Circadian rhythms and neuroendocrine dysregulation in ADHD: Therapeutic insights from omega-3 fatty acids
Ayesha Zafar Iqbal, Patricia Marest Suwindi, Sunny Yin-Shan Chen, Lindsay Liang-Tien Cho, Kuan-Pin Su, Jane Pei-Chen Chang

TL;DR
This paper explores how circadian rhythm disruptions in ADHD may be addressed with omega-3 fatty acids, which could help regulate sleep and brain function.
Contribution
The paper introduces omega-3 fatty acids as potential non-photic zeitgebers to modulate circadian dysregulation in ADHD.
Findings
Omega-3 fatty acids may influence melatonin synthesis and sleep-wake regulation in ADHD.
N-3 PUFAs could modulate circadian clock genes in the SCN and normalize HPA axis activity.
Supplementation with n-3 PUFAs may improve ADHD outcomes through circadian pathways, though clinical evidence is limited.
Abstract
Attention-deficit hyperactivity disorder (ADHD) is a common neurodevelopmental condition often accompanied by circadian rhythm disturbances, particularly delayed sleep phase. These involve suprachiasmatic nucleus (SCN) dysregulation, altered melatonin secretion, and hypothalamic–pituitary–adrenal (HPA) axis activity, which may be exacerbated by artificial light exposure. Genetic studies further implicate circadian mechanisms, linking ADHD with polymorphisms in clock genes such as PER and CLOCK. Nutraceuticals, particularly omega-3 polyunsaturated fatty acids (n-3 PUFAs), have been proposed as modulators of circadian rhythms. N-3 PUFAs are essential for brain health and may influence melatonin synthesis and sleep–wake regulation. Preclinical and clinical findings suggest that supplementation can improve cognitive and behavioral outcomes in ADHD, possibly through circadian pathways,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
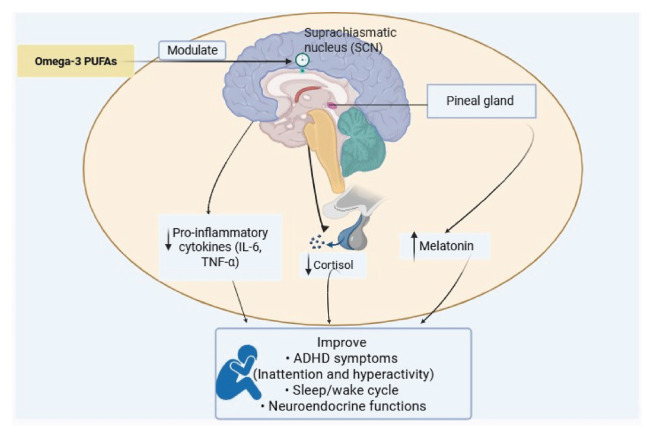 Figure 1
Figure 1- —MOST
- —National Science and Technology Council (NSTC), Taiwan
- —An-Nan Hospital, China Medical University (CMU), Tainan, Taiwan
- —Higher Education Sprout Project by the Ministry of Education (MOE), Taiwan
- —CMU, Taichung, Taiwan
- —CMU Hospital, Taichung, Taiwan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAttention Deficit Hyperactivity Disorder · Sleep and related disorders · Circadian rhythm and melatonin
Introduction
Attention-deficit hyperactivity disorder (ADHD) is one of the most common neurodevelopmental disorders that has been a burden worldwide and can potentially impact individuals and their families across the lifespan [1], with the common symptoms of inattention, hyperactivity, and impulsiveness [1]. The overall prevalence of ADHD in children and adolescents was 8.0% which is predicted to be twice as high in boys (10%) compared to girls (5%) [2]. Approximately 7.5%ADHDindividuals across all age groups had a formal sleep disorder diagnosis, and 47.5% are on prescribed sleep medications [3]. Among these disorders, delayed circadian rhythm is a common and significant feature in adults with childhood-onset ADHD. Studies indicate that up to 75% of these individuals exhibit delayed dim-light melatonin onset (DLMO), occurring about 1.5 h later than in individuals without ADHD, suggesting a significant alteration in circadian timing linked to the condition [4].
