Ecology and Diversity of Urban Drosophila Species Communities as Potential Indicators of Biodiversity Decline
Martin Kapun, Sonja Steindl, Maria Ricci, Manuel Löhnertz, Flora Strasser, Rui Qiang Chen, Lorin Timaeus, Nikolaus Szucsich, Elisabeth Haring

TL;DR
This study shows that urban fruit fly communities in Vienna have declined significantly over 30 years, with new invasive species replacing native ones, suggesting broader biodiversity loss in cities.
Contribution
The study provides novel evidence of biodiversity decline in urban areas using Drosophila as an indicator and identifies two new species records for Austria.
Findings
Species richness in Vienna's Drosophila communities has declined by over 50% compared to 30 years ago.
Non-indigenous species like D. mercatorum are replacing formerly common native species in urban areas.
D. mercatorum is highly synanthropic, thriving in urban areas with high impervious surfaces.
Abstract
Understanding the impact of ecological factors on biodiversity is central in the context of accelerating climate change and biodiversity loss. Urban areas, as landscapes under particularly strong anthropogenic influence, are undergoing rapid ecological change, yet the consequences for urban biodiversity and ecosystem functioning remain poorly understood. In this study, we focused on fruit flies of the genus Drosophila—a diverse group of dipterans with variable ecological niches and degrees of synanthropy, i.e., the adaptation to human‐modified habitats. We investigated species composition and community ecology in the metropolitan area of Vienna, Austria. With the help of numerous citizen scientists, we have collected approximately 18,000 specimens through dense spatio‐temporal sampling both indoors and outdoors of human dwellings. A total of 13 Drosophila species were identified, with…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
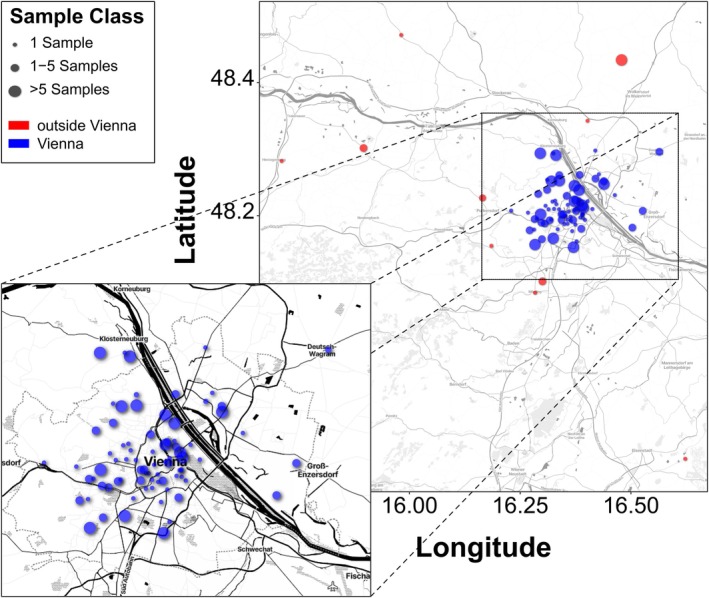 FIGURE 1
FIGURE 1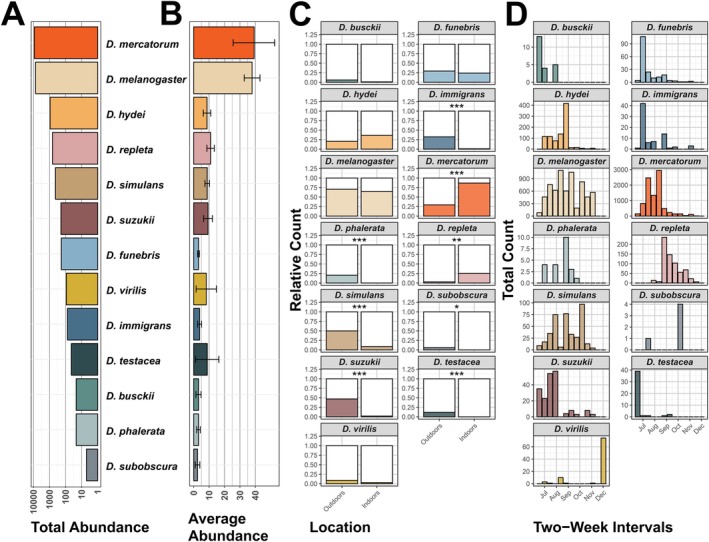 FIGURE 2
FIGURE 2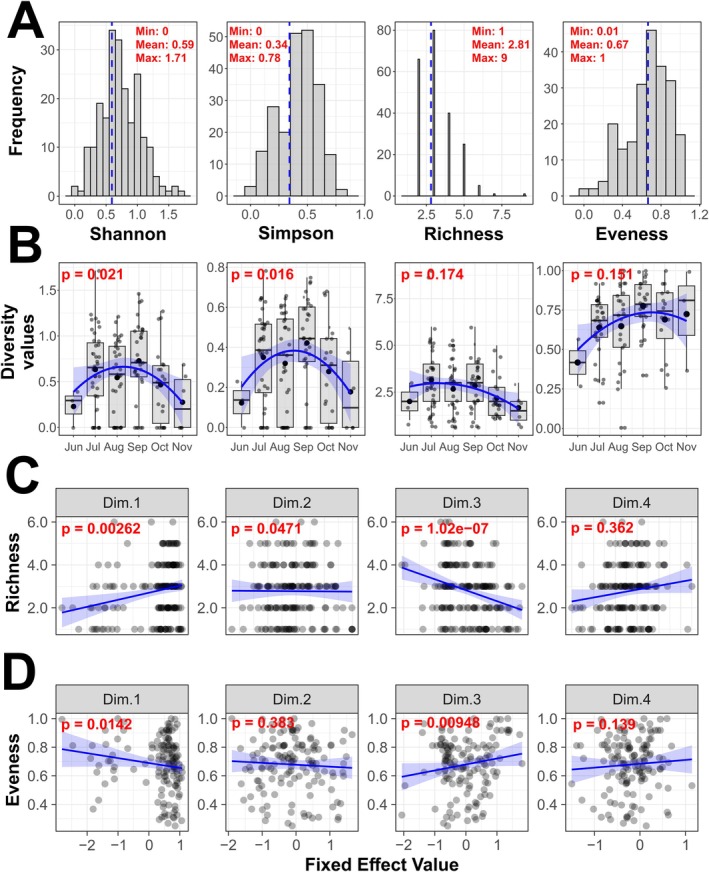 FIGURE 3
FIGURE 3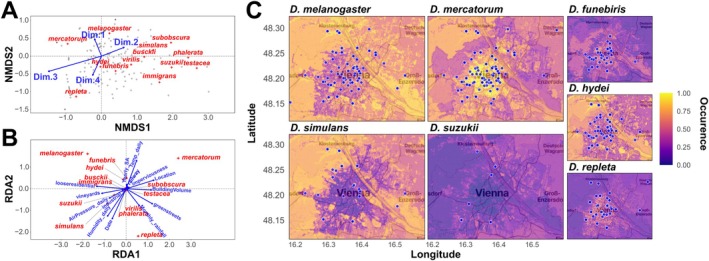 FIGURE 4
FIGURE 4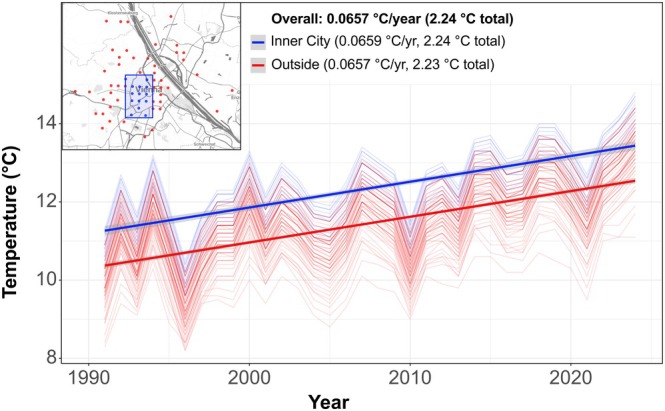 FIGURE 5
FIGURE 5| Subgenus | Species group | Species | Total abundance | Sequenced |
|---|---|---|---|---|
|
|
|
| 185 | 5 |
|
|
| 76 | 5 | |
|
|
| 22 | 3 | |
|
|
| 908 | 6 | |
|
|
| 656 | 3 | |
|
| 8851 | 6 | ||
|
|
| 44 | 3 | |
|
|
| 90 | 3 | |
|
|
|
| 6726 | 3 |
|
| 393 | 5 | ||
|
| 195 | 3 | ||
|
|
| 5 | 4 | |
|
|
| 22 | 4 |
| Species |
|
| Odds ratio (OR): In vs. Out |
|---|---|---|---|
|
| 34.93 |
| 0 (0) |
|
| 5.81 |
| 0 (0) |
|
| 17.09 |
| 0 (0) |
|
| 47.63 |
| 0.022 (45.455) |
|
| 68.40 |
| 0.023 (43.478) |
|
| 36.07 |
| 0.095 (10.526) |
|
| 1.67 | 1.97E‐01 | 0.165 (6.061) |
|
| 1.26 | 2.62E‐01 | 0.326 (3.067) |
|
| 0.21 | 6.44E‐01 | 0.757 (1.321) |
|
| 0.21 | 6.45E‐01 | 0.767 (1.304) |
|
| 2.50 | 1.14E‐01 | 2.191 (0.456) |
|
| 7.29 |
| 11.301 (0.088) |
|
| 52.64 |
| 15.692 (0.064) |
| Model | Statistic | Dataset |
|
|
|
|
|
|
|
|---|---|---|---|---|---|---|---|---|---|
| RF | AUC | training | 0.845 | 0.849 | 0.916 | 0.843 | 0.87 | 0.835 | 0.895 |
| test | 0.729 | 0.822 | 0.869 | 0.445 | 0.833 | 0.569 | 0.746 | ||
| Kappa | test | 0.382 | 0.628 | 0.598 | −0.04 | 0.622 | 0.175 | 0.473 | |
| Overfitting | 0.116 | 0.028 | 0.048 | 0.398 | 0.037 | 0.267 | 0.149 | ||
| GLM | AUC | training | 0.819 | 0.628 | 0.926 | 0.821 | 0.638 | 0.645 | 0.703 |
| test | 0.703 | 0.508 | 0.757 | 0.464 | 0.684 | 0.547 | 0.479 | ||
| Kappa | test | 0.382 | 0.095 | 0.473 | −0.066 | 0.402 | 0.096 | 0.022 | |
| Overfitting | 0.115 | 0.119 | 0.169 | 0.356 | −0.046 | 0.098 | 0.224 | ||
| GAM | AUC | training | 0.714 | 0.834 | 0.827 | 0.73 | 0.83 | 0.824 | 0.781 |
| test | 0.712 | 0.754 | 0.832 | 0.58 | 0.821 | 0.584 | 0.785 | ||
| Kappa | test | 0.452 | 0.458 | 0.393 | −0.239 | 0.526 | 0.086 | 0.61 | |
| Overfitting | 0.002 | 0.08 | −0.005 | 0.15 | 0.009 | 0.24 | −0.004 | ||
| MaxEnt | AUC | training | 0.762 | 0.882 | 0.913 | 0.833 | 0.735 | 0.858 | 0.918 |
| test | 0.61 | 0.749 | 0.792 | 0.448 | 0.519 | 0.612 | 0.735 | ||
| Kappa | test | 0.483 | 0.478 | 0.473 | −0.04 | 0.316 | 0.168 | 0.425 | |
| Overfitting | 0.152 | 0.132 | 0.121 | 0.384 | 0.217 | 0.247 | 0.183 |
- —HORIZON EUROPE Framework Programme10.13039/100018693
- —Natural History Museum10.13039/501100000831
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect behavior and control techniques · Diptera species taxonomy and behavior · Forensic Entomology and Diptera Studies
Introduction
1
The genus Drosophila comprises over 1600 described species of small dipterans, exhibiting an exceptionally wide range of ecological niches, behaviors, and morphologies (Bächli 1982; O'Grady and DeSalle 2018). Their ecological strategies range from specialists, often restricted to narrow geographic areas and specific food sources, to opportunistic generalists with broad—sometimes global—distributions (Bächli and Burla 1985; Bächli and Pite 1982; Brake and Bächli 2008). A subset of these generalists, characterized by a close association with humans (“synanthropism”), are collectively referred to as the “cosmopolitan guild” (Atkinson and Shorrocks 1977; Miller et al. 2017; Nunney 1996). These species thrive in highly disturbed environments influenced by human activities and are among the most successful biological colonizers. Accordingly, many Drosophila species exhibit a remarkable expansion potential. The most prominent example is Drosophila melanogaster , one of the best‐studied model organisms in genetics and developmental biology (Bilder and Irvine 2017; Hales et al. 2015; Jennings 2011). Originally, * D. melanogaster was* native to sub‐Saharan Africa (Lachaise et al. 1988). Its ability to adapt to diverse climates, coupled with the reliance on fermenting fruit (Mansourian et al. 2018) and other decaying organic matter, facilitated the early association of this species with human settlements (Haudry et al. 2020; Keller 2007; Markow 2015). The