The roles of TGF-β, Wnt, and MAPK signaling pathways in joint lineage specification in vitro and ex vivo
Suyash Raj, Thomas Cutia, Stefano Menghini, Mireia Alemany-Ribes, Junming Cai, Mariel Young, Sarah K. Jachim, Terence D. Capellini, April M. Craft

TL;DR
Researchers created a model using stem cells to study how joint tissues develop, finding that specific signaling pathways control joint cell formation.
Contribution
A novel dual reporter mouse embryonic stem cell line was developed to study joint lineage specification and signaling pathways.
Findings
TGF-β signaling is necessary and sufficient for inducing joint markers Gdf5 and Prg4.
Inhibiting Wnt or MAPK signaling promotes joint lineage formation in vitro and ex vivo.
mESC-derived joint lineages are transcriptionally similar to primary limb bud cells.
Abstract
The initiation of synovial joint development and subsequent differentiation of progenitor cells toward anatomically and functionally distinct joint tissues are not well understood, despite being highly relevant to joint health and disease. We generated a dual reporter mouse embryonic stem cell (mESC) line to quantify cells expressing growth differentiation factor five (Gdf5), an early marker of joint formation, and Prg4, a lubricating proteoglycan found in joint tissues. Transforming growth factor β (TGF-β) signaling was necessary and sufficient for the induction of Gdf5-RFP and Prg4-GFP. Inhibition of either Wnt or MAPK signaling significantly increased the induction of Gdf5-RFP, while activation of either pathway prohibited this induction. Single cell transcriptomics demonstrated the chondrogenic identity of Gdf5+ cells in in vitro cultures and in mouse embryonic limb buds. We…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
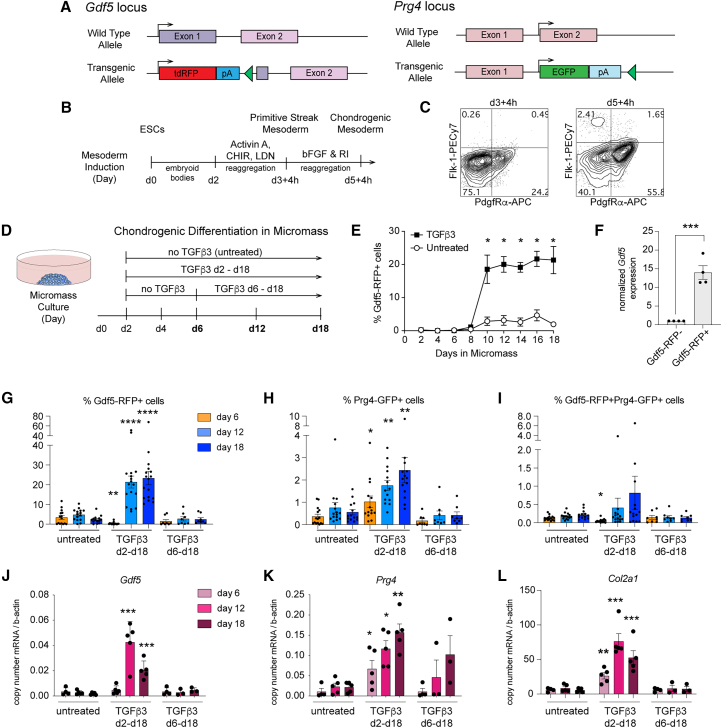 Figure 1
Figure 1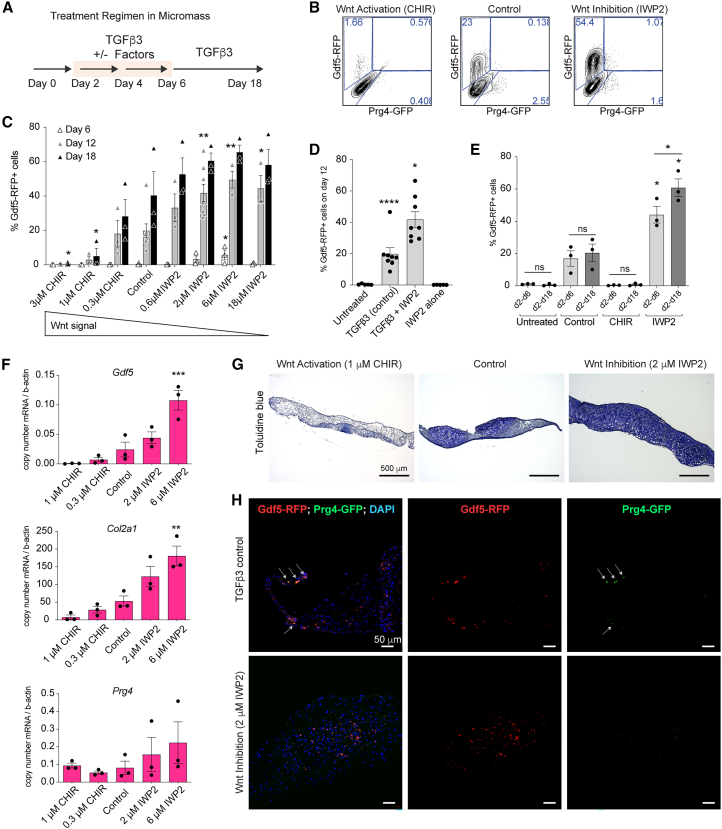 Figure 2
Figure 2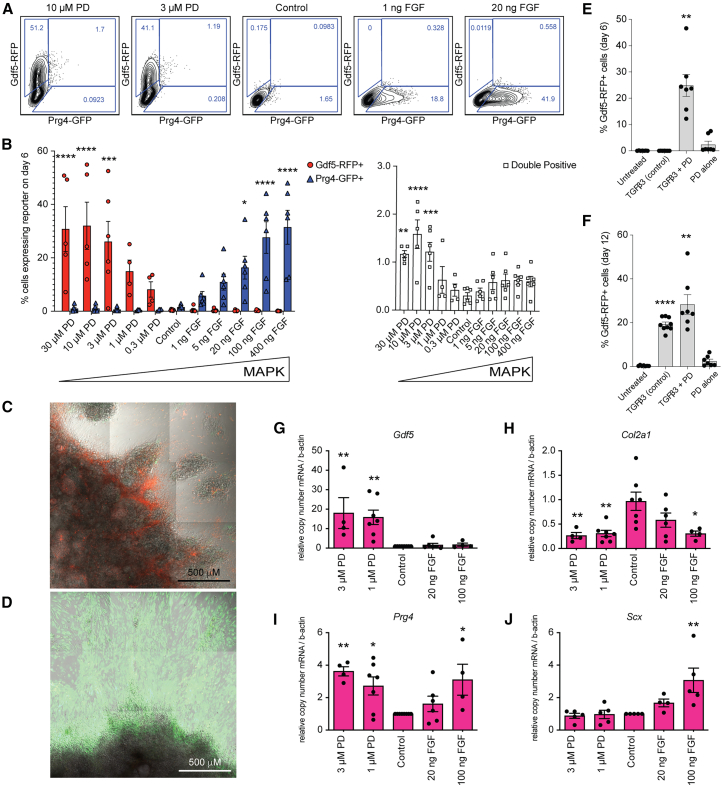 Figure 3
Figure 3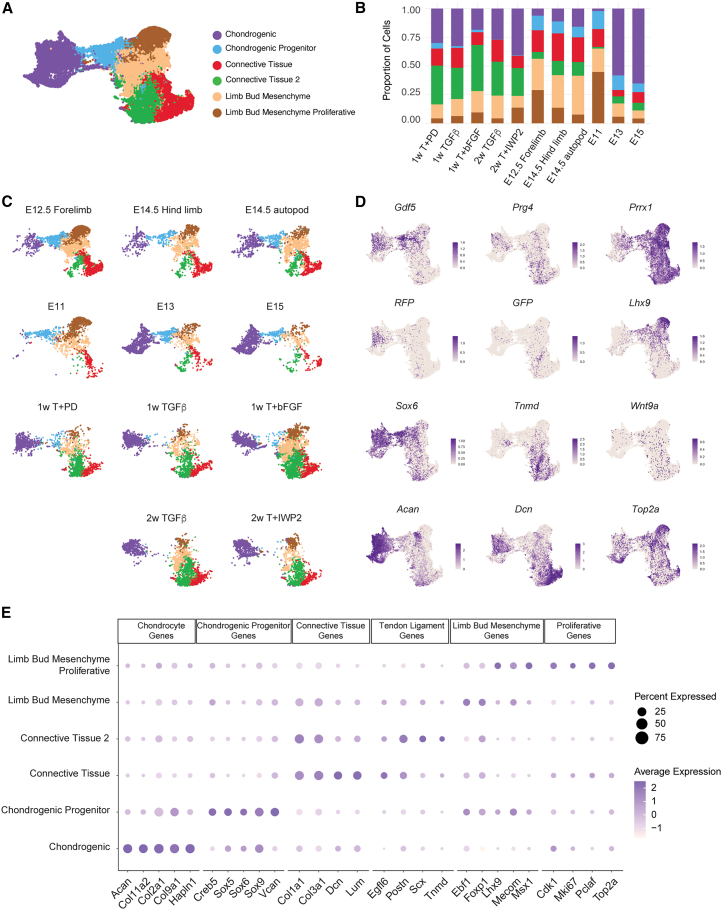 Figure 4
Figure 4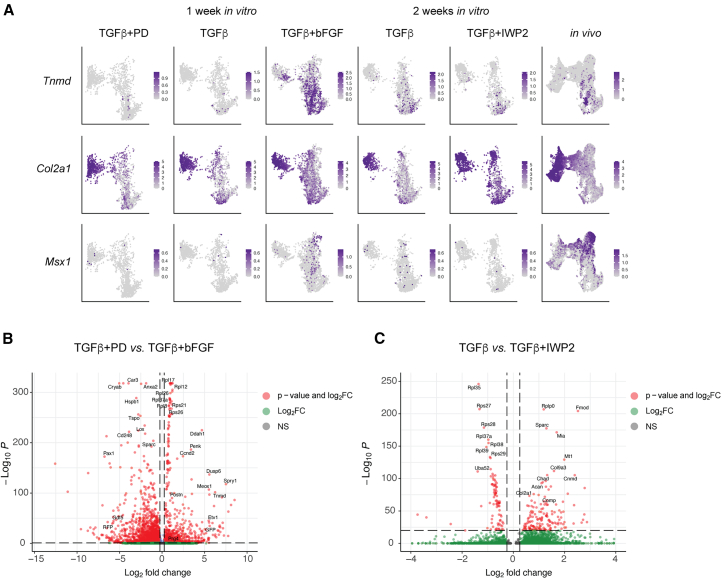 Figure 5
Figure 5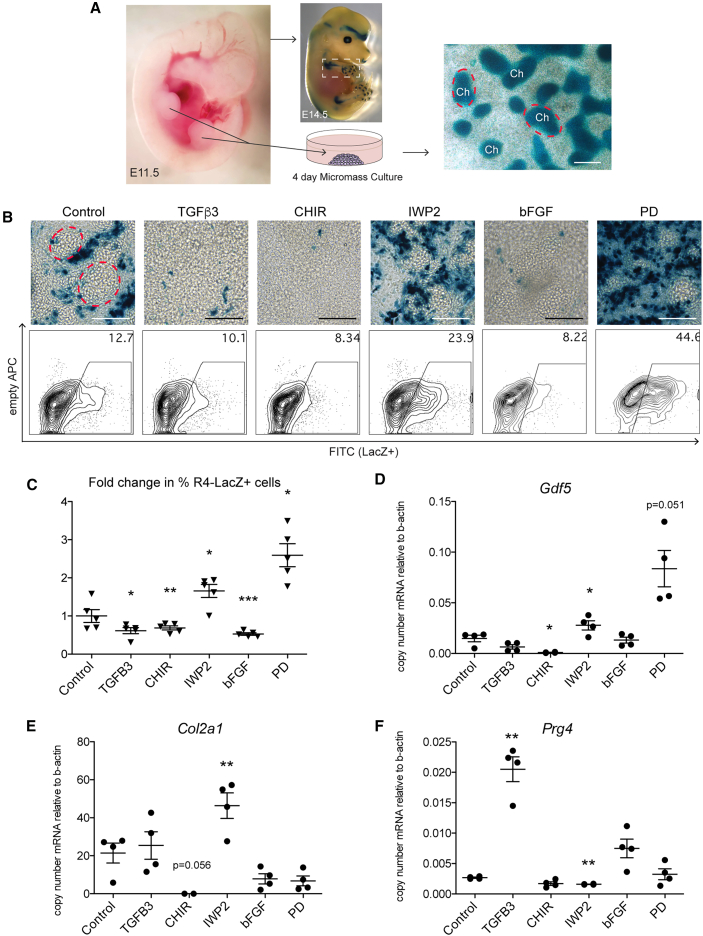 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCongenital heart defects research · Pluripotent Stem Cells Research · Cancer-related gene regulation
Introduction
Synovial joints provide smooth articulation crucial for vertebrate mobility and are found in both the axial and appendicular skeleton. During development, joints are initiated within regions of condensing chondrogenic mesenchymal structures through segmentation and emergence of the interzone. Interzone cells differentiate into chondrocytes that deposit extracellular matrix (ECM), resulting in the formation of articular cartilage, which lines the end of long bones providing a lubricating surface and resilience against mechanical load (Pacifici et al., 2006; Koyama et al., 2008). The cells of the interzone can be identified by the expression of growth differentiation factor five (Gdf5), a member of the transforming growth factor β (TGF-β) superfamily of proteins. As the joints develop, there continues to be an influx of Gdf5 positive cells that contribute to the formation of the synovial joint (Shwartz et al., 2016). Lineage tracing studies have confirmed that these *Gdf5-*expressing cell populations contribute to articular cartilage, the meniscus, ligaments, and the synovial lining. Cells at the surface of the articular cartilage continue to express Gdf5 for a short period of time postnatally, while other key proteins such as lubricin (encoded by the gene Prg4) persist throughout adulthood (Jay et al., 2000).