Furthermore, an epidemiological, genetic, etiological, and functional overlap exists between delayed circadian rhythms and ADHD symptoms across the lifespan [5]. In addition, the firstline treatment for ADHD is methylphenidate (MPH), a norepinephrine and dopamine reuptake inhibitor [6], which has been linked to adverse effects on sleep, including changes in total sleep time, sleep patterns, efficiency, and longer delay of sleep onset, potentially exacerbating sleep issues in individuals with ADHD [6]. However, these findings remain inconclusive due to conflicting results across studies and the limited availability of data. Factors such as the specific type of medication, dosage, and administration schedule warrant further investigation, as there is speculation that the pharmacological agents used may influence the chronobiology of the brain, thereby affecting sleep quality. Conversely, it is also plausible that improvements in daytime functioning resulting from medication use may subsequently lead to enhanced sleep quality during the night [7].
Circadian rhythms regulate physiological and behavioral processes on a 24-h cycle, though the actual human cycle often exceeds 24 h, requiring daily resetting [8]. This rhythm is controlled by a molecular clock involving feedback loops of core clock genes, including Circadian locomotor output cycles kaput (CLOCK), Brain and Muscle Arnt-like 1 (BMAL1), Period (PER), Cryptochrome (CRY), nuclear receptor subfamily 1 group D member 1 (REVERBα), and Retinoid-related orphan receptor alpha (RORαA). CLOCK and BMAL1 dimerize to initiate PER and CRY transcription, with the resulting PER-CRY complex inhibiting CLOCK/BMAL1 activity, creating a negative feedback loop. REV-ERB and ROR further regulate BMAL1 transcription, contributing to the precision of the circadian cycle. Post-translational modifications fine-tune protein stability and turnover. For example, casein kinase 1ε/δ (CK1ε/δ) phosphorylates PER and CRY to control nuclear entry and degradation, and F-box/LRR-repeat protein 3 (FBXL3) targets CRY for proteasomal degradation, thereby resetting the feedback loop [9]. CLOCK acetylates BMAL1 to facilitate CRY-mediated repression, whereas sirtuin 1 (SIRT1) deacetylates BMAL1 and PER2 to promote their turnover [10]. Together, these transcriptional and post-translational mechanisms ensure robust circadian rhythms, and their disruption leads to sleep and metabolic disorders.
While these molecular feedback loops constitute the circadian rhythms, their synchronization to the external environment is orchestrated by the suprachiasmatic nucleus (SCN), the central pacemaker located in the hypothalamus. The SCN synchronizes peripheral clocks mainly through light input from intrinsically photosensitive retinal ganglion cells via the retinohypothalamic tract (RHT) [11]. Although other oscillators exist outside the SCN, its role as the master pacemaker is underscored by evidence that lesions to the SCN disrupt a majority of physiological, endocrine, and behavioral rhythms [12]. Each SCN nucleus contains about 10,000 neurons and is positioned bilaterally to the third ventricle, just above the optic chiasm.
It is well-positioned to receive visual input for light-dark cycle entrainment via the RHT [13]. The melanopsin system, involving intrinsically photosensitive retinal ganglion cells, primarily mediates light's effects on the SCN [11]. These ganglion cell axons form the RHT, transmitting photic data to the SCN through a monosynaptic glutamatergic projection that includes the neuropeptide Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) [11]. Neuronal firing patterns in the SCN are crucial, as blocking SCN neuronal activity leads to behavioral arrhythmicity, which is restored upon resumption of SCN neuronal activity [14]. The SCN activity rises during the day and falls at night, following the same pattern as the clock genes and showing how the body internal clock controls SCN signals [15].
On the other hand, modern lifestyles, including irregular eating patterns, travel, caffeine intake, noise pollution, and excessive artificial light, can disrupt circadian rhythms, increasing the risk of cardiovascular diseases, cancer, neurodegenerative disorders, and ADHD [16].
Circadian rhythm disturbances in ADHD
2.1. Genetic associations of circadian rhythm disturbances and ADHD
Genome-wide association studies investigating such variants further support the involvement of disrupted molecular clock function in ADHD pathophysiology. These studies have increasingly highlighted the role of circadian CLOCK genes in ADHD, with particular attention to polymorphisms in the PER gene that influence circadian timing and behavioral regulation. A genome-wide association study investigating the impact of genetic polymorphisms on the circadian pathway in children with ADHD found a nominal association between the Period 1 (PER1) gene and ADHD risk [17]. However, no significant changes were observed in Period 2 (PER2) gene expression in cultured human dermal fibroblasts from patients with ADHD [18].
