The LINC01612-DVL2-WNT axis promotes human endoderm differentiation
Mao Li, Pei Lu, Jie Yang, Chenchao Yan, Yikang Yang, Wei Jiang

TL;DR
The study identifies a long noncoding RNA, LINC01612, that supports human endoderm differentiation by stabilizing a key WNT signaling protein called DVL2.
Contribution
LINC01612 is newly characterized as a desert lncRNA that promotes endoderm differentiation via the DVL2-WNT signaling axis.
Findings
LINC01612 depletion impairs human definitive endoderm differentiation.
LINC01612 stabilizes DVL2 protein by reducing its ubiquitination.
WNT signaling is impaired in the absence of LINC01612 or DVL2.
Abstract
Long noncoding RNAs in gene desert regions remain largely uncharacterized despite their potential regulatory roles in cell differentiation. Here, we identify LINC01612 as a crucial modulator of human definitive endoderm differentiation. LINC01612 exhibits stage-specific expression and lacks protein-coding potential during endoderm differentiation. Depletion of LINC01612, through either short hairpin RNA (shRNA)-mediated knockdown or promoter deletion, severely impairs human endoderm differentiation. Mechanistically, LINC01612 interacts with DVL2, a WNT regulator essential for early development, and enhances DVL2 protein stability by reducing its ubiquitination. Loss of LINC01612 or DVL2 impairs WNT signaling, while both WNT activation and DVL2 overexpression can rescue endoderm differentiation defect in the absence of LINC01612. These findings reveal the LINC01612-DVL2-WNT regulatory…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
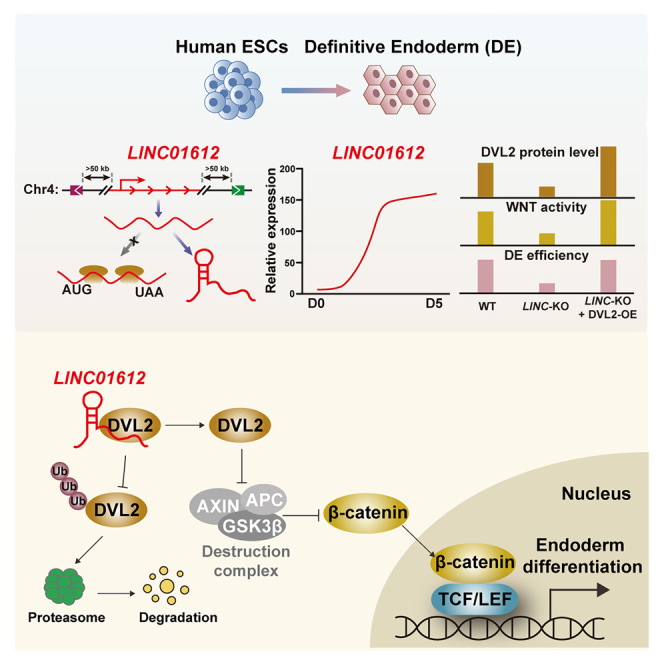 Figure 1
Figure 1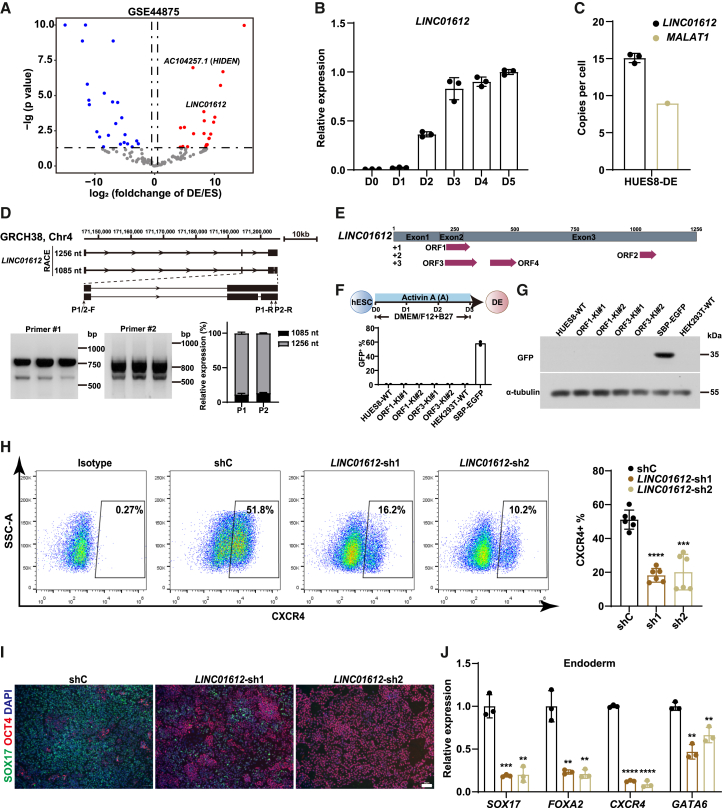 Figure 2
Figure 2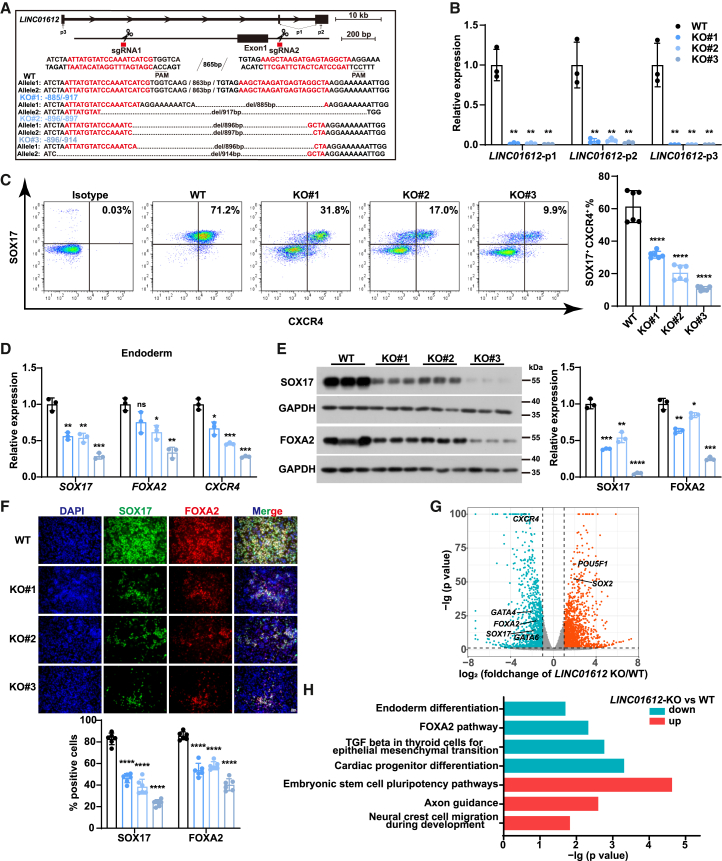 Figure 3
Figure 3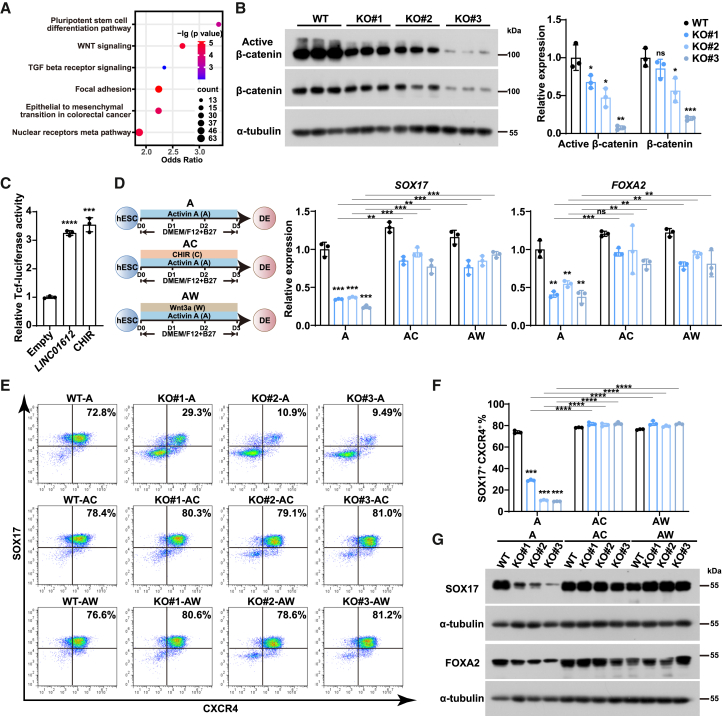 Figure 4
Figure 4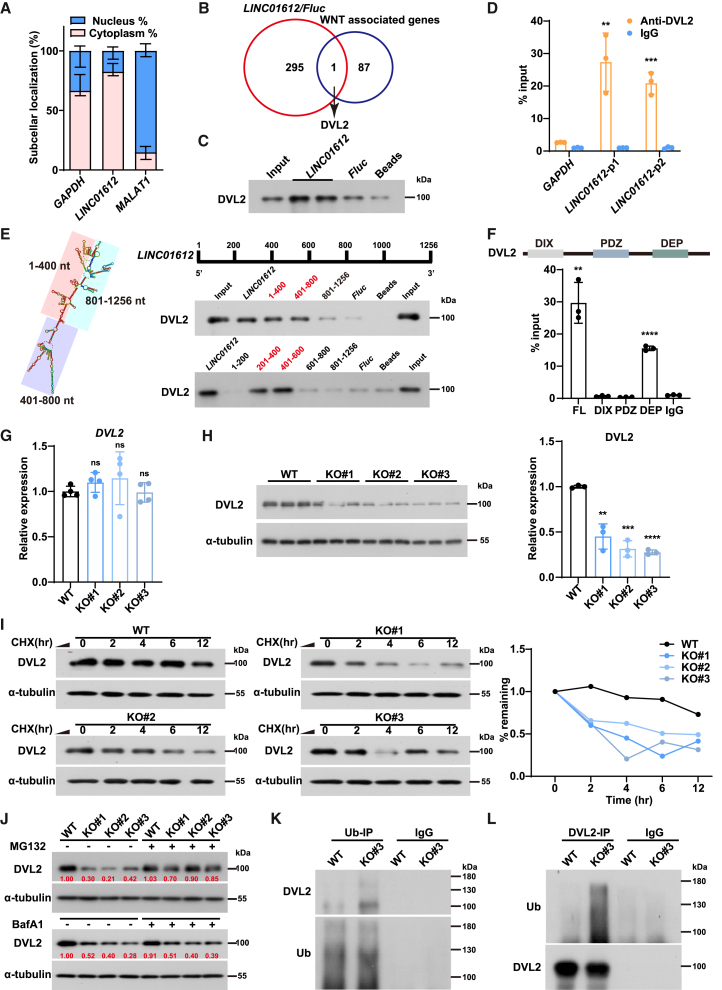 Figure 5
Figure 5 Figure 6
Figure 6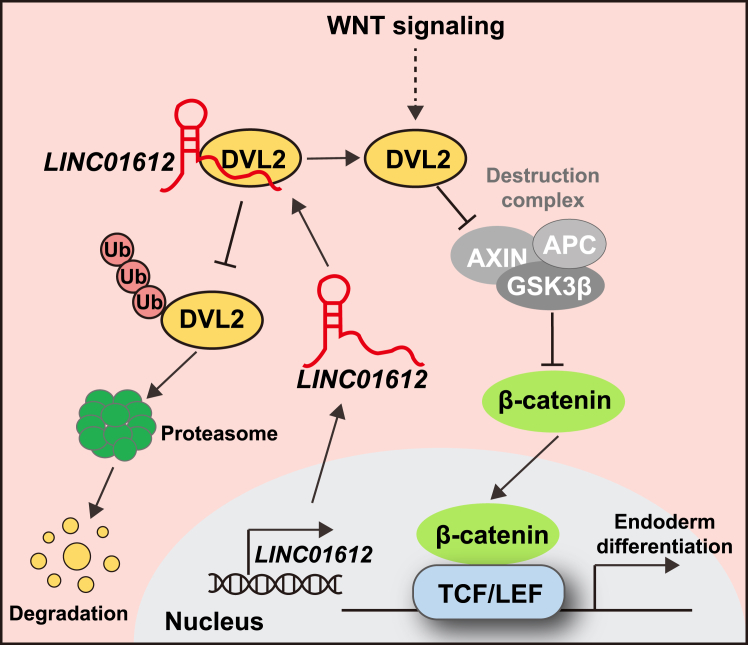 Figure 7
Figure 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCancer-related molecular mechanisms research · RNA modifications and cancer · RNA Research and Splicing
Introduction
