CRKP intestinal colonization by approximately 27% [33], L. plantarum in our study achieved a 50%reduction, along with a ~ 66.7% decrease in illness severity, and a decrease in weight loss by about 39%. These outcomes emphasize that, compared to L. fermentum, L. plantarum is more effective at suppressing CRKP colonization in the gastrointestinal tract, making it an eligible candidate for suppressing CRKP infection.
According to previous studies, the intestinal barrier plays a crucial role in maintaining gastrointestinal tract health by inhibiting the translocation of pathogens into the bloodstream. ZO−1, as the tight junction protein, is responsible for the maintenance of intestinal barrier integrity [62]. L. plantarum exerts its microbial activity by up-regulation of the tight junction protein ZO−1 gene expression, and significantly promotes the production of ZO−1 protein [63]. In this study, Western blot was used to analyze ZO−1 expression in mouse models. Our results demonstrated that L. plantarum treatment preserved ZO−1 expression, indicating its preventive effect on CRKP-induced intestinal damage and systemic spread.
Safety evaluation in alignment with international standards, including FAO/WHO and Korean MFDS guidelines [45, 46], confirmed that L. plantarum PMC105 is a non-hemolytic strain, supporting its non-pathogenic nature. The evaluation of toxin production, conducted using in vitro assays, showed no detectable toxin, confirming the strain’s safety. The strain did not produce excessive D–lactate, thereby minimizing the risk of metabolic acidosis, and it also showed low pH tolerance. In addition, its bile salt deconjugation activity indicates its survival in the gastrointestinal environment and its beneficial role in supporting healthy gut microbiota. Moreover, Antibiotic susceptibility testing revealed acceptable resistance profiles in accordance with EFSA guidelines [48]. PMC105 strain demonstrated susceptibility to a wide range of antibiotics, with no transferable resistance genes detected. Other previous studies have also reported the safety profile of L. plantarum, as it is susceptible to Tetracycline, Ampicillin, Vancomycin, Ciprofloxacin, Gentamicin, Azithromycin, and Kanamycin. Also, these studies showed that L. plantarum lacked hemolytic or in vivo toxicological activity [64]. Numerous Lactobacilli are resistant to aminoglycoside antibiotics, such as kanamycin [65, 66], likely due to reduced aminoglycoside uptake in the absence of cytochrome-mediated transport [67, 68]. For example, kanamycin resistance was observed in 79% of 187 isolates from 55 probiotic products in Europe [69]. This observation aligns with EFSA criteria, which indicate that aminoglycoside resistance in Lactobacilli is an intrinsic, non-transferable trait. Moreover, E-test susceptibility testing confirmed that L. plantarum PMC105 meets the safety standards for pharmaceutical applications.
In summary, this study demonstrated the effects of L. plantarum PMC105 on CRKP infection in vivo and in vitro and suggested that it could be a candidate for treating CRKP. This indicates it can be used as a biotherapeutic agent and in combination therapy. However, further studies are needed that focus more on the biological mechanisms underlying the beneficial effects of L. plantarum, including its interactions with the immune system, its impact on the gut microbiome, optimal doses, and possible therapeutic side effects. In addition, clinical trials will be essential to evaluate the efficacy and safety of L. plantarum in individuals affected by CRKP infections.
Conclusion
This research reveals the promising therapeutic advantage of L. plantarum PMC105 in combating CRKP infections. Further, this study provides strong evidence through both preventive and post-infection murine models, supporting the effect of L. plantarum PMC105 as a promising probiotic to prevent and suppress CRKP infections. Its safety profile, ability to maintain intestinal barrier integrity, and efficacy in reducing illness severity make it a promising candidate for further research and possible clinical applications. Importantly, both oral and intranasal delivery of PMC105 resulted in protective benefits, demonstrating flexibility in its administration route for therapeutic purposes. Fig. 7 summarizes these findings, illustrating the reduction in CRKP burden and preservation of organ health following PMC105 treatment. Future work should focus on mechanistic pathways, optimal dosing, and clinical translation to evaluate PMC105 as a scalable probiotic therapeutic candidate.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lei TY Liao BB Yang LR Wang Y Chen XB 2024 Hypervirulent and carbapenem-resistant Klebsiella pneumoniae: a global public health threat Microbiol. Res.28812783910.1016/j.micres.2024.12783939141971 · doi ↗ · pubmed ↗
- 2Chang PC Chen CC Lu YC Lai CC Huang HL Chuang YC 2018 The impact of inoculum size on the activity of cefoperazonesulbactam against multidrug resistant organisms J. Microbiol. Immunol. Infect.5120721310.1016/j.jmii.2017.08.02629037802 · doi ↗ · pubmed ↗
- 3Choby JE Howard-Anderson J Weiss DS 2020 Hypervirulent Klebsiella pneumoniae - clinical and molecular perspectives J. Intern. Med.28728330010.1111/joim.1300731677303 PMC 7057273 · doi ↗ · pubmed ↗
- 4Abbas R Chakkour M Zein El Dine H Obaseki EF Obeid ST Jezzini A 2024 General overview of Klebsiella pneumonia: epidemiology and the role of siderophores in its pathogenicity Biology (Basel)137810.3390/biology 1302007838392297 PMC 10886558 · doi ↗ · pubmed ↗
- 5Vazquez-Ucha JC Arca-Suarez J Bou G Beceiro A 2020 New carbapenemase inhibitors: clearing the way for the beta-lactams Int. J. Mol. Sci.21930810.3390/ijms 2123930833291334 PMC 7731173 · doi ↗ · pubmed ↗
- 6Abdel-Halim MS Askoura M Mansour B Yahya G El-Ganiny AM 2022 In vitro activity of celastrol in combination with thymol against carbapenem-resistant Klebsiella pneumoniae isolates J. Antibiot (Tokyo)7567969010.1038/s 41429-022-00566-y 36167781 PMC 9640353 · doi ↗ · pubmed ↗
- 7Do AD Quang HP Phan QK 2025 Probiotic cell-free supernatant as effective antimicrobials against Klebsiella pneumoniae and reduce antibiotic resistance development Int. Microbiol.2862363210.1007/s 10123-024-00575-x 39117894 · doi ↗ · pubmed ↗
- 8Hill C Guarner F Reid G Gibson GR Merenstein DJ Pot B 2014 Expert consensus document. The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic Nat. Rev. Gastroenterol. Hepatol.1150651410.1038/nrgastro.2014.6624912386 · doi ↗ · pubmed ↗