The T-allele of the rs1801260 single-nucleotide polymorphism (SNP) in the CLOCK gene has been associated with adult ADHD symptoms [19] and is also linked to evening partiality and delayed sleep timing; however, the results are inconsistent [20,21]. Although the exact functional impact of this SNP on CLOCK gene expression or activity is unclear, its location in the 3’ untranslated region suggests it may influence messenger ribonucleic acid (mRNA) stability, translation, and polyadenylation signalling [19]. In addition to circadian CLOCK gene polymorphisms, genetic variations in extracellular matrix degradation pathways have also been implicated in the vulnerability to major depressive disorder (MDD) [22]. Together, these data highlight converging molecular mechanisms ranging from circadian regulation to ECM remodelling that may underlie shared pathophysiology across neuropsychiatric disorders such as ADHD and depression.
Animal models with CLOCK gene knockouts, such as zebrafish PER1b and mouse PER1, display ADHD-like behaviors, including hyperactivity, impulsivity, and attention deficits, which provides further evidence of the connection between circadian dysfunction and ADHD. The circadian system is also believed to influence ADHD by regulating dopamine levels. A study found that zebrafish with a mutation in the PER1b gene exhibit behaviors resembling those of individuals with ADHD, such as hyperactivity, impulsivity, and attention deficits, along with lower dopamine levels. The circadian clock regulates dopamine synthesis and the development of dopaminergic neurons, and its disruption, as seen in PER1 knock-out mice, results in reduced dopamine levels and ADHD like hyperactive and inattentive behaviors, highlighting a link between circadian rhythm dysregulation and attention deficits [23].
2.2. Treatment for circadian rhythm disturbances in ADHD
Both non-pharmacological and pharmacological interventions are available to improve sleep in individuals with ADHD, targeting sleep disturbances associated with both the disorder itself and the side effects of ADHD medications. [24]. Non-pharmacological interventions include weighted blankets (a device made of weight-adding materials to cover the body evenly) [25], light therapy, acupuncture, exogenous melatonin, and other nutraceuticals. A metaanalysis showed that melatonin reduces sleep onset latency in children and adolescents with ADHD and comorbid insomnia [26]. Therefore, investigations into melatonin as a treatment for sleep disorders in ADHD patients have shown promise in improving total sleep time, shortening sleep onset delay, and enhancing emotional and behavioural regulation. However, the evidence remains limited and heterogeneous [27,28]. Moreover, nutraceuticals such as omega-3 polyunsaturated fatty acids (n-3 PUFAs) regulate neurological, inflammatory, metabolic, and cardiovascular processes through clock gene modulation, supporting their role as non-photic zeitgebers and potential therapeutics for circadian disruption-related pathologies [29]. N-3 supplementation has also shown benefits in ADHD and preventive effects against recurrent MDD, suggesting trans-diagnostic value [30]. The current work aims to provide an inclusive review of the prevailing literature on n-3 PUFAs in regulating circadian rhythms in individuals with ADHD.
Shared mechanisms between ADHD and circadian rhythm disorders
Several shared mechanisms, including the dysregulation of melatonin and hypothalamic–pituitary–adrenal (HPA) axis, have been proposed to link the pathophysiology of ADHD and circadian rhythm disorders with ADHD, further suggesting a complex interplay of biological factors in the disorder.
3.1. Melatonin dysregulation
Melatonin dysregulation may serve as a potential shared mechanism between ADHD and circadian rhythm disorders. As a key hormone modulating SCN and the circadian clock, it primarily influences the sleep-wake cycle. It is synthesized in pinealocytes from tryptophan, production peaks around 12–3 AM, and is controlled by sympathetic input from the cervical ganglion under SCN influence via GABAergic mechanisms [31]. It plays a crucial role in aligning circadian rhythms with the light-dark cycle, affecting sleep, hormone secretion, immune response, and daily activity patterns. Moreover, melatoninergic agonists have been used to treat non-24-h sleep-wake disorder in blind individuals [32].