spread of D. melanogaster likely began thousands of years ago alongside the rise of agriculture and the establishment of trade routes (Chen et al. 2024; Kapopoulou et al. 2020). By the late 19th and early 20th centuries, with the rapid expansion of global trade and transport, the species had colonized temperate and tropical regions worldwide (Arguello et al. 2019). Today it occurs on every continent except Antarctica. Several other species have similarly undergone anthropogenic expansion. For example, Drosophila simulans , the sister species of D. melanogaster (Capy and Gibert 2004), Drosophila suzukii, an invasive agricultural pest (Adrion et al. 2014), and Drosophila virilis (Mirol et al. 2008) have achieved near‐global distributions within just a few centuries or even decades, largely driven by human mobility. Because of their varying ecological specializations and responses to environmental disturbance, Drosophila species can serve as reliable bioindicators for climate change, habitat alteration, and anthropogenic impact (Parsons 1991; Poppe et al. 2013). In particular, community composition often reflects disturbance intensity: with increasing environmental degradation or anthropogenic impact, opportunistic and commensal species become more dominant (Avondet et al. 2003; Shorrocks 1977).
Urban landscapes are extreme examples of anthropogenically altered ecosystems (Vitousek et al. 2008). They are characterized by high habitat heterogeneity, fragmentation, extensive impervious surfaces, air pollution, and pronounced urban heat islands (Marzluff 2008; Szulkin et al. 2020). Green areas as well as nutritional resources are patchy, human‐generated, and often ephemeral, such as organic waste or fermenting fruit in markets, compost bins, or household kitchens (e.g., Hong et al. 2024). These conditions may offer unique opportunities for certain Drosophila species while excluding others. Urban areas might thus act both as refuges for commensal species and as barriers for specialists with narrow ecological tolerances restricted to natural and undisturbed environments. Yet, despite the long‐recognized existence of synanthropic drosophilids, systematic studies of their occurrence in urban environments remain scarce (but see, Avondet et al. 2003; Bombin and Reed 2016; Garcia et al. 2012; Ramniwas et al. 2024; Ulmer et al. 2024). Key ecological questions remain unanswered: Which urban microhabitats do different species exploit? Are urban assemblages dominated exclusively by generalists and hemerophilous species, which prefer or thrive in habitats influenced by humans, or can specialists persist in certain niches? Do species compositions differ among areas subjected to varying degrees of urban pressure, such as city centers versus peri‐urban green spaces? Moreover, species inventories for large cities are rarely updated; for example, the most recent survey of Drosophila in Vienna dates back to 1994 (Gross and Christian 1994). Recent urban Drosophila surveys have been conducted in France (Ulmer et al. 2024), southern Brazil (Hochmüller et al. 2010), and in Moscow (Gornostaev et al. 2024, 2023), but most sampling sites in these studies were relatively natural habitats embedded in urban matrices, rather than heavily built‐up areas. Consequently, we still lack a clear picture of Drosophila ecology within the most anthropogenically altered urban microhabitats—such as inside and around buildings, near waste disposal points, and in transport hubs—where commensal species may reach their highest densities. Furthermore, most previous studies have sampled flies along predefined transects rather than through quantitative random sampling across entire urban areas, potentially limiting the ability to detect general patterns in urban environments.
Even for common species, detailed ecological and biological data, as well as phylogenetic relationships within the genus (Robe et al. 2005), often remain incomplete. While broad geographic occurrence data are available for many species, fine‐scale distributional records—particularly in urban areas—are rare. In Austria, for example, GBIF lists 17 Drosophila species (DOI: 10.15468/dl.22p76q), representing about 60% of the 27 species reported for the country. Of the 157 GBIF records, the vast majority originate from the citizen science platform iNaturalist, with additional contributions from INSDC Sequences, iBOL/BOLD, the Biodiversitätsdatenbank Nationalpark Donau‐Auen, Observation.org, and the Biodiversitätsdatenbank Salzburg. Similar species inventories are found across Central Europe: Switzerland—35 species (Merz and Schweizer Zentrum für die Kartographie der Fauna 1998), Austria—27 (Szucsich, unpubl.; Franz 1989), Germany—30 (AK‐Diptera and Bächli 2023), Czech Republic—29 (Máca 2009; Máca et al. 2015), Slovakia—26 (Máca 2009; https://gd.eppo.int/reporting/article‐3303), Denmark—23, Sweden—26, Norway—19, Finland—27 (Bächli et al. 2004). The present study investigates Drosophila species distributions in urban habitats, with particular emphasis on occurrences inside and around buildings. Conducted within the framework of the citizen science project “Vienna City Fly” (Gottwald et al. 2025), this work aims to document the urban Drosophila species spectrum and to assess their ecological niches. This study is the first to focus predominantly on sampling flies within buildings, employing easily deployable funnel traps. In conjunction with outdoor collections, this approach enables a direct comparison of Drosophila communities between indoor and outdoor microhabitats. Specifically, we address the following questions: (1) Which Drosophila species occur in highly urbanized and rural areas in and around the city of Vienna, and what proportion are generalist versus specialist Drosophila species? (2) How does species composition vary with the degree of urbanization and microhabitat type? (3) Which environmental factors exert the greatest influence on species composition and abundance?