Long noncoding RNAs (lncRNAs) are a diverse class of RNA molecules that are longer than 200 nucleotides without protein-coding potential, playing crucial roles in gene regulation (Chen and Kim, 2024). Over the past decade, accumulating evidence has revealed that lncRNAs participate in a wide range of biological processes, including chromatin remodeling, transcriptional regulation, post-transcriptional modifications, and signal transduction (Lu et al., 2021). LncRNAs regulate gene expression in a context-dependent manner by interacting with DNA, RNA, or proteins to act as scaffolds, decoys, or guides (Mattick et al., 2023; Wang and Chang, 2011). In general, cis-acting lncRNAs, which constitute a substantial portion of functionally characterized lncRNAs, regulate nearby gene expression across varying genomic distances (Gil and Ulitsky, 2020; Quinn and Chang, 2016). In contrast, *trans-*acting lncRNAs, particularly those located distally from protein-coding genes (PCGs) and named as “desert lncRNAs (far away from PCGs more than 50 kb in the genome)” (Lu et al., 2023), are transcribed and translocated from their sites of origin to regulate gene expression at distant genomic loci (Kopp and Mendell, 2018; Statello et al., 2021). Our early studies showed that desert lncRNAs exhibit high expression levels specific to certain cell types or developmental stages compared to those lncRNAs near PCGs (Lu et al., 2023; Yang et al., 2025). These findings highlight potential stage-specific functions of desert lncRNAs mediated by distinct regulatory mechanisms. However, the study of desert lncRNAs remains limited, primarily because of the difficulties in uncovering their biological roles and pinpointing their downstream targets.
Embryonic stem cells (ESCs), which originate from the inner cell mass (ICM) of the blastocyst, can self-renew and differentiate into the three germ layers: ectoderm, mesoderm, and definitive endoderm (DE) (Thomson et al., 1998; Zhai et al., 2022). Due to the ethnic and technical limitations to access human materials, human ESCs offer a valuable model for studying early development, disease modeling, drug screening, and cell therapy (Yiangou et al., 2018). During embryonic patterning and the establishment of germ layers, a combination of elevated Activin or Nodal signaling with moderate WNT signaling collaboratively triggers the DE differentiation (D'Amour et al., 2005; Loh et al., 2014). Activating both signaling pathways further results in the expression of key endodermal transcription factors, such as SOX17, FOXA2, and GATA4/6 (Ang et al., 1993; Heslop et al., 2021; Viotti et al., 2014). The DE further gives rise to the respiratory and gastrointestinal systems, along with the derivate tissues and organs (Zorn and Wells, 2009), holding great significance for regenerative medicine and disease studies.