Melatonin levels in individuals with ADHD have been shown to differ from those of controls, but with inconsistent results (Table 1). One study reported higher daytime and nighttime urinary melatonin metabolite (6-OH MS) levels in ADHD individuals compared to controls [33]. At the same time, another found no difference in the daily profile between controls and ADHD [34]. Additionally, a pilot study observed a lower 24-h melatonin rhythm in ADHD cases compared to controls [35]. However, some studies reported no significant differences in night-time melatonin levels [36,37]. Another study found overall higher melatonin concentrations in ADHD patients compared to controls [38]. A more consistent finding in ADHD is the delayed DLMO (circadian phase marker) and delayed sleep and wake times, particularly in individuals with sleep onset insomnia [39]. This delay is a characteristic of the delayed sleep phase syndrome (DSPS), suggesting that sleep onset insomnia is a circadian rhythm disorder comorbid with ADHD. Notably, individuals with the inattentive subtype of ADHD exhibit fewer symptoms of sleep-onset insomnia compared to other subtypes and tend to have longer and more stable sleep patterns [40]. This aligns with reports that subjects with the inattentive subtype of ADHD are sleepier during the day and sleep longer, potentially due to melatonin dysregulation [41]. However, not all studies report delayed sleep onset timing in ADHD, indicating variability in sleep disturbances within this population [42].
Various preclinical and clinical studies have explored the impact of melatonin on sleep and behavioral parameters, particularly in individuals with ADHD and related conditions. In a zebrafish model, auriculasin treatment was found to reduce hyperactivity, increase melatonin and dopamine levels, and regulate the circadian clock gene PER1b as illustrated in Table 2 [43].
Clinical studies involving children and adults with ADHD, sleep-onset insomnia (SOI), and DSPS have consistently reported improvements in sleep parameters following melatonin supplementation [44]. Findings include reduced sleep latency, increased total sleep time, and earlier DLMO, with no significant adverse effects reported [45,46]. The outcomes of these clinical trials are summarized in Table 2, further supporting the therapeutic role of melatonin for managing comorbid sleep disturbances in ADHD.
3.2. Hypothalamic–pituitary–adrenal (HPA) axis dysregulation
Hypothalamic-pituitary-adrenal (HPA) axis dysregulation has been reported in both circadian rhythm disorders and ADHD. The rhythmic output signals of the circadian system, originating from the SCN and areas influenced by the SCN, are crucial for synchronizing peripheral oscillators [47]. Cortisol, a key adrenal glucocorticoid stress hormone in the HPA axis, is a significant output orchestrated by SCN via arginine vasopressin and corticotropin-releasing hormone (CRH) [48]. Cortisol follows the circadian rhythm, rising before morning awakening, peaking within an hour, and gradually declining throughout the day [49]. Dysfunction in the behavioral inhibition system, possibly due to abnormal HPA axis responses to stress, has been proposed as a factor contributing to ADHD [50].
Lower cortisol levels in response to stress are associated with ADHD traits such as maladaptive behaviour and poorer cognitive performance in children [51]. Although inattentive subtypes also display blunted cortisol responses [52], some studies suggest that low cortisol responsivity is specific to the ADHD-combined type in children [53]. Factors such as treatment, comorbidity, and study design may account for these discrepancies. Additionally, sex differences in cortisol response have been noted, with boys exhibiting higher early morning cortisol levels and girls exhibiting lower levels [54]. Studies specify a phase delay in cortisol rhythm relative to wake time, though post-stress cortisol profiles remain unchanged in adults with ADHD [55].
Studies have indicated that ADHD is associated with alterations in cortisol secretion patterns, including a reduced cortisol awakening response (CAR) [56,57], lower overall cortisol levels compared to controls [58,59], and disruptions in cortisol rhythmicity, such as delayed acrophase (peak secretion time) [55] (Table 3). Some studies also reported increased cumulative diurnal cortisol and elevated morning and afternoon cortisol levels in ADHD individuals [60]. Additionally, variations in key circadian rhythm markers were observed, including lower BMAL1 and PER2 rhythms, reduced melatonin amplitude, and shortened actigraphic circadian periods in ADHD patients [55].
