The splicing factor PTBP1 interacts with RUNX1 and is required for leukemia cell survival
Arjun Dhir, Alexander Ethell, Riley Watkins, Calvin Lam, Kevin Tur-Rodriguez, Jimmie Persinger, Kasidy K. Dobish, Sipra Panda, Shannon M. Buckley, Samantha A. Swenson, Sandipan Brahma, M. Jordan Rowley, R. Katherine Hyde

TL;DR
This study shows that the splicing factor PTBP1 interacts with RUNX1 and is crucial for the survival of leukemia cells by regulating metabolic genes.
Contribution
The novel contribution is identifying PTBP1 as a RUNX1 interactor and demonstrating its role in leukemia cell survival through RNA splicing and metabolic regulation.
Findings
PTBP1 interacts with RUNX1 in an HDAC1-dependent manner and co-occupies promoter regions of actively transcribed genes.
Loss of PTBP1 disrupts RNA splicing and reduces expression of metabolic genes, impairing leukemia cell metabolism and survival.
Abstract
Runt-related Transcription Factor 1 (RUNX1) is essential for definitive hematopoiesis and is among the most frequently mutated genes in leukemia. Previous work from our lab demonstrated that Histone Deacetylase 1 (HDAC1), a known RUNX1 partner, is unexpectedly required for active transcription suggesting a non-histone role for HDAC1 in regulating components of the RUNX1 complex. Here, we use proteomics, genomics, and long-read transcriptomics to identify novel RUNX1 interacting partners and decipher their role in gene regulation and RNA splicing in leukemia cells. We demonstrate that Polypyrimidine Tract Binding Protein 1 (PTBP1) interacts with RUNX1 in an HDAC1-dependent manner. Chromatin profiling revealed extensive genome-wide overlap in sites occupied by RUNX1 and PTBP1, with significant enrichment at promoters of actively transcribed genes. Loss of PTBP1 in AML cells led to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
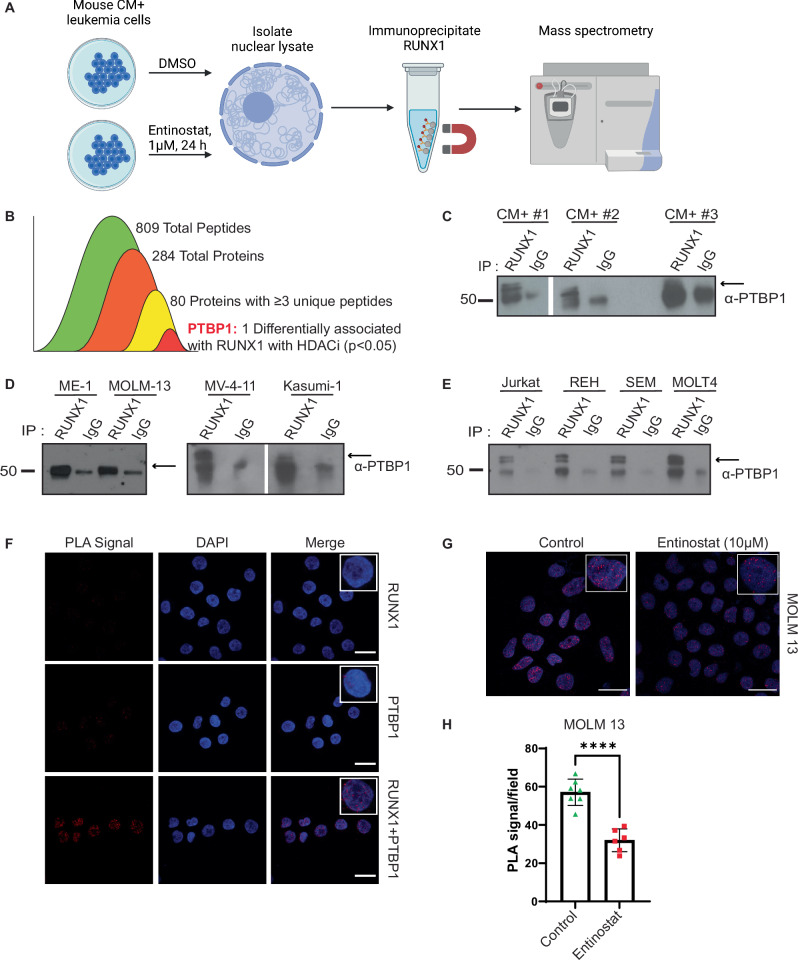 Figure 1
Figure 1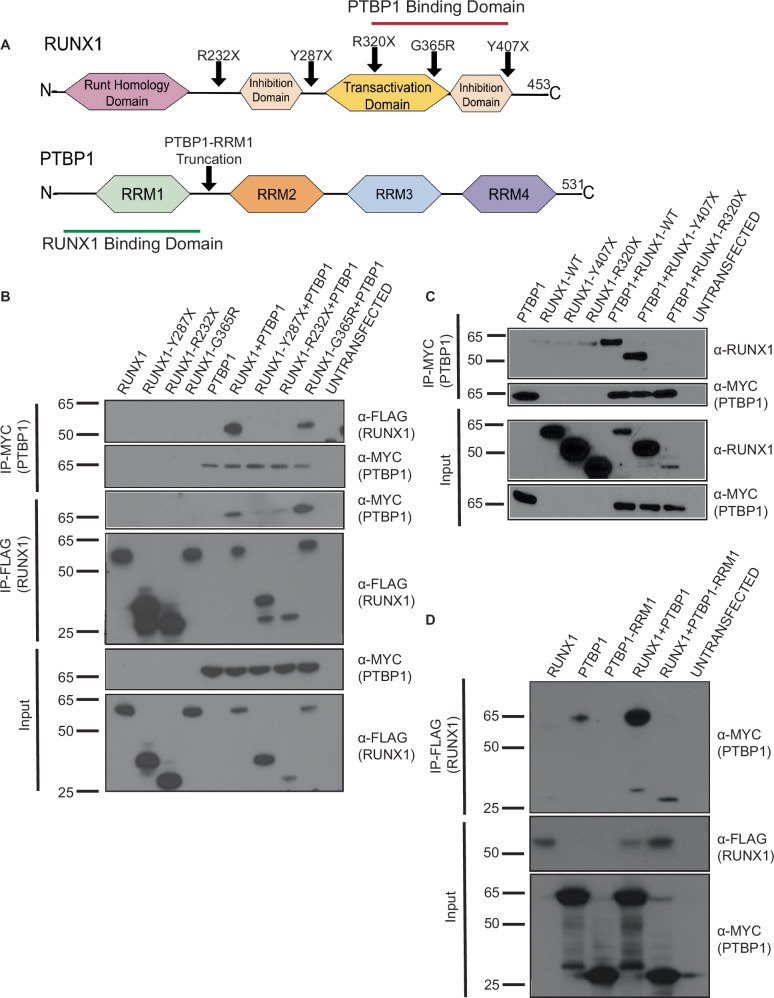 Figure 2
Figure 2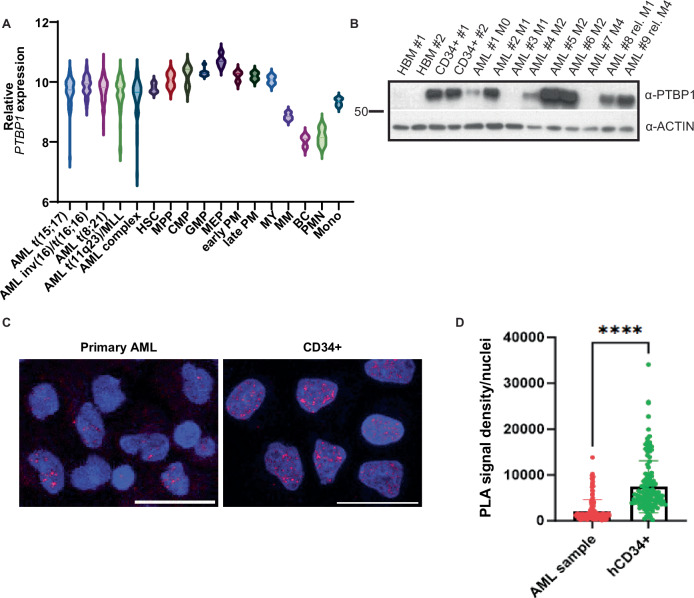 Figure 3
Figure 3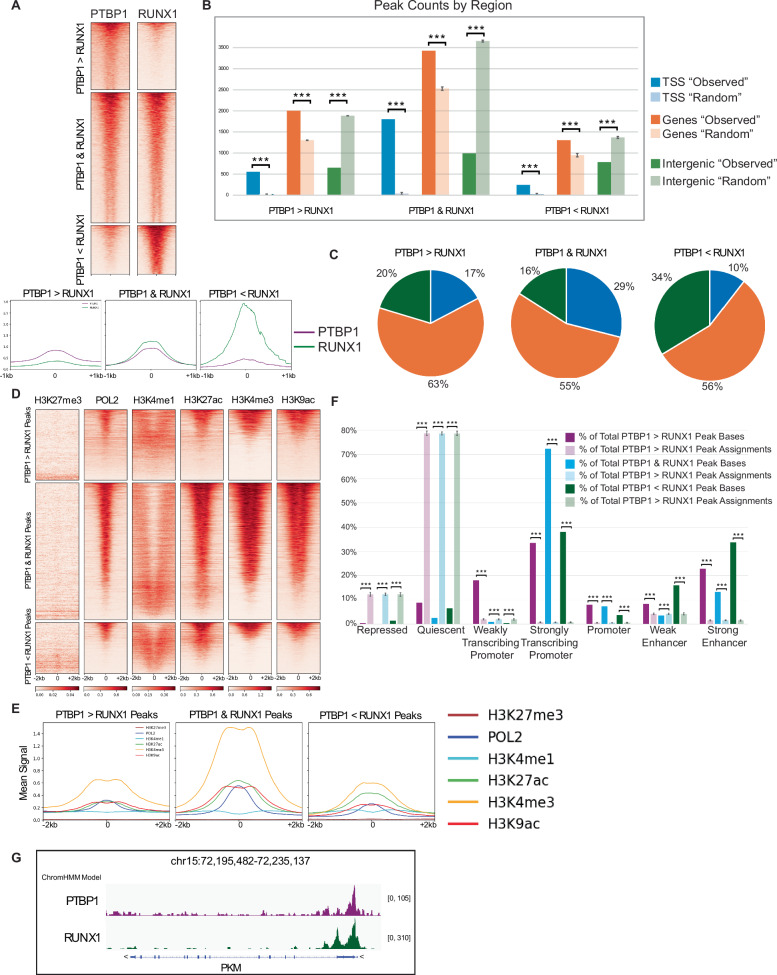 Figure 4
Figure 4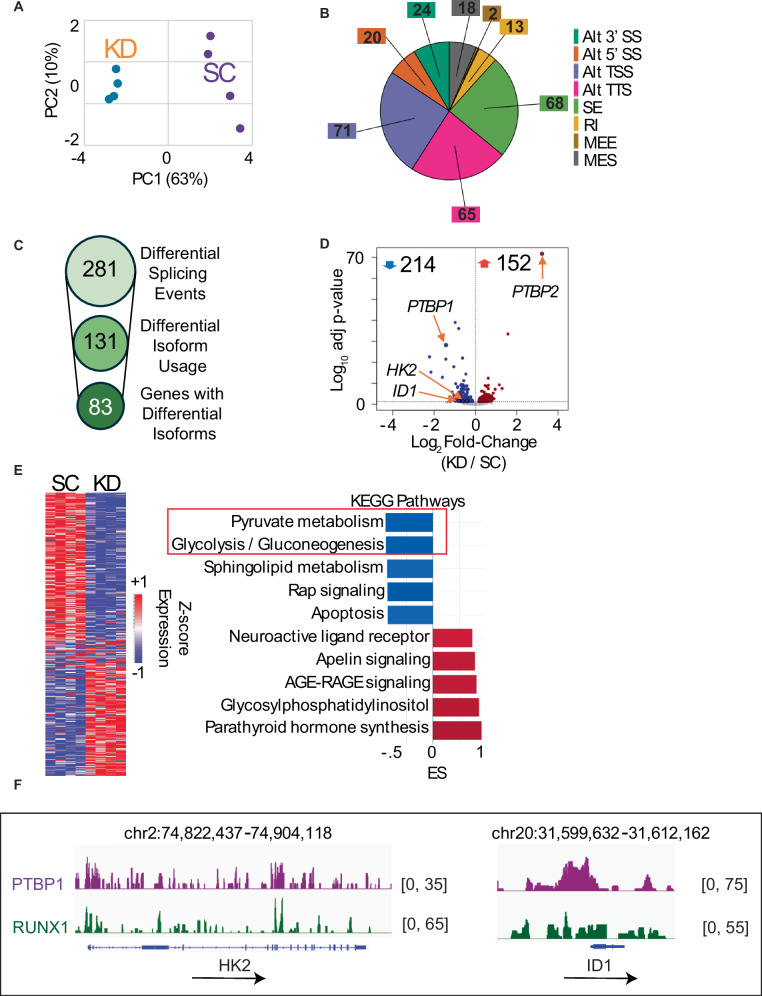 Figure 5