Accumulating evidence underscores the significance of lncRNAs as essential regulators during embryonic development and cell differentiation. LncRNAs control the lineage specification and differentiation potential of ESCs, guiding processes such as neurogenesis, myogenesis, and cardiogenesis and the differentiation into endodermal lineages (Flynn and Chang, 2014; Lu et al., 2021; Mirzadeh Azad et al., 2021; Yan et al., 2017). For instance, lncRNAs DEANR1 and GATA6-AS1 are vital for endoderm differentiation by promoting the expression of nearby DE genes FOXA2 and GATA6, respectively, by recruiting SMAD2/3 to their promoter regions (Jiang et al., 2015; Yang et al., 2020). The lncRNA DIGIT could regulate endoderm differentiation by modulating GSC transcription (Daneshvar et al., 2016); moreover, DIGIT interacts with BRD3 to promote phase separation, facilitating BRD3 binding to H3K18ac-enriched regions of endodermal transcription factors (Daneshvar et al., 2020). A notable observation is that these endoderm-associated lncRNAs identified so far are located near PCGs and exert regulatory effects on the transcription of neighboring endoderm-related genes, thereby playing a role in endoderm differentiation. Our group recently demonstrated that a desert lncRNA HIDEN enhances the interaction between IMP1 protein and FZD5 mRNA, promoting human DE differentiation (Lu et al., 2023). However, many other desert lncRNAs are still awaiting functional dissection.
This study identifies LINC01612 as a new DE-specific desert lncRNA highly expressed during human DE differentiation. Its depletion significantly impairs human endoderm differentiation by downregulating WNT activity. We further dissect the underlying mechanism and reveal that LINC01612 interacts with and stabilizes DVL2 by reducing its ubiquitination, thereby promoting WNT signaling. These findings reveal that the LINC01612-DVL2-WNT axis is essential for human DE differentiation.
Results
LINC01612 is a lncRNA highly expressed in human endoderm lineage
To investigate the role of desert lncRNAs (more than 50 kb far away from PCGs in the genome) during human early differentiation, we conducted transcriptome analysis on our published data (GEO: GSE44875) using ESCs and sorted CD184/CD117-double positive DE cells (Jiang et al., 2015), identifying 46 differentially expressed desert lncRNAs (Lu et al., 2023). Notably, LINC01612 exhibited significantly elevated expression in DE cells (Figures 1A and S1A). A time-course analysis revealed a gradual upregulation of LINC01612 during DE differentiation (Figure 1B). Furthermore, analysis of LINC01612 expression in 30 human tissues from the Genotype-Tissue Expression (GTEx) database demonstrated its high expression in endoderm-derived tissues such as the liver and lung (Figure S1B). To assess the accurate expression level of LINC01612 at the single-cell level, we performed droplet digital PCR, and the result revealed that in HUES8-DE cells, the copy number of LINC01612 is comparable to that of lncRNA MALAT1 (Figure 1C), a key lncRNA that is highly expressed in various cancers and during development and mainly localizes to nuclear speckles to regulate cell proliferation via mechanisms involving alternative splicing and transcriptional control (Amodio et al., 2018). Given the imprecise annotation of lncRNAs (St Laurent et al., 2015), we conducted 5′ and 3′ rapid amplification of cDNA ends (RACE) experiments, revealing two isoforms of LINC01612 differing by a single region: one was 1,256 nucleotides and the other was 1,085 nucleotides (Figure 1D and Table S1). Next, we further assessed the expression levels of both isoforms and found that the longer isoform accounts for more than 85% (Figure 1D). Therefore, we later concentrated on the longer isoform for our subsequent studies.Figure 1LINC01612 is a noncoding RNA highly expressed in human endoderm lineage(A) Scatterplot of differentially expressed desert lncRNAs between ESCs and DE cells. Red indicates higher expression, while blue indicates lower expression.(B) Time-course expression of LINC01612 during human endoderm differentiation, detected by RT-qPCR (n = 3 independent experiments).(C) The copy number per cell of LINC01612 in HUES8-derived DE cells, detected by droplet digital PCR. MALAT1 was used as a control.(D) Diagram of the LINC01612 gene locus. Two isoforms of LINC01612 were identified by 5′ and 3′ RACE and validated by RT-PCR. The relative expression levels of the two isoforms were calculated according to PCR analysis (n = 3 independent experiments).(E) The protein-coding potential of LINC01612 was predicted using ORF Finder online tool.(F and G) Top panel: schematic representation of endoderm differentiation of human ESCs. Bottom panel: Flow cytometric analysis (F) and western blot (G) showing the expression of ORF-EGFP (n = 2 independent experiments) in ORF1/3 knockin HUES8-DE cells. SBP-EGFP (overexpressing EGFP in HEK293T cells) was used as a positive control to confirm the expression of GFP vector.(H) Flow cytometric analysis of CXCR4-positive cells in shRNA control (shC) and LINC01612-KD DE cells. The statistical results were shown on the right (n = 3 independent experiments).(I) Immunofluorescent staining of SOX17 and OCT4 in shC and LINC01612-KD DE cells. Scale bar, 50 μm.(J) The RNA levels of endoderm genes (SOX17, FOXA2, CXCR4, and GATA6) in LINC01612-KD and control DE cells (n = 3 independent experiments).Data are presented as the mean ± SD. Significance levels are indicated as ∗∗p < 0.01, ∗∗∗p < 0.001 and ∗∗∗∗p < 0.0001.
Recently, an increasing number of lncRNAs containing small open reading frames (smORFs) have been reported to translate into functional micropeptides (Andrews and Rothnagel, 2014; Pang et al., 2018). To test such a possibility, we used ORF Finder (https://www.ncbi.nlm.nih.gov/orffinder/) to predict whether LINC01612 could encode a micropeptide. We identified four potential smORFs (Figures 1E and S1C). To examine the coding potential of LINC01612, we fused EGFP (lacking the start codon ATG) in frame to the C terminus of these four smORFs within the LINC01612 transcript, immediately upstream of their stop codons (Figure S1C). After transfecting these plasmids into HEK293T cells, we observed the expression of ORF1-EGFP and ORF3-EGFP fusion proteins (Figures S1D–S1F), similar to the reported coding micropeptide as a positive control (D'Lima et al., 2017). Moreover, when the EGFP sequence (without the start codon ATG) was fused to the C terminus of ORF1 and ORF3 following their stop codons, it abolished the expression of the ORF1-EGFP and ORF3-EGFP fusion proteins (Figures S1D and S1F), indicating that ORF1 and ORF3 likely encode micropeptides in HEK293T cells. We further performed CRISPR-Cas9-mediated in situ knockin of ORF-EGFP in HUES8 ESCs to verify the endogenous translational activity of ORF1 and ORF3 and finally generated four EGFP knockin clones (two clones for each ORF). Then, wild-type and these ORF-EGFP knockin ESCs were subjected to DE differentiation induced by Activin signaling (Figure 1F). However, we failed to observe any ORF-EGFP signal by flow cytometry analysis or western blot, although the differentiation was successfully validated by CXCR4 (CD184)-based flow cytometric analysis (Figures 1F, 1G, and S1G). Taken together, these findings indicate that LINC01612 is highly expressed but less likely encodes protein during DE differentiation, despite the fact that we could not exclude the coding potential in other contexts.