While elegant in vivo studies provide insights into the signaling pathways that might play a critical role during synovial joint development (Brunet et al., 1998; Hartmann and Tabin, 2001; Guo et al., 2004; Spagnoli et al., 2007), the signals that initiate the emergence of Gdf5-expressing cells to begin the process of joint initiation, and whether those signals also induce cells to differentiate into intra-articular Prg4-expressing cells, are less understood. TGF-β signaling was initially implicated in many human skeletal patterning disorders, and TGF-β superfamily members have long been known to promote the differentiation of chondrocytes. Spagnoli et al. identified that TGF-β signaling was not only essential to joint development in mice, but one receptor Tgfbr2 is specifically expressed in developing joints, and its deletion resulted in lack of joint interzone formation (Spagnoli et al., 2007). Consistent with this observation, the directed differentiation method we developed to generate articular cartilage tissues from human pluripotent stem cells (hPSCs), expressing both GDF5 and PRG4, also relies on TGF-β signaling in vitro (Craft et al., 2015; Li et al., 2023). In addition to TGF-β, it has been demonstrated in multiple in vivo models that joint development is altered in response to modulation of phospho-(p)ERK (Bastow et al., 2005), fibroblast growth factor (FGF) signaling (Wang et al., 2001), SOXC genes (Bhattaram et al., 2014), and Wnt9a/14 (Guo et al., 2004). Given the findings from these in vivo studies, we hypothesized that TGF-β, Wnt, and MAPK signaling pathways would influence the initiation and/or specification of joint lineage cells in vitro.
Studying joint development in vivo remains challenging due its transient nature, the minute quantity of cells available, and the high costs of generating and maintaining genetically modified animal models. Directed differentiation models using PSCs provide a means to overcome some of the limitations of in vivo models, and if directed properly with the correct signals, one could generate an unlimited amount of progenitor cells for downstream studies. While directed differentiation methods that distinguish between articular joint-lining and growth plate cartilage via lateral plate mesoderm remain limited (Smith et al., 2023), we and others have shown articular cartilage potential from PSC-derived paraxial mesoderm populations that are enriched in GDF5 expression (Craft et al., 2015; Pothiawala et al., 2022), consistent with the finding that the regulatory region of Gdf5 drives reporter expression in both synovial (e.g., costovertebral and costotransverse) and non-synovial (e.g., intervertebral) joints in the axial skeleton (Chen et al., 2016). These data support the potential use of PSCs for studying the signals that orchestrate joint initiation, and further for generating lineage-restricted intra-articular cells or tissues for replacement or repair.
We aimed to develop new tools and methods to define signals and culture conditions that are sufficient to induce expression of Gdf5, marking joint lineage cells, or Prg4, expressed by cells on the surfaces of intra-articular tissues. We generated a dual reporter mouse embryonic stem cell (mESC) line with fluorescent proteins expressed by endogenous Gdf5 and Prg4 regulatory elements. We found TGF-β signaling to be necessary and sufficient for the induction of Gdf5 expression. Inhibition of either Wnt or MAPK signals promoted the expression of Gdf5, while activation of either pathway diminished it. In contrast, activation of MAPK signaling significantly induced Prg4 expression. Joint lineage cells induced by combinations of TGF-β, Wnt, and MAPK modulators had similar transcriptomes to chondrogenic and connective tissue lineages isolated from mouse embryonic limb buds. Finally, we demonstrated similar upregulation of joint-enhancer driven Gdf5 expression upon pathway modulation in ex vivo limb bud cultures. These results highlight the utility of this novel reporter mESC line to investigate and quantify the specification of joint lineage cells, shed light on our understanding of this complex developmental process, and support the notion that there are shared mechanisms across joint cell specification in the axial and the appendicular skeleton.
