Characterizing hypoxia-orchestrated post-stroke changes in oligodendrocyte precursor cells for optimized cell therapy
Yasuhiro Kuwata, Ken Yasuda, Kazuto Tsukita, Akihiro Kikuya, Naoki Takayama, Narufumi Yanagida, Kimitoshi Kimura, Ryosuke Takahashi, Riki Matsumoto, Takakuni Maki

TL;DR
This study shows how different levels of hypoxia after stroke change oligodendrocyte precursor cells, which could help improve cell therapy for stroke recovery.
Contribution
The study identifies hypoxia as a key driver of OPC adaptation and proposes hypoxia profiling to optimize cell therapy for stroke.
Findings
Mild hypoxia promotes oligogenic OPCs that support repair, while severe hypoxia induces angiogenic OPCs.
Hypoxia-modulated OPCs improve recovery when transplanted into stroke-affected mice.
Oxygen tone is a critical regulator of OPC adaptation post-stroke.
Abstract
Oligodendrocyte precursor cells (OPCs) are highly adaptable, engaging in diverse functions beyond myelination. However, how OPCs adjust their roles after ischemic stroke and contribute to recovery remains largely unknown. To address this gap, we constructed a “transient middle cerebral artery occlusion (tMCAO) atlas” by integrating mouse single-cell RNA sequencing (scRNA-seq) datasets and combined it with ex vivo OPC cultures and in vivo cell transplantation experiments. This approach revealed the emergence of “angiogenic” OPCs in the subacute phase and “oligogenic” OPCs in the chronic phase, driven by distinct levels of hypoxia—severe hypoxia inducing angiogenic OPCs and mild hypoxia promoting oligogenic OPCs. Ex vivo, severe hypoxic preconditioning faithfully induced angiogenic OPCs, and their intravenous transplantation enhanced angiogenesis and improved recovery in tMCAO mice. These…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1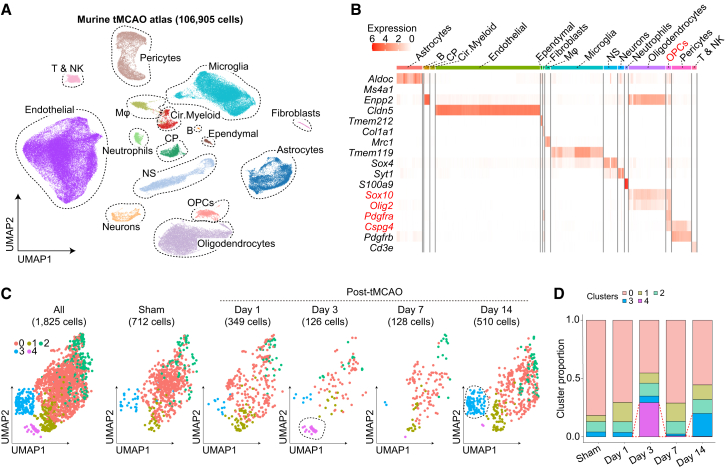 Figure 2
Figure 2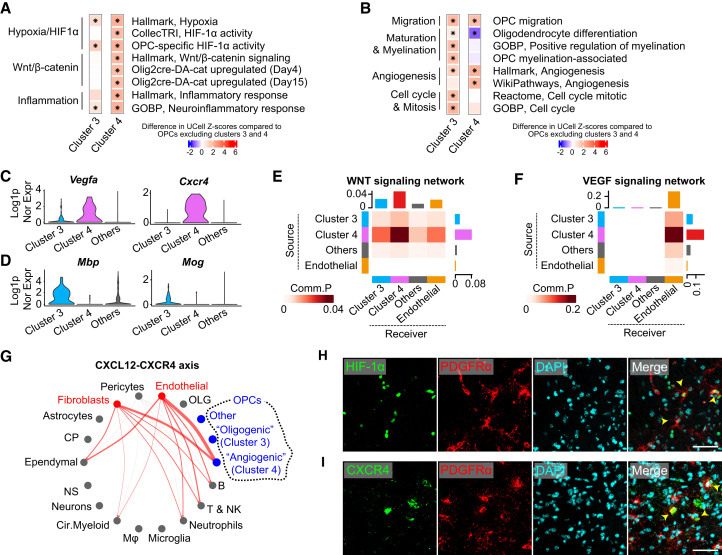 Figure 3
Figure 3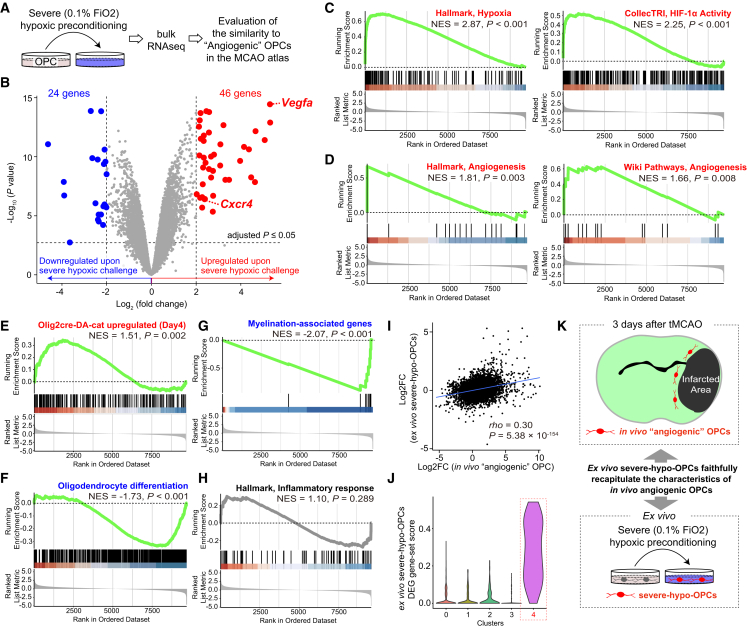 Figure 4
Figure 4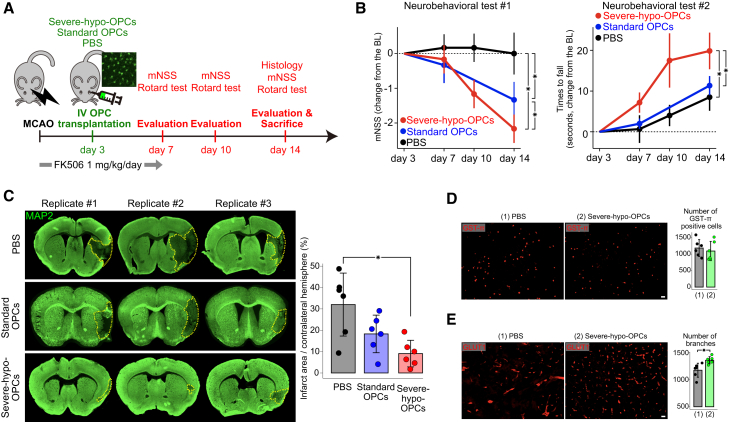 Figure 5
Figure 5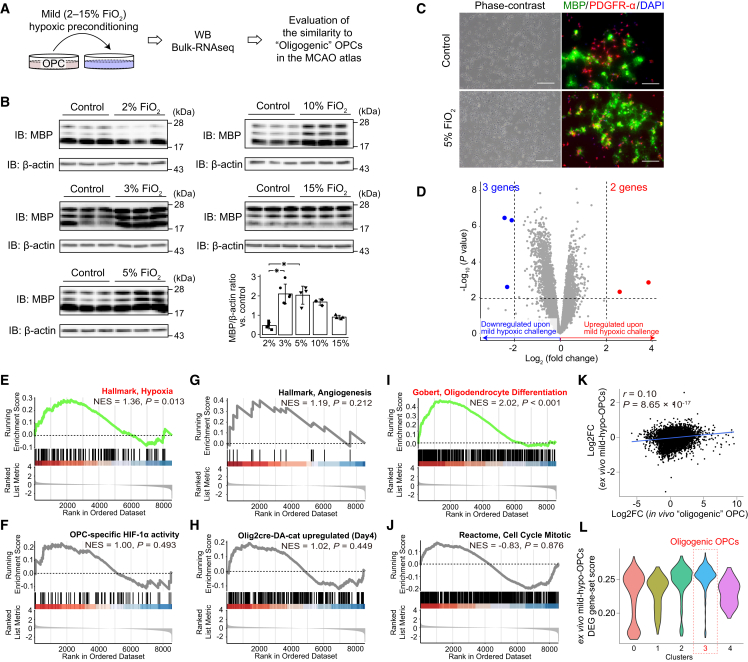 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurogenesis and neuroplasticity mechanisms · Neuroinflammation and Neurodegeneration Mechanisms · Single-cell and spatial transcriptomics
Introduction
Oligodendrocyte precursor cells (OPCs) originate during embryogenesis and persist as resident cells in the adult brain parenchyma (Akay et al., 2021; Bergles and Richardson, 2015). Initially generated in the subventricular zones of the embryonic neural tube, OPCs migrate throughout the brain and spinal cord (Tsai et al., 2016), continuing to proliferate and develop into oligodendrocytes during postnatal development (Akay et al., 2021; Bergles and Richardson, 2015). While the rate of oligodendrocyte production significantly declines as the brain matures, OPCs remain abundant in the adult brain (Akay et al., 2021; Bergles and Richardson, 2015). OPCs’ primary role is to differentiate into oligodendrocytes and contribute to myelination (Akay et al., 2021; Bergles and Richardson, 2015); however, it is increasingly recognized that OPCs actively modify their characteristics in response to the surrounding environment, participating in various aspects of brain development, structure, and function through interactions with other cells (Akay et al., 2021; Xiao and Czopka, 2023).