Figure 5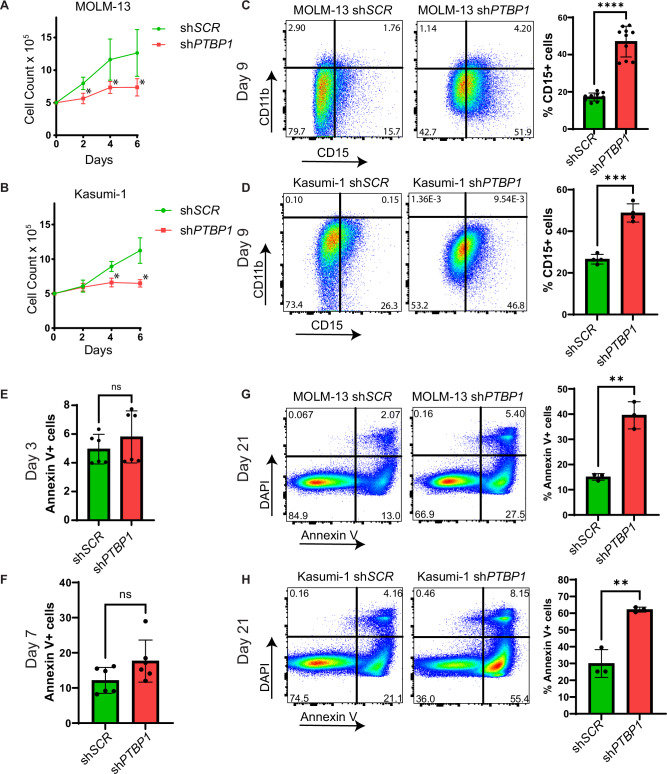 Figure 6
Figure 6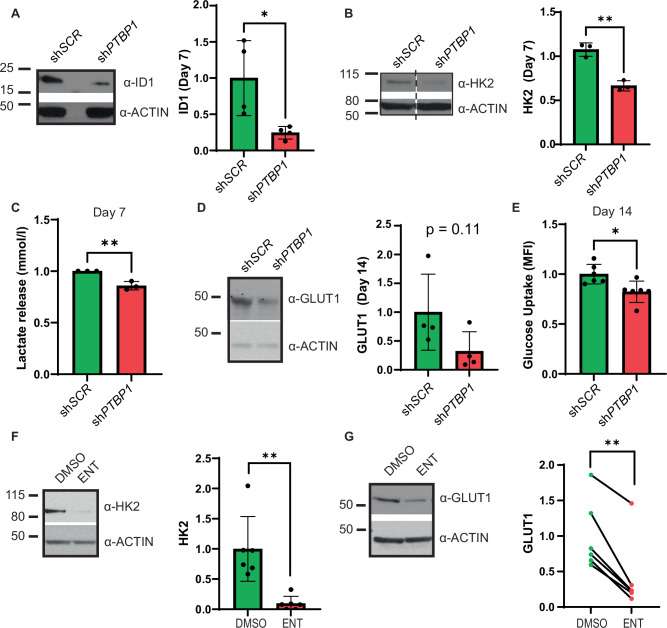 Figure 7
Figure 7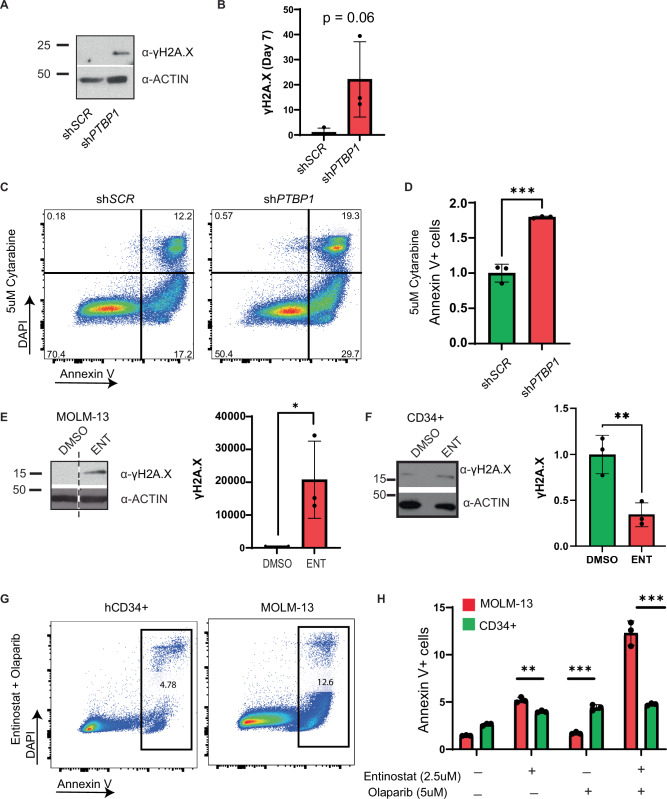 Figure 8
Figure 8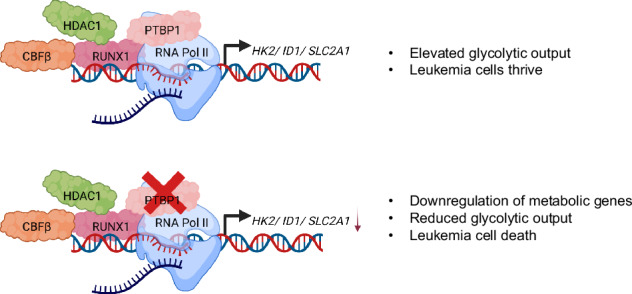 Figure 9
Figure 9- —https://doi.org/10.13039/100000054U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- —https://doi.org/10.13039/100000057U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHistone Deacetylase Inhibitors Research · Acute Myeloid Leukemia Research · Genomics and Chromatin Dynamics
Introduction
Leukemia is a cancer affecting blood and bone marrow cells, often caused by mutations involving key hematopoietic transcription factors, such as Runt-related transcription factor 1 (RUNX1, also known as AML1). Germline mutations in RUNX1 are associated with the leukemia predisposition syndrome Familial Platelet Disorder with Associated Myeloid Malignancies [1]. Somatic deletions, point mutations, and oncogenic fusion proteins involving RUNX1 (RUNX1::RUNX1T1, ETV6::RUNX1) and its obligate binding partner CBFβ (CBFβ::SMMHC) are common in Acute Myeloid Leukemia (AML) and Acute Lymphoblastic Leukemia (ALL), making RUNX1 one of the most frequently mutated genes in leukemia [2, 3].
RUNX1 is a member of the core binding factor (CBF) family of proteins, which are known to have roles in embryonic development, proliferation, differentiation, and cancer [4–6]. RUNX1 is known to interact with several other transcription factors and epigenetic modifiers, including Histone Deacetylase 1 (HDAC1) [7]. HDAC1 is best known for its role in transcriptional repression but can also mediate protein interactions and promote active transcription [8]. Our lab previously showed that HDAC1 is required for active transcription of RUNX1 target genes in a mouse model of CBFβ::SMMHC (CM) driven AML, making leukemia cells particularly sensitive to the HDAC1 inhibitor, entinostat [9]. This finding raised the possibility that HDAC1 activity may be required to mediate interactions between RUNX1 and other co-factors to promote target gene expression.