3.3. N-3 PUFAs deficiency
Another possible shared link between ADHD and circadian rhythm disorders is n-3 PUFAS deficiency. Literature supported the role of n-3 PUFAs in behavioral changes induced by inflammation in high risk groups such as children, pregnant women and patients with metabolic disorders [61]. Moreover, decreased brain-derived neurotrophic factor (BDNF) and increased pro-inflammatory cytokines such as Interleukin-6 and hsCRP have been shown in children with ADHD [59], while n-3 PUFAS have been shown to increase the expression of BDNF, tropomyosin receptor kinase B (TrkB), and cAMP response-element binding protein (CREB), which contribute to behavior and cognitive development and decrease the pro-inflammatory cytokines, including interleukin-1 (IL-1), interleukin-6 (IL-6), and tumour necrosis factor-alpha (TNF-α) in an in vitro study [62]. Previous studies demonstrated that ADHD children and adults with low n-3 index indicate more severe symptoms clinically (e.g., inattention and hyperactivity-impulsivity) [63] and physically, such as skin problems and dry eyes [64,65]. In response to this evidence, mounting studies of ADHD intervention by n-3 PUFAs supplementation, primarily with the high dosage of eicosapentaenoic acid (EPA) compared with docosahexaenoic acid (DHA), showed significant improvements in clinical symptoms [66] and cognitive performance in ADHD children and adolescents, in which particularly those with low endogenous levels of n-3 PUFAs [67].
Emerging research also highlights a potential role for n-3 PUFAs as circadian rhythm synchronizers. For instance, diets deficient in n-3 PUFAs have been shown to disrupt sleep patterns by affecting melatonin secretion and circadian clock gene expression [68]. Additionally, findings suggest a correlative association between sleep-wake cycles and fatty acid metabolism, as sleep deprivation has been linked to altered ketone body concentrations [69]. In vitro studies further demonstrate that palmitate can upregulate BMALI, a core circadian gene, but this effect is moderated in the presence of DHA, indicating that n-3 PUFAs may counteract the circadiandisrupting effects of saturated fats [70].
Based on the effects of n-3 fatty acids on the inflammatory process and cognition, which are regulated by the molecular clock, n-3 fatty acids may serve as circadian rhythm synchronizers. Various animal and human studies indicate that dietary fatty acids influence circadian rhythms. An experimental study conducted on Syrian Hamsters suggested that a diet deficient in n-3 PUFA disturbed nocturnal sleep, affecting the melatonin rhythm and also circadian clock functions [68]. Similarly, a large cohort study in type 2 diabetic patients described that marine n-3 PUFA consumption (EPA and DHA) was linked to reduced sleep impairment progression, mechanistically connected with increased central circadian regulation through upregulation of fundamental clock genes (CLOCK, BMAL1, PER2) and *RORα-*mediated BMAL1 nuclear translocation, with restoration of hypothalamic clock oscillations [71].
N-3 PUFAs in improving circadian rhythm via melatonin and HPA axis in ADHD
N-3 PUFAs may be able to improve circadian rhythm via modulation of melatonin output and HPA axis function. A study demonstrated that an n-3-deficient diet in 2-month-old rats reduced pineal DHA levels and its proportion in the phospholipids, phosphatidylcholine, and phosphatidylethanolamine, leading to lower melatonin release [71]. Another study showed that rats on an n-3-deficient diet exhibited lower nocturnal urinary excretion of 6sulfatoxymelatonin (aMT6), a melatonin metabolite, a condition that was reversible with DHA supplementation [72]. Lavialle et al. further demonstrated that an n-3-deficient diet resulted in a higher arachidonic acid (AA)/DHA ratio and reduced nocturnal melatonin peaks in Syrian hamsters, accompanied by increased striatal dopamine levels and hyper-locomotion. These alterations were linked to decreased levels of 12-Hydroxyeicosatetraenoic acid (12-HETE), which is a signaling molecule that is derived from PUFAs, and the crucial role of DHA in maintaining neuronal membrane integrity, affecting the function of enzymes, receptors, and dopamine transporters [68].
The effects of dietary n-3 fatty acids on behavior and learning in 15-month-old SenescenceAccelerated Mouse Resistant 1 (SAMR1) strain mice were studied. Mice fed a perilla oil diet displayed lower incorrect responses in learning tests and showed increased n-3 PUFAs in brain phospholipids compared to those fed a safflower oil (n-3-deficient) diet. Despite higher overall activity in the n-3-deficient group, both diet groups maintained similar circadian activity patterns. This indicates that the n-6/n-3 ratio in the diet significantly affects brain lipid composition, learning ability, and activity levels [73].