Results
TGF-β signaling is sufficient for the induction of Gdf5-RFP expressing cells from mESC-derived mesoderm
To quantitatively track the emergence of joint lineage cells, fluorescent reporter genes were introduced into the Gdf5 (tdRFP) and Prg4 loci (eGFP) using CRISPR-Cas9 methodology, leaving one allele of each intact (Figures 1A and S1). The dual reporter mESC line was specified toward PdgfRa^+^Flk-1^−^ primitive streak mesoderm and chondrogenic mesoderm using methods we previously established (Figures 1B and 1C) (Craft et al., 2013). This induction protocol includes BMP inhibitory signals that are consistent with methods developed by us and others to induce paraxial mesoderm from PSCs (Craft et al., 2015; Pothiawala et al., 2022; Tani et al., 2023; Wu et al., 2021). Following mesoderm induction, we introduced the mESC-derived cells to high density micromass culture to induce chondrogenesis, adapting the protocols we established for differentiating hPSCs toward articular cartilage (Figure 1D) (Craft et al., 2015; Richard et al., 2023).Figure 1TGF-β signaling is sufficient for induction of Gdf5-RFP expressing cells from mESC-derived mesoderm in micromass culture(A) CRISPR-Cas9 was used to introduce tdRFP into the first exon of Gdf5 and EGFP into the first coding exon (exon 2) of Prg4 in the parental E14 mESC line. The untargeted alleles remain intact.(B) Methods to induce primitive streak mesoderm and chondrogenic mesoderm from mESCs.(C) Representative flow cytometry contour plots show expression of Flk-1 and PdgfRα on cells on day 3 and day 5 of differentiation.(D) Methods to induce chondrogenesis from day 5 mesoderm cells in micromass culture.(E) Percent of Gdf5-RFP expressing cells during micromass culture in the presence or absence of TGF-β3 (10 ng/mL) from day 2 to day 18, n = 3 independent experiments, Student’s t test. Values, mean ± SEM. ^∗^p < 0.05.(F) Normalized expression of Gdf5 mRNA following cell sorting of Gdf5-RFP+ and Gdf5-RFP+ cells from 4 independent experiments (cultured in the presence of TGF-β3). Values, mean ± SEM, Student’s t test, ^∗∗∗^p < 0.001.(G) Percentage of Gdf5-RFP+ cells on days 6, 12, and 18 in micromass following indicated treatments (illustrated in D). n = 8–15 biological replicates (independent experiments) represented in (G–I); values, mean ± SEM; ANOVA with Dunnett’s multiple comparison correction vs. untreated. ^∗^p < 0.05, ^∗∗^p < 0.01, ^∗∗∗^p < 0.001, and ^∗∗∗∗^p < 0.0001 vs. untreated.(H) Percentage of Prg4-GFP+ cells on days 6, 12, and 18 in micromass following indicated treatments. Icons represent independent experiments.(I) Percentage of Gdf5-RFP+;Prg4-GFP+ cells (double-positive) on days 6, 12, and 18 in micromass following indicated treatments. Icons represent independent experiments.(J–L) Expression of indicated genes quantified by RT-qPCR on days 6, 12, and 18 following indicated treatments. n = 4–6 independent experiments. Values, mean ± SEM. ANOVA with Dunnett’s multiple comparison correction versus untreated. ^∗^p < 0.05, ^∗∗^p < 0.01, and ^∗∗∗^p < 0.001 vs. untreated. See also Figure S1.
Joint morphogenesis in the limbs in vivo and articular chondrogenesis in vitro rely on TGF-β, thus, we hypothesized that exogenous TGF-β would be sufficient to induce Gdf5 and Prg4 expression. TGF-β3 was introduced to the micromass cultures after a deliberate 48 h grace period to promote cell adherence (Figures 1D and 1E). In the absence of TGF-β3, only 3%–5% of cells express Gdf5-RFP on day 10 and onwards (Figure 1E). In the presence of TGF-β3, Gdf5-RFP was detectable by day 8 in ∼2% of cells. By day 10, over 20% of the cells were expressing Gdf5-RFP, which remained relatively stable until day 18. Expression of Gdf5 mRNA was restricted to Gdf5-RFP+ cells following fluorescence-activated cell sorting (FACS) from TGF-β3-treated micromasses on day 12 (Figure 1F). To determine whether TGF-β is required to induce Gdf5-RFP in the early stage (days 2–6) of micromass culture versus late stage (days 6–18), we delayed the introduction of TGF-β3 in the cultures until day 6, which is prior to the onset of Gdf5-RFP expression in untreated cultures. Flow cytometry on day 6, day 12 (chosen as it captures the initial peak of Gdf5-RFP expression), and day 18 (culture endpoint) showed that TGF-β3 treatment initiating on day 6 did not upregulate the reporter, suggesting that TGF-β signaling was required between days 2 and 6 of micromass culture to sufficiently induce Gdf5-RFP (Figure 1G). Prg4-GFP was also induced and was significantly higher in the cultures treated with TGF-β3 from days 2–18 (Figure 1H). Similarly, double-positive cells were in higher abundance in TGF-β3-treated cultures and peaked at about 0.8% on day 18 (Figure 1I), but this was not statistically significant. Expression of Gdf5 and Prg4 (Figures 1J and 1K), as well as the cartilage matrix gene type II collagen (Col2a1, Figure 1L), followed similar patterns, being significantly higher in continuous TGF-β3 treatment cultures. Delayed treatment with TGF-β3 on day 6 was sufficient to observe Prg4-GFP, consistent with previous reports (Zhang et al., 2021; Khalafi et al., 2007) that this gene is responsive to TGF-β signaling.
We next investigated whether other members of the TGF-β superfamily, including Gdf5 itself, could induce Gdf5-RFP (Figures S1C–S1F). We treated micromass cultures with increasing doses of TGF-β3, activin A, BMP4, and Gdf5, as well as with a chemical inhibitor of each ligand (Figures S1C–S1F). TGF-β3 was the only ligand that significantly increased the percentage of Gdf5-RFP+, Prg4-GFP+, and Gdf5-RFP+; Prg4-GFP+ cells compared to untreated cells (Figures S1G–S1I). The induction of Gdf5-RFP was dose dependent with respect to TGF-β3 (Figures S1C and S1G). At 100 ng/mL of activin A, about 10% of cells expressed Gdf5-RFP (Figures S1F and S1G). Neither BMP4 nor Gdf5 were able to induce Gdf5-RFP at any concentration tested (Figures S1D and S1E).