In this study, we identified a new RUNX1 binding partner, the splicing regulator, Polypyrimidine Tract Binding Protein 1 (PTBP1). As its name suggests, PTBP1 binds polypyrimidine-rich RNA. It is involved in pre-mRNA processing, RNA transport, and alternative splicing, and is expressed nearly ubiquitously [10–13]. PTBP1 has been implicated in the pathogenesis of solid tumors, but PTBP1’s role in the hematopoietic compartment has been less studied [14, 15]. Knockout mouse studies indicate that PTBP1 regulates hematopoietic stem cell maintenance, erythroid differentiation, and B-cell selection [16, 17]. In leukemia, PTBP1 has been linked to leukemia cell survival, metabolism, and progression [18–22]. However, the mechanisms behind how PTBP1 orchestrates these functions remain largely unknown. In this current study, we show that RUNX1 interacts with PTBP1 in both AML and ALL cells, and that this interaction requires HDAC1 activity. We also show that RUNX1 and PTBP1 co-localize at the promoters of actively transcribed genes, including key metabolic genes, and that loss of PTBP1 decreases expression of target genes, resulting in impaired glycolysis and cell death, linking RUNX1 to the control of metabolism in leukemia.
Methods
Mice
All animal experiments were approved by the University of Nebraska Medical Center IACUC in accordance with NIH guidelines. Cbfb^+/56M^, Mx1-Cre^+^ (CM^+^) or Cbfb^+/56M^, Mx1-Cre^+^, Gt(ROSA)26Sortm4(ACTB-tdTomato, -EGFP/Luo/J) (Rosa^26tdT/GFP^) (CM^+^, GFP^+^) were maintained on a mixed background (C57BL6/129S) [23, 24]. To induce leukemia, male and female mice (6-8 weeks) were treated with Polyinosine-polycytidylic acid (pI:pC) (InvivoGen, San Diego, CA, USA), intraperitoneally (250 µg/dose, three times a week, every other week for 6 weeks). Transplantations to expand leukemia cells were performed by injecting 1 × 10^5^ cells retro-orbitally into sub-lethally irradiated congenic mice, as previously described [9, 25]. At least 3 independent samples per condition were used to control for mouse-to-mouse variation. Samples were assigned to PTBP1 high or PTBP1 low groups based on western blot analysis. The investigators were not blinded.