HPA-axis activity also has a modulating effect on fatty acid metabolism. Cortisol plays a role in mobilizing, lysis, oxidation, and synthesis of fatty acids [74]. For example, Cortisol has been shown to suppress the activity of Δ5-and Δ6-desaturase, key enzymes responsible for introducing double bonds into fatty acid chains, thereby affecting the synthesis of PUFAs [75]. This inhibition seems to have differential effects on specific fatty acids, in such a way that high cortisol concentrations are associated with a decrease in n-3 PUFA concentrations [76]. Conversely, fatty acids themselves can influence HPA-axis function. Supplementation with long-chain n-3 PUFAs, such as EPA, has been found to reduce cortisol levels in both animal models and human studies [68,77]. Additionally, inadequate maternal intake of n-3 PUFAs during early development has been linked to increased HPA-axis activity in rodent offspring [78].
N-3 supplementation appears to alter HPA-axis regulation through multiple mechanisms. It increases circulating levels of EPA and DHA while reducing AA concentrations, potentially modulating stress responses in three key ways. First, the structural properties of fatty acids influence glucocorticoid receptor function depending on their chain length and degree of unsaturation [79]. Second, EPA and AA modulate the function of pglycoprotein, which plays a role in cortisol transport across the blood-brain barrier [80]. Third, the ratio of AA to EPA affects the balance of pro- and antiinflammatory eicosanoids, which can, in turn, regulate HPAaxis activity by influencing CRH secretion and glucocorticoid receptor sensitivity [81].
N-3 PUFAs play a role in modulating circadian rhythms via melatonin secretion and cortisol levels, associated with impaired sleep quality and disrupted sleep-wake cycles [71,82]. Currently, there is no clinical study specifically focused on the role of n-3 in regulating circadian rhythms in ADHD individuals; however, an animal study conducted on n-3 PUFAsdeficient diet hamsters exhibited symptoms such as disturbed melatonin rhythm and chronic locomotor hyperactivity [68] (Table 4). Differences in circadian architecture, neuroendocrine regulation, and behavioral complexity between these models and humans mean that findings may not fully extrapolate to clinical populations. Moreover, there is a relative scarcity of high quality human trials directly examining the effects of n-3 supplementation on circadian outcomes in ADHD, underscoring the need for well-designed clinical studies to validate these preclinical mechanisms. Some research has explored the impact of n-3 on sleep disorders in patients with ADHD. A study investigated the effects of n-3 PUFA supplementation containing 400 mg EPA, 40 mg DHA, in addition to 60 mg of n-6, in combination with 80 mg of magnesium and 5 mg of zinc on ADHD symptoms and sleep-related problems [83]. The study concluded that n-3 PUFAs showed promising effects in reducing ADHD symptoms, but no significant improvement was observed in sleep problems. Another study was conducted on preterm toddlers to check the effect of n-3, 6, and 9 on behavior and sleep, utilizing a supplement containing 706 mg total n-3 fatty acids (338 mg EPA, 225 mg DHA), 280 mg total n-6 fatty acids (83 mg GLA), and 306 mg total n-9 fatty acids (oleic acid), identified the improvement in socioemotional outcomes, but no effect was observed on sleep and behavior aspects [84]. Similar results were observed in another study, in which 200 mg/day DHA and 200 mg/day AA supplementation had no overall impact on child sleep. However, male children and children whose caregivers had depressive symptomatology seemed to benefit from DHA + AA supplementation in exploratory subgroup analyses, and the beneficial effects continued for some months following the end of supplementation [85]. Although, dietary n-3 PUFAs may improve cognitive regulation, behavior, sleep, and daytime functioning in ADHD, but findings remain inconsistent, largely due to methodological heterogeneity rather than lack of efficacy. Variability arises from dosage and EPA: DHA ratios, as many trials use modest doses (<1 g/day) that may be insufficient to correct deficiencies or affect circadian regulation. Higher EPA content appears more effective for core ADHD symptoms, whereas DHA is more directly linked to melatonin synthesis, membrane stability, and circadian gene modulation; thus, trials emphasizing one fatty acid may produce divergent outcomes. Intervention duration also varies, with shorter trials potentially underestimating benefits that require sustained neuronal incorporation of long-chain n-3 PUFAs. Finally, adjunctive treatments (e.g., magnesium, zinc, stimulants) complicate interpretation by obscuring the specific contribution of n-3s.
Circadian and neuroendocrine disturbances in ADHD represent an interconnected cascade rather than isolated events. N-3 PUFAs provide a modulatory link, influencing circadian timing, stress responses, and inflammation. A central premise is that n-3 PUFAs act on the master circadian clock in the SCN by regulating core clock genes such as BMAL1 and CLOCK, thereby restoring synchronized rhythms. This normalization extends to downstream neuroendocrine and inflammatory pathways.