Among various environmental cues influencing OPCs, growing evidence suggests that they utilize hypoxia-inducible factor 1α (HIF-1α), a key regulator of cellular responses to hypoxia, to shift toward an angiogenic phenotype during embryonic development (Yuen et al., 2014). In the fetal brain, extremely low oxygen levels (partial pressure of oxygen ≤ 7.60 mmHg, equivalent to a fraction of inspiratory oxygen [FiO2] ≤ 1.00%) render HIF activity indispensable for brain development (Zhang et al., 2011). During embryonic development, HIF-1α-instructed OPCs inhibit their maturation into oligodendrocytes by upregulating WNT signaling in an autocrine manner and/or by suppressing Sox10 expression via non-canonical HIF-1α targets (Allan et al., 2021; Yuen et al., 2014). At the same time, they promote angiogenesis in a paracrine manner by activating WNT and/or vascular endothelial growth factor receptor signaling in endothelial cells (Allan et al., 2021; Minocha et al., 2015; Yuen et al., 2014). Notably, autocrine activation of WNT signaling in OPCs upregulates Cxcr4, facilitating OPC association with endothelial cells through the CXCR4-CXCL12 axis (Fancy et al., 2009; 2014; Tsai et al., 2016). Thus, a key role of OPCs in embryonic brain development is to associate with vasculature and promote angiogenesis, driven by severe hypoxia in the fetal brain and coordinated by HIF-1α, WNT signaling, and Cxcr4.
Importantly, cerebral ischemia is another condition where hypoxia-driven cellular modulation becomes prominent (Baranova et al., 2007). Interestingly, although prolonged extreme hypoxia leads to cell death, sub-lethal hypoxic conditions can shift brain tissue toward greater resilience against cerebral ischemia (Rybnikova et al., 2022). Although the mechanisms of hypoxia-induced tolerance to ischemic stroke have mainly focused on neuronal cells, it has also been reported that neuronal changes alone do not fully explain the observed resilience (He et al., 2021). This highlights the important role of other cell types within the neurovascular unit, including OPCs, in stroke pathophysiology (Tiedt et al., 2022; Kishida et al., 2019; Okazaki et al., 2019; Hase et al., 2022).
Current acute ischemic stroke therapies primarily aim to rapidly restore cerebral blood flow to minimize damage (Chamorro et al., 2021). While their success depends on how quickly and effectively blood flow is restored, complete recovery is not always achievable in all patients, underscoring the need for additional therapies to provide “neuroprotection” (Chamorro et al., 2021). Among potential therapeutic approaches, stem cell transplantation is particularly promising, as it can provide the necessary cellular components that are otherwise insufficient in the post-stroke brain (Anthony et al., 2022). Although the potential of stem cell transplantation for ischemic stroke treatment is unquestionable (Anthony et al., 2022), its clinical translation to humans remains challenging (Houkin et al., 2024), highlighting the unmet need for further refinement of cell-based therapies. To this end, we propose two key steps: (1) identifying specific cells that inherently protect ischemic tissue but are insufficient on their own and (2) understanding their adaptive responses (Kakae et al., 2023; Kumar Podder et al., 2024). Following ischemic stroke, the brain undergoes dynamic biological changes driven by both local interactions within the neurovascular unit and systemic metabolic shifts (Hosoki et al., 2020; Tiedt et al., 2022). Therefore, a thorough understanding of these biological events is crucial to identifying protective responses that should be supported. In this context, a promising strategy to improve stem cell transplantation is to modify these cells ex vivo to mimic in vivo adaptations, thereby enhancing their protective functions and optimizing therapeutic outcomes.
Here, we focused on OPCs for two reasons: (1) under physiological conditions, only a limited number of OPCs migrate to ischemic regions (Wang et al., 2022); and (2) our previous studies demonstrated that ex vivo OPCs preconditioned with severe hypoxia secrete a variety of pro-angiogenic factors (Kishida et al., 2019). These findings suggest that hypoxia-induced phenotypic changes in OPCs can enhance ischemic tolerance in brain tissue, though their natural response remains suboptimal.
In this study, we first compiled and analyzed publicly available single-cell RNA sequencing (scRNA-seq) datasets to construct a “transient middle cerebral artery occlusion (tMCAO) atlas” and profile OPC transcriptional changes during ischemic stroke. We chose tMCAO as the model for acute ischemic stroke as it is a well-established and widely used model (Frazier et al., 2023; Kim et al., 2022; Nakahashi-Oda et al., 2021; Shi et al., 2021a; 2021b; Wu et al., 2023; Zeng et al., 2023; Zheng et al., 2022). This model induces ischemic injury in the ipsilateral cortex, striatum (including the caudoputamen), and adjacent white matter, thereby closely reproducing the distribution of infarcts typically observed in patients with middle cerebral artery territory stroke. Importantly, tMCAO allows reperfusion, which is highly relevant to current clinical practice involving thrombolysis and thrombectomy. Using the tMCAO atlas, we found that a subset of OPCs altered their transcriptome in the subacute stage after tMCAO (day 3), adopting a distinct hypoxia-induced, HIF-1α-driven “angiogenic” transcriptional profile that mirrors OPC characteristics reported in embryonic brain development (Yuen et al., 2014). In contrast, in the chronic stage after tMCAO (day 14), OPCs transitioned into a distinct “oligogenic” transcriptional profile. Importantly, ex vivo, we successfully induced OPCs to acquire transcriptomic characteristics similar to those of in vivo “angiogenic” OPCs by utilizing severe hypoxia preconditioning. Notably, intravenous transplantation of these cells efficiently promoted angiogenesis, reduced infarct volume, and ameliorated functional decline after tMCAO. Finally, we demonstrated that varying oxygen levels critically influence OPC maturation ex vivo, with mild hypoxia optimally promoting OPC maturation and, at least in part, contributing to the emergence of in vivo “oligogenic” OPCs. These findings underscore the crucial role of “oxygen tone” in shaping temporally dynamic reparative OPC phenotypes and highlight the therapeutic potential of hypoxia-preconditioned OPCs for enhancing post-stroke recovery.
Results
Creation of the tMCAO atlas
We systematically searched the BioProject database to identify scRNA-seq datasets that investigated post-tMCAO transcriptional changes and utilized the droplet-based 10× Genomics Chromium approach (see Methods) (Frazier et al., 2023; Kim et al., 2022; Nakahashi-Oda et al., 2021; Shi et al., 2021a; 2021b; Wu et al., 2023; Zeng et al., 2023; Zheng et al., 2022). To minimize batch effects across studies as much as possible, we downloaded raw FASTQ data and processed all datasets using the same pipeline for mapping, followed by uniform and stringent quality control according to single-cell best practices (see Methods) (Heumos et al., 2023). For our meta-analysis, we analyzed scRNA-seq datasets derived primarily from the ipsilateral hemisphere after tMCAO, encompassing both the infarct core and peri-infarct regions. Contralateral hemispheres from tMCAO mice were excluded, and control datasets were obtained from the ipsilateral hemisphere of sham-operated mice. Ultimately, we included 21 samples from 6 studies that provided sufficient OPCs to construct the tMCAO atlas (Table S1). After quality control, our tMCAO atlas comprised 106,905 cells from sham-operated, 1-day, 3-day, 7-day, and 14-day post-tMCAO groups (Figures 1A and 1B).Figure 1. Murine transient middle cerebral artery occlusion atlas identifies distinct subclusters of oligodendrocyte precursor cells(A) The transient middle cerebral artery occlusion (tMCAO) atlas comprises 106,905 cells from 21 oligodendrocyte precursor cell (OPC)-containing samples across six studies.(B) OPCs were extracted based on the expression of Sox10, Olig2, Pdgfra, and Cspg4 genes.(C) Dimensional reduction using uniform manifold approximation and projection (UMAP) reveals distinct OPC clusters, including cluster 4 at day 3 post tMCAO and cluster 3 at day 14 post tMCAO.(D) The proportion of cluster 4 increases on day 3 post tMCAO, whereas cluster 3 becomes more prominent on day 14 post tMCAO. Abbreviations: B, B cells; CP, choroid plexus cells; Cir.Myeloid, circulating myeloid cells; Endothelial, endothelial cells; Ependymal, ependymal cells; Mφ, macrophages; NP, neural progenitor cells; T & NK, T cells and NK cells.