Cell culture
Cbfb^+/56M^, Mx1-Cre^+^ cells were cultured in StemSpan SFEM I (Stemcell Technologies, Vancouver, Canada), supplemented with 1% Lipid Mixture (Sigma-Aldrich, St. Louis, MO, USA), 1% penicillin/streptomycin/L-glutamine (PSL), recombinant murine IL-3 (10 ng/mL), IL-6 (10 ng/mL) and SCF (20 ng/mL) (Gibco, Grand Island, NY, USA). Entinostat was procured from Cayman Chemical (Ann Arbor, MI). ME-1 cells were a kind gift from P. Liu, NGHRI, NIH, and authenticated by karyotype analysis. Authenticated Kasumi-1, MV-4-11, REH, MOLT-4, and Jurkat cells were obtained from ATCC (Manassa, VA, USA). Authenticated SEM and MOLM-13 cells were obtained from DSMZ (Leibniz Institute, Germany). All cell lines were reauthenticated by karyotype or STR analysis every 2 years and tested for mycoplasma contamination monthly. Deidentified, mobilized CD34+ cells from healthy donors were obtained from Fred Hutchinson Cancer Center Co-operative Center for Excellence in Hematology. Deidentified patient AML cells were obtained from the Stem Cell and Xenograft Core at the University of Pennsylvania Perelman School of Medicine and Cureline Translation CRO and cultured as previously described [26]. All human samples were obtained after informed consent under an approved protocol by each institution’s IRB committee.
Colony forming assay
Colony forming assays were performed using MethoCult GF M3434 (StemCell Technologies). CM^+^, GFP^+^ cells were sorted to isolate leukemic cells. 100,000 CM^+^, GFP^+^ cells were plated in 500 µl of MethoCult/well of a 24-well plate, in triplicate, and counted 10 days later.
Plasmid constructs
The Myc tag WT PTBP1 construct was purchased from Addgene (Plasmid# 23024). The FLAG-RUNX1 WT, FLAG-RUNX1-R232, FLAG-RUNX1-Y287, and FLAG-RUNX1-G365R constructs were a kind gift from J. Yang, St. Jude’s [27]. The WT-RUNX1 and RUNX1-Y407 constructs were generously shared by Paul Liu at NIH [28, 29]. The RUNX1-R320 construct was a kind gift from Dong-Er Zhang at the University of California, San Diego [30].
Site directed mutagenesis
Myc tag WT PTB (Addgene) was mutated to Myc-PTB-RRM1 (Y193) using the Phusion Hot Start II High-Fidelity PCR Master Mix (Thermo, Waltham, MA, USA). Primer sequences used to generate mutant constructs are available upon request.
Lentiviral mediated shRNA knockdown
HEK293T cells were co-transfected with second-generation lentiviral plasmids, pPax2 and MD2.G (5 µg each), and 10 µg of the PTBP1 or Scrambled (SCR) shRNA plasmid to generate lentivirus [22]. The PTBP1 knockdown and control plasmids, which express the shRNAs and the ΔLNGFR receptor under the control of a doxycycline-inducible promoter, were modified by GenScript (Piscataway, New Jersey, USA), replacing puromycin resistance gene for mCherry. Virus was concentrated using Amicon® Ultra-15 Centrifugal Filter Units (MilliporeSigma, Burlington, MA, USA). Transduction protocol for leukemia cells is available on request.
Immunoprecipitation and Western blot
Protein overexpression studies were carried out in HEK293T cells using Lipofectamine-2000 or Lipofectamine-3000 (Thermo). Protein lysates were incubated with primary antibodies followed by Protein A/G Dynabeads (Thermo) or with anti-FLAG M2 magnetic beads (Sigma-Aldrich). Additional details on antibodies are provided in Supplementary Table 1.
Mass spectrometry
Mouse CM^+^ cells were incubated with entinostat or DMSO for 24 h, and nuclear lysates were prepared and immunoprecipitated with an anti-RUNX1 antibody (Abcam, Cambridge, UK, Supplementary Table 1) and analyzed by the UNMC Mass Spectrometry Core Facility. Quantitative data analysis was performed using Progenesis QI Proteomics 4.2 (Nonlinear Dynamics). Statistical analysis was performed using ANOVA with the Benjamini-Hochberg (BH) correction for multiple testing caused false discovery rate. The adjusted p ≤ 0.05 was considered significant. Mass spectrometry proteomics data has been deposited to the ProteomeXchange Consortium via the PRIDE partner repository and is available with identifier PXD068027.
Confocal microscopy
Lineage depletion was performed using the EasySep™ Mouse Hematopoietic Progenitor Cell Isolation Kit (StemCell Technologies). Proximity Ligation Assay (PLA) was performed using Sigma-Aldrich’s Duolink PLA kit. Slides for both IF and PLA were imaged on the AiryScan 800 (63x magnification) at the UNMC Advanced Microscopy Facility. Quantification of PLA, puncta per field, was carried out using the Image J plug-in, Andy’s Algorithms [31]. Quantification of the PLA per nuclei was performed using ImageJ and processed using Z-projection with max intensity. DAPI signal was used to outline individual nuclei, and the PLA density within each nuclei was determined.
Flow cytometry and cell sorting
Cells were stained with Annexin V, DAPI, CD15, CD11b, and CD271 (ΔLNGFR) (BD Biosciences, Franklin Lakes, NJ, USA, and Biolegend, San Diego, CA, USA, Supplementary Table 1) and analyzed on a BD LSRII. Cell sorting was performed on FACSAria I or FACSAria II (BDBiosciences) or BigFoot (ThermoFisher Scientific). Data was analyzed in FlowJo v.10.0.8 (FlowJo, LLC, Ashland, OR, USA).
Cleavage under targets and tagmentation (CUT&Tag)
CUT&Tag was performed on MOLM 13 cells as previously described [32]. Primary antibodies were used at the following dilutions: PTBP1 (Thermo): 1:50; RUNX1 (Abcam): 1:20; rabbit/mouse IgG (CST, Danvers, MA): 1:100, and H3K27me3 (CST): 1:100. For secondary antibodies (1:100), we used guinea pig anti-rabbit antibody (Antibodies Online, Pottstown, PA, USA) and rabbit anti-mouse antibody (Abcam). Refer to Supplementary Table 1 for details. Library quality and concentrations were determined using the D1000 TapeStation system (Agilent). Libraries were sequenced in 150-bp paired-end mode on the Illumina NextSeq 550 (UNMC Genomics core) or NovaSeqX 10B (Novogene) platforms, and data was analyzed as previously described [33]. Adapters were clipped and paired-end reads were mapped to UCSC Hg38 using Bowtie2 [34] with parameters: --end-to-end --very-sensitive --no-mixed --no-discordant -q --phred33 -I 10 -X 700. Spike-in reads were mapped to E. coli K-12 genome with parameters: --end-to-end --very-sensitive --no-overlap --no-dovetail --no-unal --no-mixed --no-discordant -q --phred33 -I 10 -X 700. Continuous-valued data tracks (bedGraph and bigWig) were generated using genomecov in bedtools v2.30.0 (-bg option) and calibrated using total number of spike-in reads. Aligned reads were filtered for mapq>30, deduplicated, underwent peak identification using MACS3 [35] q-value threshold 10^−7^, and differential peaks were identified using MAnorm [36]. CUT&Tag data has been deposited at NCBI Geo and is available with identifier GSE308248.