Material and Methods
2
Collection of Flies
2.1
Collections of fly samples were carried out between July and December 2024 as part of a large‐scale citizen science campaign entitled “Vienna City Fly” (Gottwald et al. 2025), which involved 89 citizen science volunteers from the city of Vienna, but also from other Austrian federal states, who caught drosophilid vinegar flies using commercial traps that were placed within kitchens and/or gardens for a maximum of 14 days. The traps consisted of two parts, a translucent plastic container on the outside and a yellow opaque plastic funnel with fine slits at the side to maintain airflow and three holes at the bottom of the funnel which allowed flies to enter the traps. Slices of fruit (e.g., banana or apple), which were placed at the bottom of the plastic container below the funnel, acted as bait and attracted flies into the trap. Behavioral constraints such as negative geotaxis prevented flies from escaping the trap. Caught specimens usually tried to leave the trap at the top of the plastic container, which was sealed by the funnel, and did not find the hole at the bottom of the funnel through which they initially had entered the trap. However, we observed that tiny individuals of species with overall small body size, such as Drosophila melanogaster , occasionally managed to escape through the air vents.
Throughout the sampling season, collectors returned or exchanged filled traps against new ones at the Natural History Museum of Vienna and provided information on collector's ID (referring to the collecting site) and sampling duration (Gottwald et al. 2025). We explicitly requested additional information about the absence of flies, that is, empty traps, and obtained this absence data from several collectors. The filled traps were then placed at −4°C for at least 30 min to anesthetize the flies prior to transferring all insects within a trap to Eppendorf tubes filled with 96% Ethanol. All collected flies were incorporated into the Diptera collection of the NHMW.
Sampling Area—The City of Vienna and Its Vicinity
2.2
We concentrated our sampling efforts in Vienna, a metropolitan area encompassing approximately 415 km^2^ and comprising over 176,000 buildings (Bauer et al. 2024). The urban landscape included in our sampling is predominantly characterized by multi‐family apartment buildings, including numerous public housing complexes, while single‐family homes are more commonly found in peripheral districts. Although more than half of the total area of Vienna is classified as green space—including parks, forests, vineyards, and urban gardens—these are largely concentrated in the outer districts. The study primarily targeted the urbanized zones of the city, which present a highly heterogeneous mosaic of environments, ranging from densely built‐up and impervious areas to green spaces and low‐density residential neighborhoods. We also included samples collected in suburban or rural areas outside of Vienna, in the neighboring regions of Lower Austria and Burgenland, which are predominantly farmland and characterized by a humid continental climate (Köppen classification Dfb and Cfb; Köppen 2011).
Species Identification
2.3
All sampled drosophilid flies were identified to species level under a light microscope following the identification key of Bächli and Burla (1985). We furthermore used additional literature to specifically distinguish species of the repleta group (Beuk 1993). All identified specimens were then sorted, counted, and stored in 96% Ethanol at 4°C for each trap and each species separately.
DNA Barcoding
2.4
For each Drosophila species, we further characterized several specimens from various localities by DNA barcoding using the mitochondrial (mt) cytochrome c oxidase subunit 1 gene (COX1) standard marker (Folmer et al. 1994). DNA extraction was performed in the DNA clean room of the Natural History Museum Vienna under strict contamination control protocols. The DNeasy Blood & Tissue Kit (Qiagen) was used for DNA extraction, following the manufacturer's instructions, with a final elution volume of 50 μL. A negative control extraction was carried out without a DNA sample to detect potential contamination in the extraction reagents. These control extractions were subsequently included in the PCR analyses. All post‐extraction work (thermocycling and post‐PCR processing) was performed in a separate laboratory.
A partial region of the mitochondrial cytochrome c oxidase subunit 1 gene (COX1) was amplified using the PCR primers LCO1490 5′‐GGTCAACAAATCATAAAGATATTGG‐3′ and HCO2198 5′‐TAAACTTCAGGGTGACCAAAAAATCA‐3′ (Folmer et al. 1994) resulting in an amplicon 709 bp in length that was used as a genetic marker sequence (alignment length 658 bp). PCR reactions were conducted using the Multiplex PCR Kit (Qiagen, Hilden, Germany) in 25 μL reaction volumes containing 12.5 μL of Multiplex PCR Master Mix, 0.5 μM of each primer, and 2 μL of template DNA. The thermocycling conditions were as follows:for all reactions: an initial denaturation at 94°C for 15 min, followed by 40 cycles of 94°C for 30 s, 52°C for 30 s, and 72°C for 30 s, with a final extension at 72°C for 10 min. Negative controls were included in all PCRs: one PCR reaction using the control extraction as a template and a PCR reaction with template DNA. PCR products were purified using the QIAGEN PCR Purification Kit and sequenced by SANGER sequencing in both directions using the original PCR primers (at Microsynth Austria).
The generated DNA barcodes contribute to the Austrian Barcode of Life (ABOL) initiative dedicated to recording Austrian biodiversity. DNA barcodes can be found at the Barcode of Life database BOLD (https://v5.boldsystems.org) under the accession numbers listed in Table S1.
Environmental Variables
2.5
To identify potential links between Drosophila community composition and environmental factors, we obtained gridded climatic data from the data hub of GeoSphere Austria (https://data.hub.geosphere.at/) and demographic and administrative data from the data repository provided by the city of Vienna (https://www.data.gv.at/). Climatic data were obtained from two different sources: The SPARTACUS v. 2.0 dataset (Hiebl and Frei 2016) from GeoSphere Austria consists of yearly (average temperature [TA; °C], total rainfall [RR; mm/m^2^] and total sunshine hours [SA; hours]), monthly (average temperature [TA; °C], total rainfall [RR; mm/m^2^] and total sunshine hours [SA; hours]), and daily (minimum daily temperature [TM; °C], maximum daily temperature [TX; °C], total precipitation [RR; mm/m^2^] and total sunshine hours [SA; hours]) datasets. The INCA dataset (Haiden et al. 2011) consists of hourly estimates of rainfall rate [RR; kg/m^2^], global radiation [GL; W/m^2^], relative humidity [RH2M; %], wind speed in eastward direction [UU; m/s], wind speed in northward direction [VV; m/s], dew point temperature [TD2M; °C], air temperature [T2M; °C], and air pressure [P0; Pa]. Both the INCA and the SPARTACUS datasets have a spatial resolution of 1 × 1 km grid cells. We programmatically obtained the raw datafiles in NetCDF format from the datahub using R. Subsequently, we projected all layers to EPSG:4326 (WGS84) using the project() function of the R package terra (Hijmans et al. 2025) and restricted all datasets to a geographical bounding‐box (Longitude: 15.5° E—16.7° E; Latitude: 47.5° N—48.5° N) that includes the area covered by the city of Vienna and neighboring villages using the crop() function of the raster package in R. Using the resample function of the same package, we further interpolated the gridded datasets to obtain a higher resolution raster of 100 × 100 m grid cells for each continuous variable. For the yearly datasets, we extracted the single yearly value of the grid‐cell closest to the latitude/longitude coordinate pair for each of the samples using the extract() function from the R package raster (Hijmans and van Etten 2012). For the monthly datasets, we further extracted the values for the corresponding sampling month of the closest grid cell as described before. Finally, for the hourly and daily datasets, we averaged all values in a 14‐days interval prior to each collection date in the closest grid cells, since we assume that the weather conditions up to 2 weeks before the sampling may strongly influence the species composition in a given sample. We obtained land cover and land use data from the Austrian data portal (https://www.data.gv.at/) and from Copernicus Land Monitoring Service (CLMS, https://land.copernicus.eu/en). The Austrian data portal provides the land use map of Vienna, with 32 land use classes, in vector format. We defined a grid with 100 × 100 m cells and for all cells computed the share of each class separately. We then projected the 32 raster layers thus obtained in EPSG:4326. We processed the “Protected Areas” layer in the same way. Similarly, we used the buildings inventory of the city of Vienna to compute the share of built‐up area and built‐up volume in the 100 × 100 m cells.
The CLMS Imperviousness density and Tree‐cover density layers are raster layers providing the sealing resp. tree cover density in the range from 0% to 100% at 10 m resolution. They were resampled at 100 × 100 m and projected to EPSG:4326. In total we incorporated 58 independent variables into our analyses, including the information if flies were collected inside or outside of buildings, latitude, longitude, and collection date of samples.