Transcriptomic characterization of angiogenic OPCs on day 3 and oligogenic OPCs on day 14 after tMCAO
We identified OPCs based on the expression of Pdgfrα, Cspg4, Sox10, and Olig2 genes and primarily focused on OPCs for subsequent analyses (Figure 1B). Notably, we noted two distinct clusters: one that increased on day 3 after tMCAO (cluster 4) and another that increased on day 14 after tMCAO (cluster 3) (Figures 1C and 1D). Importantly, neither cluster 4 nor cluster 3 expressed genes specific to non-OPC cell types (Figure S1A), and both were consistently detected in samples at 3 and 14 days post tMCAO, respectively (Figure S1B), indicating that these clusters likely represent significant time-course-specific biological changes rather than batch effects.
To investigate the overall functional properties of these distinct clusters, we conducted gene set scoring analysis using pathways retrieved from the Molecular Signatures Database (Castanza et al., 2023), including Hallmark gene sets, Gene Ontology gene sets, and WIKI pathways gene sets. Additionally, to infer transcription factor activity, we used the CollecTRI database for transcription factor regulons (Müller-Dott et al., 2023). Finally, to examine OPC-specific functions, we manually curated gene sets, including OPC migration/myelination-associated gene sets (Hamanaka et al., 2023) and OPC-specific HIF-1α target gene sets (Allan et al., 2021; Hamanaka et al., 2023). We also extracted signature of WNT-activated OPCs via reanalyzing microarray datasets of mouse WNT-activated OPCs from Olig2cre-DA-cat mice (Fancy et al., 2009; 2014; Tsai et al., 2016), in which dominant-active β-catenin allows constitutive activation of the WNT pathway only in oligodendrocyte lineage cells.
These analyses revealed that OPCs in cluster 4, a distinct population present 3 days post-tMCAO, specifically upregulate HIF-1α activity, as evidenced by increased expression of both canonical and OPC-specific HIF-1α target genes (Figure 2A), including marked upregulation of the Vegfa gene (Figure 2C). Consistent with previous reports on HIF-1α-instructed OPCs (Yuen et al., 2014), this cluster also exhibits heightened WNT pathway activity (Figure 2A), with a drastic increase in Cxcr4 expression (Figure 2C) (Tsai et al., 2016). Collectively, these findings collectively indicate that severe hypoxia plays a key role in inducing cluster 4 of OPCs.Figure 2. Bioinformatic analyses predict the functions of distinct OPC subclusters as angiogenic and oligogenic(A) Gene set scoring analyses revealed upregulated pathways in OPC subclusters compared to other OPCs outside clusters 3 and 4. Mann-Whitney U test with multiple comparisons adjusted using the Benjamini & Hochberg method, p < 0.05.(B) Gene set scoring analyses identified upregulated OPC-relevant functional properties in OPC subclusters compared to other OPCs outside clusters 3 and 4. Mann-Whitney U test with multiple comparisons adjusted using the Benjamini and Hochberg method, p < 0.05.(C) Vegfa and Cxcr4 were specifically upregulated in cluster 4 OPCs.(D) Mog and Mbp were specifically upregulated in cluster 3 OPCs.(E and F) Cell-cell interaction analyses revealed that cluster 4 OPCs act as critical signaling sources in the WNT and VEGF signaling networks, with endothelial cells serving as key recipients.(G) The CXCL12-CXCR4 axis was specifically upregulated from endothelial cells and fibroblasts to cluster 4 OPCs.(H and I) Immunohistochemistry demonstrated upregulated HIF-1α activity and CXCR4 expression in subsets of OPCs accumulating at the ischemic border. Scale bars, 50 μm.
Notably, cluster 4 also exhibits specific upregulation of genes involved in the inflammatory response (Figure 2A) and is predicted to be influenced by cytokines, such as interleukin-1β (IL-1β), tumor necrosis factor alpha (TNF-α), and interferon-γ (IFN-γ), which have been widely implicated in the pathological processes during the acute phase of ischemic stroke (Doll et al., 2014; Li et al., 2001) (Figure S1C). This suggests that inflammation associated with the acute phase of ischemic stroke may also modify the characteristics of this cluster.
Functionally, cluster 4 upregulated genes associated with OPC migration (Figure 2B). In line with the characteristics of HIF-1α-instructed OPCs observed in the developing brain (Allan et al., 2021; Yuen et al., 2014), OPCs in cluster 4 appeared to halt gene programs related to oligodendrocyte differentiation and subsequent myelination (Figure 2B), as evidenced by the reduced expression of myelination-associated Mbp and Mog genes (Figure 2D). Instead, the defining feature of OPCs in this cluster appears to be their active involvement in angiogenesis (Figure 2B). Consistently, cell-cell interaction analyses revealed that OPCs in this cluster closely associate with endothelial cells, acting as a signaling source through the WNT and vascular endothelial growth factor (VEGF) pathways (Figures 2E and 2F), both of which are essential for OPC-driven angiogenesis in the developing brain (Allan et al., 2021; Yuen et al., 2014). Conversely, endothelial cells appear to attract OPCs in cluster 4 via the CXCL12-CXCR4 axis (Figure 2G), a key pathway for the close anatomical association between OPCs and endothelial cells in the developing brain (Tsai et al., 2016). It is also noteworthy that, even in comparison to other cell types, OPCs in cluster 4 represent a significant source of Vegfa at 3 days after tMCAO (Figure S1E), alongside astrocytes and fibroblasts, and are the primary source of Wnt7a at 3 days after tMCAO (Figure S1F), both of which are key factors in post-stroke angiogenesis (Hu et al., 2024). Altogether, we concluded that cluster 4 represents the OPCs primarily instructed by HIF-1α, resembling the angiogenesis-inducing OPCs previously characterized in the developing brain (Allan et al., 2021; Minocha et al., 2015; Tsai et al., 2016; Yuen et al., 2014), with additional functional modulation likely influenced by inflammation. We therefore annotated cluster 4 as angiogenic OPCs.
To validate the presence and investigate the spatial distribution of angiogenic OPCs, we next conducted immunofluorescence analysis 3–5 days post tMCAO. Nuclear HIF-1α signals were observed in a subset of platelet-derived growth factor receptor alpha (PDGFRα)-positive cells exhibiting morphological characteristics consistent with OPCs at the peri-infarct region (Figure 2H). Additionally, in the peri-infarct cortex, we identified cells double-positive for VEGFA and OLIG2 (Figure S2A), as well as cells double-positive for CXCR4 and PDGFRα (Figure 2I)—a combination that effectively distinguishes angiogenic OPCs from other cell types (Figures S1E, S1G, and S1H)—but not in the ischemic core or in the peri-infarct corpus callosum with its adjacent regions (Figure S2B).
Interestingly, OPCs in cluster 3, a distinct population observed 14 days post tMCAO, exhibit functional characteristics that are markedly different from the angiogenic OPCs seen at 3 days post tMCAO. This cluster appears to be only mildly affected by hypoxia, moderately upregulates OPC-specific HIF-1α target genes, and slightly increases the expression of angiogenesis-related genes (Figure 2A). However, its defining feature is the elevated expression of genes involved in oligodendrocyte differentiation and myelination (Figure 2B), such as Mbp and Mog genes (Figure 2D), accompanied by pronounced cellular renewal through cell division (Figure 2B). Prominently, Myrf, a key transcription factor for oligodendrocyte myelination and myelin maintenance (Emery et al., 2009; Qian et al., 2021), and Bcas1, a marker of the active phase of oligodendrocyte generation and myelination, are specifically upregulated in OPCs in cluster 3 (Fard et al., 2017) (Figure S1D). Additionally, these cells are predicted to be influenced not only by acute-phase cytokines like IL-1β, TNF-α, and IFN-γ but also by cytokines more associated with the chronic phase, such as IL-10 and transforming growth factor β (Doll et al., 2014; Hu et al., 2024) (Figure S1C). Based on these observations, we annotated cluster 3 as oligogenic OPCs, which are likely to play an integral role in remyelination following the acute stage of ischemic stroke. This role appears to be shaped by a milder hypoxic environment than that of angiogenic OPCs, as well as the influence of chronic-phase cytokines.