Long-read RNA sequencing
RNA extraction was carried out using Qiagen’s RNeasy Mini Kit (Qiagen, Germantown, Maryland) following manufacturer’s instructions. RNA was isolated from PTBP1 knockdown and control MOLM-13 cells after 9 days of doxycycline treatment. Long-read RNA sequencing was performed by CD Genomics, Shirley, NY, using cDNA libraries prepared using the ONT SQK-PCS109 + SQK-PBK004 kits and sequenced on the PromethION platform. Reads were mapped to the Hg38 genome build using minimap2 [37], and filtered with a long-read mapq>50. Differentially expressed genes were identified using Stringtie [38] to generate transcript counts, followed by DESeq2 with an adj. p value <0.05. Differential isoform usage was performed by IsoformSwitchAnalyzeR [39] with a q-value >0.05. Functional categorization and overrepresentation analysis were performed using EnrichR [40]. RNA sequencing data has been deposited at NCBI Geo and is available with identifier GSE308249.
Gene expression data
Publicly available PTBP1 gene expression data was accessed from “Normal hematopoiesis with AMLs” data set in BloodSpot 3.0, a specialized gene and protein expression database of normal and malignant hematopoietic cells [41].
Metabolic assays
Lactate release and glucose uptake assays were performed using the Lactate Assay kit and the Glucose Uptake Assay Kit–Green (Dojindo, Rockville, MD, USA).
Statistics
Data was analyzed using either the two-sided Student’s t-test, ANOVA with Turkey post-hoc (for data with normal variation), or Wilcoxon-Rank Sum (for data with non-normal distribution) as appropriate, using GraphPad Prism 10.0.2. Exact N numbers are indicated in the figure legends. Data are reported as mean ± standard deviation. P values ≤ 0.05 were considered significant.
Results
PTBP1 binds RUNX1 in a HDAC1 dependent manner in leukemia
Work from our lab showed that HDAC1 binds CBFβ::SMMHC (CM) and is required for active transcription of RUNX1/CM target genes [9]. Canonically, HDAC1 is associated with transcriptional repression, but can also regulate the formation of protein complexes that promote gene expression [8]. We performed mass spectrometry to determine if HDAC1 activity influences the recruitment of proteins to the RUNX1 complex in leukemia cells. Five independent mouse CM^+^ leukemia samples were treated with 1 µM entinostat, an HDAC1 inhibitor, or DMSO for 24 h, nuclear lysates prepared, RUNX1 immunoprecipitated, and the precipitate analyzed by mass spectrometry (Fig. 1A). After filtering for known non-specific contaminants, we identified 809 peptides, that mapped to 284 different proteins, 80 of which were identified with at least 3 unique peptides. From this list, one protein showed a statistically significant difference between control and entinostat treated samples: Polypyrimidine Tract Binding Protein 1 (PTBP1) (Fig. 1B). We confirmed the interaction by showing higher co-immunoprecipitation of PTBP1 with RUNX1, compared to IgG, in 3 independent mouse CM^+^ leukemia samples (Fig. 1C). We also performed immunoprecipitations/westerns (IP/WB) using the human CM expressing AML cell line, ME-1, and found that RUNX1 interacts with PTBP1 in human leukemia cells, as well. To test if this interaction is unique to cells expressing the CM fusion gene, we performed IP/WB using a panel of human AML and ALL cell lines. We observed co-immunoprecipitation of PTBP1 and RUNX1 in all the cell lines tested (Fig. 1D, E), indicating that the interaction between PTBP1 and RUNX1 is present in both myeloid and lymphoblastic leukemia cells.Fig. 1PTBP1 binds RUNX1 in a HDAC1-dependent manner.A Schematic of experimental workflow. B Diagram showing filtering of peptides from mass spectrometry. C Representative western blot (WB) of lysates isolated from CM^+^ cells, D human AML cell lines, or (E) human ALL cell lines immunoprecipitated with anti-RUNX1 or anti-IgG antibodies and blotted for PTBP1. F Confocal images of Proximity Ligation Assays (PLA) performed in MOLM-13 cells using the indicated antibodies. G Confocal images of PLAs in MOLM-13 cells treated with vehicle or 10 µM entinostat for 48 h. H Graph quantifying PLA signal per field (minimum of 50 cells) from cells treated as in (G). N = 5 for mass spectrometry. N = 3 for IP, WBs, and PLA. * = p < 0.05. **** = p < 0.0001. Scale bar = 20 µm.
Both PTBP1 and RUNX1 can reside in the nucleus and the cytoplasm. To determine in which cellular compartment the proteins are interacting, we performed proximity ligation assays (PLA). We observed very few PLA puncta in RUNX1 or PTBP1 only antibody controls, but observed multiple puncta in the nuclei of cells with both PTBP1 and RUNX1 antibodies (Fig. 1F). To confirm that HDAC1 activity regulates the interaction between RUNX1 and PTBP1, we performed PLA on MOLM-13 and REH cells treated for 48 h with vehicle or entinostat. We found that in both cell lines (Fig. 1G, H and Supplementary Fig. 1A, B) there were significantly fewer PLA puncta in cells treated with entinostat, consistent with our mass spectrometry results. These findings indicate that RUNX1 and PTBP1 interact in the nucleus of both AML and ALL cells, and this interaction requires HDAC1 activity.
The N-terminus of PTBP1 and the C-terminus of RUNX1 are required to interact
To ascertain protein domains responsible for the interaction between RUNX1 and PTBP1, we used expression constructs with full length, wild type (WT) FLAG tagged RUNX1, and a series of point and deletion mutations found in patients with Familial Platelet Disorder with Associated Myeloid Malignancy (FPDMM) (Fig. 2A) [28–30, 42]. We observed co-IP of Myc-PTBP1 with WT FLAG-RUNX1, FLAG-RUNX1-G365R and FLAG-RUNX1-Y407X, but not with FLAG-RUNX1-R232X, FLAG-RUNX1-Y287X, and RUNX1-R320X (Fig. 2B, C). Even with a three-fold excess of the RUNX1-R320X plasmid, we did not observe co-IP with PTBP1 (Supplementary Fig. 1C). This implies that PTBP1 binds between amino acids 320 and 407 of RUNX1. To test which region of PTBP1 is required to bind RUNX1, we generated a PTBP1 truncation mutant that contains only the N-terminal region and the first RNA recognition motif (RRM), Myc-PTBP1-RRM1 (Fig. 2A). We observed co-IP of WT Flag-RUNX1 with full length Myc-PTBP1 and Myc-PTBP1-RRM1, implying that the N-terminus of PTBP1 binds RUNX1 (Fig. 2D). Collectively, these results indicate that the N-terminus of PTBP1 and the C-terminus of RUNX1 are required for the interaction between RUNX1 and PTBP1.Fig. 2. The N-terminus of PTBP1 binds the C-terminus of RUNX1.A Schematic representation of RUNX1 and PTBP1 proteins showing binding domains, and the relevant mutations indicated with arrows. B–D Representative western blots from HEK293T cells transfected with expression constructs for the indicated proteins, immunoprecipitated (IP’d) and probed with the indicated antibodies. N = 3.