Biodiversity Data Analyses
2.6
Based on the count data of each species, we first assessed the species‐specific total abundance and average abundance of each species per trap. To assess sampling completeness in the dataset, we performed species accumulation curve analysis using the vegan package in R (Dixon 2003). Species accumulation curves were generated using random sampling methods, and asymptotic species richness was estimated by fitting Michaelis–Menten models to the accumulation data. Bootstrap confidence intervals (n = 1000 replicates) were calculated for total richness estimates, and sampling completeness was assessed using bootstrap Z‐tests comparing observed richness to predicted asymptotic values. Statistical significance of sampling gaps was evaluated at α = 0.05 to determine whether additional sampling effort would likely yield new species discoveries. Species data were aggregated by sampling locations and further separated by “indoors” or “outdoors” collections, with zero‐abundance samples excluded. Subsequently, we calculated several estimators for α‐diversity in each of the traps using the R package vegan. These estimators include the species richness (SR), that is, the absolute number of species per trap, and the Shannon index (H′), which both estimate species richness and are thus strongly influenced by the number of rare species (Magurran 2004). Furthermore, we calculated the Evenness (EV) and the Simpson diversity indices (1‐D), which both summarize how evenly species are distributed in a sample (Magurran 2004).
We additionally estimated differences in community composition, i.e., β‐diversity, among samples based on Bray‐Curtis distances in R using the vegan package and employed non‐metric multidimensional scaling (NMDS) to visualize the projected distances among the samples along the first two NMDS axes. We furthermore included species positions in NMDS plots, which are computed as weighted averages based on the abundances of species in the samples along the ordination coordinates. Subsequently, we scaled the 58 environmental variables with point estimates for each of the sampling sites in and around Vienna using the R function scale() and performed PCA using the PCA() function from the FactoMineR package (Lê et al. 2008). We then used the envfit() function from the vegan package in R to fit the first four PC axes onto the NMDS ordination, using 99,999 permutations to assess the significance of correlations between the variables and the ordination axes.
Furthermore, we examined seasonal variation in species composition and abundance by plotting total abundance across the entire sampling season in 2‐week intervals for each identified species. To avoid spatial pseudoreplication, we focused specifically on 14 sampling locations that were sampled at least five times per season. We then used the adonis2() function of the vegan package to perform a PERMANOVA with 9999 permutations to test whether sampling month, sampling location, and trap placement (indoors vs. outdoors) influenced species composition. To evaluate whether diversity patterns changed significantly over the course of the season, we fitted linear and quadratic (polynomial) mixed models, which include the sampling location as a random factor to the monthly α‐diversity indices as described above and compared model performance based on Akaike Information Criteria (AIC).
Finally, we tested for spatial autocorrelation among sampling sites by calculating Moran's I (Moran 1948), using the R package spdep (Pebesma and Bivand 2023), based both on α‐diversity indices and species abundances. To define spatial relationships among samples, we used the knearneigh() function to generate a spatial weights matrix from the geographic coordinates and assessed the significance of Moran's I using randomization tests.
Ecological Data Analysis
2.7
For a more in‐depth assessment of links between environmental variables and the abundance of Drosophila species, we employed Redundancy Analysis (RDA), which is a multivariate statistical method that combines elements of multiple regression and principal component analysis to explore and quantify the relationships between explanatory (i.e., environmental data) and response matrices (i.e., species abundance data). Since certain environmental datasets were not available for all sampling sites, we generated geographical subsets of the sampling data and analyzed these datasets separately. Before conducting the RDA, we tested for and visualized intercorrelations among environmental factors in R using the cor() function and manually removed environmental factors including daily wind speed, global radiation, temperature, rainfall rate, monthly temperature means, and specific land use categories (outdoor sports, vineyard, water bodies, commercial areas) with high levels of positive or negative correlation (|r| > 90%) with at least one other factor and retained one of the intercorrelated factors. Subsequently, we standardized the values of the remaining environmental factors using the scale() function in R for z‐score normalization and Hellinger‐transformed the abundance data of the Drosophila species to reduce the impact of highly abundant species and excess zeros (e.g., Blanchet et al. 2014) while rendering the data appropriate for linear ordination methods such as principal component analysis (PCA) and redundancy analysis (RDA). We then used the rda() function from the vegan package in R to compute a full model including all environmental variables, as well as a null model conditioning on participant ID to control for collector effects. Subsequently, we performed forward model selection using ordiR2step() with 999,999 permutations to identify the combination of environmental variables that yielded the most informative model. To assess the significance of the most informative model and of individual terms, we conducted permutation tests with 9999 iterations using the anova.cca() function from vegan. Finally, we visualized the RDA results using the ordiplot() function.
Complementary to the analyses above, we compared four different approaches for species distribution modeling (SDM) to predict habitat suitability for Drosophila species across Vienna. Six rare Drosophila species with fewer than 20 observations (D. busckii, D. immigrans , D. phalerata, D. subobscura, D. testacea , and D. virilis ) were excluded from these analyses, as the very limited sample sizes would offer insufficient predictive power and could bias or distort habitat predictions. We first used the rast() function from the terra package (Hijmans et al. 2025) in R to combine land use, administrative boundaries, and annual average climate data (all in GeoTIFF format) into a comprehensive environmental predictor stack consisting of 44 distinct layers at a resolution of 230 × 300 grid cells, each of 100m^2^ size. We then aligned all raster layers to a common spatial template through resampling and cropped the whole stack to the study area extent. Species abundance data from the cleaned dataset restricted to Vienna were Hellinger‐transformed to account for zero‐inflation as described above, and spatial coordinates were converted to sf objects with an EPSG:4326 projection using the st_as_sf() function of the sf package (Pebesma and Bivand 2023).
Subsequently, we extracted environmental predictor values at each sampling location and performed species distribution modeling using a comprehensive comparative approach. Species were classified as “generalists” (present at > 50% of sampling sites; D. melanogaster and D. mercatorum ) or “specialists” (present at ≤ 50% of sampling sites; D. funebris , D. hydei , D. repleta , D. simulans , and D. suzukii) to account for differences in spatial occupancy patterns and to optimize model parameterization for random forest modeling.
We employed four different modeling frameworks to ensure robust species‐habitat relationship inference: (1) Generalized Linear Models (GLM) with a binomial error structure for baseline linear relationships, (2) Random Forest (RF) machine learning algorithms calculated with the randomForest package (Liaw and Wiener 2002) with species‐specific overfitting controls (generalists: 300 trees, maxnodes = 8, nodesize = 15; specialists: 500 trees, maxnodes = 15, nodesize = 10; mtry set to the square root of the predictor count) and balanced class weights to handle prevalence differences, (3) Generalized Additive Models (GAM) with smoothing splines (k = 3) as implemented in the mgcv package (Wood 2011) to capture non‐linear environmental responses while avoiding overfitting, and (4) Maximum Entropy models (MaxEnt) via MaxEnt.jar (Phillips et al. 2006), which was executed using the dismo package (Hijmans et al. 2024), a presence‐background approach commonly used in ecology. For all approaches, we used stratified sampling conducted with the caret package (Kuhn 2008) to partition the full dataset into 80% training and 20% testing subsets.
Model performance was evaluated using two complementary metrics on the independent test set: (1) Area Under the Receiver Operating Characteristic Curve (AUC), calculated with the pROC package (Robin et al. 2011), which measures the model's ability to discriminate between presences and absences (values range from 0.5 for random prediction to 1.0 for perfect discrimination), and (2) Cohen's Kappa statistic as implemented in the caret package, which assesses practical classification accuracy while correcting for chance agreement (values range from −1 to 1, with 0 indicating no better than random and values > 0.4 indicating moderate to substantial agreement). We also quantified overfitting by calculating the difference between training and test AUC, with lower differences indicating better model generalization. Based on this comprehensive model comparison, we selected the best‐performing approach for generating habitat suitability predictions across the entire study area using raster‐based environmental layers with the terra package, which were exported as GeoTIFF files. Results were visualized using ggplot2 with Stadia basemaps obtained via the ggmap package (Kahle and Wickham 2013), displaying predicted abundance values.
Results
3
During the sampling period, which spanned from late June to early December 2024, we received a total of 278 fly traps from 89 participants across the whole sampling area in Austria and 252 traps from 75 participants located in or near the city of Vienna, Austria (Figure 1). Most participants (n all = 49 and n Vienna = 40) returned a single trap, and the remaining participants returned more, several of them even between 10 and 33 traps throughout the sampling season resulting in an average of three traps per participant—both for the whole sampling area and for the city of Vienna, respectively. Samples outside of Vienna were collected in the Federal Provinces Burgenland (n = 1) and Lower Austria (n = 25).
Sample distribution. Geographic distribution of sampling sites in Vienna and surrounding areas. Locations for which detailed land use and environmental data are available are highlighted in blue. Sampling sites outside this core area are shown in red. The size of the circle corresponds to the number of repeated sampling at a given location.