To explore the function of LINC01612 in DE differentiation, we established two stable LINC01612-knockdown (KD) ESC lines using shRNAs, achieving at least 85% knockdown efficiency as determined by RT-qPCR (Figures S1H and S1I). No significant differences in pluripotent markers, such as OCT4 (POU5F1) and SSEA4, were observed between control and LINC01612-KD ESCs (Figure S1H). Importantly, we found that LINC01612-KD cells exhibited lower DE differentiation efficiency, evident by CXCR4-based flow cytometric analysis (Figure 1H) and further supported by the expression of SOX17 using immunofluorescence analysis (Figure 1I). RT-qPCR assay showed that the RNA levels of endoderm genes (SOX17, FOXA2, CXCR4, and GATA6) were significantly reduced (Figure 1J) and the expression of pluripotent genes (SOX2 and OCT4) and mesodermal genes (MIXL1 and T) were increased in LINC01612-KO DE cells compared with control (Figure S1J). Taken together, these data indicate that LINC01612 KD severely impairs human ESC differentiation toward DE.
LINC01612 is required for endoderm differentiation
Furthermore, we employed CRISPR-Cas9-mediated genomic deletion to completely abolish LINC01612 function. Based on the observed signal enrichment of assay for transposase-accessible chromatin using sequencing (ATAC-seq) (GEO: GSE285132) indicating chromatin accessibility and H3K27ac chromatin immunoprecipitation sequencing (ChIP-seq) peaks (data from ENCODE: https://www.encodeproject.org/experiments/ENCSR200ETW/) marking transcriptionally active regions (Figure S2A), we defined the genomic region adjacent to exon 1 of LINC01612 as its promoter and accordingly designed two single guide RNAs (sgRNAs) to delete this region to achieve complete ablation (Figures 2A and S2A). Genomic PCR and RT-qPCR results validated successful knockout and complete deletion of LINC01612 transcript in three LINC01612-KO clones with different genotypes (Figures 2A, 2B, and S2B). We found that the mRNA and protein expression levels of pluripotency-associated genes were not significantly altered upon LINC01612-KO (Figures S2C and S2D). Next, we subjected the LINC01612-KO and wild-type ESCs to DE differentiation. Flow cytometric analysis showed that DE differentiation was compromised (Figure 2C), and the expression of key endodermal genes, including SOX17, FOXA2, and CXCR4, was significantly reduced upon LINC01612-KO, whereas pluripotency- and mesoderm-associated genes were upregulated (Figures 2D and S2E). Similarly, western blot analysis and immunofluorescence staining revealed a significant reduction of SOX17 and FOXA2 protein levels in LINC01612-KO DE cells (Figures 2E and 2F). Taken together, the loss of LINC01612 results in a pronounced impairment in DE differentiation of human ESCs.Figure 2. Knockout of LINC01612 severely affects DE differentiation(A) The genotypes of the three LINC01612-KO HUES8 ESC lines. The sgRNA target sequences and PAM sequences were indicated in red and underlined, respectively.(B) The LINC01612 expression levels in wild-type and LINC01612-KO DE cells, examined by RT-qPCR using three sets of primers illustrated in (A) (n = 3 independent experiments).(C) Flow cytometric analysis of SOX17- and CXCR4-positive cells in wild-type and LINC01612-KO DE cells. The statistical results of double-positive cells were shown on the right (n = 6 independent experiments).(D) The RNA levels of endoderm genes (SOX17, FOXA2 and CXCR4) in wild-type and LINC01612-KO DE cells (n = 3 independent experiments).(E) The protein levels of DE markers (SOX17 and FOXA2) in wild-type and LINC01612-KO DE cells, determined by western blot. Quantitative results were shown on the right (n = 3 independent experiments).(F) Immunofluorescent staining of DE markers (SOX17 and FOXA2) in wild-type and LINC01612-KO DE cells. Quantitative results were shown at the bottom (n = 6 images). Scale bar, 50 μm.(G) Volcano plot showing differentially expressed genes identified by RNA-seq of wild-type and LINC01612-KO DE cells. Upregulated and downregulated genes upon LINC01612-KO were shown in red and green, respectively.(H) GO enrichment analysis of upregulated and downregulated genes in LINC01612-KO DE cells compared to wild-type, respectively.Data are presented as the mean ± SD. Significance levels are indicated as ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001 and ∗∗∗∗p < 0.0001; ns, not significant.
Next, we performed RNA sequencing (RNA-seq) to evaluate the transcriptomic change upon LINC01612 deletion during DE differentiation (Table S2). We identified 1,079 downregulated genes and 1,313 upregulated genes, and consistent with RT-qPCR results, the DE marker genes (SOX17, FOXA2, CXCR4, and GATA4/6) were downregulated, while the pluripotent genes (POU5F1 and SOX2) were upregulated (Figure 2G). In addition, gene ontology (GO) analysis revealed that the terms for downregulated genes in LINC01612-KO DE cells enriched in endoderm differentiation, FOXA2 pathway, and transforming growth factor β (TGF-β) for EMT. In contrast, the upregulated genes were related to ESC pluripotency pathways, axon guidance, and neural crest cell migration during development (Figure 2H and Table S3). Gene set enrichment analysis (GSEA) also showed a significant downregulation of DE-specific genes and an upregulation of ESC-specific genes in LINC01612-KO DE cells (Figure S2F).
To verify the important role of LINC01612 in lineage differentiation, we performed embryoid body (EB) differentiation and analyzed gene expression by RT-qPCR (Figures S3A and S3B). Compared to wild-type cells, LINC01612-KO cells showed reduced expression of endoderm markers, while ectoderm markers were obviously upregulated (Figure S3B), supporting the crucial role of LINC01612 in the development of endoderm lineages. However, the expression of mesoderm markers was somehow chaotic: T was significantly upregulated, while MIXL1 was downregulated to a certain extent (Figure S3B). To further investigate the role of LINC01612 during the progressive differentiation of mesendodermal cells into more specific lineages, we further induced endodermal pancreatic differentiation (Li et al., 2024) and mesodermal heart progenitor differentiation (Yang et al., 2024) in wild-type and LINC01612-KO cells. We observed significant downregulation of pancreatic transcription factors in LINC01612-KO cells, including PDX1 at pancreatic progenitor (PP) 1 stage and PDX1 and NKX6-1 at PP2 stage (Figures S3C and S3D). Similarly, the expression of heart progenitor markers (MEF2C, ISL1, GATA4, and TNNT2) was reduced as well (Figure S3E). Collectively, these results underscore the essential function of LINC01612 in human mesendodermal lineage differentiation.