The PTBP1/RUNX1 interaction occurs in healthy hematopoietic stem and early progenitor cells
RUNX1 is known to have roles in both leukemia and healthy hematopoietic stem and progenitor cells (HSPCs). To address whether RUNX1 binds PTBP1 in these cells, we compared the expression of PTBP1 in leukemia cells and HSPCs. Using publicly available gene expression data, we found that PTBP1 is expressed at similar levels in leukemia cells and healthy HSPCs but decreases with differentiation (Fig.3A). To examine PTBP1 protein expression, we performed WBs with bone marrow (HBM) and CD34+ cells from healthy volunteers, and primary AML patient samples. We found that PTBP1 is undetectable in HBM but highly expressed in CD34+ cells. PTBP1 was detectable in most patient samples, but its expression levels showed considerable variation, with no apparent correlation with French-American-British (FAB) classification (Fig. 3B). To test if RUNX1 binds PTBP1 in healthy hematopoietic cells, we performed PLA on mouse lineage depleted (lin-) bone marrow cells and human CD34+ cells. Primary mouse CM^+^ cells and patient AML cells were included for comparison. We observed significantly more PLA puncta in mouse CM^+^ leukemia compared to wild type lin- bone marrow cells (Supplementary Fig. 2A, B). Human CD34+ showed significantly higher levels of PLA puncta compared to primary human AML cells (Fig. 3C, D). These results indicate that RUNX1 and PTBP1 likely interact in the nuclei of healthy HSPCs.Fig. 3. The PTBP1-RUNX1 interaction occurs in healthy hematopoietic stem and early progenitor cells.A Violin plot depicting relative PTBP1 expression in human hematopoietic populations and AML subtypes (HSC- Hematopoietic Stem Cell, MPP- Multipotent Progenitor, CMP- Common Myeloid Progenitor, GMP- Granulocyte-Monocyte Progenitor, MEP- Megakaryocyte-Erythrocyte Progenitor, early PM- Early Promyelocyte, late PM- Late Promyelocyte, MY- Myelocyte, MM- Metamyelocytes, BC- Band cell, PMN- Polymorphonuclear cells, Mono- Monocytes). B Western blot (WB) of PTBP1 protein in healthy bone marrow (HBM), sorted CD34^+^ cells from healthy individuals, and primary diagnosis and relapse (rel) AML samples of the indicated FAB stage. C Representative confocal images and (D) graph of PLA intensity/nuclei (minimum of 50 cells) in human primary AML and healthy human CD34+ cells. N = 3. **** = p < 0.0001. Scale bar −20 µm.
Previous work in genetic mouse models indicates a role for PTBP1 in healthy hematopoietic stem cells, raising the possibility that PTBP1 could also regulate leukemia stem cell (LSC) activity [16]. For this, we used mouse CM^+^ leukemia cells expressing GFP from the Rosa26 locus (CM^+^, GFP^+^), and took advantage of the natural variability in PTBP1 expression levels in primary leukemia cells (Supplementary Fig. 2C). CM^+^, GFP^+^ leukemia cells were sorted for GFP, and WBs were used to identify mouse leukemia samples with PTBP1 expression above (PTBP1 high) and below (PTBP1 low) the median expression level (Supplementary Fig. 2D). Equal numbers of cells from 3 independent PTBP1 high and PTBP1 low samples were plated for colony formation assay. We observe significantly more colonies in samples expressing high levels of PTBP1, implying that increased PTBP1 expression is associated with increased LSC activity (Supplementary Fig. 2E, F). We next asked if the PTBP1/RUNX1 interaction is enriched in the LSC compartment. We sorted CM^+^, GFP^+^ leukemia cells for KIT and CSF2RB, markers we previously showed distinguish between these two populations in this mouse model, and performed PLA for RUNX1 and PTPB1 [43]. We observed no difference in the PLA puncta per nuclei between the LSC and non-LSC cells, implying that either the RUNX1/PTBP1 interaction has different activities in the different populations, or that regulation of LSC-activity by PTBP1 is RUNX1-independent (Supplementary Fig. 2G, H).
PTBP1 co-localizes with RUNX1 at promoter regions
To determine if PTBP1 co-localizes with RUNX1 on chromatin, we performed CUT&Tag (Cleavage Under Targets and Tagmentation) in MOLM-13 cells using antibodies against, RUNX1, PTPB1, and the appropriate IgG controls. We observed extensive overlap between PTBP1 and RUNX1 peaks, with RUNX1 signal at nearly all PTBP1 peaks and vice-versa (Fig. 4A). We categorized peaks into three groups: the largest category being those with equivalent binding of both PTBP1 and RUNX1 (PTBP1 & RUNX1), with fewer regions showing either stronger PTBP1 (PTBP1 > RUNX1) or stronger RUNX1 (PTBP1 < RUNX1). To identify the types of genomic regions bound by PTBP1 and RUNX1, we compared enrichment of signal in peaks at annotated Transcription Start Site (TSS), genes, and intergenic regions. We found that for all three categories of peaks, PTBP1 and RUNX1, were significantly enriched at TSS and gene bodies compared to Monte Carlo permutations (Fig. 4B, C). To test whether PTBP1 and RUNX1 binding is associated with active transcription, we compared our CUT&Tag data to publicly available RNA Polymerase II and histone modification data from MOLM-13 cells [44]. Regions bound by both PTBP1 and RUNX1 show strong enrichment of RNA Pol II and histone modifications associated with active transcription (H3K9ac, K3K27ac, H3K4me1, and H3K4me3) but not marks of transcriptional repression (H3K27me3) (Fig. 4D, E and Supplementary Fig. 3A–C). Further characterization of chromatin states using ChromHMM [45] revealed that PTBP1 and RUNX1 together are significantly enriched at strongly transcribing promoters (Fig. 4F, G and Supplementary Fig. 3D).Fig. 4PTBP1 co-localizes with RUNX1 at promoter regions.A Heatmaps and mean signal intensities from PTBP1 and RUNX1 CUT&Tag, centered on the peak, with 0 indicating the midpoint (±1 kb) grouped into regions with PTBP1 high (PTPB1 > RUNX1), PTBP1 and RUNX1 high (PTBP1 & RUNX1), and RUNX1 high (PTBP1 < RUNX1). Peaks were called using MACS3, and the three groups of bound regions defined using MAnorm. Heatmaps were sorted by decreasing signal intensity. B Location of peaks relative to annotated genomic loci; lightly colored bars display results of Monte Carlo permutations (n = 1000), demonstrating likelihood against random relocation of peaks. TSS- Transcription Start Site. C Pie chart representing occupancy of either RUNX1, or PTBP1, or both at annotated genomic loci. D Heatmap and E mean signal intensities of RNA Polymerase 2 (POL2) and histone modifications, specifically H3K27me3, H3K4me1, H3K27ac, H3K4me3, H3K9ac centered on the midpoint of the peak (±2 kb). F Percent distribution of peaks over ChromHMM-generated states; lightly colored bars display results of Monte Carlo permutations (n = 1000), demonstrating likelihood against random relocation of peaks. G IGV snapshot showing high PTBP1 & high RUNX1 signal at PKM gene TSS. *** = p ≤ 0.001. Both CUT&Tag and CUT&RUN were performed in duplicate.
