Characterization of extended-spectrum cephalosporin-resistant Klebsiella recovered from dairy manure in Southern Ontario, Canada
Rebecca E. V. Anderson, Gabhan Chalmers, Andrew Scott, Roger Murray, Michael Mulvey, Edward Topp, Patrick Boerlin, Nicole Ricker

TL;DR
This study examines antibiotic-resistant Klebsiella bacteria in dairy manure from Ontario, revealing diverse resistance genes and plasmids that could spread between animals and humans.
Contribution
The study provides the first characterization of ESC-resistant Klebsiella from Canadian livestock, including plasmid and sequence type diversity.
Findings
Eighty-six percent of Klebsiella isolates from dairy manure were multi-drug resistant.
Variants of CTX-M resistance genes were found on multiple plasmid types and the chromosome.
Conjugation of resistance plasmids occurred between Klebsiella and E. coli, highlighting potential for gene spread.
Abstract
Extended-spectrum cephalosporin (ESC)-resistant Klebsiella pneumoniae are a problem in human patients and have been studied extensively. However, there is a paucity of information regarding ESC-resistant K. pneumoniae from livestock in general, and in Canada in particular. This study characterized ESC-resistant K. pneumoniae recovered from dairy manure in Ontario, Canada, and their ESC-resistance plasmids. ESC-resistant K. pneumoniae (n = 73) and K. quasipneumoniae (n = 11) isolates were screened by PCR for blaCTX-M, blaCMY and blaSHV prior to undergoing antimicrobial susceptibility testing using disk diffusion. Isolates from dairy manure carrying blaCTX-M (n = 74), and additional isolates from turkeys (n = 8) and dogs (n = 2), underwent short-read whole genome sequencing (WGS) and a subset of these (n = 35) had additional long-read sequencing and hybrid assembly for confirmation.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
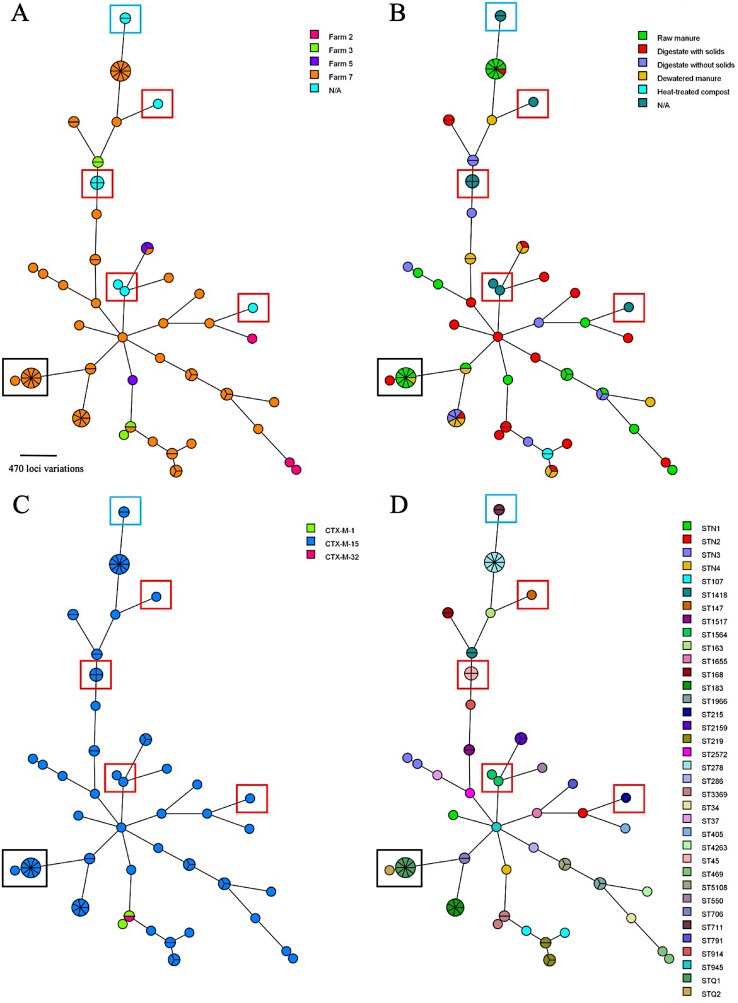 Fig 1
Fig 1 Fig 2
Fig 2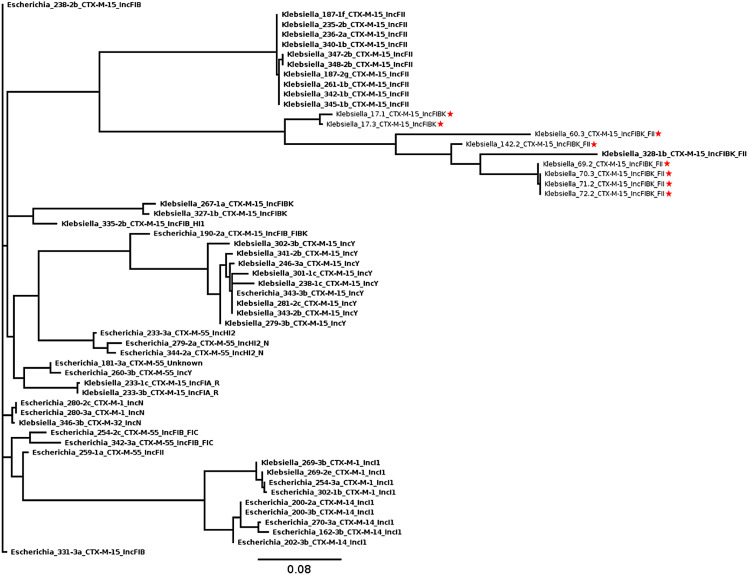 Fig 3
Fig 3 Fig 4
Fig 4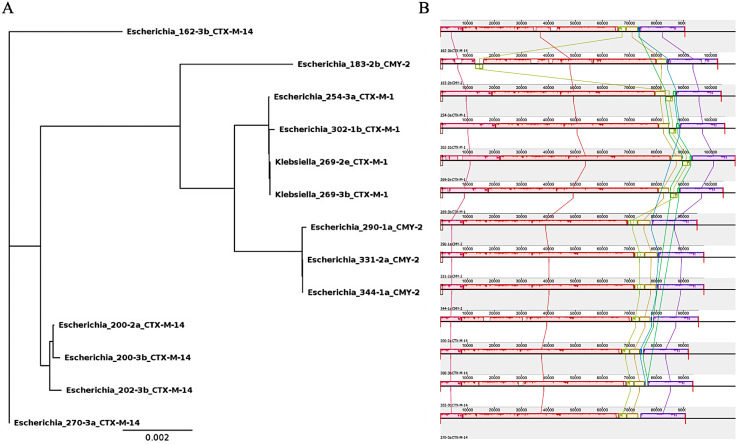 Fig 5
Fig 5- —Canadian Institute of Health Research (CIHR)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Antibiotic Use and Resistance · Antimicrobial agents and applications
Introduction