WNT signaling pathway acts as the downstream of LINC01612
To understand how LINC01612 affected human endoderm differentiation, we performed GO analysis of differentially expressed genes in LINC01612-KO cells, showing enrichment in pathways related to pluripotent stem cell differentiation pathway, WNT signaling, and TGF-β receptor signaling (Figure 3A and Table S3). Given the critical role of the WNT signaling pathway in endoderm differentiation (Dziedzicka et al., 2021; Jiang et al., 2013), we hypothesized that the WNT signaling pathway might function downstream of LINC01612. Therefore, we analyzed the protein levels of active (unphosphorylated, nuclear-located) β-catenin, the key effector of the WNT signaling pathway, in wild-type and LINC01612-KO DE cells. Both active and total β-catenin levels were significantly diminished in LINC01612-KO DE cells compared to wild-type cells (Figure 3B). We also observed a significant increase in both active and total β-catenin protein levels in LINC01612-overexpressing HEK293T cells (Figures S4A and S4B). Additionally, the β-catenin/TCF-responsive luciferase reporter assay in HEK293T cells revealed that LINC01612 overexpression resulted in significantly increased TCF luciferase activity (Figure 3C). These findings suggest that LINC01612 depletion, indeed, impairs WNT signaling pathway during DE differentiation.Figure 3WNT signaling pathway acts as the downstream of LINC01612(A) GO enrichment analysis of differentially expressed genes in wild-type and LINC01612-KO DE cells.(B) The levels of active β-catenin and total β-catenin in wild-type and LINC01612-KO DE cells, determined by western blot. Quantitative results were shown on the right (n = 3 independent experiments).(C) The TCF-luciferase activity in HEK293T cells transfected with LINC01612. The sample treated with 1 μM CHIR-99021 was used as a positive control (n = 3 independent experiments).(D) Left panel: schematic representation of endoderm differentiation of human ESCs. A, Activin A; AC, Activin A and CHIR-99021; AW, Activin A and Wnt3a. Right panel: the mRNA levels of endoderm marker genes, including SOX17 and FOXA2, detected by RT-qPCR (n = 3 independent experiments).(E and F) Flow cytometric analysis of SOX17-and CXCR4-positive cells (E) in wild-type and LINC01612-KO DE cells treated with WNT activators during DE differentiation. The statistical results of double-positive cells (F) were shown (n = 3 independent experiments).(G) The protein levels of DE markers (SOX17 and FOXA2) in wild-type and LINC01612-KO DE cells treated with WNT activators during DE differentiation, determined by western blot.Data are presented as the mean ± SD. Significance levels are indicated as ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001 and ∗∗∗∗p < 0.0001; ns, not significant.
WNT signal activation is essential for efficient DE differentiation, particularly during the early mesendoderm differentiation phase (Jiang et al., 2013; Loh et al., 2014). We wondered whether the activation of WNT signalg could rescue the impaired DE differentiation observed in LINC01612-KO cells. Therefore, we treated LINC01612-KO cells with either GSK3 inhibitor CHIR-99021 or WNT ligand protein Wnt3a (both function as WNT signal activators) (Figure 3D). RT-qPCR analysis revealed a significant restoration of DE-associated genes (SOX17 and FOXA2) in LINC01612-KO DE cells treated with CHIR-99021 or Wnt3a (Figure 3D). Consistently, flow cytometry and western blot analyses also showed significant restoration of SOX17 and FOXA2 expression in LINC01612-KO DE cells treated with CHIR-99021 or Wnt3a (Figures 3E–3G and S4C). These results suggest that modulating the WNT signaling pathway can resolve the DE differentiation defects induced by LINC01612 disruption. This supports the notion that WNT signaling is the functional downstream of LINC01612, critical in regulating DE differentiation.
LINC01612 physically interacts with and stabilizes DVL2 protein by reducing ubiquitination
To investigate how LINC01612 affects WNT activity and contributes to DE differentiation, we first assessed its subcellular localization in DE cells. We separated the cytoplasmic and nuclear fractions of DE cells and found that the LINC01612 transcript was predominantly localized in the cytoplasmic fraction (Figure 4A). Thus, we performed RNA pull-down assay using biotin-labeled LINC01612 and the negative control luciferase (Fluc) mRNA in DE cells to identify its interacting proteins (Figure S5A). The whole immunoprecipitated extracts of LINC01612 and Fluc were then subjected to label-free quantitative mass spectrometry (Table S4). We identified 296 potential interacting proteins, among which DVL2, a WNT-associated protein, was found to directly interact with LINC01612 (Figure 4B). The interaction of LINC01612 with DVL2 was confirmed by western blot analysis following RNA pull-down (Figure 4C). Moreover, RNA immunoprecipitation (RIP) assay further confirmed the interaction between LINC01612 and DVL2 (Figure 4D). These results indicate that LINC01612 interacts with DVL2 protein.Figure 4LINC01612 physically interacts with and stabilizes DVL2 protein by reducing its ubiquitination(A) Subcellular localization of LINC01612 in DE cells by RT-qPCR (n = 6 independent experiments).(B) Venn diagram indicating the overlapped hits identified by label-free quantitative mass spectrometry (log_2_(fold-change of LINC01612/Fluc) > 1) and WNT-associated genes.(C) Immunoblot for DVL2 after LINC01612 RNA pull-down in DE cells. Beads and Fluc were used as negative controls.(D) RIP assays showed that DVL2 enriched LINC01612 in DE cells (n = 3 independent experiments).(E) Left: the secondary structure of LINC01612, predicted via RNAfold. Right: immunoblot for DVL2 after truncated LINC01612 fragment RNA pull-down in HEK293T cells.(F) RIP assays were performed to identify the regions in DVL2 that mediate their interactions with LINC01612 in HEK293T cells transfected with FLAG-tagged DVL2 and truncation mutants of DVL2.(G) The RNA levels of DVL2 in wild-type and LINC01612-KO DE cells, determined by RT-qPCR (n = 4 independent experiments).(H) The protein levels of DVL2 in wild-type and LINC01612-KO DE cells, determined by western blot analysis. Quantitative results were shown on the right (n = 3 independent experiments).(I) Effect of LINC01612 on the endogenous protein levels of DVL2 in wild-type and LINC01612-KO DE cells treated with cycloheximide. Quantitative results were shown on the right.(J) Effect of LINC01612 on the endogenous protein levels of DVL2 in wild-type and LINC01612-KO DE cells treated with MG132 or BafA1.(K and L) Ubiquitination-immunoprecipitation (K) and DVL2-IP (L) assays, showing the ubiquitination levels of DVL2 in wild-type and LINC01612-KO DE cells.Data are presented as the mean ± SD. Significance levels are indicated as ∗∗p < 0.01, ∗∗∗p < 0.001 and ∗∗∗∗p < 0.0001; ns, not significant.
To identify the direct binding region between LINC01612 and DVL2, we first employed RNAfold (http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi) to predict the secondary structure of LINC01612 (Figure 4E). Based on the predicted secondary structure, we divided LINC01612 into three fragments: 1–400 nucleotides (nts), 401–800 nts, and 801–1,045 nts. We then generated biotin-labeled truncated fragments of LINC01612 for RNA pull-down assays in HEK293T cells. The results revealed that the 1-400-nt and 401-800-nt regions of LINC01612 can significantly bind to DVL2 (Figure 4E). We further truncated the 1-400-nt and 401-800-nt regions of LINC01612 into four smaller fragments and found that the fragments of 201–400 nts and 401–600 nts were primarily responsible for the interaction between LINC01612 and DVL2 (Figure 4E). On the other hand, we truncated the DVL2 protein based on its domain annotation, which includes an N-terminal DIX domain, a central PDZ domain, and a C-terminal DEP domain (Boutros and Mlodzik, 1999). Each truncated construct was FLAG tagged and co-expressed with LINC01612 in HEK293T cells (Figures 4F and S5B). Subsequent RIP-qPCR results showed that LINC01612 was most highly enriched in the DVL2 truncation containing the DEP domain, compared to the other mutants (Figure 4F). These results suggest that the 201- to 600-nt fragment of LINC01612 and the DEP domain of DVL2 contribute to the physical interaction with each other.