Species Occurrence and Abundance
3.1
Our sampling effort yielded a total of 18,173 flies representing thirteen species of the genus Drosophila (Table 1, Table S2). We identified several species of the Sophophora subgenus, including two members of the melanogaster group ( Drosophila melanogaster , Drosophila simulans ), one member of the suzukii group (Drosophila suzukii), and one member of the obscura group (Drosophila subobscura). Furthermore, we found species of the subgenus Drosophila, including Drosophila virilis, several species of the immigrans‐tripunctata radiation lineage, including Drosophila funebris , Drosophila immigrans , Drosophila phalerata, and Drosophila testacea , and members of the repleta species group, including Drosophila hydei , Drosophila mercatorum , and Drosophila repleta , as well as one species from the subgenus Dorsilopha ( Drosophila busckii ).
Eleven species were found in Vienna as well as in some surrounding localities. Only two species, D. phalerata and D. virilis, were found exclusively within the city borders. Species accumulation curve analysis indicated that sampling completeness was high (88.6%), with 13 observed species compared to a predicted total richness of 14.7 species (95% CI: 10.3–16.5). Bootstrapping indicated no statistically significant sampling gap (Bootstrap Z‐test, p = 0.336), which suggests that our funnel‐trap sampling method captured most of the Drosophila species diversity present in the Vienna study area that is detectable with this approach.
To verify morphological species identifications, DNA barcoding was conducted on 2–6 individuals per species, resulting in a total of 53 barcoded individuals (Table 1). All obtained sequences confirmed our morphological determinations. Along with corresponding reference specimens, these data contribute to the Austrian Barcode of Life initiative (ABOL; see BOLD identification numbers in Table S1).
To our knowledge, this study is the first to scientifically document the occurrence of two previously unrecorded species in Austria: D. mercatorum and D. virilis . One of these neozoans, D. mercatorum (n = 8851), as well as D. melanogaster (n = 6726) was the most frequent species (Figure 2A) in our samples. In contrast, D. subobscura was, by far, the rarest with only five individuals across all the collections. In line with their overall abundance, D. melanogaster and D. mercatorum were consistently found in high numbers across many traps. Conversely, we found that certain species, such as D. virilis and D. funebris were overall rare but sampled in high numbers in only a few traps, as indicated by the large standard deviations shown in Figure 2B. We further observed that the location of the trap had a significant influence on the occurrence of certain species. While D. suzukii, D. immigrans , and D. simulans were approximately 45, 43, and 11 times more likely to be caught outdoors, respectively, D. mercatorum and D. repleta were about 11 and 16 times more likely to be found indoors than outdoors (Table 2 and Figure 2C). The three species D. phalerata, D. testacea , and D. subobscura, which were overall rare, were only caught outdoors.
*Abundance and phenology of the investigated Drosophila species. (A) Total number of individuals recorded per species across all sampling events. (B) Mean number of individuals per species per trap, with error bars representing standard deviations. (C) Proportion of traps (indoor vs. outdoor) in which each species was detected. Asterisks indicate significant differences in presence between trap locations (*p < 0.05, **p < 0.01, **p < 0.001). (D) Temporal distribution of species: Histograms showing absolute counts per species in two‐week intervals over the full sampling period.
TABLE 2: Results of χ 2 tests comparing the counts of samples collected indoors (n = 233) and outdoors (n = 45) for each species.
Furthermore, we observed differences in the phenology of the different species (Figure 2D). D. busckii , D. funebris , and D. testacea were predominantly trapped early in the year. Among the abundant species, D. melanogaster and D. simulans occurred commonly across the whole sampling season. Conversely, we found that D. suzukii was mostly caught early in the season, while D. virilis occurred only very late during the sampling season. However, we caution that our random sampling scheme at different locations and time‐points may contribute to sampling bias that could influence the seasonality patterns observed here.
Species Diversity and Community Composition
3.2
We next assessed α‐diversity across samples and found that traps generally exhibited moderate levels of diversity (Figure 3A). The mean Shannon index (H′) was 0.59 (SD = ±0.40), and the mean Simpson diversity index (1 − D) was 0.34 (SD = ±0.23), reflecting moderate species diversity across the dataset. Species richness ranged from 1 to 9 species per trap, with a mean of 2.81 (SD = ±1.36). The distribution of richness was positively skewed, indicating that most traps were dominated by just a few species, while only a minority harbored more diverse communities. Evenness was overall similarly moderate, with a mean of 0.66 (SD = ±0.22), suggesting that, on average, no single taxon dominated the assemblages. However, unlike richness, evenness values were negatively skewed. This pattern indicates that most samples had relatively balanced species abundances, while a smaller subset of traps exhibited low evenness—likely due to one or a few species dominating the community, despite higher overall richness.
α‐diversity and the influence of the environment on species diversity. (A) Histograms illustrating the distribution of α‐diversity metrics across all traps. Insets display the minimum, mean, and maximum values, with vertical red dashed lines indicating the mean for each metric. (B) Boxplot showing temporal dynamics of α‐diversity plotted in monthly intervals based on 14 sampling locations that were sampled at least five times per season. The p‐values shown at the top of the plots correspond to the fit of the quadratic function highlighted as blue lines. The other two subpanels show correlations between Species Richness (C) and Evenness (D) and the first four principal components (PCs) from a PCA based on 58 environmental variables (see also Figure S1). p‐values were derived from multiple ANOVA models, with α‐diversity metrics as dependent variables and latitude, longitude, and the four PC axes as independent variables. The four PC‐axes were dominated by short‐term weather conditions, long‐term climatic conditions, urbanization and aerial factors, such as wind and rainfall, respectively (see Table S7).
We also observed pronounced seasonal variation in biodiversity and community composition. AIC‐based model comparisons indicated that quadratic functions provided a better fit than linear models for all diversity indices (Table S3). However, we found that only Shannon and Simpson diversity indices exhibited significant temporal dynamics, with values peaking in mid‐summer and declining at the beginning and end of the season (Figure 3B). Conversely, Eveness and Species Richness were not significant and did not exhibit a convex seasonal pattern time (Table S3, Figure 3B). Moreover, our analyses revealed highly significant spatial autocorrelation among sampling locations in species richness (Moran's I = 0.230, p < 10^−15^), Shannon diversity (Moran's I = 0.128, p < 10^−5^), and Simpson diversity (Moran's I = 0.085, p = 0.001). At the species level, common cosmopolitan species exhibited stronger spatial clustering than local environmental specialists (Table S4). Given these strong spatial signals, we further fitted linear mixed‐effects models (LMEs) to test for the influence of location (indoors vs. outdoors), latitude, longitude, and sampling month on these two α‐diversity metrics (Table S5). Neither latitude nor longitude had a significant effect, indicating that spatial autocorrelation is not necessarily structured linearly along urban gradients.
Ecology and Environmental Preferences
3.3
To test if environmental factors had a significant influence on α‐diversity, we focused our analysis on samples collected within and around the city of Vienna, where detailed information on land use, climate, and weather conditions at the sampling time‐points were available. PCA of 58 environmental factors (Table S6), which were included in the analyses, revealed that the information of the first four axes (dimensions) was dominated by (1) short‐term weather conditions, (2) long‐term climatic conditions, (3) anthropogenic influence, including land use, as well as (4) wind, precipitation, and air pressure, respectively (Figure S1 and Table S7). Multiple linear regression revealed that Species Richness was significantly correlated with the first three principal component axes (Dim.1, Dim.2, and Dim.3; Figure 3C; Table S8). Notably, we found a strong negative correlation with Dim.3, which is primarily associated with anthropogenic factors. Moreover, the positive correlation with Dim.1 indicates that Species Richness increases with lower average annual temperature, higher average precipitation, and altitude. We likewise found significant correlations of Dim.3 with the Shannon and the Simpson diversity indices (Table S8). Conversely, we found the opposite correlation patterns for Eveness, suggesting that urban areas are dominated by a few, evenly distributed Drosophila species (Figure 3D; Table S8).
We further assessed β‐diversity by calculating pairwise Bray‐Curtis distances among all samples. The subsequent ordination with NDMS and correlation of species and PC axes from the environmental data with the two NMDS axes revealed substantial species‐specific differences in the occurrence among the traps. Along the first NMDS axis we observed a strong gradient ranging from D. mercatorum and D. repleta , which we previously identified to predominantly occur indoors (see above), to D. suzukii, D. phalerata, and D. testacea , which all showed a strong preference for outdoor habitats in our dataset (Figure 4A). Consistent with this observation, we found that Dim.3 of the environmental PCA, which was influenced by environmental factors associated with land use and human activity, was most significantly associated with the ordination space, Dim.3 explained about 14.3% of the variation in the ordination, which was statistically highly significant (p < 1e‐5; Table S9). The remaining three PC axes each explained 1.6% (Dim.1; p > 0.05), 2.11% (Dim.4; p > 0.05), and 2.79% (Dim.2; p = 0.044) and thus had either no or only very small influence on the species diversity. While not the dominant drivers of the variation, this result further indicates a substantial influence of urbanization on community composition and species distribution.