Klebsiella pneumoniae is a member of the ESKAPE group of priority pathogens [1] and is of clinical relevance for both humans and animals. In humans, K. pneumoniae is a common cause of urinary tract infections, septicemia, and abdominal infections [2,3] and is amongst the top five causes of bacterial pneumonia in Canadian hospitals [4]. In animals, K. pneumoniae is associated with mastitis in dairy cattle, urinary tract infections (UTIs) in dogs, pneumonia in horses, and a variety of other opportunistic infections [5–7]. Farm animals such as dairy cattle may also play a role in the dissemination of enteric organisms into the environment and subsequently into the food chain through manure treatment onto crops, such as has been documented for E. coli transfer to lettuce [8].
Antimicrobial resistance (AMR) is a rising global health issue in humans and animals, with resistance to the critically important extended-spectrum cephalosporins (ESCs) documented originally in human clinical cases, and subsequently in companion animals and livestock [9]. More research is needed to identify the role K. pneumoniae plays in the spread of AMR especially since, in recent years, the frequency of multi-drug resistant (MDR) K. pneumoniae strains, including ESC-resistance, observed among human clinical infections has increased [1,10]. However, most research surrounding ESC-resistance in non-human sources has focused on Escherichia coli and Salmonella enterica. Ceftiofur, an ESC, is an important antimicrobial in the dairy industry for the treatment of severe clinical mastitis caused by Klebsiella spp. [11], particularly if the strain is resistant to first line antimicrobials [12]. Despite these concerns, there is little information regarding ESC-R K. pneumoniae from animals and the environment, including in Canada.
Recent studies have shown that ESC-resistance in K. pneumoniae is usually associated with blaCTX-M, blaCMY and the intrinsic, blaSHV [10,13,14] genes. The most frequent variants of CTX-M in K. pneumoniae from human and animal sources are CTX-M-14, −15 and −55 [15–19]. K. pneumoniae may therefore have a role in the dissemination of AMR genes through horizontal gene transfer (HGT) [20,21], as CTX-M-14 and −15 have been associated with conjugative plasmids in diverse hosts within the Enterobacterales [22].