DVL2, as a component of the WNT signaling pathway, plays a pivotal role in stabilizing β-catenin by disrupting the β-catenin degradation complex (Kang et al., 2022). The time course analysis showed that DVL2 exhibited a progressive upregulation during DE differentiation (Figure S5C). To test whether LINC01612 regulates DVL2, we examined the mRNA and protein levels of DVL2 in LINC01612-KO DE cells. Interestingly, although RT-qPCR results showed that the mRNA level of DVL2 was unaltered (Figures 4G and S5D), the protein level of DVL2 was significantly downregulated in LINC01612-KO DE cells compared to wild-type (Figure 4H). Meanwhile, we observed a significant increase in DVL2 protein levels, but not in its mRNA expression, upon LINC01612 overexpression in HEK293T cells (Figures S5E and S5F). These results indicate that LINC01612 may interact with DVL2 protein and regulate its protein stability.
To confirm the protein stability regulation of DVL2 by LINC01612, we performed the protein synthesis inhibitor cycloheximide chase assays, revealing that LINC01612-KO indeed reduced DVL2 stability in DE cells (Figure 4I). This conclusion was further confirmed by the result that LINC01612 overexpression enhanced DVL2 stability in HEK293T cells (Figure S5G). Since protein degradation is primarily mediated by the ubiquitin-proteasome system or the autophagy-lysosome pathway (Pohl and Dikic, 2019), we subsequently employed proteasome inhibitor MG132 or lysosome inhibitor bafilomycin A1 (BafA1) to elucidate the mechanism underlying DVL2 degradation. The results showed that MG132 obviously restored the change of endogenous DVL2 protein levels in both DE cells due to depletion of LINC01612 and in HEK293T cells with LINC01612 overexpression, but the lysosome inhibitor BafA1 had no effect on the expression of DVL2 (Figures 4J and S5H). In addition, both ubiquitination-immunoprecipitation (IP) and DVL2-IP assays showed that LINC01612-KO resulted in increased ubiquitination of DVL2 (Figures 4K and 4L). These findings indicate that LINC01612 interacts with DVL2 protein and contributes to the stabilization of DVL2 by impeding ubiquitin-mediated proteasomal degradation.
LINC01612 regulates endoderm differentiation through DVL2
Since the involvement of the LINC01612-interacting protein DVL2 in DE differentiation was yet undocumented, we also explore whether DVL2 contributes to this process. We established three stable ESC lines of DVL2-KD without affecting the expression of pluripotency-associated genes (Figures S6A–S6C). Next, we subjected the DVL2-KD ESCs to DE differentiation. Intracellular flow cytometric analysis showed a significant decrease in the number of SOX17 and CXCR4 double-positive cells (Figure 5A). Consistently, RT-qPCR analysis showed a corresponding decrease in the expression of endodermal genes (SOX17, FOXA2, and CXCR4), along with elevated levels of pluripotent genes (OCT4, SOX2, and NANOG) and disrupted expression of mesodermal genes (EOMES and T) in DVL2-KD DE cells (Figure S6D). Furthermore, differentiated DE cells upon DVL2 depletion exhibited decreased SOX17, FOXA2, and active and total β-catenin protein levels (Figures 5B and 5C), which phenocopied the LINC01612 depletion. These results together confirm the essential role of DVL2 in endoderm fate determination.Figure 5DVL2 counteracts the effects of LINC01612 deficiency(A) Flow cytometric analysis of SOX17- and CXCR4-positive cells in shRNA control (shC) and DVL2-KD DE cells. The statistical results were shown on the right (n = 3 independent experiments).(B) The protein levels of DVL2, SOX17, and FOXA2 in DVL2-KD and control DE cells. Quantitative results were shown on the right (n = 3 independent experiments).(C) The levels of active β-catenin and total β-catenin in DVL2-KD and control DE cells, determined by western blot. Quantitative results were shown at the bottom (n = 3 independent experiments).(D) Venn diagram indicating the significant overlap of differentially expressed genes due to LINC01612-KO and DVL2-KD in DE cells.(E) GO enrichment analysis of overlapped differentially expressed genes in LINC01612-KO and DVL2-KD DE cells.(F) Flow cytometric analysis of the SOX17-/CXCR4-double positive cells in differentiated wild-type, LINC01612-KO, and DVL2-overexpressing cells with LINC01612-KO (n = 3 independent experiments).(G) The RNA levels of endoderm genes (SOX17, FOXA2, and CXCR4) for the samples shown in (F) (n = 3 independent experiments).(H) Immunofluorescent staining of FOXA2 for the samples shown in (F). Quantitative results were shown on the right (n = 5 images). Scale bar, 100 μm.(I) Western blot analysis of DVL2 and active β-catenin levels for the samples shown in (F).Data are presented as the mean ± SD. Significance levels are indicated as ∗∗p < 0.01, ∗∗∗p < 0.001 and ∗∗∗∗p < 0.0001; ns, not significant.
Next, we performed RNA-seq using DVL2-KD and control DE cells and identified 681 downregulated genes and 1,442 upregulated genes (Table S2). Consistent with RT-qPCR results (Figure S6D), the DE marker genes (SOX17, FOXA2, and CXCR4) were downregulated, while the pluripotent genes (POU5F1 and NANOG) were upregulated upon DVL2-KD (Figure S6E). GO analysis further showed downregulated genes in DVL2-KD DE cells enriched in terms including endoderm differentiation, WNT signaling, heart development, and EMT (Figure S6F and Table S3), similar to the results in LINC01612-KO DE cells (Figure 2H). GSEA also indicated the downregulation of DE-specific genes and upregulation of ESC-specific genes upon DVL2 KD (Figure S6G). To further investigate the relationship between LINC01612 and DVL2, we performed an integrated analysis of RNA-seq datasets derived from LINC01612-KO and DVL2-KD DE cells. We found that there was a significant overlap between LINC01612- and DVL2-regulated genes, with 326 genes co-downregulated, including DE marker genes (SOX17, FOXA2, and CXCR4), and 724 genes co-upregulated, encompassing pluripotent genes (POU5F1, DNMT3B, and FOXD3) (Figure 5D). Furthermore, GO analysis revealed that co-downregulated genes enriched in terms of FOXA2 pathway, mesodermal commitment pathway, and endoderm differentiation. In contrast, the co-upregulated genes were associated with ESC pluripotency pathways, neural crest cell migration during development, regulation of actin cytoskeleton, and synaptic vesicle pathway (Figure 5E and Table S3). In addition to their cooperative role in endoderm differentiation, LINC01612 and DVL2 also independently regulate distinct sets of genes involved in various metabolic and signaling pathways (Figure S6H). These findings suggest that LINC01612 and DVL2 function within the same pathway to influence endoderm differentiation.
To determine whether LINC01612 exerts its function through DVL2, we performed DVL2 overexpression experiments in LINC01612-KO cells. Significantly, DVL2 overexpression effectively restored DE differentiation in LINC01612-KO cells and rescued the expression of DE markers at both mRNA and protein levels (Figures 5F–5H and S6I). Additionally, DVL2 overexpression was able to increase the active β-catenin protein level and rescue WNT signaling activity (Figure 5I). Altogether, these data demonstrate that LINC01612 enhances WNT signaling activity and DE differentiation via stabilizing DVL2 protein (Figure 6).Figure 6. Model of the role of LINC01612-DVL2-WNT axis in endoderm differentiationDuring human endoderm differentiation, LINC01612 enhances WNT signaling activity by binding to DVL2 protein, reducing its ubiquitination, and thereby protecting DVL2 protein from proteasomal degradation.