Loss of PTBP1 induces widespread changes in splicing and gene expression
The co-localization of PTBP1 with RUNX1 at active promoters raises the possibility that PTBP1 has a role in the expression and splicing of target genes. To test this possibility, we used a doxycycline (dox) inducible shRNA against PTBP1 (shPTBP1) or a scrambled shRNA (shSCR) [46]. These lentiviral constructs constitutively express mCherry and express ΔLNGFR along with the shRNA after dox treatment (Supplementary Fig. 4A). After confirming efficient knockdown (KD) of PTBP1 (Supplementary Fig. 4B), we performed long-read RNA sequencing in control and PTBP1 KD MOLM-13 cells. Principal component analysis showed that the shPTBP1 (KD) samples cluster together, distinct from the control shSCR (SC) samples (Fig. 5A). We also found that loss of PTBP1 induced changes in splicing, with the most common changes involving the use of alternative transcriptional start sites (Alt TSS) (25%), alternative termination sites (Alt TTS) (23%), and skipped exons (24%) (Fig. 5B), with 131 significant differential isoforms associated with 83 genes (Fig. 5C). Analysis of our CUT&Tag data for the isoform switched genes shows this gene set is enriched for RUNX1, PTBP1, and RNA Pol II binding, as well as histone modifications associated with active transcription, at the TSS (Supplementary Fig. 5A, B). Comparing the list of isoform switched genes to publicly available PTBP1 eCLIP data, we found that the majority of these transcripts (77 of 83) are bound by PTBP1, indicating they are PTBP1 splicing targets (Supplementary Fig. 5C–E) [47].Fig. 5. Loss of PTBP1 induces widespread changes in splicing and gene expression.A Principal component (PC) analysis of MOLM-13 shSCR (SC) and shPTBP1 (KD) cells from long-read RNA sequencing data. B Pie chart revealing altered splicing events in response to PTBP1 KD in MOLM-13s (Alt 3’ SS – Alternative 3’ Splice Site, Alt 5’ SS – Alternate 5’ Splice Site, Alt TSS – Alternate Transcription Start Site, Alt TTS – Alternate Transcription Termination Site, SE – Skipped Exon, RI – Retained Intron, MEE – Mutually Exclusive Exon, MES – Multiple Exon Skipping). C Analysis of the splicing events in (B) revealing genes with differential isoform expression in response to PTBP1 KD in MOLM-13s. D Volcano plot showing differentially expressed genes in response to PTBP1 KD in MOLM-13s. Highlighted genes include Inhibitor of Differentiation 1 (ID1), Hexokinase-2 (HK2), PTBP1, and PTBP2. E Analysis of differentially expressed genes by EnrichR showing top KEGG pathways affected after PTBP1 KD. F Integrated Genomics Viewer (IGV) tracks depicting the binding of RUNX1 and PTBP1 at HK2 and ID1 gene loci. N = 4 for long-read RNA sequencing.
In addition to altered isoform usage, PTBP1 KD resulted in the downregulation of 214 and upregulation of 152 genes, with the most highly upregulated gene being the fellow PTBP family member, PTBP2 (Fig. 5D). Among the downregulated genes, we identified key metabolic genes including Hexokinase-2 (HK2), Inhibitor of DNA Binding 1 (also known as Inhibitor of Differentiation 1) (ID1) and Solute Carrier 2A1 (SLC2A1), as well as PTBP1. (Fig. 5D). Functional categorization and overrepresentation analysis of Kegg pathways revealed that the genes downregulated with PTBP1 KD are associated with metabolic pathways critical for leukemia cell growth, including pyruvate metabolism and glycolysis (Fig. 5E). Our CUT&Tag data identified RUNX1 and PTBP1 co-localization at several deregulated genes associated with these pathways, including ID1, HK2 (Fig. 5F and Supplementary Fig. 6A), and SLC2A1, which encodes Glucose Transporter 1 (GLUT1) (Supplementary Fig. 6B).
PTBP1 promotes leukemia cell survival by regulating expression of metabolic genes
Downregulation of genes associated with glycolysis and pyruvate metabolism with PTPB1 KD implies a role for PTBP1 in regulating leukemia cell growth. To test this, we treated shPTBP1 and control shSCR cells with dox for 2 weeks and performed growth assays. We observed a significant reduction in cell numbers in PTBP1 KD cells (shPTBP1) compared to control cells (shSCR) (Fig. 6A, B). To test if the decreased growth is associated with differentiation to a post-mitotic, mature myeloid phenotype, we examined the expression CD11b and CD15. We found that 9 days of dox treatment induced a statistically significant increase in CD15 expression in shPTBP1 cells, as compared to shSCR cells (Fig. 6C, D). To test if cell death also contributes to the growth defect in PTBP1 KD cells, we used flow cytometry. At days 3 and 7 of PTBP1 KD, we observed no significant effect on viability in MOLM-13s (Fig. 6E, F). In contrast, at day 21, there was significantly increased Annexin V+ staining in shPTBP1 cells indicating that prolonged knockdown of PTBP1 induces apoptosis (Fig. 6G, H).Fig. 6PTBP1 promote leukemia cell survival by regulating expression of metabolic genes.A Growth curve of MOLM-13 (N = 3) and B Kasumi-1 (N = 3) shSCR and shPTBP1 cells. C Representative plots and quantification from flow cytometry analysis of CD15 and CD11b of MOLM-13 (N = 10) and D Kasumi-1 (N = 4) shSCR and shPTBP1 cells. E Representative plots and quantification of Annexin V+ cells in MOLM-13 shSCR and shPTBP1 cells on day 3 (N = 6) and F day 7 (N = 6) of doxycycline induction. G Representative plots and quantification of flow cytometry analysis of Annexin V and DAPI on day 21 of doxycycline induction in MOLM-13 (N = 3) and H Kasumi-1 (N = 3) shSCR and shPTBP1 cells. % = percentage. For each experiment “N” refers to separate doxycycline inductions * = p < 0.05, ** = p < 0.01. **** = p < 0.0001.
To determine if growth defects caused by PTBP1 KD are accompanied by metabolic dysfunction, we first performed western blot analysis of metabolism related proteins found deregulated in our RNA-seq analysis. We found that ID1 and HK2 showed significantly decreased expression after 7 days of KD (Fig. 7A, B). At this time point, PTBP1 KD cells showed significantly lower lactate release into the media, consistent with decreased glycolysis (Fig. 7C). On day 14 of PTBP1 KD, we also observed a trend towards decreased GLUT1 expression (Fig. 7D). Using a flow cytometry-based glucose uptake assay, we observed a significant reduction in glucose uptake specifically in viable, DAPI-, shPTBP1 cells at this time point (Fig. E). Collectively, these results indicate that PTBP1 regulates the expression of key metabolic genes required for efficient glycolysis and survival in leukemia cells.Fig. 7. Loss of PTBP1 causes metabolic defects in leukemia cells.Loss of PTBP1 causes metabolic defects in leukemia cells: A Representative WB and quantification of relative abundance of Inhibitor of DNA binding 1 (ID1) (N = 4) and B Hexokinase-2 (HK2) (N = 3) on day 7 of PTBP1 KD in MOLM-13s. Dashed line indicates non-adjacent lanes from the same gel. C Quantification of lactate from lactate release assays in MOLM-13 shSCR and shPTBP1 cells at day 7 of KD (N = 3). D Representative WB and quantification of relative abundance of Glucose transporter-1 (GLUT1) on day 14 of PTBP1 KD in MOLM-13s (N = 4). E Quantification of mean fluorescence intensity (MFI) from glucose uptake assays on day 14 of doxycycline treatment in MOLM-13 shSCR and shPTBP1 cells (N = 6). F Representative WB and quantification of relative abundance of HK2 (N = 6) and G GLUT1 (N = 6) in MOLM 13 cells treated with 5 µM Entinostat (ENT) or DMSO for 48 h. *p ≤ 0.05. ** = p < 0.01. For (A–E), “N” refers to separate doxycycline inductions. For (F and G), “N” refers to separate entinostat treatments.
To address whether the interaction with RUNX1 may contribute to PTBP1’s role in the expression of these genes, we treated MOLM-13 cells with 5 µM entinostat for 48 h to disrupt the RUNX1/PTBP1 interaction. We observed a significant decrease in HK2 and GLUT1 protein in entinostat treated cells as compared to control, consistent with a role for the RUNX1/PTBP1 interaction in promoting expression of their target genes (Fig. 7F, G). To determine whether disruption of the RUNX1/PTBP1 interaction differentially affected the LSC and non-LSC cells, we treated mouse CM^+^ leukemia cells with entinostat for 24 h, and examined glucose uptake in these populations. We found that entinostat led to a significant decrease in glucose uptake in both populations, implying that the RUNX1/PTBP1 interactions similarly promotes glycolysis in both LSCs and non-LSCs (Supplementary Fig. 7).