Farm animals play a role in the One Health cycle due to their intimate interactions with humans (both farm workers and consumers) and the environment through manure application. The overarching goal of this study was to gain insights into the role that K. pneumoniae plays in the context of antimicrobial resistance and One Health in relation to dairy cattle. The objectives of this study were threefold. The first was to assess the diversity and distribution of ESC-resistant K. pneumoniae in a sample of dairy farms in Ontario. The second objective was to characterize the diversity and distribution of blaCTX-M-harbouring plasmids within the dairy manure collection compared to other relevant sources, including other animal species in our collection and sequenced plasmids available through public databases. Finally, the last objective was to understand the transferability of blaCTX-M-harbouring plasmids between K. pneumoniae and E. coli. The results of the study will contribute to a better understanding of the epidemiology of ESC-resistance in K. pneumoniae in Canada, and of transmission of ESC-resistance plasmids between bacterial species.
Materials and methods
Farm sampling and bacterial isolates
K. pneumoniae isolates (n = 120) were screened using polymerase chain reaction (PCR) for blaCTX-M, blaCMY, and blaSHV (Table 1). PCR positive isolates (n = 84) were identified from 34 manure samples recovered in four of six sampled farms in Southern Ontario between November 2018 and August 2019 [23]. Manure samples were taken before treatment, during anaerobic digestion at mesophilic and thermophilic temperatures, and at the final output stages including the liquid manure applied to land under cultivation to grow crops, and the dewatered material used for livestock bedding [23]. Additionally, ESC-resistant K. pneumoniae from turkey fecal samples (*n *= 23) [24] and dog clinical samples (n = 13) [25] were also investigated. Species identification was performed using matrix-assisted laser desorption ionization time of flight (MALDI-TOF; Bruker Daltonik GmbH, Bremen, Germany) at the Animal Health Laboratory (AHL), University of Guelph, ON.
Table 1: Primers used for PCR screening to characterize ESC-resistant isolates.
Characterization of ESC-resistant K. pneumoniae
Antimicrobial susceptibility profiles were determined using the disk diffusion method, following guidelines and interpretation criteria from CLSI [28,29] for cefoxitin (FOX), cefotaxime (CTX), ceftazidime (CAZ), ertapenem (ETP), gentamicin (GEN), ampicillin (AMP), amoxicillin-clavulanic acid (AMC), sulphonamide compounds (SUL), sulfamethoxazole-trimethoprim (SXT), tetracycline (TET), streptomycin (STR), kanamycin (KAN), ciprofloxacin (CIP) and chloramphenicol (CHL). Isolates resistant to antimicrobials from three or more antimicrobial classes, not including intermediate phenotypes, were classified as MDR. For the sake of defining MDR, the ß -lactamase category was divided into the following sub-categories: cephems (CTX, CAZ and FOX), ß-lactam/ß-lactamase inhibitor combinations (AMC), carbapenems (ETP) and other ß-lactams (AMP). AMC was not considered in the determination of multi-drug resistance since carrying blaCMY would naturally result in multi-drug resistance (i.e., resistance to cephems, other ß-lactams, and ß-lactam/ß-lactamase inhibitor combinations).
Whole genome sequencing (WGS)
K. pneumoniae carrying blaCTX-M from dairy manure (n = 74), turkey (n = 8) and dog (n = 2) underwent short read WGS. Preparation of DNA for WGS was done using Epicentre MasterPure^TM^ DNA Purification kit (Epicentre, Madison, WI) following the manufacturer’s instructions. Sequencing was done either with Illumina Miseq (PE300; Illumina, San Diego, CA) at the Advanced Analysis Centre, University of Guelph, Guelph, ON, or Illumina NextSeq (PE150) at the National Microbiology Laboratory in Winnipeg, MB, (NML Winnipeg).
A subset of isolates carrying blaCTX-M from dairy manure (*n *= 27), in addition to the isolates from turkey feces (n = 8) and dog clinical samples (n = 2), were also sequenced using the MinION Platform (Oxford Nanopore Technologies). The subset of dairy manure isolates were selected using a phenetic tree created with the inhibition zone diameters from antimicrobial susceptibility testing. This tree was created in BioNumerics v7.6 (Applied Maths, Austin TX. USA) using Pearson correlation coefficients and cluster analysis with the unweighted pair group method with arithmetic mean. One or two isolates were selected from each cluster defined at a 94% cut off, to include the highest diversity of strains from different time periods and treatment processes. Preparation of DNA samples was performed as above. Sequencing libraries and barcoding preparation were done using the SQK-LSK109 and EXP-NBD104/114 ligation and native barcoding kits (Oxford Nanopore Technologies) according to the manufacturer’s instructions. Flow cells (version FLO-MIN106 R9.4) were run for 72h each, or until exhausted. Basecalling of fast5 files and demultiplexing was performed using Guppy Basecaller v3.3 (Oxford Nanopore Technologies) with barcode trimming enabled.