β‐diversity and the influence of environmental factors on species distribution. (A) NMDS plot based on Bray‐Curtis dissimilarities, with gray dots representing individual traps. Red crosses indicate the average position of each Drosophila species along the two NMDS axes, while blue arrows represent correlations with the first four principal components (PCs) from a PCA of 58 environmental variables (see also Figure S1). (B) Redundancy Analysis (RDA) plot showing the first two canonical axes. Red crosses indicate species' positions in RDA space, and blue arrows represent correlations with 14 environmental variables selected by forward regression. The maps in (C) display results from species distribution models based on random forest modeling. Color gradients indicate predicted occurrence probabilities, ranging from 0 to 1 (see also Table 3). Sampling locations of the corresponding species are indicated by blue dots.
Complementing the previous analyses, we applied redundancy analysis (RDA) to assess the influence of specific environmental factors on species composition. Using forward selection, we identified 14 environmental variables that contributed to the best‐fitting model, which was constrained by latitude and longitude (Table S10). This model explained approximately 26.9% of the variation in species composition (adjusted R ^2^ = 0.2690) and the most significant factors were “Location” (“indoors/outdoors”; F 1,213 = 21.74, p < 0.001), “sampling date” (F 1,213 = 20.3, p < 0.001), the proportion of greened road space (“greenstreets”; F 1,213 = 11.98, p < 0.001), the building volume in a grid cell (“BuildingVolume”; F 1,213 = 9.69, p < 0.001), the proportion of loosely built residential areas (“looseresidential”; F 1,213 = 7.84, p < 0.001), and monthly precipitation (“Monthly_RR”; F _1,213=_5.66, p = 0.002). In the RDA plot (Figure 4B), species positioned at the periphery typically occurred across many samples, while those near the center were often rare and observed in only a few samples. Notably, the two most common species, D. melanogaster and D. mercatorum , were located at opposite ends of the first RDA axis but shared similar positions along the second axis. In contrast, D. repleta appeared at the opposite end of the second axis, suggesting ecological differences among these three species. Most of the other species clustered near the center of the plot, reflecting their broader and more uniform distribution across environmental gradients or their rareness. Consistent with the results above, we found that the distribution of D. mercatorum on the RDA plot was positively correlated with several environmental factors, such as sampling location, building volume, and daily temperature. This is indicated by arrows pointing toward the species' position on the RDA plot along the first or second RDA axis, providing further evidence that D. mercatorum has a strong ecological preference for habitats with a high anthropogenic influence and elevated daily temperature. The second most common species, D. melanogaster , which is commonly assumed to be an obligatory human commensal, appears to be likewise, but less strongly influenced by temperature. In contrast to * D. mercatorum, D. melanogaster
- seems to be negatively correlated with building volume but more common in loosely built residential areas, which may suggest that it prefers less urban but rather sub‐ and peri‐urban habitats. Both D. simulans and D. repleta , which represent less common but still abundant species, were positively influenced by monthly rainfall and daily humidity but negatively correlated with daily temperature, which may indicate that these species prefer a cooler and more humid climate than the two aforementioned species. However, we caution that these analyses may be confounded by the unequal number of samples provided by citizen scientists, which may bias toward locations that have been sampled multiple times. To test the robustness of our results, we repeated the RDA analysis after removing daily and monthly climatic variables and collapsing all samples collected at the same sampling site. The collapsed abundance data were Hellinger‐transformed to account for unequal sample sizes. Consistent with the previous results, we found a qualitatively similar distribution of D. mercatorum , D. melanogaster , and D. simulans in the RDA plot (Figure S2). The main differences were that D. repleta was located closer to D. mercatorum , and D. hydei was positioned on the opposite side of the second RDA axis, relative to D. mercatorum . Moreover, D. repleta and D. melanogaster were farther from the plot center. Furthermore, we again found that “Location” (indoors/outdoors; F 1,74 = 7.04, p < 0.001) and building volume per grid cell (“BuildingVolume”; F 1,74 = 4.26, p = 0.01) were significantly associated with the distribution of D. mercatorum (Table S11). Overall, the model explained 14.6% of the variance in species distribution (adjusted R ^2^ = 0.14581). We therefore consider our main findings from the full model to be robust.
We subsequently performed a comprehensive species distribution modeling (SDM) analysis using four widely applied approaches—generalized linear models (GLM), generalized additive models (GAM), MaxEnt, and Random Forest—based on presence‐absence data from 222 samples collected within and around Vienna and 58 gridded environmental predictors (Table S6). In contrast to the abundance‐based models reported earlier, this expanded SDM framework enabled a direct comparison of different modeling paradigms and revealed consistent differences in predictive performance across species. Across seven species with sufficient sample size (≥ 20 presences) that we included in our analyses, Random Forest consistently achieved the strongest predictive performance, yielding the highest test AUC and Cohen's Kappa values for five of the seven species analyzed. GAM outperformed the other methods only for a single species, while GLM and MaxEnt rarely achieved the highest scores for any species (Table 3). Model comparison further revealed pronounced differences in overfitting among methods. MaxEnt exhibited the highest mean overfitting (20.5%), followed by Random Forest (14.9%) and GLM (14.8%), whereas GAM showed the lowest overfitting (6.8%) but at the cost of lower predictive accuracy for most species. At the species level, some models displayed substantial train/test divergence, indicating instability in learned relationships. Random Forest showed low overfitting for D. suzukii, D. simulans , and D. mercatorum , but considerably higher overfitting for several other species, reflecting variability in model generalization across taxa (see Table 3).
The species‐specific comparisons confirmed that only a subset of species achieved high test AUC values across models, whereas others performed poorly regardless of method. For example, Random Forest models for D. suzukii, D. simulans , and D. mercatorum produced high test AUC (> 0.8) and Kappa (~0.6) values with comparatively low overfitting, while D. melanogaster and D. funebris showed only moderate performance (test AUC = 0.729 and AUC = 0.746, respectively). In contrast, all four modeling approaches produced low predictive performance for D. hydei and * D. repleta—*in some cases yielding even negative Kappa values, accompanied by strong overfitting in both training and cross‐validation metrics. These outcomes indicate that the distributions of these latter species are either only weakly associated with the measured environmental variables or driven by unmeasured fine‐scale habitat features. The spatial predictions from the Random Forest models, as shown in Figure 4C, complement and support our previous findings based on abundance data as shown above. Drosophila mercatorum was strongly concentrated in highly urbanized areas characterized by elevated imperviousness and human activity, whereas D. melanogaster was broadly distributed across Vienna, occurring in a wide variety of habitats. Conversely, D. suzukii and D. simulans showed clear hotspots outside the city center corresponding to their expected ecological requirements as fruit specialists that are rarely found inside human houses.
Discussion
4
The Influence of Urbanity on Drosophila Communities
4.1
Urban landscapes are often considered ephemeral environments with a high turnover rate that negatively affect biodiversity, while favoring ecological generalists and neobiota (Johnson and Munshi‐South 2017; Lewthwaite et al. 2024; McGlynn et al. 2019; Sukopp 2008; Szulkin et al. 2020). To investigate these ecological dynamics and better understand the impact of human activity on biodiversity in urban landscapes, we analyzed the composition of Drosophila species communities in the metropolitan area of Vienna in Austria. Specifically, we focused on samples collected in close proximity to human settlements, both indoors (within kitchens, living or dining rooms) and outdoors (on balconies and in gardens).
Consistent with previous studies (for a review, see Sukopp 2008) and our expectations, we found that Drosophila species richness was negatively correlated with urbanity. The Drosophila community of the Vienna city area was dominated by synanthropic and cosmopolitan members of the “cosmopolitan guild”, namely D. melanogaster , D. simulans , D. funebris , D. hydei , D. immigrans , and D. repleta , and a few specimens of D. busckii , which are characterized as generalist species with strong associations to humans (Atkinson and Shorrocks 1977; Miller et al. 2017; Nunney 1996). As shown by previous studies, for example, in France (Ulmer et al. 2024), North America (Avondet et al. 2003; Bombin and Reed 2016), and with the exception of D. funebris also in South America (Ferreira and Tidon 2005; Garcia et al. 2012; Gottschalk et al. 2007), these species have been found to be common in world‐wide urban areas. This highlights the strong synanthropic and hemerophilous character of these species and underscores their ability to thrive in highly disturbed environments, such as urban habitats, even across different climatic zones (Atkinson and Shorrocks 1977; Nunney 1996).