PTBP1 KD induces DNA damage and sensitizes leukemia cells to chemotherapy
Germline mutations in RUNX1 lacking the C-terminus, which would disrupt the interaction with PTBP1, have been reported in patients with Familial Platelet Disorder with associated Myeloid Malignancies. Interestingly, these mutations have also been shown to induce DNA damage via an unknown mechanism [48]. Additionally, PTBP1 has been implicated in the DNA damage response, raising the possibility that the disruption of the RUNX1/PTBP1 interaction may lead to the accumulation of DNA damage [49]. To test this possibility, we examined the expression of the DNA damage marker, phosphorylated γH2A.X, in PTBP1 KD and control cells. We observed elevated levels of phospho-γH2A.X in shPTBP1 cells (Fig. 8A, B). To test if the increased DNA damage sensitizes shPTBP1 cells to chemotherapy, we treated shSCR and shPTBP1 cells with 5 µM of cytarabine for 3 days. We observed a significant increase in the Annexin V+ population in shPTBP1 cells as compared to control, indicating that loss of PTBP1 sensitizes leukemia cells to chemotherapy (Fig. 8C, D). To determine if disruption of the RUNX1/PTBP1 interaction also leads to increased DNA damage, we examined phospho-γH2A.X expression in control and entinostat treated MOLM-13s. We observed a significant increase of phospho-γH2A.X in entinostat treated cells compared to the control (Fig. 8E). Interestingly, treatment with entinostat did not induce an increase in phospho-γH2A.X in healthy CD34+ cells (hCD34+), implying that disruption of the RUNX1/PTBP1 interaction induces double stranded breaks in leukemia cells, but not in healthy HSPCs (Fig. 8F). This result raises the possibility that loss of the RUNX1/PTBP1 interaction may make leukemia cells dependent on double strand break repair pathways. Therefore, we treated MOLM-13 and healthy CD34+ cells (CD34 + ) with entinostat and the FDA approved PARP inhibitor, olaparib, either individually or in combination. We observed a modest increase in Annexin V+ cells in entinostat treated MOLM-13s compared to hCD34+ cells. However, combination treatment of entinostat and olaparib caused a dramatic, statistically significant increase in apoptosis in MOLM-13s, but not in hCD34+ cells (Fig. 8G, H). This result implies that the combination of entinostat and olaparib may be effective treatment for AML patients.Fig. 8PTBP1 KD sensitizes leukemia cells to chemotherapy.A Representative WB and B quantification of (S139) phosphorylated ɣH2A.X levels after 7 days of PTBP1 KD in MOLM-13 cells (N = 3). C Representative flow cytometry analysis plots and D quantification of Annexin V and DAPI staining in MOLM-13 shSCR and shPTBP1 cells treated with 5 µM cytarabine for 72 h after 7 days of PTBP1 KD (N = 3). E Representative WB and quantification of (S139) phosphorylated ɣH2A.X levels after 48 h of entinostat treatment in MOLM-13 and F healthy CD34+ (hCD34+) cells (N = 3). G Representative flow cytometry analysis plots and H quantification of Annexin V and DAPI stained MOLM-13 AML cells and hCD34+ cells treated with entinostat and Olaparib (N = 3). *** = p < 0.001. For (A–D), “N” refers to separate doxycycline inductions. For (E–H), “N” refers to separate treatments.
Collectively, our findings indicate that RUNX1 and PTBP1 interact in an HDAC1 dependent manner at the promoters of key metabolic genes critical for leukemia cells survival and that disruption of this association induces double strand DNA breaks, sensitizing leukemia cells to inhibitors of the DNA repair machinery. This work supports a model in which recruitment of PTBP1 to RUNX1 target genes promotes their efficient expression and maintains robust glycolytic output, highlighting a previously unrecognized aspect of RUNX1’s central role in leukemia pathogenesis.
Discussion
HDAC1 is canonically considered a repressor of gene expression, but can also mediate protein interactions. Previous work from our lab showed that HDAC1 is required for active transcription of a subset of RUNX1 target genes in leukemia cells. To address this contradiction, we tested whether inhibition of HDAC1 alters the RUNX1 interactome. We found that PTBP1 binds RUNX1 in an HDAC1-dependent manner and is required for expression of genes bound by both factors, providing a potential mechanism for HDAC1’s non-histone role in gene regulation. In addition, we identified the regions in RUNX1 and PTBP1 required for this interaction. Interestingly, both domains include multiple lysine residues that could be targets of HDAC1 mediated deacetylation. In the case of PTBP1, lysines (K92 and K94) in the RUNX1 interaction domain have been shown to be acetylated, providing further support for this potential mechanism [50].
We also showed that both RUNX1 and PTBP1 exhibit a strong bias for binding the transcription start site of actively transcribed genes, including those involved in metabolism. Previous work has implicated PTBP1 in the regulation of glycolysis by altering the splicing of pyruvate kinase (PKM) in leukemia [18, 22]. In the current study, we identify additional metabolic genes dependent on PTBP1 for expression, including SLC2A1, which encodes the glucose transporter GLUT-1, Hexokinase 2 (HK2), an enzyme in the glycolysis pathway, and Inhibitor of DNA Binding 1 (ID1), an upstream regulator of glycolysis [51]. Interestingly, analysis of transcriptomic data from Rehn et al. indicates that these genes are not similarly downregulated in the HSPCs of Ptbp1 knockout mice, implying that control of metabolic genes by the RUNX1/PTBP1 complex may be unique to leukemia cells [16].
It is notable that the most upregulated gene in PTBP1 KD cells was its family member, PTBP2. This observation is consistent with work in neuronal cells showing that PTBP1 represses PTBP2 expression by promoting splicing of an alternative isoform containing a premature termination codon and triggering non-sense mediated decay of the PTBP2 transcript [52]. The high expression of PTBP2 also raises the possibility that it may be compensating for the loss of PTBP1. Indeed, PTBP1 and 2 have been shown to have both overlapping and distinct activities [53–55]. In addition, PTBP2 has been shown to regulate survival and proliferation in Chronic Myeloid Leukemia, making it likely that PTPB2 at least partially compensates for PTBP1 loss in AML cells [56].
We also found that loss of PTBP1, as well as disruption of the RUNX1/PTBP1 interaction using entinostat, induced decreased expression of these factors at the RNA and protein level and caused metabolic defects. It is notable that with PTBP1 KD, we largely observed decreased expression of these genes, rather than the presence of alternative or mis-spliced isoforms. As PTBP1 is not thought to have any transactivation activity on its own, the most likely explanation for this finding is that these transcripts were subject to nonsense-mediated decay. Another explanation is that PTBP1 promotes the stability of these transcripts, a PTBP1 function observed in other cancers [57]. Importantly, the changes in expression and the associated metabolic defects were observed prior to induction of apoptosis, implying that the decreased expression of the RUNX1/PTBP1 target genes is the cause, and not the consequence, of cell death in PTBP1 KD cells. Finally, we showed that disruption of the interaction induces DNA damage and that inhibiting the DNA damage response in these cells leads to increased apoptosis in AML cells compared to healthy HSCs. Our findings imply that treatment with HDAC1 inhibitors may sensitize leukemia cells to PARP inhibitors, a class of drugs not previously thought to be effective in treating AML.
Our study demonstrates a novel facet of RUNX1’s role in leukemia through its interaction with PTBP1 and the regulation of metabolism. Our findings lead us to propose the following mechanism: PTBP1, which does not have a DNA binding domain, binds RUNX1 and is recruited to the promoter regions to drive expression of genes essential for leukemia cell survival. As a result, loss of PTBP1 leads to decreased expression of these target genes, leading to reduced metabolic output and ultimately cell death (See Graphical Abstract). Collectively, our results provide new insights into the intricacies of co-transcriptional splicing in leukemia and hematopoiesis, further underscoring the central role RUNX1 plays in both these processes.
Supplementary information
Supplemental Material