Sequence analysis
Short reads were first used to assemble the genome sequences in BioNumerics v7.6 (Applied Maths, Austin TX. USA) using the SPAdes K. pneumoniae de novo assembler, and assembly-free and assembly-based allele calling. Resistance genes and plasmid markers were identified using the E. coli functional genotype plug-in for BioNumerics at a 90% similarity which utilizes ResFinder and PlasmidFinder databases from the Center for Genomic Epidemiology (Technical University of Denmark, DTU). Multi-locus sequence types (MLSTs) were manually assigned using a MLST finder database also from the Center for Genomic Epidemiology [30,31] with paired end Illumina reads. Any isolates with an unknown sequence type (ST) due to a novel allele or novel allelic configuration, were thoroughly analyzed to identify any errors in allelic regions by reference mapping of short reads. If no errors were present, and no ST was assigned, these strains were submitted to the Pasteur Institute (https://bigsdb.pasteur.fr/klebsiella/) and assigned novel ST designations.
Plasmid assembly and analysis
Long and short reads were assembled using hybrid assembler Unicycler v0.4.8 [32], in parallel with Flye v2.6 [33] and were polished with short reads using Racon v1.4.0 [34] and Pilon v1.23 [35]. Both Flye and Unicycler pipelines were used for quality control purposes; however, Unicycler assemblies were used for all downstream analysis. Assemblies were visualized using Bandage v0.8.1 [36] and annotated with ABRicate v0.9.8 [37] against the ResFinder v4.1 [38], PlasmidFinder v2.1 [39] and VFDB [40] databases. Sequenced genomes were analyzed using Geneious v9.1.8 (Biomatters, Auckland, New Zealand) and plasmids were aligned and visualized using Mauve plug-in v2.3.1 [41]. Alignment of plasmids was performed using EasyFig [42].
Analysis of genetic relationships
Minimum spanning trees (MSTs) were generated using the whole genome multi-locus sequence typing (wgMLST) (core enterobase) function for core genome MLST (cgMLST) trees in BioNumerics v7.6 (Applied Maths, Sint-Martens-Latem, Belgium) with 10x bootstrapping and no multithreading. This approach utilized the Klebsiella pneumoniae WGS scheme, which includes 634 core loci.
Core gene phylogenetic analyses on isolates recovered from dairy manure were also performed using Snippy v4.4.5 according to developer’s guidelines, under Core SNP Phylogeny (https://github.com/tseemann/snippy) with K. pneumoniae type strain ATCC 13883 (GenBank Accession #JOOW00000000.1) and K. quasipneumoniae subsp. quasipneumoniae strain 01A030 (GenBank Accession #GCA_000751755.1) as reference strains. The clean.full.aln files were analyzed with Gubbins v2.4.0 [43] and the output clean.core.aln file was analyzed with SNP-sites v3.0 [44]. FastTree v2.2.11 [45] was used to generate the trees which were visualised in Geneious v9.1.8.
In order to compare blaCTX-M harbouring plasmids, phylogenetic SNP analyses of the hybrid assembled ESC-resistance plasmids were conducted using a pre-determined protocol [46]. A gene presence/absence analysis was first conducted on all ESC-resistance plasmids independently of replicon types, including the ESC-plasmids in Klebsiella spp. from dairy cattle and turkey, in addition to ESC-resistance plasmids in E. coli from the same dairy manure samples [23]. These plasmids (n = 65) were first analyzed with Prokka v1.14.6 (https://github.com/tseemann/prokka) with the output.gff files analyzed using Roary v3.13.0 [47] A SNP analysis was then performed individually on IncY replicon, IncI1 and IncFII plasmids. Analysis for both IncY replicons and IncI1 plasmids also incorporated plasmids from E. coli isolated in a parallel study [23] using the SNP-sites generated from the core.gene.alignment output file from Roary. The SNP sites along with the core.gene.alignment file, were used to create a maximum likelihood tree with IQtree v2.0.3 [48]. All trees were visualized in Geneious v9.1.8 (Biomatters, Auckland, New Zealand).
Conjugation assays
To determine inter-species transfer of blaCTX-M harbouring plasmids through conjugation, K. pneumoniae (n = 3) were used as donors and E. coli CV601gfpKan^R^Rif^R^ as recipient strain [49]. Transfer of blaCTX-M plasmids from E. coli donors (n = 3) to the K. pneumoniae ATCC13883∆Nal^R^ recipient was also assessed. K. pneumoniae and E. coli donors included isolates which carried either IncI1 or IncF plasmids from previous studies [23].