We next turned to immunohistochemical analyses to confirm the presence of oligogenic OPCs in vivo and assess their spatial distribution 14 days post tMCAO. In contrast to angiogenic OPCs, BCAS1^+^OLIG2^+^ oligogenic OPCs were more prominently increased in the peri-infarct corpus callosum with its adjacent regions, relative to the peri-infarct cortex, at day 14 post tMCAO (Figures S2C–S2E). These results confirmed the presence of the identified OPC subtypes and further indicated that the temporal transitions of OPC phenotypes after ischemic stroke are coupled with distinct spatial distributions, with angiogenic OPCs predominantly localized to peri-infarct cortical regions at day 3 and oligogenic OPCs enriched in peri-infarct white matter at day 14.
Interestingly, trajectory analysis predicted two distinct differentiation trajectories (Figures S2F and S2G). The first follows an angiogenic trajectory, culminating in angiogenic OPCs (Figure S2H), while the second represents an oligogenic trajectory, leading to oligogenic OPCs (Figure S2I). In both trajectories, key genes characteristic of each OPC population show progressively increased expression as differentiation advances (Figure S2J).
Ex vivo induction of HIF-1α-instructed angiogenic OPCs via severe hypoxic preconditioning
Encouraged by the in vivo detection of HIF-1α-instructed angiogenic OPCs after tMCAO, we reanalyzed bulk RNA sequencing (RNA-seq) data of OPCs subjected to severe hypoxic preconditioning ex vivo (severe-hypo-OPCs), previously generated by our group (Figure 3A) (Kishida et al., 2019).Figure 3Ex vivo OPCs acquire functional characteristics similar to in vivo angiogenic OPCs following severe hypoxic preconditioning(A and B) Bulk RNA-seq-based differentially expressed gene (DEG) analysis revealed marked transcriptional changes in ex vivo severe hypoxia-preconditioned OPCs (severe-hypo-OPCs), including significant upregulation of Vegfa and Cxcr4 and hallmark genes of in vivo angiogenic OPCs. Red points indicate significantly different genes identified using a linear model with multiple comparisons adjusted by the Benjamini and Hochberg method, with significance set at p < 0.05 and log_2_ fold change >2. n = 3 (control, biological replicates) and n = 3 (0.1%, biological replicates).(C–H) Gene set enrichment analysis (GSEA) revealed increased hypoxia and HIF-1α pathway activities (C), angiogenesis pathway activities (D), and WNT pathway activities (E); decreased OPC differentiation toward oligodendrocytes (F); decreased myelination-associated gene expression (G) in severe-hypo-OPCs, faithfully mirroring the functional characteristics of in vivo angiogenic OPCs. However, no upregulation of the inflammatory response was detected (H).(I) Correlation analysis revealed a significant positive correlation of log_2_ fold changes (log_2_FC) between in vivo angiogenic OPCs and ex vivo severe-hypo-OPCs.(J) Gene set scores calculated from DEGs in ex vivo severe-hypo-OPCs were specifically upregulated in cluster 4 OPCs in the transient middle cerebral artery occlusion (tMCAO) atlas.(K) Illustration showing that ex vivo severe-hypo-OPCs faithfully recapitulate the characteristics of in vivo angiogenic OPCs, which emerge at the peri-infarct region 3 days post tMCAO. FiO_2_, fraction of inspiratory oxygen; NES, normalized enrichment score; Hallmark, Hallmark gene sets; CollecTRI, CollecTRI database; DA-cat, dominant-active β-catenin; GOBP, Gene Ontology Biological Process; WikiPathways; rho, Spearman’s rank correlation coefficient.
First, we aimed to confirm that our ex vivo OPC cultures faithfully reflect the transcriptional characteristics of in vivo OPCs in the murine brain. Analysis of scRNA-seq data from murine oligodendrocyte lineage cells (Marques et al., 2016) demonstrated that our ex vivo OPCs closely resemble the transcriptional profile of in vivo OPCs, although some cells exhibit signs of differentiation into committed oligodendrocyte precursors and newly formed oligodendrocytes (Figure S3A). These findings validate our ex vivo OPC cultures as a suitable model for evaluating functional changes in OPCs in vivo.
Crucially, differentially expressed gene (DEG) analysis revealed substantial transcriptional changes within severe-hypo-OPCs compared with standard OPCs, including significant upregulation of Vegfa and Cxcr4 (Figure 3B). As expected, gene set enrichment analysis (GSEA) suggested that hypoxia-induced changes underlie these marked transcriptional alterations, and HIF-1α activation played an orchestrating role (Figure 3C). Notably, severe-hypo-OPCs dramatically upregulated angiogenesis activity (Figure 3D) and WNT pathway activity (Figure 3E), along with apparent downregulation of genes associated with oligodendrocyte differentiation and subsequent myelination (Figures 3F and 3G), mirroring the functional characteristics of in vivo angiogenic OPCs detected 3 days post tMCAO. Although we did not observe upregulation of inflammatory response genes in severe-hypo-OPCs (Figure 3H), correlation analyses between the log2-fold changes of DEGs in these ex vivo severe-hypo-OPCs and those in the in vivo angiogenic OPCs revealed significant positive correlations (rho = 0.30, Figure 3I). Given that cross-platform correlations between scRNA-seq and bulk RNA-seq log_2_ fold changes are typically around rho ≈ 0.6 even for identical samples—primarily due to the substantial difference in sequencing depth between the two platforms (Liu et al., 2023)—the observed correlation of rho = 0.30 should be interpreted as biologically relevant. Furthermore, the gene set score calculated from DEGs in ex vivo severe-hypo-OPCs confirmed a marked increase in the coordinated upregulation of these DEGs in in vivo angiogenic OPCs (Figure 3J).
To further investigate the contribution of the inflammatory response in shaping in vivo angiogenic OPCs, we reanalyzed bulk RNA-seq data from OPCs treated with IFN-γ (pro-inflammatory) or dexamethasone (DEX; anti-inflammatory) (Meijer et al., 2022). In both conditions, there were no changes in the expression of Vegfa and Cxcr4 (Figures S3B and S3C), indicating that inflammation is unlikely to be the main driver of gene expression changes in in vivo angiogenic OPCs. Nevertheless, IFN-γ treatment did suppress the expression of myelination-associated genes (Figures S3D and S3F), and the gene expression changes observed in ex vivo IFN-γ- and DEX-treated OPCs showed weak positive and negative correlations, respectively, with those observed in in vivo angiogenic OPCs (Figures S3E and S3G). Collectively, we concluded that ex vivo severe-hypo-OPCs faithfully replicate the functional characteristics of in vivo angiogenic OPCs detected 3 days post tMCAO, although additional modifications influenced by inflammatory responses may further alter the properties of in vivo angiogenic OPCs (Figure 3K).
Transplantation of severe-hypo-OPCs effectively ameliorated tMCAO outcome through increasing angiogenesis
Considering the well-known benefits of angiogenesis (Fang et al., 2023; Kanazawa et al., 2019; Krupinski et al., 1994), we hypothesized that in vivo angiogenic OPCs play a protective role. Encouraged by the finding that ex vivo severe-hypo-OPCs exhibit similar functional characteristics to in vivo angiogenic OPCs, we transplanted these severe-hypo-OPCs and compared their efficacy with standard OPCs. The angiogenic OPCs emerge at 3 days post tMCAO, and angiogenesis is reported to be detectable around 3–4 days after tMCAO (Fang et al., 2023; Kanazawa et al., 2019; Krupinski et al., 1994); therefore, we chose to administer severe-hypo-OPCs at 3 days post tMCAO. While previous OPC transplantation studies have primarily employed intracranial injection (Chen et al., 2015; Li et al., 2021; Wang et al., 2022), we opted for intravenous transplantation due to the following reasons: (1) severe-hypo-OPCs are likely to interact readily with brain endothelial cells and fibroblasts through the CXCR4-CXCL12 axis (Figure 2G); (2) the tMCAO atlas indicates upregulation of Cxcl12 expression in brain endothelial cells and fibroblasts at 3 days post tMCAO (Figure S4A); and (3) brain endothelial cells express Cxcl12 at higher levels than endothelial cells from other organs, as shown by public scRNA-seq data on multi-organ endothelial cells (Figures S4B and S4C) (Bondareva et al., 2022). Importantly, intravenous injection also has far greater clinical translation potential compared to intracranial injection.