Discussion
This study dissects the biological function and molecular mechanisms of a new desert lncRNA LINC01612 during human endoderm differentiation. Our findings demonstrate that as a highly expressed lncRNA during DE differentiation, LINC01612 is essential for the process, evident by the data that the disruption through shRNA KD or promoter deletion significantly impairs DE differentiation (Figures 1 and 2). Furthermore, depletion of LINC01612 leads to reduced WNT signaling activity, coupled with the restoration of impaired DE differentiation by WNT signaling activators, strongly suggesting that the WNT signaling pathway serves as the functional downstream of LINC01612 in promoting DE differentiation (Figure 3). Mechanistically, we show that LINC01612 physically interacts with and stabilizes DVL2 protein by reducing its ubiquitination (Figure 4). DVL2-KD results in suppression of endoderm differentiation. At the same time, DVL2 overexpression rescues the defects caused by LINC01612 deficiency, further demonstrating that DVL2 is the functional target of LINC01612 (Figure 5). Our study highlights the critical regulatory role of LINC01612-DVL2-WNT axis in human endoderm differentiation (Figure 6).
LncRNAs are transcribed by RNA polymerase II and lack long and conserved ORFs (Chen and Kim, 2024). However, it has been increasingly recognized that a subset of lncRNAs containing smORFs can give rise to micropeptides (Orr et al., 2020). In our results, LINC01612 contains four predicted smORFs, and the exogenous overexpression of ORF-EGFP fusion proteins revealed that ORF1 and ORF3 showed the capacity to be translated into micropeptides (Figures S1C–S1F). However, further in situ EGFP knockin experiments demonstrated that neither produced micropeptides endogenously (Figures 1F, 1G, and S1G). This suggests that utilizing a strong promoter for ORF-EGFP overexpression markedly increases its expression level and that the differences in cellular environments do not accurately reflect the endogenous translation events. Therefore, a comprehensive approach is crucial for determining whether lncRNAs can be translated into micropeptides, combining bioinformatic analysis, ribosome profiling, mass spectrometry, proteomics, monoclonal antibodies, and in situ fusion protein detection.
We have noticed that LINC01612 might have other functions in different biological contexts. An expression analysis suggested that LINC01612 is significantly downregulated in gastric cancer samples and closely linked to enhanced metastasis and advanced tumor stage (Song et al., 2016). Moreover, LINC01612 inhibits gastric cancer cell proliferation, induces apoptosis by binding to ATR and suppressing CHK1 phosphorylation, and enhances oxaliplatin sensitivity (Liu et al., 2024). In p53-expressing hepatocellular carcinoma cells, LINC01612 promotes ATF3 expression by sponging miR-494 and activating the p53 pathway, while in p53-null cells, LINC01612 binds to YBX1 and promotes its degradation to suppress tumor progression (Liu et al., 2022). In our case, LINC01612 stabilizes DVL2 protein, a key WNT pathway regulator, by reducing its ubiquitination, thereby playing a crucial role in the differentiation process. Such cell-type specificity stems from the distinct expression patterns of lncRNAs, their interactions with different proteins, and their involvement in signaling pathways adapted to cellular contexts. Whether the LINC01612-DVL2-WNT regulatory axis is conserved in certain developmental or pathological contexts involving WNT signaling awaits further investigation.
DVL2 is an essential modulator of the WNT signaling pathway, playing a role in maintaining β-catenin stability by disrupting the degradation complex formed by APC, AXIN, CK1α, and GSK3β (Kang et al., 2022; Schubert et al., 2022). In addition, DVL2/WNT/β-catenin axis has been identified as a driver of progression mediated by KLF12 in pancreatic cancer (He et al., 2019). Here, we discovered that depleting DVL2 impairs WNT signaling activity, subsequently affecting endoderm differentiation (Figure 5). Of note, in addition to their cooperative role in endoderm differentiation, LINC01612 and DVL2 independently regulate distinct sets of genes involved in various metabolic and signaling pathways as well (Figure S6H). Specifically, Qu and colleagues demonstrated the downregulation of genes in the Hippo/YAP signaling pathway and upregulation of genes involved in cell metabolism upon macrophage DVL2 deficiency (Qu et al., 2025), aligning well with our results. This consistency reinforces the role of DVL2 in regulating these pathways and supports the biological relevance of our observations.
Increasing evidence suggests that multiple E3 ligases, such as NEDD4L, ITCH, and TRIM56, bind to the C terminus of DVL2 and mediate DVL2 ubiquitination, ultimately promoting its degradation and attenuating WNT signaling activity (Ding et al., 2013; Wei et al., 2012; Yan et al., 2021). Meanwhile, USP14 acts as a deubiquitinating enzyme for DVL, facilitating its deubiquitylation and consequently enhancing WNT signaling activity (Jung et al., 2013). Our results showed that LINC01612 interacts with DVL2 via the 201- to 600-nt fragment of LINC01612 and the DEP domain of DVL2 (Figure 4). The DEP domain is essential for WNT signal transduction to the nucleus (Gammons et al., 2016) and harbors its ubiquitination site (Ding et al., 2013; Wei et al., 2012; Yan et al., 2021). It has been reported that lncRNAs can influence the ubiquitination of their associated proteins and regulate their stability (Bian et al., 2024). Nevertheless, it remains to be determined whether LINC01612 affects the ubiquitination of DVL2 through their interaction by either occupying the binding site on DVL2 for E3 ubiquitin ligases, thereby preventing ubiquitination, or by recruiting deubiquitinating enzymes to DVL2 via the non-interacting region of LINC01612 to reduce its ubiquitination. Further studies are needed to understand how LINC01612 regulates DVL2 protein stability.
In summary, here we report the role of a newly discovered desert lncRNA, LINC01612, in the human endoderm differentiation. LINC01612 binds to and stabilizes DVL2 protein, and the depletion of LINC01612 or DVL2 compromises WNT signaling activity and abolishes endoderm differentiation. These results reveal the role and action of the desert lncRNA, LINC01612, in early embryonic lineage specification and provide deeper insights into the regulatory mechanisms underlying cell fate decisions.
Materials and methods
Cell culture and differentiation
The human ESC line HUES8 from Harvard University was maintained in mTeSR1 medium (STEMCELL Technologies, Cat#85850) and 1% PS (penicillin-streptomycin) (Gibco, Cat#10378016) on Matrigel-coated plates at 37°C with 5% CO_2_. Cells were passaged every 3–4 days at 1:20 ratio by incubating cells with Accutase (Sigma, Cat#A6964) for 3 min. All the HUES8 lines tested negative for mycoplasma contamination. Our work on human ESCs is approved by the Biomedical Ethics Committee of Wuhan University (WHU-LFMD-IRB2024026). HEK293T cells were cultured in DMEM (Gibco, Cat#C11995500BT) supplemented with 10% FBS (fetal bovine serum) (Gibco, Cat#10100147) and 1% PS at 37°C with 5% CO_2_.