Wnt inhibition in the presence of TGF-β significantly increases the percentage of Gdf5-RFP+ cells
Wnt signaling has been implicated in joint initiation in the limb (Guo et al., 2004), and inhibition of Wnt has been previously shown to improve chondrogenesis from PSCs by minimizing off-target cell types (Wu et al., 2021). We sought to define the role of Wnt in the induction of Gdf5-RFP and Prg4-GFP by using agonists or antagonists between days 2 and 6 of micromass culture (Figure 2A). In the absence of TGF-β3, neither the small molecule Wnt agonist CHIR99061 (CHIR; a GSK3β inhibitor) nor the Wnt antagonist IWP2 (PORCN inhibitor) induced Gdf5-RFP (data not shown, and Figure 2D). In the presence of TGF-β3, addition of CHIR resulted in a loss of Gdf5-RFP+ cells, while inhibition of Wnt with IWP2 significantly increased the percentage of Gdf5-RFP+ cells (up to 2-fold higher) on day 18 (Figures 2B and 2C). By day 12 the induction of Gdf5-RFP became dose dependent on IWP2, where concentrations of 2 μM or higher were significantly higher than TGF-β3 treatment alone (control). Conversely, as CHIR concentrations were increased, the percentage of Gdf5-RFP+ cells was significantly decreased compared to control. Early Wnt inhibition (days 2–6) was sufficient to increase the proportion of Gdf5-RFP+ cells on day 18, while continued IWP2 treatment (days 2–18) led to a slight but significant increase compared to early Wnt inhibition (Figure 2E), suggesting that Wnt inhibition was effective at promoting Gdf5-RFP throughout the differentiation. Similar responses in the percentage of Gdf5-RFP+ cells were observed following the addition of an alternative Wnt antagonist XAV939 or agonist Wnt3a (Figure S2A). Interestingly, the percentage of Prg4-GFP+ cells increased slightly in cultures treated with CHIR on day 18 compared to control, and the percentage of double-positive cells remained unchanged (Figures S2B and S2C).Figure 2. Inhibition of Wnt in the presence of TGF-β significantly increases the percentage of Gdf5-RFP expressing cells and chondrogenic potential in a dose-dependent manner(A) Treatment scheme of micromass cultures with Wnt pathway agonist (CHIR99021) and antagonist (IWP2).(B) Representative flow cytometry contour plots show expression of Gdf5-RFP and Prg4-GFP on day 18 of micromass culture.(C) Percentage of Gdf5-RFP expressing cells on days 6, 12, and 18 of micromass culture in the presence or absence of increasing concentrations of the agonist CHIR99021 or the antagonist IWP2. Values, mean ± SEM. n = 3–8 independent experiments, ANOVA with Dunnett’s multiple comparison correction versus control (TGF-β3 alone). ^∗^p < 0.05 and ^∗∗^p < 0.01.(D) Percentage of Gdf5-RFP expressing cells on day 12 of micromass culture following IWP2 treatment in the presence or absence of TGF-β3 from 8 independent experiments. Values, mean ± SEM. ANOVA with Dunnett’s multiple comparison correction versus untreated. ^∗^p < 0.05 and ^∗∗∗∗^p < 0.0001.(E) Percentage of Gdf5-RFP expressing cells on day 18 of micromass culture following indicated treatment either from day 2–6 (early, light gray bars) or from day 2–18 (continuous, dark gray bars). Icons represent 3 independent experiments. ANOVA with Dunnett’s multiple comparison correction for significance versus respective early or continuous control (TGF-β3 alone). Student’s t test to compare early to continuous within each treatment. ^∗^p < 0.05.(F) RT-qPCR of Gdf5, Col2a1, and Prg4 on day 12 of micromass culture from 3 independent experiments. Values, mean ± SEM. ANOVA with Dunnett’s multiple comparison correction versus control. ^∗∗^p < 0.01 and ^∗∗∗^p < 0.001.(G) Sections of representative micromass tissues were stained with toluidine blue (purple stain indicates sulfated glycosaminoglycans) on day 32. Scale bars, 500 μm.(H) Gdf5-RFP-expressing and Prg4-GFP-expressing cells in sections of above micromasses were visualized by confocal fluorescence microscopy. Nuclei are stained with DAPI. Scale bars, 50 μm. Cells expressing both Gdf5-RFP and Prg4-GFP are indicated with gray arrows. See also Figure S2.
To identify the fate of these cells, we performed quantitative quantitative reverse-transcription PCR (RT-qPCR) on day 12 of micromass culture (Figure 2F). We observed significant increases in Gdf5 and Col2a1 in the presence of IWP2, and increased Prg4 expression compared to TGF-β3 alone. With CHIR, expression of these genes was either decreased or similar to cultures treated with TGF-β3 alone (Figure 2F). Sulfated glycosaminoglycans (sGAGs), which are abundant in cartilage, were found throughout the TGF-β3+IWP2-treated micromasses using the metachromatic toluidine blue stain (i.e., purple color in Figure 2G). TGF-β-treated tissues also contained areas with abundant sGAGs, while these were absent in CHIR-treated tissues. Gdf5-RFP+ cells were generally found to be evenly distributed within the cartilaginous regions of the TGF-β3-treated and TGF-β3+IWP2-treated tissues (Figure 1H). As expected, very few Prg4-GFP+ cells were observed, but were most abundant at the periphery of the micromass, where they co-expressed Gdf5-RFP (Figure 1H, white arrows in TGF-β3-treated tissue). Prg4-GFP+ cells isolated from micromasses treated with TGF-β3 showed restricted expression of Prg4 mRNA, but Gdf5 mRNA was not enriched in either GFP+ or GFP− cells (Figure S2D).
MAPK inhibition in the presence of TGF-β induces early Gdf5-RFP in the in vitro cultures
It has been previously reported that MAPK signaling plays a role in chondrogenic differentiation from bone marrow derived mesenchymal stem cells (Ma et al., 2019; Muddasani et al., 2007), and FGF signaling is active during chondrogenesis in appendicular skeletal elements. Here, we used our in vitro model to assess if MAPK signaling plays a role in Gdf5 induction and joint lineage specification. We introduced an MAPK agonist, basic fibroblast growth factor (FGF-2, bFGF), and an MAPK antagonist (MEK inhibitor PD0325901, referred to as “PD”) on day 2 and day 4 of micromass culture in the presence of TGF-β3. Inhibition of MEK with PD resulted in the upregulation of Gdf5-RFP by day 6 in a dose dependent manner (Figures 3A and 3B). Greater than 26% (and up to 55% in individual experiments) of cells were expressing Gdf5-RFP with PD concentrations of 3 μM or higher (Figure 3B), and was significantly higher than untreated cultures on days 6 and 12 (Figures 3E and 3F). PD-treatment alone, however, was not sufficient to induce Gdf5-RFP. Cells expressing both Gdf5-RFP and Prg4-GFP (double-positive) were also significantly higher with PD at 3 μM or higher concentrations, but did not exceed an average of 1.6% (Figure 3B). The introduction of bFGF resulted in significant increases in the percentage of Prg4-GFP+ cells by day 6 in a dose dependent manner, with greater than 16% and up to 48% of the cells expressing Prg4-GFP at concentrations between 20 and 400 ng/mL (Figures 3A and 3B). Prg4-GFP+ cells remained significantly higher in these bFGF-treated cultures compared to control on days 12 and 18 (Figure S3B), while levels of Gdf5-RFP plateaued to those observed with TGF-β3 alone (Figure S3A) and double-positive cells were not significantly different (Figure S3C).Figure 3. Modulation of MAPK signaling results in the expression of Gdf5-RFP and Prg4-GFP(A) Representative flow cytometry contour plots show expression of Gdf5-RFP and Prg4-GFP on day 6 of micromass culture following indicated treatment. All treatments are in the presence of TGF-β3 (10 ng/mL). PD, PD0325901; control, TGF-β3 treatment alone.(B) Percentage of Gdf5-RFP and/or Prg4-GFP expressing cells on day 6 of micromass culture in indicated conditions. Icons represent independent experiments. Values, mean ± SEM. ANOVA with Dunnett’s multiple comparison correction, ^∗^p < 0.05, ^∗∗^p < 0.01, ^∗∗∗^p < 0.001, and ^∗∗∗∗^p < 0.0001 versus control.(C) Confocal fluorescence and bright field microscopy image of TGF-β+PD (10 mM) treated micromass on day 6.(D) Confocal fluorescence and bright field microscopy image of TGF-β+bFGF (20 ng/mL) treated micromass on day 6.(E) Percentage of Gdf5-RFP expressing cells on day 6 of micromass culture following indicated treatment. Icons represent independent experiments. ANOVA with Dunnett’s multiple comparison correction, ^∗∗^p < 0.01 versus control.(F) Percentage of Gdf5-RFP expressing cells on day 12 of micromass culture following indicated treatment. Icons represent independent experiments. ANOVA with Dunnett’s multiple comparison correction, ^∗∗^p < 0.01 and ^∗∗∗∗^p < 0.0001 versus control.(G–J) RT-qPCR quantified expression of Gdf5 (G), Prg4 (H), Col2a1(I), and Scx (J) on day 6 of micromass culture. Icons represent independent experiments. Values, mean ± SEM. ANOVA with Dunnett’s multiple comparison correction, ^∗^p < 0.05 and ^∗∗^p < 0.01 versus control. See also Figure S3.
Gdf5-RFP+ cells were found in and between chondrogenic areas of PD-treated micromasses on day 6, while Prg4-GFP+ cells were highly prevalent in the fibroblast-like cells outgrowing from bFGF-treated micromasses, similar to their peripheral location in Figure 1H (Figures 3C and 3D). RT-qPCR for Gdf5 on day 6 corroborated the increase in Gdf5-RFP+ cells we observed by flow cytometry in the PD-treated cells and confirmed no effect with bFGF (Figure 3G). bFGF caused a significant increased in Prg4 mRNA compared to control cultures (Figure 3I), but expression was also increased in PD-treated cultures, possibly reflecting the higher percentage of double-positive cells (Figure 3B). However, neither PD nor bFGF resulted in significant increases in Col2a1 compared to TGF-β3 alone (Figure 3H), rather the highest concentrations of both reduced Col2a1 expression. From these data, the fate of the PD and bFGF-treated cells remained unclear. As both TGF-β and FGF signaling have been implicated in tendon/ligament lineages (Havis et al., 2016), we quantified the expression of scleraxis (Scx), a transcription factor expressed in early tendon and ligament fibroblasts (Brent and Tabin, 2004). We observed a significant increase in Scx expression at the highest dose of bFGF (100 ng/mL) (Figure 3J), suggesting that the Prg4-GFP+ cells may be of tendon/ligament lineage.