We administered a retro-orbital injection of 7.5 × 10^5^ CAG-enhanced green fluorescent protein (EGFP)-tagged severe-hypo-OPCs suspended in 5 μL PBS (severe-hypo-OPC group), CAG-EGFP-tagged standard OPCs suspended in 5 μL PBS (standard OPC group), or 5 μL PBS alone (control group) on day 3 post tMCAO. Neurological function was assessed over time, and on day 14, we measured the percentage of infarcted areas in each mouse, a commonly used endpoint around the time when brain repair processes typically activate (Clarkson et al., 2010; Shi et al., 2021a) (Figure 4A). At 14 days after transplantation, co-staining with microtubule-associated protein-2 (MAP-2) revealed that GFP+-transplanted OPCs were distributed not only within the ischemic core and border but also extended into the peri-infarct penumbra, indicating that the transplanted OPCs survived and remained engrafted in the host tissue at day 14 (Figure S4D). Neurobehavioral tests revealed that the severe-hypo-OPC group exhibited the greatest recovery compared to the standard OPC and control groups. This was evident both in modified neurological severity scores (mNSS) (at day 14: −2.17 ± 0.41 [severe-hypo-OPC] vs. −1.33 ± 0.52 [standard OPC] vs. 0.00 ± 0.60 [control]) and in the Rotarod test (time to fall at day 14: 19.83 ± 4.49 [severe-hypo-OPC] vs. 11.33 ± 2.34 [standard OPC] vs. 8.50 ± 3.34 [control]) (Figure 4B). Critically, the severe-hypo-OPC group exhibited the smallest infarcted area among the three groups (% infarcted area at day 14: 10.57 ± 6.36 [severe-hypo-OPC] vs. 18.28 ± 8.80 [standard OPC] vs. 32.02 ± 14.74 [control]).Figure 4. Intravenous transplantation of severe-hypo-OPCs facilitates post-tMCAO recovery by promoting angiogenesis(A) Intravenous (i.v.) transplantation of OPCs (severe-hypo-OPCs, standard OPCs, and PBS) was performed on day 3 post tMCAO. Serial neurobehavioral assessments and histological quantifications of the infarcted area were subsequently conducted.(B) Neurobehavioral tests confirmed accelerated recovery in the severe-hypo-OPC group. Circles indicate the mean, and error bars represent the standard deviation. Mann-Whitney U test with multiple comparisons adjusted using the Benjamini and Hochberg method, p < 0.05; n = 12 (control, biological replicates), n = 6 (severe-hypo-OPC, biological replicates), n = 6 (standard OPC, biological replicates).(C) MAP-2 immunofluorescence-based quantification of the infarcted area showed a reduced infarct size in the severe-hypo-OPC group. The yellow dashed line indicates the infarcted area. The tops of the bars represent the mean, and error bars indicate the standard deviation. Mann-Whitney U test with multiple comparisons adjusted using the Benjamini and Hochberg method, p < 0.05; n = 6 (control, biological replicates), n = 6 (severe-hypo-OPC, biological replicates), n = 6 (standard OPC, biological replicates).(D) The number of GST-π-positive cells remained unchanged between the control (phosphate-buffered saline [PBS]) and severe-hypo-OPC groups. The tops of the bars indicate the mean, and the error bars indicate the standard deviation. Scale bars, 50 μm. n = 3 (severe-hypo-OPC), n = 3 (standard OPC).(E) The number of branches in blood vessels was significantly higher in mice intravenously injected with severe-hypo-OPCs. The tops of the bars indicate the mean, and the error bars indicate the standard deviation. Scale bars, 50 μm. Mann-Whitney U test, ^∗^p < 0.05; n = 3 (severe-hypo-OPC), n = 3 (standard OPC).
Immunofluorescence analyses revealed that transplanted severe-hypo-OPCs expressed PDGFRα (Figure S4E) but did not express O4 or glutathione-S transferase (GST)-π (Figure S4F), suggesting that they remained immature and did not differentiate into oligodendrocytes. Approximately half of the transplanted severe-hypo-OPCs colocalized with ionized calcium-binding adaptor molecule-1 (Iba-1)-positive cells (Figure S4E), indicating that they were phagocytosed by microglia. Regarding the therapeutic mechanism of transplanted severe-hypo-OPCs, immunofluorescence showed no change in the number of GST-π-positive cells (Figure 4D), but vascular density and branching were significantly increased following severe-hypo-OPC transplantation (Figure 4E). Thus, these findings collectively indicated that intravenous transplantation of severe-hypo-OPCs efficiently reduced infarct size and improved stroke outcomes primarily by promoting angiogenesis rather than oligodendrogenesis.
Oxygen tone contributes to regulating OPC phenotypic change during the tMCAO time course
Although the hypoxic response in in vivo oligogenic OPCs appeared lower compared to angiogenic OPCs, we still observed a marginally higher hypoxic response in in vivo oligogenic OPCs (cluster 3), as evidenced by OPC-specific HIF-1α activity (Figure 2A). This prompted us to investigate whether varying oxygen levels might differentially regulate OPC phenotypes and specifically contribute to the induction of in vivo oligogenic OPCs. To test this, we adjusted the oxygen concentration (2%, 3%, 5%, 10%, and 15% FiO_2_) in our ex vivo OPC cultures and quantified MBP protein levels as a marker of OPC maturation (Figure 5A). Strikingly, MBP protein levels varied significantly with oxygen concentration, with 3%–5% oxygen most effectively increasing MBP levels (Figures 5B and 5C), indicating that mild hypoxic preconditioning may enhance OPC maturation ex vivo.Figure 5. Mild hypoxic preconditioning directs ex vivo OPCs toward oligogenic OPCsTo examine how mild hypoxia affects OPC maturation, we conducted MBP western blotting (WB), immunostaining, and bulk RNA-seq, comparing ex vivo mild hypoxia-preconditioned OPCs (mild-hypo-OPCs) with in vivo oligogenic OPCs.(A) Experimental design overview.(B) WB analysis showed that FiO_2_ levels of 3%–5% significantly increased MBP expression. Error bars represent standard deviation. Mann-Whitney U test with multiple comparisons adjusted using the Benjamini and Hochberg method, p < 0.05; n = 5 (control in 2%, biological replicates), n = 5 (2%, biological replicates), n = 6 (control in 3%, biological replicates), n = 6 (3%, biological replicates), n = 4 (control in 5%, biological replicates), n = 4 (5%, biological replicates), n = 3 (control in 10%, biological replicates), n = 3 (10%, biological replicates), n = 4 (control in 15%, biological replicates), n = 4 (15%, biological replicates).(C) Immunostaining confirmed robust MBP induction at 5% FiO_2_.(D) Bulk RNA-seq-based differentially expressed gene (DEG) analysis revealed minimal transcriptomic changes in ex vivo mild-hypo-OPCs. n = 3 (per group, biological replicates).(E–J) Gene set enrichment analysis (GSEA) showed increased activity in general hypoxia-related pathways (E), but no upregulation of HIF-1α signaling (F), angiogenesis (G), or WNT signaling (H). Importantly, GSEA revealed enhanced OPC differentiation toward oligodendrocytes (I), consistent with an oligogenic phenotype, without increases in mitotic activity (J).(K) DEG log_2_ fold changes between in vivo oligogenic OPCs and ex vivo mild-hypo-OPCs were weakly but significantly correlated.(L) Gene set scoring analysis in the transient middle cerebral artery occlusion (tMCAO) atlas showed a marginally increased DEG score of ex vivo mild-hypo-OPCs in oligogenic OPCs. NES, normalized enrichment score; Hallmark, Hallmark gene sets; DA-cat, dominant-active β-catenin; Reactome, Reactome pathway database.