As the SCN regulates melatonin, n-3 deficiency has been associated with reduced melatonin rhythm and nocturnal sleep disturbances, whereas supplementation restores secretion and supports timely sleep. Similarly, the HPA axis and its cortisol rhythms are under circadian control. By dampening inflammation and lowering cytokines such as IL-6 and TNF-α, n-3 PUFAs indirectly stabilize cortisol release, often disrupted in ADHD. As summarized in Fig. 1, n-3 PUFAs act as central regulators, modulating circadian gene expression and aligning neuroendocrine and inflammatory outputs. This integrated mechanism addresses multiple facets of ADHD pathophysiology, highlighting the potential of n-3 supplementation as a holistic therapeutic strategy.
Conclusion
Sleep and circadian rhythm disruptions are known to exacerbate ADHD symptoms, highlighting the need for targeted clinical interventions. Emerging evidence indicates that n-3 PUFAs may represent a safe and well-tolerated therapeutic strategy for managing both ADHD symptoms and circadian rhythm disturbances. This review integrates ADHD, circadian mechanisms and n-3 PUFAs to propose a novel framework in which n-3 PUFAs offer a mechanistic bridge between metabolic, immune and chronobiological regulation in ADHD However, the lack of clinical trials directly assessing circadian endpoints in ADHD patients receiving n-3 supplementation remains a critical gap. Future studies should prioritize well designed clinical trials to evaluate the specific effects of n-3 supplementation on circadian rhythm disorders in individuals with ADHD, with a focus on determining the optimal dosage, timing, and formulation to maximize therapeutic outcomes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1American Psychiatric Association Diagnostic and Statistical Manual of Mental Disorders: DSM-5 5th ed Arlington, VA American Psychiatric Association 2013 10.1176/appi.books.9780890425596 · doi ↗
- 2Ayano G Demelash S Gizachew Y Tsegay L Alati R The global prevalence of attention deficit hyperactivity disorder in children and adolescents: an umbrella review of meta-analyses J Affect Disord 2023 339 860 6 10.1016/j.jad.2023.07.071 37495084 · doi ↗ · pubmed ↗
- 3Ahlberg R Garcia-Argibay M Taylor M Lichtenstein P D'Onofrio BM Butwicka A Prevalence of sleep disorder diagnoses and sleep medication prescriptions in individuals with ADHD across the lifespan: a Swedish nationwide register-based study BMJ Ment Health 2023 26 e 300809 10.1136/bmjment-2023-300809 PMC 1057771037657817 · doi ↗ · pubmed ↗
- 4Lunsford-Avery JR Kollins SH Editorial Perspective: delayed circadian rhythm phase: a cause of late-onset attention-deficit/hyperactivity disorder among adolescents? JCPP (J Child Psychol Psychiatry) 2018 59 1248 51 10.1111/jcpp.12956 30176050 PMC 6487490 · doi ↗ · pubmed ↗
- 5Bijlenga D Vollebregt MA Kooij JJS Arns M The role of the circadian system in the etiology and pathophysiology of ADHD: time to redefine ADHD? Atten Defic Hyperact Disord 2019 11 5 19 10.1007/s 12402-018-0271-z 30927228 · doi ↗ · pubmed ↗
- 6StorebøOJ Storm MRO Pereira Ribeiro J Skoog M Groth C Callesen HE Methylphenidate for children and adolescents with attention deficit hyperactivity disorder (ADHD) Cochrane Database Syst Rev 2023; 3 CD 009885 10.1002/14651858.CD 009885.pub 3 PMC 1004243536971690 · doi ↗ · pubmed ↗
- 7Sobanski E Schredl M Kettler N Alm B Sleep in adults with attention deficit hyperactivity disorder (ADHD) before and during treatment with methylphenidate: a controlled polysomnographic study Sleep 2008 31 375 81 10.1093/sleep/31.3.375 18363314 PMC 2276739 · doi ↗ · pubmed ↗
- 8Potter GD Skene DJ Arendt J Cade JE Grant PJ Hardie LJ Circadian rhythm and sleep disruption: causes, metabolic consequences, and countermeasures Endocr Rev 2016 37 584 608 10.1210/er.2016-1083 27763782 PMC 5142605 · doi ↗ · pubmed ↗