All conjugation assays were quantitative and conducted in biological triplicate at 30˚C and 37˚C. For broth conjugations, cultures were grown overnight at 37˚C with shaking in 5 ml LB broth containing appropriate selective antimicrobials for donor E. coli and K. pneumoniae strains (CTX 2 mg/ml), recipient E. coli (RIF 50 mg/ml and KAN 50 mg/ml) and recipient K. pneumoniae (NAL 64 mg/ml) for the donor and recipient strains. Cultures were pelleted, washed three times with LB broth containing no antibiotics and resuspended. Donor (200µl) and recipient (200µl) strains were added to 1.6 ml LB broth and incubated for 16h without shaking at 30˚C or 37˚C. Conjugations were serially diluted and plated (100µl from each dilution) on appropriate selective and non-selective media. Three colonies per transconjugant selective medium were grown in pure cultures and confirmed to carry blaCTX-M using PCR. Final conjugation populations were calculated based on the ratio of transconjugants to recipient CFU/mL [49]. In addition to the broth mating for E. coli donors to K. pneumoniae recipients, surface mating was also conducted. Donor (20µl) and recipient (20µl) strains were inoculated onto filter paper (1 cm^2^) on LB agar and incubated for 8h at 30˚C or 37˚C. Filter papers were removed with sterile forceps and added to 1 ml of LB broth, vortexed and serially diluted on appropriate selective media.
The transferability of blaCTX-M-15-harbouring IncY plasmids was assessed through quantitative filter-mating conjugation experiments performed in biological triplicate and technical duplicate. E. coli isolate 343-3b [23], and K. pneumoniae isolate 343-2b were used as donor strains, while E. coli CV601gfpKan^R^Rif^R^ [49] was used as the recipient strain. Strains were inoculated into 20 mL LB broth supplemented with cefotaxime (2 mg/L) for donor strains and rifampicin (16 mg/L) for the recipient strain. Filter plating was performed as described above, except that 100 μL of donor and recipient were plated and conjugation was allowed to continue for 24 hours before plating. Two independent transconjugant colonies (one circular, and one punctiform) from the K. pneumoniae to E. coli and the E. coli to E. coli conjugations were extracted using the MasterPure™ Complete DNA and RNA Purification Kit (Lucigen, cat no MC85200) before long read sequencing was performed using a MinION Mk1B device (Oxford Nanopore Technologies, Oxford, United Kingdom).
Results
Antimicrobial susceptibility and multi-drug resistance
Klebsiella spp. carrying blaCTX-M, blaCMY or blaSHV (n = 84) from dairy cattle were identified through PCR screening from multiple stages of manure treatment across four Ontario dairy farms. All isolates were primarily identified as K. pneumoniae using MALDI-TOF. However, further analysis using full length 16S rRNA gene read mapping identified eleven isolates as Klebsiella quasipneumoniae subspecies quasipneumoniae, all of which carried blaOKP instead of the intrinsic blaSHV gene, as identified through WGS. The K. pneumoniae and K. quasipneumoniae will be described accordingly throughout this study. Only two farms (n = 5 isolates) were positive for Klebsiella spp. carrying blaCMY. Eighty-three percent (70/84) of the Klebsiella spp. isolates from dairy manure carrying blaCTX-M and blaSHV or blaOKP were multi-drug resistant (MDR) (Table 2). As expected, all isolates from dairy cattle carrying blaCTX-M (77 total) were resistant to ampicillin and cefotaxime, while 90.9% (70/77) were resistant to ceftazidime and 50.6% (39/77) had an intermediate phenotype for amoxicillin-clavulanic acid. Only one isolate (1.3%; 1/77) was fully resistant to amoxicillin-clavulanic acid and it did not carry any additional ß-lactamase genes. Among Klebsiella spp. carrying blaCTX-M, no resistances to ertapenem or cefoxitin were observed, and the most frequent resistances other than to ß-lactams were to tetracyclines (77.9%; 60/77), streptomycin (64.9%; 50/77), sulfonamide (57.1%; 44/77) and sulfamethoxazole-trimethoprim (54.5%; 42/77). The least common resistances were to chloramphenicol (9.1%; 7/77), kanamycin (6.5%; 5/77) and gentamicin (2.6%; 2/77). Although 27.3% (21/77) of the isolates had an intermediate susceptibility phenotype for ciprofloxacin, none were fully resistant. All blaCTX-M-positive isolates were subjected to WGS; however, three were removed from further analysis due to low sequencing coverage, leaving 63 K. pneumoniae and 11 K. quasipneumoniae in the WGS analysis. Isolates carrying blaCMY from dairy manure and isolates solely carrying blaSHV underwent susceptibility testing to the panel of antimicrobials but were not included in WGS. Among these, most isolates carrying blaCMY with blaSHV (3/5) and all isolates carrying blaSHV only (2/2), were MDR. Among K. pneumoniae from turkey feces (n = 23), all isolates carrying blaCTX-M, blaCMY, or blaSHV were MDR (Table 2). Similarly, isolates from dog clinical samples (n = 13) were all MDR except for one isolate carrying blaCMY with no detectable blaSHV (Table 2).