Single cell RNA-seq identified similar cell populations in in vitro generated micromasses and mouse embryonic limb buds
To identify the cell types and developmental stages of Gdf5-RFP+ and Prg4-GFP+ cells, we performed single cell transcriptomic analysis of micromasses when reporter genes were expressed: TGF-β3, TGF-β3+PD (high Gdf5-RFP), and TGF-β3+bFGF (high Prg4-GFP) on day 7, and micromasses treated with TGF-β3 and TGF-β3+IWP2 (high Gdf5-RFP) on day 14. As in vivo controls, we sequenced cells isolated from mouse embryonic limb buds at the onset of Gdf5 expression in the developing joints (E12.5 forelimb) and two days later when specialized joint cell types were arising (E14.5 hindlimb and the autopod of E14.5 hindlimb) (Table S1). Each dataset was first evaluated individually, and cell clusters were identified using differentially expressed genes (DEGs) and known marker genes (Figure S4). We identified diverse cell types (e.g., connective tissue, chondrogenic, myogenic, neural, limb bud mesenchyme, endothelial, among others) in the limb buds, as expected. In vitro, the most prominent cell types were chondrogenic, connective tissue-like, and cells that co-express mesoderm and chondrogenic genes (Alexandrovich et al., 2008; Almubarak et al., 2021) (Figures S4A and S4D). Small proportions of cells with neuronal or mitochondrial signatures were termed “off-target” (Figure S4; Table S1).
To comparatively analyze mESC-derived cells and mouse embryonic limb bud cells, we integrated the chondrogenic, connective tissue, and mesoderm/mesenchyme clusters from each sample (Figures 4A–4C). E11, E13, and E15 hindlimb bud cells, sequenced by Kelly et al. (2020) were also integrated to represent additional and intermediate time points of in vivo limb development. Six clusters were identified (Figures S5A and S5B; Table S2), reflecting chondrogenic cells (expressing Col2a1, Acan, Col9a1, Col11a1, and Hapln1), chondrogenic progenitor cells (expressing Sox5, Sox6, and Sox9) (Bhattaram et al., 2010), two distinct connective tissue populations (expressing Col1a1, Col3a1, Dcn, Lum or Egfl6, Postn, Scx, and Tnmd) (Markman et al., 2023; Ying et al., 1997; Raouf et al., 2002; Liu et al., 2015; Sakai and Kumagai, 2025), and limb bud mesenchyme/proliferative lineages (expressing Prrx1, Lhx9, Mecom, Ebf1, Foxp1, Msx1, and Top2a) (Leussink et al., 1995; Martin et al., 1995; Satoh et al., 2007; Tzchori et al., 2009) (Figures 4A–4D, and 4E). Chondrogenic cells were present in all in vitro treatments, suggesting that a chondrogenic fate was being induced as early as 1 week of culture (Figure 4C). TGF-β3+IWP2 cultures proportionally had more chondrogenic cells compared to TGF-β-3 alone, consistent with the flow cytometry and gene expression data (Figure 4B). TGF-β3+PD (high Gdf5-RFP+) and TGF-β3+bFGF (high Prg4-GFP+) micromass cells after 1 week also showed contrasting cell composition, with PD treatment having increased chondrogenic cells, and bFGF treatment having proportionally more cells within the connective tissue 2, limb bud mesenchyme and limb bud mesenchyme proliferative clusters (Figure 4B).Figure 4. Overview of single cell RNA sequencing analysis(A) Integrated UMAP of in vitro and in vivo datasets.(B) Bar graph illustrates the proportion of each cluster per sample.(C) Individual (split) UMAPs illustrate the general proportion of cells with each cell cluster. E11, E13, and E15 hindlimb samples were previously published (Kelly et al., 2020).(D) Feature plots illustrate the level of indicated genes’ expression in each cell in integrated RNA object.(E) Dot plot shows the percent of cell expressing and the average normalized expression levels of indicated genes in each cluster of the integrated RNA object. See also Figure S4.
Gdf5-expressing cells were largely clustered with chondrogenic progenitors and chondrocytes (Figure 4D). mESC-derived cells expressing RFP mRNA overlapped with cells expressing Gdf5 mRNA, consistent with cell sorting (Figure 1F) and histological results (Figure 2G). Gdf5 expression did not overlap significantly with the canonical Wnt ligand Wnt9a, which induced ectopic Gdf5 expression in the developing limb (Guo et al., 2004) (Figure 4D). Prg4-expressing cells and GFP-expressing cells were found in all clusters, consistent with a variety of joint tissue lineages known to express Prg4. Notably, both Prg4+ and GFP+ cells are found in the connective tissue 2 cluster, which co-expressed Tnmd and Scx (Figures 4D and 4E). The specificity of Prg4 expression to mESC-derived cells expressing GFP was confirmed following cell sorting and qPCR (Figure S2D).
We split the UMAPs by sample/treatment to specifically interrogate if changes in gene expression arose in one or more clusters following differential treatment (Figure 5A). While all in vitro cultures had cells with a limb bud mesenchyme-like signature, the TGF-β3+bFGF micromass had the highest number of cells expressing the limb bud, somite, and craniofacial mesenchyme gene Msx1. All cultures also contained cells within the two connective tissue clusters, however only TGF-β3+bFGF-treated cultures contained a large proportion of cells expressing the tendon/ligament-specific gene Tnmd (Figure 5A). The majority of Tnmd expressing cells were located within the connective tissue 2 cluster, which we subset from TGF-β3+PD (high Gdf5-RFP+) and TGF-β3+bFGF (high Prg4-GFP+) treatments to investigate changes in gene expression (Figure 5B). MAPK target genes such as Dusp6, Spry1, and Etv1 (Janknecht, 1996; Hanafusa et al., 2002; Ahmad et al., 2018) were among the highest DEGs in the TGF-β3+bFGF treatment, along with GFP, consistent with flow cytometry data. Connective tissue 2 cells in the TGF-β3+PD treatment had significantly higher levels of Gdf5 and RFP, as expected, and genes associated with ECM organization like Lox (Ahsan et al., 2005; Makris et al., 2013) and Sparc (Mundlos et al., 1992) and the glycoprotein Cd248 (Hong et al., 2024). The chondrogenic cluster in TGF-β3 or TGF-β3+IWP2 micromasses were also specifically subset to quantify gene expression changes in the chondrocyte lineage (Figure 5C). Col2a1, Col9a3, Acan, and Cnmd were among the most significant genes elevated in the TGF-β3+IWP2 treatment (Figure 5C). While it remained clear that the cells within this cluster in the TGF-β3 micromass were also chondrocytes, genes more highly expressed encoded for ribosomal proteins.Figure 5. Transcriptomic changes that occur in mESC-derived micromass cultures as a result of treatment conditions(A) Feature plots illustrate the level of indicated genes’ expression in each cell in UMAPs split by sample.(B) Volcano plot illustrates differentially expressed genes.in the connective tissue 2 clusters from TGF-β+PD and TGF-β+bFGF samples. Gdf5 and RFP were enriched in cells treated with TGF-β+PD, while MAPK signaling target genes (e.g., Dusp6, Etv1, and Spry1) and genes associated with the tendon/ligament lineage (e.g., Tnmd and Prg4) were enriched in cells treated with TGF-β+bFGF. Cutoffs at log(2)FC >1 and padj <0.005.(C) Volcano plot illustrates differentially expressed genes in the chondrogenic clusters from TGF-β and TGF-β+IWP2 samples after two weeks of treatment. Chondrogenic genes such as Col2a1, Comp, Cnmd, and Acan are enriched in cells treated with TGF-β+IWP2. Cutoffs at log(2)FC > 1 and padj <0.005. Genes encoding ribosomal proteins were not omitted from the datasets for DEG analyses. See also Figure S5.
Overall, these data identified similar cell populations within in vitro generated mESC-derived micromass cultures and mouse embryonic limb buds, highlighting the in vivo relevance of this in vitro experimental model and suggest we can use the mESC model to study these phenomena.