DE differentiation was performed following a previously described protocol (Lu et al., 2023; Yang et al., 2020). Briefly, 5∼8 × 10^4^ cells were seeded onto Matrigel-coated 24-well plates in mTeSR1. On the following day, the culture medium was replaced with DE induction medium consisting of DMEM/F12 (Gibco, Cat#C11330500BT), 0.2% BSA (bovine serum albumin) (YEASEN, Cat#B57370), 1% B27 without vitamin A (Shanghai BasalMedia, Cat#S441J7), and 100 ng/mL Activin A (PeproTech, Cat#120-14P). Cells were collected on day 3 for immunofluorescence, RT-qPCR, western blot, or flow cytometric analysis. The pancreatic lineage and heart progenitor differentiation were performed according to the published differentiation protocol (Li et al., 2024; Yang et al., 2024) and EB formation was carried out as previously described (Lan et al., 2022).
Plasmid constructs of shRNA knockdown and gene overexpression
The shRNAs targeting LINC01612, DVL2, and a scramble control were inserted into the lentiviral vector pLKO.1 plasmid. HEK293T cells were co-transfected with these lentiviral plasmids, expressing either the specific shRNAs or the scramble control (shC), along with lentiviral packaging plasmids (psPAX2 and pMD2.G) for lentivirus production. DVL2 CDS sequence was inserted into the doxycycline-induced pCW plasmid and co-transfected with lentiviral packaging plasmids into LINC01612-KO ESCs cells. Stable ESC lines were established through selection with 2 μg/mL puromycin for about 7 days. The targeting sequences were provided in Table S5.
CRISPR-Cas9-mediated knockout and knockin in human ESCs
Pairwise guide RNAs (gRNAs) targeting LINC01612 were designed using CHOPCHOP web tool (https://chopchop.cbu.uib.no/), and they were constructed in pX459 (pSpCas9(BB)-2A-Puro, Addgene, Cat#48139) expression plasmid. As for knockin, about 2,000-bp double-stranded donor DNA was gel-purified from the PCR products of genome sequence carrying EGFP. Briefly, 1 million HUES8 cells were electroporated with a total of 10 μg pX459 and donor DNA. The cells were then seeded onto a Matrigel-coated 6-well plate in mTesR1 medium supplemented with 10 μM Y-27632 (Selleck, Cat#S1049). After 24 h of culture, puromycin (1 μg/mL) (Santa Cruz Biotechnology, Cat#58582) was added for an additional 48 h. Single cells were subsequently sorted into Matrigel-coated 96-well plates using FACSAria III. Surviving colonies were expanded and genotyped through Sanger sequencing. The sequences of all sgRNAs and the primers for genomic sequencing were listed in Table S5.
RNA-seq and data analysis
Total RNA was sent to YINGZI GENE (Wuhan, China) for library preparation and sequencing. RNA-seq data were aligned to the human genome (hg38) using HISAT2 (Kim et al., 2019). FeatureCounts (v2.0.1) was used to count reads based on the GENCODE V29 gene transfer format, and gene expression was quantified using transcripts per million (TPM). Differential gene expression analysis for binary comparisons was carried out using the DESeq2 R package, with a fold-change cutoff >1, p value <0.05, and average TPM>10 (Love et al., 2014). GO analysis was performed using Enrichr (https://maayanlab.cloud/Enrichr/) (Kuleshov et al., 2016).
Statistical analysis
Data were presented as means ± SD from at least three independent experiments. The significance levels were determined using an unpaired t test in GraphPad Prism 9.5. p values were shown as ^∗^p < 0.05, ^∗∗^p < 0.01, ^∗∗∗^p < 0.001, and ^∗∗∗∗^p < 0.0001, with “ns” indicating no statistical significance.
Resource availability
Lead contact
Further information and requests for resources and reagents should be directed to the lead contact, Wei Jiang ([email protected]).
Materials availability
Plasmids and cell lines generated in this study are available from the lead contact upon request.
Data and code availability
The RNA-seq data in this study have been uploaded to the Gene Expression Omnibus (GEO) database under accession code GEO: [GSE291901](GSE291901).
Acknowledgments
We would like to thank laboratory members for technical help and insightful discussion. We thank the Core Facility of Medical Research Institute of Wuhan University and Large-Scale Instrument and Equipment Sharing Foundation of Wuhan University for technical support and equipment grant. This work was supported by grants from the National Natural Science Foundation of China (No. 32270857 to W.J. and No. 32400672 to P.L.), the China Postdoctoral Science Foundation (GZC20251853 and 2025M772807 to J.Y.) and the Fundamental Research Funds for the Central Universities in China (2042022dx0003 to W.J.).
Author contributions
W.J. conceived the project and designed the experiment together with M.L., P.L., and J.Y.; M.L. performed most of the bench experiments with help from P.L. and J.Y.; P.L. performed the initial screening, LINC01612-KD, and nucleo/cytoplasmic separation experiments; J.Y. performed the 5′ and 3′ RACE experiments; Y.Y. performed the ORF-EGFP experiments; C.Y. and J.Y. analyzed the next-generation sequencing data; M.L. drafted the manuscript, and W.J. and M.L. finalized the manuscript. All authors contributed to and approved the final manuscript.
Declaration of interests
The authors declare no competing interests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Amodio N.Raimondi L.Juli G.Stamato M.A.Caracciolo D.Tagliaferri P.Tassone P.MALAT 1: a druggable long non-coding RNA for targeted anti-cancer approaches J. Hematol. Oncol.1120186310.1186/s 13045-018-0606-429739426 PMC 5941496 · doi ↗ · pubmed ↗
- 2Andrews S.J.Rothnagel J.A.Emerging evidence for functional peptides encoded by short open reading frames Nat. Rev. Genet.15201419320410.1038/nrg 352024514441 · doi ↗ · pubmed ↗
- 3Ang S.L.Wierda A.Wong D.Stevens K.A.Cascio S.Rossant J.Zaret K.S.The formation and maintenance of the definitive endoderm lineage in the mouse: involvement of HNF 3/forkhead proteins Development 11919931301131510.1242/dev.119.4.13018306889 · doi ↗ · pubmed ↗
- 4Bian Z.Yang F.Xu P.Gao G.Yang C.Cao Y.Yao S.Wang X.Yin Y.Fei B.Huang Z.LINC 01852 inhibits the tumorigenesis and chemoresistance in colorectal cancer by suppressing SRSF 5-mediated alternative splicing of PKM Mol. Cancer 2320242310.1186/s 12943-024-01939-738263157 PMC 10807094 · doi ↗ · pubmed ↗
- 5Boutros M.Mlodzik M.Dishevelled: at the crossroads of divergent intracellular signaling pathways Mech. Dev.831999273710.1016/s 0925-4773(99)00046-510507837 · doi ↗ · pubmed ↗
- 6Chen L.L.Kim V.N.Small and long non-coding RN As: Past, present, and future Cell 18720246451648510.1016/j.cell.2024.10.02439547208 · doi ↗ · pubmed ↗
- 7D'Amour K.A.Agulnick A.D.Eliazer S.Kelly O.G.Kroon E.Baetge E.E.Efficient differentiation of human embryonic stem cells to definitive endoderm Nat. Biotechnol.2320051534154110.1038/nbt 116316258519 · doi ↗ · pubmed ↗
- 8D'Lima N.G.Ma J.Winkler L.Chu Q.Loh K.H.Corpuz E.O.Budnik B.A.Lykke-Andersen J.Saghatelian A.Slavoff S.A.A human microprotein that interacts with the m RNA decapping complex Nat. Chem. Biol.13201717418010.1038/nchembio.224927918561 PMC 5247292 · doi ↗ · pubmed ↗