Table 2: Antimicrobial susceptibility profiles for ESC-resistant Klebsiella spp. Recovered from dairy manure, turkey feces and dog clinical samples. Antimicrobial susceptibility profiles were based on disk diffusion using cefoxitin (FOX), cefotaxime (CTX), ceftazidime (CAZ), ertapenem (ETP), gentamicin (GEN), ampicillin (AMP), amoxicillin-clavulanic acid (AMC), sulfonamide (SUL), sulfamethoxazole-trimethoprim (SXT), tetracycline (TET), streptomycin (STR), kanamycin (KAN), ciprofloxacin (CIP) and chloramphenicol (CHL). Antimicrobials were grouped to determine multi-drug resistance; aminoglycosides (KAN, GEN, STR), tetracyclines (TET), quinolones (CIP), folate pathway inhibitors (SUL, SXT), phenicols (CHL), cephems (CTX, CAZ, FOX), carbapenems (ETP) and other ß-lactams (AMP). Intermediate was considered susceptible for the multi-drug resistance classification and AMC was excluded from multi-drug resistance classification. Abbreviations: An.: animal source; DM: dairy manure; T: turkey; D: dog; MDR: multi-drug resistant.
ESC-resistance gene diversity
Among the three animal sources, three blaCTX-M variants were identified using WGS which included blaCTX-M-15 (n = 81), blaCTX-M-1 (n = 2) and blaCTX-M-32 (n = 1). Of the 81 isolates carrying blaCTX-M-15 (70 K. pneumoniae and 11 K. quasipneumoniae), 73 also carried blaTEM-1 and eight of these also carried blaOXA-1. The isolates carrying blaCTX-M-1 or blaCTX-M-32 did not carry either blaTEM-1 or blaOXA-1 (Table 3). Most dairy manure ESC-R isolates investigated (73/84, 87%) carried a blaSHV gene. The remaining 11 all carried blaOKP and were shown by WGS to be K. quasipneumoniae. Of the blaSHV gene variants identified, the only known extended-spectrum ß-lactamase (ESBL) was SHV-27, which was found in 4 isolates. There were also 3 novel SHV variants identified, with isolates 233-1c and 233-3c having the same variant (Table 3). These two isolates were from the same sample and have the same ST, suggesting they could be the same strain. The variants of blaSHV found in K. pneumoniae from turkey feces (blaSHV-201 and blaSHV-148) and from dog samples (blaSHV-187) were not found in any Klebsiella spp. from dairy manure (Table 3). There were also two isolates from dairy cattle manure that had truncated blaSHV genes.
Table 3: Source and characteristics of Klebsiella spp. isolates carrying blaCTX-M recovered from dairy cattle manure, turkey feces or dog clinical samples in Canada. Isolates carrying blaOKP instead of blaSHV are K. quasipneumoniae subsp. quasipneumoniae and bolded STs are those recovered in samples from more than one farm source. SHVΔ refers to truncated SHV genes.
The majority of isolates carrying blaCTX-M were from the largest farm of the study (Farm 7, 64/74 isolates). Although ESC-R isolates were found throughout the manure treatment process, recovery was consistently low and highly variable, which did not allow for delineation of any clear trends with respect to manure treatment stages. Isolates positive for blaCTX-M were found in raw manure in three farms, in the liquid end-product to be applied to the environment in two farms and in the dried end-product to be recycled as bedding in two farms. The eleven K. quasipneumoniae isolates were also all from Farm seven. They were found mainly in raw manure, but also in two different downstream processing stages (Table 3).
Sequence type diversity among blaCTX-M positive isolates
Among isolates from dairy manure, 30 STs were identified through multi-locus sequence typing (MLST) (Table 3). Additionally, five isolates were identified as belonging to four different novel STs, ST7678, ST7679, ST5837 and ST6320 (S1 Table). The K. quasipneumoniae (*n *= 11) were identified as having novel MLST profiles and were assigned ST5682 using the MLST scheme for K. pneumoniae as a foundation for differentiation (S2 Table). The most frequently recovered STs from dairy manure were ST278 (6 samples), ST183 (4 samples) and ST219 (4 samples) (Table 3), all of which were isolated from the largest farm of the study (Fig 1A). Many STs were only identified once (Table 3). Two K. pneumoniae STs, ST2159 and ST3369, were found on more than one farm (Fig 1A). None of the STs found in turkey feces or dog urine samples overlapped with those found in dairy manure (Fig 1D). The STs recovered from turkey feces included ST45 (n = 4), ST1564 (n = 2), ST215 (n = 1) and ST147 (n = 1). The two isolates from dog urine samples were both ST711 and were recovered from urine samples taken at different times from the same patient diagnosed with a UTI. It is unknown whether the UTI was due to a persistent infection or a re-infection with the same strain.