Effect of TGF-β, Wnt, and MAPK modulation in ex vivo embryonic limb bud cultures
In vitro cell culture can introduce artifacts that are not present in vivo, and likewise, there are signals present in vivo that are not precisely recapitulated in vitro. Given the transcriptional similarities of cells within mESC-derived micromasses and embryonic limb buds, we sought to determine whether signaling pathways we identified in vitro could induce Gdf5 expression in primary cells ex vivo using a well-established limb bud micromass assay (Kulyk et al., 1989; Seo and Serra, 2007) (Figure 6A). We first assessed the baseline expression patterns of Gdf5 in this ex vivo micromass culture using two different transgenic mice with LacZ reporter genes under the control of (1) a 200 kb BAC containing all known upstream and downstream regulatory elements of Gdf5 (Chen et al., 2016; Capellini et al., 2017) or (2) an enhancer element, R4, that drives expression specifically in cells within the joint regions in developing mice, also referred to as PHC21 strain (Chen et al., 2016; Richard et al., 2020). Embryonic limb bud cells were isolated at E11.5, prior to the onset of Gdf5 expression, and cultured in micromass (Figure 6A). Remarkably, LacZ expression patterns in the micromass cultures of each strain were reminiscent of their in vivo expression, where expression in the 200 kb BAC transgenic mouse at E14.5 was broader than that of the Gdf5-R4-LacZ transgenic mouse (Figures S6A and S6B). Micromasses from the 200 kb BAC transgenic mouse had LacZ+ cells primarily within chondrogenic nodes and less frequently in cells adjacent to and surrounding chondrogenic nodes (Figure S6B). The majority of Gdf5-R4-LacZ-derived positively stained cells, in contrast, were adjacent to and in between chondrogenic nodes (Figure S6B), similar to the specificity of this enhancer-reporter in developing joints in vivo. Thus, to understand the initiation of Gdf5 expression in developing joint cells, we tested the effects of modulating TGF-β, Wnt, and MAPK signaling using the Gdf5-R4-LacZ mouse strain (Figure 6).Figure 6. Gdf5-R4-LacZ reporter expression and chondrogenesis in embryonic limb bud micromass cultures following TGF-β, Wnt, and MAPK modulation(A) Limb buds from Gdf5-R4-LacZ transgenic mice were microdissected on embryonic day 11.5 (E11.5) and isolated cells were cultured in micromass for 4 days in the presence or absence of factors: TGF-β3 10 ng/mL, CHIR 1 μM, IWP2 2 μM, bFGF 20 ng/mL, and PD 3 μM. Chondrogenic nodes (subset indicated with dashed red line and/or “Ch”) in limb bud micromass cultures stain positively with Alcian blue indicating presence of proteoglycans/glycosaminoglycans in cartilage matrix. Scale bars, 200 μm.(B) Gdf5-R4-LacZ-expressing cells were visualized by X-gal staining of micromass cultures after 4 days. Dashed red lines in the control image indicate representative chondrogenic nodes. Scale bars, 75 μm. Representative contour plots indicate the percentage of cells expressing the LacZ reporter quantified by flow cytometry using the FDG reagent.(C) Fold change in the percentage of Gdf5-R4-LacZ-expressing cells within each litter (n) is depicted. Each treatment value was normalized to untreated control culture within the same litter, n = 5 litters (independent experiments). Values, mean ± SEM. ANOVA with Dunnett’s multiple comparison correction. ^∗^p < 0.05, ^∗∗^p < 0.01, and ^∗∗∗^p < 0.001 vs. control.(D–F) RT-qPCR of Gdf5, Col2a1, and Prg4 following 4 days of micromass culture, n = 4 litters (independent experiments), Values, mean ± SEM. ANOVA with Dunnett’s multiple comparison correction. ^∗^p < 0.05 and ^∗∗^p < 0.01. See also Figure S6.
The initial requirement of TGF-β signaling Gdf5-RFP induction in mESC-derived cells suggested that TGF-β would increase the percentage of Gdf5-R4-LacZ expressing cells in the limb bud micromasses. However, a small but significant reduction in the number of LacZ+ cells was observed after 4 days compared to untreated controls (Figures 6B and 6C). This unexpected result could reflect the endogenous (i.e., sufficient) levels of TGF-β signaling in the chondrogenic mesenchyme in E11.5 limb buds or the differences in regulatory control of the R4 enhancer versus the endogenous Gdf5 locus. The expression of Gdf5 mRNA in TGF-β-treated micromasses was not significantly different from control (Figure 6D), supporting the latter. To address the former, we quantified the expression of several genes known to be regulated by TGF-β signaling in isolated limb bud cells compared to mESC-derived chondrogenic mesoderm cells prior to and after TGF-β treatment in micromass (Figures S6C and S6D). The expression of Snai1 (Basu et al., 2015), Ccn2 (Igarashi et al., 1993; Untergasser et al., 2005), Pai1 (Geng et al., 1999; Hirashima et al., 2003), and Smad7 (Afrakhte et al., 1998) were all significantly higher in E11.5 limb bud cells compared to chondrogenic mesoderm cells on day 5 of differentiation (Figure S6C). Following 6 days in micromass with TGF-β, mESC-derived cells upregulated all 4 genes, and only Pai1 was significantly different from the E11.5 limb bud cells, being higher in the mESC-derived cells (Figure S6D). These results suggest that endogenous TGF-β signaling was sufficient to permit expression of Gdf5-R4-LacZ ex vivo; therefore, we tested the effects of modulating Wnt and MAPK signaling in the absence of exogenous TGF-β.
Limb bud cells isolated from 5 independent litters were treated in micromass with CHIR (1 μM), IWP2 (2 μM), bFGF (20 ng/mL), or PD (3 μM), and the expression of Gdf5-R4-LacZ was quantified by flow cytometry and microscopy on day 4 (Figures 6B and 6C). Similar to the results observed in mESC-derived micromasses, activation of Wnt with CHIR or MAPK with bFGF significantly reduced the percentage of Gdf5-R4-LacZ+ cells while inhibition of Wnt with IWP2 or inhibition of MAPK signaling with PD significantly increased the percentage of Gdf5-R4-LacZ+ cells (Figure 6C). Gdf5 expression was increased in replicate cultures from each litter with IWP2 and PD treatment (Figure 6D), with PD being significantly higher than control. CHIR significantly reduced Gdf5 expression, and no significant changes were observed with TGF-β or bFGF treatment. Similar to the chondrogenic-promoting effects of Wnt inhibition in mESCs, IWP2 treatment led to a significant increase in Col2a1 in the limb bud cells, which was reduced in the CHIR treatment (Figure 6E). We confirmed, in non-transgenic limb bud cells, that recombinant proteins that activate (Wnt3a) or inhibit Wnt (DKK1) behaved similarly to the small molecules used in this study (Figure S6E). bFGF and PD treatments both resulted in decreases in Col2a1 compared to untreated controls, but not significantly (Figure 6F). Finally, as expected, TGF-β3 and bFGF treatments both increased Prg4 expression (Figure 6G), with the TGF-β3-mediated increase being significant. These data support the notion that signals we identified to induce Gdf5 and Prg4 expression in mESCs in vitro were able to induce similar expression and cell fate changes in mouse embryonic limb bud cells ex vivo.
Discussion
In this study, we generated a dual fluorescent reporter mESC line to study the role of key signaling pathways in joint development. We utilized directed differentiation methodology and demonstrated that TGF-β signaling is necessary for induction of Gdf5 expression in vitro, and modulation of Wnt and MAPK signaling pathways significantly impacted the onset of both Gdf5 and Prg4 expression. Single cell transcriptomic analysis revealed that joint lineage cells induced by TGF-β, Wnt, and MAPK modulation had similar transcriptomic profiles to chondrogenic and connective tissue lineages in mouse embryonic limb buds. Finally, we corroborated our in vitro findings regarding modulation of these pathways in ex vivo limb bud cultures, highlighting the utility of this dual reporter mESC line in studying specification of joint progenitor cells and joint developmental processes.
Our finding that canonical Wnt signaling prohibits chondrogenesis in both mESC and limb bud micromasses is consistent with previous reports (Koyama et al., 1996; Serra et al., 1997; Wu et al., 2021). Building on this, we found that inhibition of Wnt promoted Gdf5 expression and chondrogenesis, which upon face value is inconsistent with previous reports of Wnt9a/14 ligand expression in the interzone, and that ectopic Wnt9a/14 expression can induce Gdf5 expression (Guo et al., 2004; Hartmann and Tabin, 2001; Bhattaram et al., 2014). We suggest these data support a dynamic temporal role for Wnt signaling in joint development. In vivo, Wnt activation may be required to suppress chondrogenesis, as suggested by high endogenous expression of TGF-β target genes and the fact that exogenous TGF-β was not necessary for Gdf5 expression ex vivo as it was in vitro. At later times, Wnt signaling must be dampened to support chondrogenesis of those same progenitors. In vitro, however, the Wnt-activating function of chondrogenic repression was not required, as suggested by significantly lower basal levels of TGF-β target genes and the requirement of exogenous TGF-β to induce Gdf5 expression and chondrogenesis.