To confirm whether mild hypoxic preconditioning truly accelerates OPC maturation and to determine whether the transcriptomic changes resemble those of in vivo oligogenic OPCs, we conducted bulk RNA-seq analysis of these mild hypoxic-preconditioned OPCs (mild-hypo-OPCs). DEG analysis revealed minimal overall gene expression changes compared to those induced by severe hypoxic preconditioning (Figure 5D). GSEA and DEG analyses relative to standard OPCs showed that mild hypoxic preconditioning induced subtle hypoxia-associated transcriptomic changes (Figure 5E) but did not significantly increase HIF-1α activity (Figure 5F), angiogenesis (Figure 5G), or WNT pathway activity (Figure 5H). Consistent with the observed increase in MBP protein levels (Figure 5B), GSEA analysis demonstrated a clear promotion of OPC differentiation toward oligodendrocytes (Figure 5I). Although no upregulation of cell cycle activity was observed in mild-hypo-OPCs (Figure 5J), correlation analysis of log2 fold changes in DEGs between mild-hypo-OPCs and in vivo oligogenic OPCs revealed a weak but significant positive correlation (rho = 0.10, Figure 5K). Furthermore, gene set scoring analysis showed a mild upregulation of DEG scores from ex vivo mild-hypo-OPCs in in vivo oligogenic OPCs (Figure 5L).
Consistent with our prediction from the tMCAO atlas that the characteristics of in vivo oligogenic OPCs may be influenced by cytokines (Figure 2A), correlation analysis of log2 fold changes in DEGs between ex vivo IFN-γ-treated OPCs, DEX-treated OPCs, and in vivo oligogenic OPCs revealed significant positive correlations for IFN-γ treatment and negative correlations for DEX treatment (Figure S5A). Cell-cell interaction analysis of the tMCAO atlas indicated increased interaction strength between oligogenic OPCs and various glial cells, including microglia, astrocytes, and neurons (Figure S5B), consistent with the notion that oligogenic OPCs may promote myelination through interactions with multiple cell types during the subacute and chronic phases (Shi et al., 2021a). Thus, these findings collectively suggest that while mild hypoxia likely contributes, at least partially, to the emergence of in vivo oligogenic OPCs in the chronic stage post tMCAO, mild hypoxia preconditioning alone was insufficient to faithfully recapitulate the characteristics of in vivo oligogenic OPCs in our ex vivo OPC cultures. Therefore, cytokines and interactions with other cell types also play a crucial role in the induction of in vivo oligogenic OPCs.
Discussion
In this study, by assembling and analyzing scRNA-seq data after tMCAO, we successfully transcriptionally characterized distinct OPC populations that preferentially emerged at specific time points following acute ischemic stroke. One population, termed angiogenic OPCs, appeared 3 days post tMCAO and was predicted to be induced by severe hypoxia. These OPCs appeared to be regulated by HIF-1α, promoting angiogenesis through the WNT and VEGF signaling pathways while also being attracted to endothelial cells via the CXCR4-CXCL12 axis. Notably, severe hypoxic preconditioning enabled ex vivo OPCs to acquire transcriptomes faithfully mirroring those of in vivo angiogenic OPCs. Importantly, transplantation of these ex vivo severe-hypo-OPCs effectively mitigated tMCAO-induced motor deficits and reduced infarct size, primarily through their pro-angiogenic activity. It is noteworthy that in the tMCAO model, at day 3–5 after stroke, the infarct is not yet fully matured; additional neuronal loss continues beyond this time point (Bellut et al., 2023; Buscemi et al., 2019). Therefore, it is reasonable to consider that OPC transplantation at day 3 may not directly rescue pan-necrotic tissue but could instead promote the initiation of repair processes, such as angiogenesis, thereby preventing ongoing secondary neuronal injury and facilitating tissue recovery. Another population, termed oligogenic OPCs, emerged 14 days post tMCAO and was predicted to contribute to myelination. Integration with ex vivo experiments suggested that mild hypoxic preconditioning partially contributed to the generation of in vivo oligogenic OPCs. Thus, our study highlights oxygen tone as a crucial determinant in OPCs acquiring protective phenotypes appropriate to specific time points after ischemic stroke, underscoring that a deep understanding and replication of this adaptive response are key to successfully developing OPC-based cell transplantation therapies.
Extensive previous research has documented the dynamic and complex biological changes occurring over the course of acute ischemic stroke, using various models, including the widely used tMCAO model. In the acute phase (within 24 h), there is a marked upregulation of inflammation- and apoptosis-related genes in the ischemic brain (Lu et al., 2004), driven by the infiltration of inflammatory cells, predominantly neutrophils and other innate immune cells (Cai et al., 2020), along with substantial neuronal loss (Buscemi et al., 2019; Garcia et al., 1995). In the subacute phase (3–7 days later), angiogenesis becomes detectable (Kanazawa et al., 2019). In the chronic phase, the infiltration of adaptive immune cells increases, and brain repair processes, such as remyelination, are activated (Ito et al., 2019; Shi et al., 2021a). Regarding OPCs, existing evidence indicates that OPCs proliferate, migrate toward the lesion, and accumulate in the peri-infarct region following cerebral ischemia (Bonfanti et al., 2017; Jiang et al., 2011). Notably, prior reports suggest that nearly all OPCs migrating into the peri-infarct region fail to differentiate or mature until the chronic phase (Bonfanti et al., 2017; Jiang et al., 2011), implying functional roles beyond myelination during the acute and subacute phases.
Despite this, surprisingly few studies have specifically examined the spatiotemporal transcriptional changes in OPCs after tMCAO. This lack of research likely stems from the fact that OPCs represent a relatively small and underrepresented cell population, making them challenging to study in isolation. Consequently, a meta-analysis approach, such as ours, provides a powerful strategy to uncover the dynamic changes within these rare cell populations (Oki et al., 2018; Zou et al., 2024). Importantly, our meta-analysis approach, for the first time at the transcriptomic level, highlights the remarkable adaptability of OPCs as they dynamically shift their functions—playing a critical role in angiogenesis during the subacute phase and contributing to remyelination during the chronic phase. Moreover, our study reveals that oxygen tone serves as a crucial factor sensed by OPCs to spatiotemporally acquire specific characteristics post stroke, a phenomenon partially shared with the developing brain (Allan et al., 2021; Yuen et al., 2014), to facilitate recovery after acute ischemic stroke. Critically, our tMCAO atlas revealed that these functional adaptations naturally occur in only a small subset of OPCs in the adult brain following ischemic stroke. This limited natural adaptation underscores the therapeutic potential of our proposed strategy—preconditioning OPCs ex vivo to induce “protective functional adaptations” before transplantation—positioning them as an ideal target for cell-based therapies aimed at enhancing stroke recovery. In this context, given that (1) both human and mouse studies have shown that post-stroke angiogenesis correlates with improved outcomes (Berkhemer et al., 2016; Greenberg, 2015), (2) our findings indicate that angiogenic OPCs are among the primary contributors to angiogenesis within brain-resident cells, and (3) severe hypoxic preconditioning ex vivo faithfully induced angiogenic OPCs, we propose that enhancing angiogenesis through the transplantation of ex vivo severely hypoxia-preconditioned angiogenic OPCs represents a theoretically robust strategy.
We have to acknowledge that although this study successfully characterized dynamic OPC functional adaptations at the transcriptomic level, many questions remain unanswered. The factors limiting the broader induction of angiogenic and oligogenic OPC phenotypes remain unclear. One possible contributor is inflammation, though other mechanisms may also play a role, warranting further investigation. Additionally, exploring the potential mutual transformation between angiogenic OPCs and oligogenic OPCs is particularly intriguing. Our results showed that transplanted angiogenic OPCs did not express maturation markers, even after entering the chronic phase post tMCAO, and trajectory analysis predicted distinct developmental paths for angiogenic and oligogenic OPCs.
Another unresolved question concerns the origin of angiogenic and oligogenic OPCs. Resident OPCs expand in the peri-infarct region, while subventricular zone (SVZ) progenitors can migrate into ischemic areas and generate oligodendrocyte lineage cells (Menn et al., 2006). Their relative contribution appears to be lesion dependent—peri-striatal infarcts involving more SVZ input—whereas distal cortical lesions may rely predominantly on resident OPCs, possibly with additional input from meningeal-derived OPCs (Maki et al., 2013). Our histochemical analyses showed that angiogenic OPCs preferentially reside in the peri-infarct cortex, whereas oligogenic OPCs were enriched at the peri-infarct corpus callosum with its adjacent regions, in proximity to the SVZ. These findings suggest that the origins of the two OPC subtypes may differ; however, rigorous lineage-tracing studies will be required to definitively resolve this question. Finally, we should acknowledge that although our study focused exclusively on tMCAO, other models—such as permanent middle cerebral artery occlusion (MCAO), thromboembolic occlusion, or photothrombosis—offer complementary strengths (Macrae, 2011; Matur et al., 2023), and future studies using these approaches will be valuable to confirm the external validity of our findings.