We also detected recent additions to the neobiotic Drosophila fauna of Vienna, such as D. suzukii, a fruit pest originating from Asia that was first reported in Austria in 2012 (EPPO Global Database [https://gd.eppo.int/]; Asplen et al. 2015). In addition, we found two species not previously reported in the scientific literature to occur in Austria to date: D. mercatorum and D. virilis . Drosophila mercatorum , which was the most common species in our collection, was originally described from California and is distributed throughout the United States, Mexico, and South America. It has been introduced to Europe in the last century, first recorded in Spain (Prevosti 1953). Subsequent reports from various countries—from Portugal to Ukraine—document the continued expansion of its range (Adaschkiewitz and Gossner 2013; Ivannikov and Zakharov 2000; Kraaijeveld 1992; Pité 1972; Widmann and Bächli 2022). In Austria, a few records on the citizen‐science platform iNaturalist and a sample barcoded by L. Timaeus already in 2022 (ABOL‐BioBlitz22‐0415; https://portal.boldsystems.org/record/TDAOE2549‐23) indicate that the species' presence was not entirely unnoticed. However, as D. mercatorum is easily confused with D. repleta , it may have remained overlooked in many locations for a longer time. For instance, Vilela (1983) reported this species from Zimbabwe based on a specimen originally collected in 1935. Drosophila virilis , the second newly reported species for Austria which was rather rare in our collections, originated in Asia—likely from forested regions of China or the arid zones of Iran and Afghanistan (Throckmorton 1982)—and only recently expanded its range as a human commensal, now occurring widely across the Northern Hemisphere (Mirol et al. 2008; Vieira and Charlesworth 1999; Wang et al. 2006).
We failed to find several Drosophila species that have been reported in a study on drosophilids of the Vienna area 34 years ago (Gross and Christian 1994). This study likewise used funnel traps to sample Drosophila, but placed them along predefined transects including a greater proportion of more natural habitats than our own sampling—such as park areas, open meadows, riverbeds, and forests. Only 10 species— D. busckii , D. funebris , D. hydei , D. immigrans , D. melanogaster , D. phalerata, D. repleta , D. simulans , D. subobscura, and D. testacea —were also identified in our survey. In total, 12 species reported by Gross and Christian (1994) were not detected in the present study. Among them were nine Drosophila species— D. ambigua , D. bifasciata , D. confusa , D. deflexa, D. helvetica, D. kuntzei, D. limbata , D. obscura , D. rufifrons , D. subsilvestris, D. tristis , and *D. tsigana—*as well as three taxa that are no longer classified within the genus Drosophila. D. rufifrons and D. deflexa are now placed in Scaptodrosophila (Grimaldi 1990), while D. confusa is assigned to Hirtodrosophila (Grimaldi 1990). Thus, nine Drosophila species appear to be absent, which corresponds to a 47.4% Drosophila species reduction over time. Two of these belong to the quinaria group, and one to the melanica group. The most dramatic changes were observed in the obscura group. Seven members of the obscura group were not observed in the present study. These include D. obscura , a species native to Europe and reported as very common by Gross and Christian (1994). Moreover, D. subobscura, which was the most common species in the previous study, was the rarest in our survey, represented by only five specimens. Such a strong reduction in abundance may indicate that D. subobscura is under particular selection pressure due to global warming. This hypothesis is supported by a recent study that identified strong genomic signals of rapid adaptation for increased thermal tolerance in numerous European D. subobscura populations (Rodríguez‐Trelles and Tarrío 2024). We further found that D. deflexa and D. tsigana (Burla and Gloor 1952) – two Drosophila species that were previously detected for the first time in Austria or even in Central Europe (Gross and Christian 1994) – were absent from our collections, which may suggest that these species either failed to establish in the study area or that our sampling design was not sensitive enough to detect these species in close proximity to human housing.
In summary, the comparison of species composition over the past three decades may indicate a substantial turnover and reduction in Drosophila diversity in the metropolitan area of Vienna, consistent with the accelerating global biodiversity loss observed in recent decades (Drenckhahn et al. 2020; Isbell et al. 2023). The reasons for these changes are likely manifold: (1) Driven by global climate change, the average annual temperature in the inner city and the Vienna catchment area has risen by more than 2°C during this period (see Figure 5), which may have reduced the competitiveness of Drosophila species with narrow temperature niches (Capek et al. 2025; Kim et al. 2020; Sillero et al. 2014). (2) Furthermore, increasing temperatures may have triggered indirect ecological effects, such as shifts in food availability or changes in the prevalence of parasites, parasitoids and insect diseases, which could differentially affect the Drosophila community (Corcos et al. 2019). (3) Alterations in human activities, such as waste management practices or the use of insecticides, may have disproportionately impacted the species that are now absent compared to those still detected in our survey (e.g., Collins et al. 2024). (4) Finally, competition with newly introduced Drosophila species occupying overlapping niches may have contributed to the decline of formerly common species (Krijger et al. 2001; Rombaut et al. 2023). In addition to these biological and ecological explanations, we cannot exclude the possibility that differences in sampling design also contributed to the observed patterns. In contrast to Gross and Christian (1994), who predominantly collected their 28 spatio‐temporal samples outdoors, most of our samples (83.8%, n = 233) were collected indoors, with only 45 samples obtained outdoors. Moreover, Gross and Christian (1994) placed their traps in more rural and semi‐natural landscapes, which may have facilitated the capture of less synanthropic species compared to our study. However, seasonal differences, bait choice and subtle differences in the trap design may have favored or repelled different species in this study and the former fly collections. We therefore speculate that the observed differences in species abundance and community composition may reflect major ecological and biological changes, potentially reinforced by differences in sampling methods between the present and the previous study by Gross and Christian (1994).
Annual average temperature change in Vienna. Line plot depicting annual average temperatures at the sampling locations since 1991. Blue lines represent samples from the central urban region with a high degree of imperviousness (highlighted by a blue box in the map at the top left). Red lines represent the remaining sampling locations. Bold straight lines indicate regression lines from linear models calculated separately for the two groups. Insets at the top show the average annual temperature increase as well as the total increase across the whole time span, both for the full dataset and for the two subsets.
Ecology of Urban and Suburban Drosophila
4.2
We observed pronounced seasonal changes in biodiversity. Both Shannon and Simpson indices peaked in mid‐summer, indicating that this period provides good ecological conditions for most species in an urban environment and results in the highest levels of α‐diversity. This pattern aligns with phenology patterns shown in Figure 2D, where most species exhibit partial overlap in July and August. In contrast, we did not observe similarly pronounced convex seasonal patterns in Species Richness or Evenness. Instead, both metrics showed more linear trends: Species Richness decreased over time, whereas Evenness increased. This suggests that the community became dominated by fewer species with relatively similar abundances in the autumn months.
The temporal patterns observed based on Simpson and Shannon indices, however, contrast with those of Gleason et al. (2019), who reported a linear increase in α‐diversity based on Simpson and inverse‐Simpson indices over time in a Drosophila community sampled in the temperate zone of the United States. Several factors may explain this discrepancy: (1) Gleason et al. (2019) sampled fly species in a rural area of Kansas, USA, where urban environmental influences—likely more prominent in our study—are reduced and where overall environmental conditions may differ substantially from those in Vienna, Austria; (2) the species composition differed markedly, with only five synanthropic species overlapping between the two studies; and (3) Gleason et al. (2019) focused on a single sampling site, whereas our study included multiple locations, potentially capturing broader and more generalizable trends. Nevertheless, denser temporal sampling across additional urban and suburban habitats will be essential to determine whether the seasonal patterns we observed are specific to our study area or reflect a more widespread feature of urban ecosystems. Such broader datasets would also help identify the ecological factors driving seasonal dynamics.
These ecological factors are closely tied to differences in ecological niches, which can strongly influence the composition of Drosophila communities across environments. Previous studies have shown that many Drosophila species, even within the cosmopolitan guild that commonly coexist sympatrically, exhibit substantial differences in their food preferences and breeding sites. For example, D. melanogaster , D. simulans , and D. suzukii—all members of the Sophophora subgenus—prefer fruits at different stages of decay as food sources (Atkinson and Shorrocks 1977; Brake and Bächli 2008). In contrast, species of the obscura group are typically associated with forest habitats and are thought to rely on fungal substrates, although detailed information on their breeding sites remains scarce (e.g., Kimura 1980). Members of the quinaria group, such as D. phalerata and D. testacea (Scott Chialvo et al. 2019), are recognized mushroom feeders, whereas D. virilis preferentially feeds from slime flux of trees but also breeds on other substrates (Spieth 1979). Meanwhile, members of the repleta group, such as D. repleta and D. mercatorum , utilize decaying plant material, including rotting cacti in their native habitats in the Americas (Hasson et al. 1992; Oliveira et al. 2012).
Because the traps provided by citizen scientists contained a wide variety of baits—including citrus fruits, berries, apples, bananas, and other fruits—it was not possible to directly assess the food preferences of the investigated fruit fly species in this study. To account for the fact that ecological niches are shaped by more than food sources alone, we complemented our survey with a comprehensive environmental dataset comprising climatic, weather, and land‐use variables at the sampling locations to analyze the ecological preferences of the observed Drosophila species.
Complementary analyses based on redundancy analysis and environmental correlations with Bray–Curtis distances revealed that short‐term weather conditions, such as daily temperature and rainfall, strongly influenced species composition and abundance. However, the strongest impact was associated with the degree of urbanization, measured, for example, by land‐use types as well as by building volume and area at the sampling sites.