MAPK inhibition, via MEK inhibition, strongly promoted the expression of Gdf5 in both mESC-derived and ex vivo limb bud cultures, a finding that provides additional insights around joint cavity formation. The joint line that will ultimately result in the creation of the joint cavity is known to be driven through the activation of the MEK-ERK pathway (Bastow et al., 2005), and this niche inhibits Gdf5 expression to prevent joint fusion (Lamb et al., 2003). The role of joint line cells in promoting cavitation is thus distinguishable from the Gdf5-expressing cells that will emerge and undergo differentiation. The former exhibit activated MEK/ERK, while the latter present with decreased MEK/ERK signaling. This phenomenon is consistent with the observed acceleration of Gdf5 expression and chondrogenesis upon MEK inhibition in vitro and ex vivo in this study.
Lineage tracing in mice have reproducibly shown that Gdf5-lineage cells contribute to the formation of permanent articular cartilage, and not to the chondrogenic mesenchyme/growth plate cartilage involved in endochondral ossification (Koyama et al., 2008; Shwartz et al., 2016). It is thus justly hypothesized that Gdf5-lineage cells would be an optimal progenitor cell source to not only study intra-articular tissue development, but to generate permanent articular cartilage cells and tissues for replacement or repair. hESC-derived populations enriched for GDF5 expression preferentially generated chondrocytes lacking a growth plate-like phenotype compared to those enriched for SOX9 expression (Pothiawala et al., 2022). We performed similar experiments in which we demonstrated that articular chondrocytes derived from hESCs, which expressed GDF5 at earlier time points, remained cartilaginous in vivo, and did not initiate ossification following subcutaneous injection in mice (Craft et al., 2015). Following implantation of these same articular cartilage tissues in focal cartilage defects in the rat, however, regions exposed to bone marrow spaces did undergo remodeling and hypertrophy as one might expect from a more naive cartilage anlage (Gardner et al., 2019). In earlier studies using chondrocytes derived from mESCs, we found ossification was only prevented when grafts were physically protected from invading host vasculature (Craft et al., 2013). In contrast, GDF5 also participates in promoting vascularization in a rabbit long-bone defect model (Kleinschmidt et al., 2014), and both recombinant GDF5 (Coleman and Tuan, 2003) and forced expression of GDF5 can promote chondrocyte hypertrophy (Tsumaki et al., 1999). Thus, we do not expect that chondrocytes enriched in Gdf5/GDF5 expression will remain as permanent cartilage in a scenario where vascularization is permitted or if Gdf5/GDF5 expression persists. The optimal progenitor cell is important, but the recipient environment must be optimal as well.
Through our manipulation of the MAPK pathway, we observed that treatment with basic bFGF increased expression of Prg4-GFP and Prg4 mRNA in a dose dependent manner, concomitant with evidence of a tendon/ligament lineage. Expression of Prg4 mRNA was also increased in PD-treated cultures to the same extent, while these cultures lacked a significant proportion of Prg4-GFP+ cells. Prg4 expression in PD-treated cultures could reflect the higher percentage of double-positive cells, but it is possible that reporter expression is not perfectly aligned with endogenous gene expression from the untargeted allele, raising concerns about the conclusions we have made. In a recent study using a complementary Col2a1-IRES-nGFP;Prg4-tdTomato mESC reporter line, we found Prg4, in a scenario where Col2a1 expression is absent, to be a useful surrogate marker for tendon-like cells (Niu et al., 2024). In support of our findings here, activation of MAPK in mESC-derived progenitors with either cyclic (c)AMP agonists or bFGF in this study also resulted in the generation of Prg4-expressing cells that expressed tendon/ligament gene repertoires (e.g., Scx and Tnmd). Thus, while we showed that MAPK plays a role in promoting the tendon-ligament lineage, further investigation is needed to delineate the discrepancy between the GFP reporter and Prg4 mRNA expression in this mESC line.
Synovial joints are present in both the appendicular and axial skeleton and prior work has shown that regulatory elements within the upstream BAC and downstream 200 kB BAC drive Gdf5-lacZ expression in both anatomical locations in mice (Chen et al., 2016). In vitro, directed differentiation methodologies to induce paraxial mesoderm, which gives rise to cartilage in the vertebrae and ribs, versus lateral plate mesoderm, responsible for cartilage formation in the limbs, have become increasingly refined over time (Chal et al., 2018; Pothiawala et al., 2022; Kattman et al., 2011; Loh et al., 2016). Paraxial mesoderm, often induced with Wnt activation and BMP inhibition, can be identified in in vitro cultures by the expression of Pdgfra/PDGFRA and lack of Flk-1/KDR, while lateral plate mesoderm induction protocols include active BMP signaling, resulting in the expression of both of these markers. However, the important distinction between articular cartilage, which is derived from a joint cell progenitor, and growth plate cartilage, which is not (Koyama et al., 2008) has been more extensively described using paraxial mesoderm induction protocols (Craft et al., 2015; Pothiawala et al., 2022). The protocol used herein also induced a Pdgfra^+^Flk-1^−^ mesoderm population with BMP inhibition and endogenous Wnt signaling, suggesting a paraxial mesoderm intermediate, although this was not shown definitively. Thus, we cannot claim that cells expressing Gdf5-RFP or Prg4-GFP are of the same anatomical origin as the ex vivo limb bud cells. We did however corroborate findings from the in vitro micromass culture to ex vivo mouse embryonic limb bud cultures under a number of different treatment conditions. These results suggest that there may be shared mechanisms of joint cell specification in developing axial and appendicular synovial joints, and this ESC model may recapitulate fundamental concepts regarding the molecular regulation of this process.
The epigenetic regulation of Gdf5 expression is anatomically complex, and one caveat to this study is that the expression of the fluorescent reporters in the mESC line was not confirmed to track with gene expression in the mouse embryo. The Gdf5 R4 enhancer element resides within a much larger regulatory locus, encapsulated by at least 200 kb of genomic sequence containing many additional tissue-specific regulatory elements, including GROW1, which activates Gdf5 in chondrogenic long bone/growth plate regions (Capellini et al., 2017; Muthuirulan et al., 2021; Chen et al., 2016; Coveney et al., 2025). It was remarkable to find that the pattern of LacZ expression driven by the larger 200 KB (BAC) reflected a broader domain, as expression resided both adjacent to (as driven by the R4 sequence) and within chondrogenic nodes. In vivo, the R4 regulatory element drives strong Gdf5 and LacZ expression in joint interzones and then in the developing joint tissues through early post-natal life, which correlates with its expression in cells adjacent to and along the periphery of the chondrogenic nodes of limb bud micromasses. Interestingly, loss of this enhancer in mice and human cells alters joint morphology and contributes to osteoarthritis risk (Richard et al., 2020). These regulatory elements and their specific expression domains were also identified during human chondrogenesis and joint formation (Richard et al., 2020; Muthuirulan et al., 2021), indicating that such signal-regulatory element interactions are conserved across mammals, and likely across amniotes.
Since the discovery of Gdf5 as the earliest marker of joint initiation (Storm and Kingsley, 1999; Koyama et al., 2008), we have made limited progress in deciphering the cues underlying the specification of these progenitor cells that give rise to joint tissues required for mobility. As injuries to joint tissues are prevalent in all demographics, it is important that we investigate these developmental processes to support the evolution of novel therapeutics. This study introduced new experimental platforms that successfully elucidated mechanisms for promoting joint cell identities, thereby illuminating a poorly understood area of cell fate determination and furthers our ability to generate joint lineage cells from pluripotent stem cells.
Methods
Generation of dual mESC reporter line
The coding sequences for tdRFP and eGFP were introduced into the Gdf5 and Prg4 loci, respectively, of E14 mESCs using CRISPR-Cas9 (Cong et al., 2013) to generate the BC29 dual reporter line (Figures S1A and S1B). Clones were confirmed by Sanger sequencing. We note that the untargeted allele of Prg4 is missing one cysteine codon in the signal peptide.