In conclusion, by assembling and analyzing scRNA-seq data, we delineated the temporal transcriptional changes of OPCs following ischemic stroke, identifying two functionally distinct populations: angiogenic OPCs, which support angiogenesis in the subacute phase, and oligogenic OPCs, which contribute to remyelination in the chronic phase. Furthermore, we successfully induced ex vivo OPCs with transcriptomic characteristics similar to angiogenic OPCs through severe hypoxic preconditioning. Importantly, intravenous transplantation of these hypoxia-conditioned OPCs significantly enhanced post-stroke recovery by promoting angiogenesis in a stroke mouse model. Finally, we demonstrated that mild hypoxia partially contributes to the generation of oligogenic OPCs. Thus, after acute ischemic stroke, OPCs sense oxygen tone and undergo phenotypic shifts that enable them to fulfill stage-specific reparative roles, thereby facilitating recovery. Understanding and harnessing these adaptations offer promising therapeutic avenues for ischemic stroke, beyond current treatments that primarily focus on rapid reperfusion.
Methods
Experimental design
We assembled publicly available scRNA-seq data from the in vivo tMCAO murine model to characterize OPC temporal transcriptomic change. Controlled laboratory experiments were then conducted, including immunohistochemical staining of the in vivo tMCAO murine model, ex vivo perturbations of OPCs, and their subsequent transplantation back into the in vivo tMCAO murine model.
tMCAO atlas
To identify relevant datasets, we searched the NCBI BioProject database on December 1, 2023, using the following search terms:
(“tMCAO” [All Fields] AND “single cell” [All Fields]) OR (“middle cerebral artery occlusion” [All Fields] AND “single cell” [All Fields]) OR (“tMCAO” [All Fields] AND “scRNAseq” [All Fields]) OR (“middle cerebral artery occlusion” [All Fields] AND “scRNAseq” [All Fields]).
Inclusion criteria required raw FASTQ data generated using the 10× Genomics Chromium platform to ensure uniform pipeline processing. Only datasets with multiple samples from multiple studies at the same time point were included to distinguish biological changes from batch effects. For eligible datasets, raw FASTQ files were mapped to the mouse reference transcriptome (mm10) using the pre-built reference from the 10× Genomics website (https://www.10xgenomics.com/) via the cellranger count command in CellRanger v.7.2.0. Each count matrix was analyzed using Seurat (v.5.0.1) in R (v.4.3.0), following best practices for scRNA-seq (Heumos et al., 2023). Detailed uniform quality control procedures and bioinformatics analyses for scRNA-seq data, as well as bulk RNA-seq data processing and analysis, are described in the supplemental methods.
Animals
Male C.B-17/Icr-+/+Jcl mice (10–12 weeks old, 18–33 g; Clea Japan, Tokyo, Japan, RRID: IMSR_JCL:JCL:mID-0004) were used. Our study exclusively examined male mice. It is unknown whether the findings are relevant for female mice.
tMCAO procedure
The standard intraluminal tMCAO method was employed (Kishida et al., 2019). Mice were anesthetized with 2%–4% isoflurane (FUJIFILM Wako, Osaka, Japan; catalog no. 099-06571) in a 50% N_2_O/50% O_2_ mixture. Body temperature was maintained at 37°C–38°C using a heating pad and lamp (UNIQUE MEDICAL, Tokyo, Japan; catalog no. ATC-101B-MS). A midline incision was made to expose the left common carotid artery, where a 0.22 mm Doccol filament (Doccol, Sharon, MA, USA; catalog no. 602256PK10) was inserted up to the middle cerebral artery origin. Reperfusion was established after 60 min by filament withdrawal. tMCAO success was confirmed by laser Doppler flowmetry (Omegawave, Tokyo, Japan; catalog no. FLO-C1), ensuring at least an 80% reduction in cerebral blood flow during ischemia. Of 28 mice, 20 underwent successful tMCAO, while 8 died postoperatively.
Immunohistochemistry
Mouse brains were collected after perfusion with PBS (Nacalai Tesque, Kyoto, Japan; catalog no. 14249-24) and 4% PFA (Nacalai Tesque; catalog no. 09154-85) and then fixed in 4% PFA for 24 h and preserved in 20% sucrose. Coronal sections (20 μm) were rinsed with PBS, blocked with 3% BSA (Nacalai Tesque; catalog no. 01281-26), and incubated overnight at 4°C in 0.3% BSA with primary antibodies, including the following:
- (1)Anti-MAP-2 (Proteintech; catalog no. 17490-1-AP; RRID: AB_2137880) at 1:500
- (2)Anti-PDGFRα (Bio-Techne; catalog no. AF1062; RRID: AB_2236897) at 1:200
- (3)Anti-glucose transporter-1 (GLUT-1) (Merck Millipore; catalog no. 07–1401; RRID: AB_1587074) at 1:200
- (4)Anti-GST-π (MBL; catalog no. 311-H; RRID: AB_591790) at 1:200
- (5)Anti-doublecortin (Abcam; catalog no. ab18723; RRID: AB_732011) at 1:200
- (6)Anti-Iba-1 (FUJIFILM Wako; catalog no. 011-27991; RRID: AB_2935833) at 1:200
After three PBS washes, sections were incubated for 1 hour at room temperature with secondary antibodies, including the following:
- (1)Anti-rabbit Alexa Fluor 594 (Thermo Fisher Scientific; catalog no. A-21207; RRID: AB_141637) at 1:500
- (2)Anti-goat Alexa Fluor 594 (Thermo Fisher Scientific; catalog no. A-11058; RRID: AB_2534105) at 1:500
The sections were washed again, mounted with DAPI (Thermo Fisher Scientific; catalog no. 62247), and imaged using a fluorescence microscope (KEYENCE BZ-X710, Keyence, Osaka, Japan) or a confocal microscope (Olympus FV1000, Olympus, Tokyo, Japan).
Isolation of primary oligodendrocyte lineage cells
OPCs were isolated as previously described (Kishida et al., 2019). Cerebral cortices from P1 or P2 wild-type Sprague-Dawley rats (Shimizu Laboratory Supplies, Kyoto, Japan; RRID:MGI:5651135) were dissected, minced, and digested into a single-cell suspension using 0.25% trypsin-EDTA (Thermo Fisher Scientific; catalog no.25200056) and DNase I (Sigma-Aldrich; catalog no. D5219-500μg) at 37°C for 15 min. After filtration through a 40-μm cell strainer (Corning; catalog no. 352340), cells were plated in poly-D-lysine-coated (Sigma-Aldrich; catalog no. P0421-100MG) flasks with DMEM (FUJIFILM Wako; catalog no. 043-30085), 1% penicillin/streptomycin (Nacalai Tesque; catalog no. 26253-84), and 20% heat-inactivated fetal bovine serum (Biosera; catalog no. FB-1365/500). Once confluent (∼10 days), microglia were removed by shaking at 220 rpm for 1 h at 37°C, followed by a medium change and overnight shaking (20 h). Non-adherent cells were plated on uncoated culture dishes for 1 h at 37°C to eliminate residual astrocytes and microglia. The remaining cells were seeded on poly-L-ornithine-coated (Sigma-Aldrich; catalog no. P3655-100MG) plates at 20,000 cells/cm^2^ in Neurobasal Medium (Thermo Fisher Scientific; catalog no. 21103049) with 1% penicillin/streptomycin, 2 mM glutamine (Nacalai Tesque; catalog no.16948-04), 10 ng/mL FGF-2 (Thermo Fisher Scientific; catalog no. PTI-100-18B-50), 10 ng/mL PDGF-AA (Thermo Fisher Scientific; catalog no.PTI-100-13A-10), and 2% B27 supplement (Thermo Fisher Scientific; catalog no.17504044). Six days post plating, OPCs underwent 0.1% oxygen-glucose deprivation for 6 h to induce severe hypoxia, followed by exposure to 2%, 3%, 5%, 10%, or 15% FiO_2_ for 6 days under mild hypoxia. Due to the high cytotoxicity of 0.1% oxygen-glucose deprivation, cells were only assessed at 6 h. For transplantation, hypoxic-preconditioned OPCs were dissociated using 5 mL Accumax (Innovative Cell Technologies; catalog no. AM105-500ML), and 7.5×10^5^ cells were suspended in 5 μL PBS.