Minimum spanning trees created using cgMLST with 634 core genome loci for K. pneumoniae (n = 63) and K. quasipneumoniae (n = 11) isolates from dairy manure, turkey feces (n = 8) and dog urine (n = 2). Trees are colour coded based on A) dairy farm, B) dairy manure treatment process, C) blaCTX-M variant and D) MLST profile. The black square encapsulates the K. quasipneumoniae isolates (n = 11), blue square encapsulates K. pneumoniae from dog urine samples and the red squares indicate isolates from turkey feces. On all farms, digestate with solids is the first anaerobically digested manure and digestate without solids is environmentally applied manure. On farm five, dewatered manure is used as recycled bedding whereas on farm seven, dewatered manure is thermophilically treated to create compost, which is then used as recycled bedding.
Of the STs found in our study, seven (ST107, ST1966, ST34, ST37, ST405, ST45 and ST711) were also found in the human clinical isolate database (National Center for Biotechnology Information (NCBI) BioProject PRJNA717739 [50]). Of these 7 human-associated isolates, four ST groups (ST37, ST405, ST45, and ST711) had similar ESC-resistance profiles to our K. pneumoniae samples from dairy manure and were resistant to ceftazidime, ceftriaxone and cefepime. From the human clinical isolates database, ST37 was from a urine sample in Asia in 2019, ST45 was from a respiratory sample in North America in 2013, ST405 was from a respiratory sample in South America in 2013, and ST711 was from a wound sample in Africa in 2019 [50].
The genomic similarities of isolates from all three animal sources were visualized using cgMLST Minimum Spanning Trees (MSTs) to identify any strain clustering among or between sources (Fig 1). Additionally, Klebsiella spp. isolates from dairy manure were further analyzed using the more discriminatory core SNP analysis to better understand distribution and persistence throughout manure treatments (Fig 2). Clusters of isolates were found to originate from several samples and therefore are not likely to be replicates. Clusters in both trees are supportive of one another. As mentioned above, only two clusters of isolates were found on more than one farm (Fig 1A) corresponding with ST2159 and ST3369. The ST2159 cluster includes three blaCTX-M-15-carrying isolates that are closely related and had no core SNP differences identified (246-3a, 246-3b, and 280-1b in Fig 2A). The ST3369 cluster includes two highly related isolates carrying blaCTX-M-1 (269-2e and 269-3b which have no SNP differences) from the same farm and sample, and one slightly different isolate (346-3b, 164 SNP differences from the other two isolates), which is from another farm and carried blaCTX-M-32 (Figs 1C and 2).
Single nucleotide polymorphism (SNP) maximum likelihood phylogenetic tree using 128,910 core SNP sites from K. pneumoniae isolates (n = 63) carrying blaCTX-M in reference to K. pneumoniae type strain ATCC 13883 (GenBank Accession #JOOW00000000.1). Isolate identifications are comprised of the sample number, followed by replicate number and colony ID. Therefore, isolates with the same three digits are from the same sample and possibly clones.
Two dairy manure isolates (ST219) carried blaCTX-15 on the chromosome and are linked by their source (Figs 1A and 1B), which was also the case for the two dog isolates (ST711). The two isolates from dairy manure (302-2b and 304-2b) (ST219, Fig 2) were from the same farm and sampling date, but different manure treatment stages. They had no core genome SNPs and had the same AMR gene profile. ISEcp1 insertion sequences were located upstream of the blaCTX-M-15 on the four chromosomes. For the dairy isolates, insertion had occurred within a conserved region of the chromosome and interrupted an aldose-1-epimerase gene. For the dog isolates, the insertion was instead in a different region with multiple IS elements and resistance genes, suggesting high plasticity.
Persistence through dairy manure treatments
Using core genome SNP analysis on all sequenced K. pneumoniae isolates from dairy manure, excluding the eleven K. quasipneumoniae, nine clusters are evident (Fig 2). Of these, five [ST278 (0–31 SNPs, ST183 (0–31 SNPs), ST1966 (30–81 SNPs), ST706 (51 SNPs) and ST469 (20 SNPs)] contain closely related isolates which were recovered from different treatment stages and sampling dates. Two of the nine clusters [ST7679 (10 SNPs) and ST1966 (51–81 SNPs)] contain closely related isolates recovered from both the input and output stages of manure treatment from the same farm. Three identical ST5108 isolates were recovered from raw manure at different sampling periods but not found at other treatment stages. Additionally, closely related strains ST107 (n = 2) and ST219 (n = 5), were recovered from various stages of the treatment process (Fig 2). Within this branch, two ST219 (261-1a and 261-1b; 0 SNPs), likely to be the same clone, were recovered from heat treated compost in March, and were distinguishable by 288 SNPs from the other three ST219 isolates recovered from digestate with solids (302-2b in June) and dewatered manure (282-2b in April and 304-2b in June).