The neozoan fly D. mercatorum , and to a lesser degree D. repleta , clearly stood out as isoanthropic species. In particular, D. mercatorum was rarely captured outdoors and was found predominantly in highly urbanized regions with the greatest levels of imperviousness. By contrast, D. melanogaster —commonly regarded as a strict human commensal with a high degree of hemerophily—showed a preference for more suburban areas with substantial green space. Our analyses further indicated marked ecological differences between D. melanogaster and its sister species D. simulans , the latter being found mainly in rural residential areas and only rarely indoors. At the opposite end of the spectrum, we observed species such as D. phalerata and D. testacea , which are considered native to Central Europe and were never detected indoors or in highly impervious areas.
Theoretical frameworks from urban ecology provide a useful lens for interpreting these patterns. Urban environments act as strong environmental filters, sorting species into “urban avoiders, adapters, and exploiters (or dwellers)” based on their ecological traits and tolerance of human‐modified habitats (Blair 1996; Fischer et al. 2015). Our finding that cosmopolitan, synanthropic species dominate present‐day Vienna, while several formerly common species—such as D. subobscura and D. obscura —have disappeared, is consistent with this filtering process, whereby urban avoiders and weak adapters decline as natural habitats become increasingly fragmented or lost. Conversely, the successful establishment of neozoan species such as D. mercatorum aligns with evidence that urbanization can facilitate the introduction and persistence of non‐native, disturbance‐tolerant taxa (Carlon and Dominoni 2024). However, quantitative abundance data from Drosophila communities in other cities are needed to determine whether the community structures reported here represent general patterns.
Conclusion
5
In this study, we comprehensively investigated the Drosophila community through dense spatiotemporal sampling in and around the metropolitan area of Vienna, Austria—a region characterized by a diverse range of landscapes and land‐use types.
Our analyses provide evidence for highly distinct ecological niches among most of the investigated species. This is particularly remarkable given that the majority of specimens in this study were collected indoors and were thus only indirectly exposed to weather conditions and climatic factors. Our findings therefore suggest that flies are highly mobile and primarily transient visitors to human dwellings, spending much of their life cycle outdoors where they are directly influenced by the environmental factors shaping their niches. Given the broad ecological spectrum of the species identified in this study, ranging from strict specialists to common generalists, our results further underscore the versatility of the genus Drosophila as bioindicators. While additional data from other urban areas are needed to establish general patterns, Drosophila community composition holds promise as a reliable indicator of the degree of anthropogenic disturbance in an ecosystem.
Our results further highlight the power of citizen science for biodiversity assessment, made possible through the coordinated efforts of many contributors who collected samples randomly but followed standardized protocols. Benefitting from the participation of 89 citizen scientists, we collected and analyzed approximately 18,000 specimens from over 280 individual fly traps. An in‐depth comparison with a biodiversity survey conducted more than 30 years ago revealed significant changes in species abundance and community composition, underscoring the potential of Drosophila as a versatile model for monitoring biodiversity loss. Moreover, since Drosophila communities consist of species ranging from wild and local to highly synanthropic with global distributions, they provide a powerful system for assessing the degree of anthropogenic impact on ecosystems. Taken together, our study demonstrates that long‐term monitoring of Drosophila, empowered by citizen science, represents an effective and scalable approach for tracking biodiversity change in a rapidly transforming world.
Author Contributions
Martin Kapun: conceptualization (equal), data curation (equal), formal analysis (equal), funding acquisition (equal), investigation (equal), methodology (equal), project administration (lead), software (equal), supervision (lead), visualization (equal), writing – original draft (equal). Sonja Steindl: conceptualization (equal), data curation (equal), formal analysis (equal), investigation (equal), methodology (equal), software (equal), writing – original draft (equal). Maria Ricci: data curation (equal), formal analysis (equal), investigation (equal), methodology (equal), resources (equal), software (equal), writing – original draft (equal), writing – review and editing (equal). Manuel Löhnertz: data curation (equal), formal analysis (equal), investigation (equal), methodology (equal), resources (equal), software (equal), writing – original draft (equal), writing – review and editing (equal). Flora Strasser: data curation (equal), investigation (equal), writing – review and editing (equal). Rui Qiang Chen: data curation (equal), investigation (equal), writing – review and editing (equal). Lorin Timaeus: investigation (equal), validation (equal), writing – review and editing (equal). Nikolaus Szucsich: investigation (equal), methodology (equal), validation (equal), writing – review and editing (equal). Elisabeth Haring: conceptualization (equal), formal analysis (equal), investigation (equal), methodology (equal), resources (equal), supervision (equal), writing – original draft (equal), writing – review and editing (equal).
Funding
This work was supported by HORIZON EUROPE Framework Program (101059238).
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Figure S1: Principal Component Analysis (PCA) of environmental factors. Biplots display the loadings of individual environmental variables along the first two principal components (Dim.1 vs. Dim.2, left) and the third and fourth components (Dim.3 vs. Dim.4, right). Arrows beside each axis summarize the most influential environmental factors contributing to the corresponding dimensions, providing a visual overview of key gradients represented by each principal component. Figure S2: Redundancy Analysis (RDA) of collapsed abundance data. RDA plot showing the first two canonical axes similar to Figure 4B. Red labels indicate the positions of each species in RDA space, and blue arrows represent correlations with four environmental variables (see Table S2) selected by forward regression.
Table S1: Table containing barcoded Drosophila specimens, including BOLD accession numbers, Sample Ids and Collection Date. All voucher specimens are stored at the Natural History Museum Vienna. Table S2: Coordinates, sampling dates, species counts, and environmental factors for all samples included in the dataset. Table S3: Table showing AIC and p‐values of linear and quadratic models that test for assoications between sampling months and diversity indices. Table S4: Table showing species‐specific values of Moran's I and corresponding p‐values which quantify the level of spatial autocorrelation among sampling sites. Table S5: Table showing the results of general linear models assessing the influence of space and and time on diversity indices. Table S6: Environmental factors used for species distribution modeling, including their identifier, description, properties, data source, and usage. Table S7: Loadings of variables on the first four principal component (PC) axes. Values represent correlations, with positive loadings shown in blue and negative loadings in red. Table S8: Results of an Analysis of Deviance for generalized linear models testing the effect of environmental factors on alpha‐diversity measures, showing each factor, test statistic, p‐value, and significance. Table S9: Results of the envfit analysis testing associations between environmental factors and NMDS axes based on Bray–Curtis distances. Table S10: RDA results showing the relative importance of environmental factors in explaining Drosophila species composition in Vienna. Table S11: RDA results showing the relative importance of environmental factors in explaining Drosophila species composition in Vienna, with species abundances summed by sampling site (see also Table S6).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adaschkiewitz, W. , and M. M. Gossner . 2013. “Einige Arten aus den Familien Anthomyiidae, Calliphoridae, Dolichopodidae, Drosophilidae, Muscidae und Phoridae (Diptera) neu für Deutschland.” Studia Dipterologica 20: 214.
- 2Adrion, J. R. , A. Kousathanas , M. Pascual , et al. 2014. “ Drosophila suzukii: The Genetic Footprint of a Recent, Worldwide Invasion.” Molecular Biology and Evolution 31: 3148–3163. 10.1093/molbev/msu 246.25158796 PMC 4245814 · doi ↗ · pubmed ↗
- 3AK‐Diptera , and G. Bächli . 2023. “Checkliste der Taufliegen Deutschlands (Diptera: Drosophilidae).” 10.20363/CDG.DROSOPHILIDAE.2023.IX.25. · doi ↗
- 4Arguello, J. R. , S. Laurent , and A. G. Clark . 2019. “Demographic History of the Human Commensal Drosophila melanogaster .” Genome Biology and Evolution 11: 844–854. 10.1093/gbe/evz 022.30715331 PMC 6430986 · doi ↗ · pubmed ↗
- 5Asplen, M. K. , G. Anfora , A. Biondi , et al. 2015. “Invasion Biology of Spotted Wing Drosophila (Drosophila suzukii): A Global Perspective and Future Priorities.” Journal of Pest Science 88: 469–494. 10.1007/s 10340-015-0681-z. · doi ↗
- 6Atkinson, W. , and B. Shorrocks . 1977. “Breeding Site Specificity in the Domestic Species of Drosophila .” Oecologia 29: 223–232. 10.1007/BF 00345697.28309117 · doi ↗ · pubmed ↗
- 7Avondet, J. L. , R. B. Blair , D. J. Berg , and M. A. Ebbert . 2003. “ Drosophila (Diptera: Drosophilidae) Response to Changes in Ecological Parameters Across an Urban Gradient.” Environmental Entomology 32: 347–358. 10.1603/0046-225X-32.2.347. · doi ↗
- 8Bächli, G. 1982. “On the Type Material of Palearctic Species of Drosophilidae (Diptera).” Beiträge Zur Entomologie = Contributions to Entomology 32: 289–301.