ESC maintenance and differentiation
mESCs were maintained in a modified serum-free (SF), feeder-free culture system (i.e., 2i media) as described previously (Sim et al., 2017; Craft et al., 2013; Gadue et al., 2006; Ying et al., 2003). For differentiation, ESCs were cultured in suspension in SF differentiation medium (SFD) (Gouon-Evans et al., 2006) without additional factors for 48 h. Embryoid bodies were treated with activin A, CHIR99021, and noggin or LDN-193189 for 28 h to induce primitive streak mesoderm, then resuspended in SFD containing bFGF and Y-27632 for 48 h. High density micromasses were generated from mesoderm cells on day 5 (Craft et al., 2015) and maintained in chondrogenic media consisting of high glucose DMEM with 1× Insulin Transferrin Selenium (ITS) supplement, ascorbic acid (50 μg/mL), proline (40 μg/mL), and dexamethasone (0.1 μM) with indicated factors. Recombinant growth factors and small molecules were purchased from R&D Systems and Sigma-Aldrich. Details including concentrations of all factors are in the supplemental information.
Mouse embryonic limb bud cultures
Animal studies were performed in compliance of ethical regulations and were approved by Animal Resources at Children’s Hospital or Harvard University. Wild-type FVB (friend virus B) strain mice, transgenic 200 kb BAC Gdf5 LacZ mice, and PHC21 (Gdf5-R4-LacZ) mice (both FVB strain) were used for these studies. From each pregnant dam, forelimb and hindlimb buds were micro-dissected from all mouse embryos, pooled prior to dissociation with 0.2% type I collagenase for two hours at 37°C and cells were plated in micromass.
Flow cytometry and cell sorting
The following antibodies were used for flow cytometry: anti-mouse Flk-1-biotin, anti-mouse Pdgfrα (CD140a)-allophycocyanin (APC; clone APA5), streptavidin-phycoerythrin (PE) or streptavidin-PE-Cy7 (BD Pharmingen). Micromasses were dissociated with collagenase prior to analysis. Limb bud cells expressing beta-galactosidase (Gdf5-R4-LacZ transgene) were detected using the fluorescein di-V-galactoside (FDG) reagent within the FluoReporter LacZ flow cytometry kit (F1930, Thermo Fisher Scientific) following manufacturer’s instructions. Cells were analyzed or sorted using a BD FACS Fortessa and BD Melody (Becton Dickinson), analysis was performed using FlowJo (Tree Star), and mean, standard errors and all statistical tests were calculated in Prism (GraphPad).
Quantitative reverse-transcription PCR (RT-qPCR)
Total RNA was extracted using the MagMAX mirVana Total RNA kit (Applied Biosystems), and reverse transcribed with Superscript IV VILO reverse transcriptase with ezDNase enzyme (Invitrogen). RT-qPCR was performed on a ViiA 7 Real-Time PCR System using PowerUp SYBR Green PCR kit (Applied Biosystems). The copy number of each gene shown is normalized to that of β-actin. Biological replicates/independent experiments are indicated in figures or legends. Primer sequences are in the supplemental material. Mean, standard errors, and all statistical tests were calculated in Prism (GraphPad).
Histology, staining, and imaging
Micromasses from limb bud cells were fixed in 4% paraformaldehyde (PFA) prior to standard X-gal and Alcian blue staining. 5–10 μm sections of micromasses derived from mESCs were cryosectioned, fixed in 4% PFA, and mounted with DAPI-containing media or stained with toluidine blue. Imaging was performed on an EVOS FL Auto 2 or a Zeiss LCM800 confocal microscope.
scRNA-seq methods and analysis
scRNA-seq libraries were prepared using Single Cell 3′ Library & Gel Bead Kit v2 (10× Genomics) and sequenced on an Illumina NextSeq 500 to ∼30,000 reads per cell. Reads were processed by Cell Ranger (version 3.0.0), and aligned to the GRCm38 (version 100) genome amended with RFP and eGFP transgenes. Cells with <500 UMIs, >70,000 UMIs, or >20% mitochondrial DNA were removed. The cell matrix was normalized using SCTransform (Hafemeister and Satija, 2019), and uniform manifold approximation and projection (UMAP) was used for dimensional reduction and visualization (McInnes et al., 2018). Clustering and DEGs were defined using Leiden algorithm (Traag et al., 2019) and the model-based analysis of single-cell transcriptomics (MAST) framework (Finak et al., 2015). Indicated cell types were integrated using reciprocal PCA (Butler et al., 2018), using the E14.5 limb samples as reference. Details including all packages and versions used can be found in the supplemental information.
Resource availability
Lead contact
Requests for further information and resources should be directed to and will be fulfilled by the lead contact, April Craft ([email protected]).
Materials availability
Plasmids and the mESC line generated in this study are available for research purposes with a completed materials transfer agreement.
Data and code availability
Raw single-cell transcriptomic data used in this publication are deposited at GEO: [GSE274060](GSE274060). All code and analyzed R objects are available on Github at github.com/suyraj/Mouse-Cartilage-scRNAseq/tree/main. Requests for additional information should be directed to the corresponding author.
Acknowledgments
The authors would like to acknowledge Dr. Patrick Tschopp (University of Basel) for thoughtful discussions regarding the research project, and Dr. Clarissa Coveney and members of the Craft lab for critical reading of the manuscript. This project was supported with 10.13039/100000052NIH/10.13039/100000069NIAMS R01-AR073821 [A.M.C.] and R01-AR070139 [T.D.C.] and 10.13039/100000964Arthritis National Research Foundation (A.M.C.) funding. We thank the BPF Genomics Core Facility at 10.13039/100006691Harvard Medical School (RRID: SCR_007175) and the Center for Musculoskeletal Research (P30 AR075042) for expertise that supported this work.
Author contributions
Conceptualization, A.M.C.; formal analysis, S.R., T.C., A.M.C., and M.A.-R.; funding acquisition, A.M.C. and T.D.C.; investigation, S.R., T.C., S.M., M.A.-R., J.C., and S.K.J., methodology, S.R., T.C., and A.M.C.; resources, S.R., M.Y., and T.D.C.; visualization, S.R., T.C., and A.M.C.; writing – original draft, S.R., S.K.J., and A.M.C.; writing – review & editing, S.R., S.K.J., T.D.C., and A.M.C.
Declaration of interests
The authors declare no competing interests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Afrakhte M.Morén A.Jossan S.Itoh S.Sampath K.Westermark B.Heldin C.H.Heldin N.E.ten Dijke P.Induction of inhibitory Smad 6 and Smad 7 m RNA by TGF-beta family members Biochem. Biophys. Res. Commun.2491998505511971272610.1006/bbrc.1998.9170 · doi ↗ · pubmed ↗
- 2Ahmad M.K.Abdollah N.A.Shafie N.H.Yusof N.M.Razak S.R.A.Dual-specificity phosphatase 6 (DUSP 6): a review of its molecular characteristics and clinical relevance in cancer Cancer Biol. Med.15201814282954596510.20892/j.issn.2095-3941.2017.0107 PMC 5842331 · doi ↗ · pubmed ↗
- 3Ahsan T.Harwood F.Mc Gowan K.B.Amiel D.Sah R.L.Kinetics of collagen crosslinking in adult bovine articular cartilage Osteoarthr. Cartil.132005709715
- 4Alexandrovich A.Qureishi A.Coudert A.E.Zhang L.Grigoriadis A.E.Shah A.M.Brewer A.C.Pizzey J.A.A role for GATA-6 in vertebrate chondrogenesis Dev. Biol.31420084574701819112010.1016/j.ydbio.2007.12.001 · doi ↗ · pubmed ↗
- 5Almubarak A.Lavy R.Srnic N.Hu Y.Maripuri D.P.Kume T.Berry F.B.Loss of Foxc 1 and Foxc 2 function in chondroprogenitor cells disrupts endochondral ossification J. Biol. Chem.2972021101020
- 6Bastow E.R.Lamb K.J.Lewthwaite J.C.Osborne A.C.Kavanagh E.Wheeler-Jones C.P.D.Pitsillides A.A.Selective Activation of the MEK-ERK Pathway Is Regulated by Mechanical Stimuli in Forming Joints and Promotes Pericellular Matrix Formation J. Biol. Chem.280200511749117581564728610.1074/jbc.M 414495200 · doi ↗ · pubmed ↗
- 7Basu M.Bhattacharya R.Ray U.Mukhopadhyay S.Chatterjee U.Roy S.S.Invasion of ovarian cancer cells is induced by PITX 2-mediated activation of TGF-β and Activin-A Mol. Cancer 1420151622629839010.1186/s 12943-015-0433-y PMC 4546816 · doi ↗ · pubmed ↗
- 8Bhattaram P.Penzo-Méndez A.Kato K.Bandyopadhyay K.Gadi A.Taketo M.M.Lefebvre V.SOXC proteins amplify canonical WNT signaling to secure nonchondrocytic fates in skeletogenesis J. Cell Biol.20720146576712545238610.1083/jcb.201405098 PMC 4259807 · doi ↗ · pubmed ↗