OPC transplantation
tMCAO was performed in 32 mice, with 8 excluded due to death before day 3. The remaining 24 mice were randomly assigned on day 3 to one of three groups: control (n = 12), standard OPC (n = 6), and severe-hypo-OPC (n = 6). Under anesthesia, all mice received orbital injections as follows:
- (1)Control group: 5 μL PBS
- (2)Standard OPC group: 7.5 × 10^5^ CAG-EGFP-tagged standard OPCs in 5 μL PBS
- (3)Severe-hypo-OPC group: 7.5 × 10^5^ CAG-EGFP-tagged severe-hypo-OPCs in 5 μL PBS
All mice received intraperitoneal FK506 (1 mg/kg/day; FUJIFILM Wako; catalog no. 063-06071) and 500 μL normal saline daily from days 1–7 post tMCAO.
Neurobehavioral and infarct volume measurements
We assessed mNSS by a blinded evaluator on days 3, 7, 10, and 14, following a standardized 0–18 scale that includes motor, sensory, beam balance, and reflex tests. Rotarod tests were conducted on the same days using an accelerating rotarod (Ugo Basile; catalog no. 47650) set to increase from 0 to 40 rpm over 4 min. The average time to fall was recorded over two trials. Mice were sacrificed on day 14, and brains were sectioned into 20 μm coronal slices. Five sections (+1.0, +0.5, 0, −0.5, and −1.0 mm relative to bregma) were stained with anti-MAP-2. A blinded investigator captured images using a fluorescence microscope (KEYENCE BZ-X710) and analyzed them in TIFF format using ImageJ 1.53a. The infarct area was calculated as follows:
- (1)The contralateral hemisphere area (A) and the ipsilateral hemisphere area excluding the infarct zone (B) were measured.
- (2)Infarct area = (A − B)/A (%).
- (3)Infarct areas were multiplied by the section interval (0.5 mm) and summed to determine total infarct volume.
Evaluation of angiogenesis and oligodendrogenesis
Coronal brain sections (20 μm) were stained with anti-GLUT-1 (1:200) or anti-GST-π (1:200). Four penumbra regions surrounding the infarct were analyzed in sections at +0.5, 0, and −0.5 mm relative to the bregma. Images were captured at 20× magnification and analyzed using ImageJ. Angiogenesis was assessed by counting the total number of vessel branches in 12 regions (4 per section, 3 sections). Oligodendrogenesis was quantified by counting GST-π-positive oligodendrocytes in the same regions.
Evaluation of spatial distribution of angiogenic and oligogenic OPCs
Coronal brain sections (20 μm) were stained with anti-BCAS1 (1:200), anti-VEGFA (1:200), and anti-OLIG2 (1:200). Four penumbra regions surrounding the infarct were analyzed in sections at +0.5 mm relative to the bregma. Images were captured at 20× magnification and analyzed. On day 3 after MCAO, the proportion of VEGFA^+^ OLIG2^+^ angiogenic OPCs among total OLIG2^+^ oligodendrocyte lineage cells was compared between the peri-infarct cortex and the peri-infarct corpus callosum with its adjacent regions. On day 14 after MCAO, the proportion of BCAS1^+^ OLIG2^+^ oligogenic OPCs was compared between these regions.
Statistical analysis
Statistical analyses were primarily conducted by K.T., who is certified by the Japan Statistical Society (grade 2), using custom R scripts. The Mann-Whitney U test and Spearman’s rank correlation coefficients (rho) were used for between-group comparisons and correlation analyses, respectively, as the assumption of normal distribution was often not met. Throughout the study, multiple testing was adjusted using the Benjamini and Hochberg method. Two-sided p values <0.05 were considered statistically significant. Values are presented as mean ± standard deviation.
Study approval
All procedures adhered to Kyoto University’s animal experimentation guidelines and were approved by its Ethical Committee.
Resource availability
Lead contact
Further information and requests for resources and reagents should be directed to and will be fulfilled by the lead contact, Takakuni Maki ([email protected]).
Materials availability
All materials and lines generated in this study are available from the lead contact.
Data and code availability
- •All data are available in the main text or supplemental information including Supporting Data Values file. The MCAO atlas datasets—including both the full dataset and the OPC-only subset—have been deposited in the Single-Cell Portal under accession numbers SCP3078 (https://singlecell.broadinstitute.org/single_cell/study/SCP3078) and SCP3080 (https://singlecell.broadinstitute.org/single_cell/study/SCP3080), respectively.
- •All R code, statistical source data, and data used to generate the figures have been deposited in the Open Science Framework (https://osf.io/a3tc8/).
- •Sequence data generated in our study have been deposited in GEO under accession number [GSE275670](GSE275670).
Acknowledgments
The authors would like to thank other members of our department for their support. This work was funded by the 10.13039/501100001700Ministry of Education, Culture, Sports, Science and Technology Japan (Grant-in-Aid for Scientific Research C) 20K06853 (T.M.), 10.13039/501100002241Japan Science and Technology Agency (Moonshot R&D) JPMJMS2024 (T.M.), and 10.13039/501100001691Japan Society for the Promotion of Science (Grant-in-Aid for Young Scientists) JP22K18178 (K.T.). The graphical abstract preparation was supported by Editage.
Author contributions
Conceptualization, Y.K., K.Y., K.T., and T.M.; methodology, Y.K., K.Y., K.T., and T.M.; investigation, Y.K., K.Y., K.T., and T.M.; visualization, Y.K., K.T., and A.K.; funding acquisition, K.T. and T.M.; project administration, R.T. and T.M.; supervision, R.T. and T.M.; writing – original draft, Y.K., K.T., and T.M.; writing – review and editing, Y.K., K.T., and T.M.
Declaration of interests
The authors declare no competing interests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Akay L.A.Effenberger A.H.Tsai L.-H.Cell of all trades: oligodendrocyte precursor cells in synaptic, vascular, and immune function Genes Dev.35202118019810.1101/gad.344218.12033526585 PMC 7849363 · doi ↗ · pubmed ↗
- 2Allan K.C.Hu L.R.Scavuzzo M.A.Morton A.R.Gevorgyan A.S.Cohn E.F.Clayton B.L.L.Bederman I.R.Hung S.Bartels C.F.Non-canonical Targets of HIF 1a Impair Oligodendrocyte Progenitor Cell Function Cell Stem Cell 282021257272.e 1110.1016/j.stem.2020.09.01933091368 PMC 7867598 · doi ↗ · pubmed ↗
- 3Anthony S.Cabantan D.Monsour M.Borlongan C.V.Neuroinflammation, Stem Cells, and Stroke Stroke 5320221460147210.1161/STROKEAHA.121.03694835380050 PMC 9038685 · doi ↗ · pubmed ↗
- 4Baranova O.Miranda L.F.Pichiule P.Dragatsis I.Johnson R.S.Chavez J.C.Neuron-specific inactivation of the hypoxia inducible factor 1 alpha increases brain injury in a mouse model of transient focal cerebral ischemia J. Neurosci.2720076320633210.1523/JNEUROSCI.0449-07.200717554006 PMC 6672155 · doi ↗ · pubmed ↗
- 5Bellut M.Bieber M.Kraft P.Weber A.N.R.Stoll G.Schuhmann M.K.Delayed NLRP 3 inflammasome inhibition ameliorates subacute stroke progression in mice J. Neuroinflammation 202023410.1186/s 12974-022-02674-w 36600259 PMC 9811791 · doi ↗ · pubmed ↗
- 6Bergles D.E.Richardson W.D.Oligodendrocyte Development and Plasticity Cold Spring Harbor Perspect. Biol.82015 a 02045310.1101/cshperspect.a 020453 · doi ↗
- 7Berkhemer O.A.Jansen I.G.H.Beumer D.Fransen P.S.S.van den Berg L.A.Yoo A.J.Lingsma H.F.Sprengers M.E.S.Jenniskens S.F.M.Lycklama À Nijeholt G.J.Collateral Status on Baseline Computed Tomographic Angiography and Intra-Arterial Treatment Effect in Patients With Proximal Anterior Circulation Stroke Stroke 47201676877610.1161/STROKEAHA.115.01178826903582 · doi ↗ · pubmed ↗
- 8Bondareva O.Rodríguez-Aguilera J.R.Oliveira F.Liao L.Rose A.Gupta A.Singh K.Geier F.Schuster J.Boeckel J.-N.Single-cell profiling of vascular endothelial cells reveals progressive organ-specific vulnerabilities during obesity Nat. Metab.420221591161010.1038/s 42255-022-00674-x 36400935 PMC 9684070 · doi ↗ · pubmed ↗