Plasmid diversity
A subset of 35 isolates were sequenced with both long and short reads to enable ESC-resistance plasmid assembly (Table 4). Plasmids in isolates from dairy manure carried blaCTX-M-15 on IncY (n = 8), IncFII (n = 10), IncFIB(K) (n = 2), cointegrated IncR/IncFIA/Col (n = 2), IncFIB/IncHI1 (n = 1), and IncFIB(K)/IncFII (n = 1) replicons. Two of the isolates harbouring IncY replicons carrying blaCTX-M-15 were K. quasipneumoniae. Plasmids carrying blaCTX-M-15 from turkey samples were IncFIB(K)/IncFII (n = 6) or IncFIB(K) (n = 2). Two K. pneumoniae isolates from dairy manure carried blaCTX-M-1 on IncI1/ST3 plasmids and one isolate carried a blaCTX-M-32 on an IncN plasmid.
Table 4: Characteristics of blaCTX-M plasmids of K. pneumoniae and K. quasipneumoniae isolated from dairy cattle (DC) manure on dairy farms or turkey (T) feces from Canadian sources.
The IncY replicons carrying blaCTX-M-15 (n = 8) were all from the same farm except for 246-3a. These IncY replicons were recovered from seven different STs (Table 4), and the samples they originated from spanned the entire manure treatment pipeline. However, the two isolates from raw input manure were both K. quasipneumoniae (301-1c and 279-3b). Despite being harbored by different hosts and on two separate farms, the IncY replicons were practically identical, with minor variations in copy number and location of insertion sequences (S1A Fig, S1B Fig). The IncY replicons identified in this study were annotated as conjugative plasmids, unlike many other documented IncY elements that have been classified as prophages or phage-plasmids [51], and they carried an extensive AMR region, including genes conferring resistance to tetracycline, sulfonamides, streptomycin, and aminopenicillins (S1 C Fig). Nearly identical IncY replicons from E. coli (MW077912.1; 99.99% nucleotide identify to the full length) were identified in the NCBI database and highly similar elements were also identified in a Klebsiella pneumoniae (CP132612.1; 100% identity, 91% coverage) and several Salmonella enterica isolates (e.g., CP173313.1; 100% identity, 94% coverage), Notably, a highly similar IncY- blaCTX-M-15 replicon was also found in an E. coli isolate from the same farm in our previous study [23], suggesting recent transfer between species. Conversely, an IncY-blaCTX-M-55 from another E. coli in that study did not carry conjugation genes, highlighting the functional diversity within this family of mobile elements.
Assessment of plasmid similarity using the presence and absence of genes shows a good correlation between clusters and replicon types among isolates recovered from dairy cattle and turkey, and also with ESC-resistance plasmids identified in E. coli from the same dairy manure samples [23] (Fig 3). IncF plasmids were dominant and generally clustered according to their replicon subtypes, with some exceptions, which likely result from the high heterogenicity in this family of plasmids. Plasmids from the turkey samples remained distinct from those of dairy manure, with the exception of 328-1b, which was the only IncFIB(K)/FII isolated from dairy manure. Notably, one IncFIB plasmid was found at the root of the branch containing all of the IncY replicons. An alignment of these plasmids revealed that the conjugation genes associated with the IncY plasmids had a high similarity to this IncFIB plasmid. Further alignment with a non-conjugative IncY plasmid (CP117017.1) confirmed homology from the replication gene to the antibiotic resistance followed by the addition of the conjugation module in our plasmids (Fig 4)
Phenetic tree based on presence and absence of 2,064 genes, including core genes (n = 330) and cloud genes (n =1734), among plasmids carrying blaCTX-M in K. pneumoniae and E. coli from dairy manure (boldface) and turkey feces (red stars). All isolates were recovered from sources in Ontario, Canada with the exception of isolates from turkey feces in British Columbia (17.1 and 17.3) and Quebec (69.2, 70.3, 71.2, 72.2 and 142.2).
Alignment of one representative blaCTX-M-15 IncY plasmid from the dairy isolates aligned to a blaCTX-M-15 IncF plasmid from E. coli in the same samples and a non-conjugative blaCTX-M-15 IncY plasmid (CP117017.1).The IncY plasmid from our study is highly similar to the non-conjugative IncY from the replication genes until the antibiotic resistance region and then contains conjugation genes with high similarity to the IncFIB plasmid.
The IncFII-CTX-M-15 plasmids were among the most frequent ESC-resistance replicons of the plasmids recovered from Klebsiella spp. in this study (n = 10). They were all carried by isolates recovered from farm seven, spanning the whole one-year sampling period, from December 2018 to October 2019. Based on core gene SNP analysis, these plasmids formed two main subclusters, with the exception of 261-1b (ST219) which was recovered in heat-treated compost (S2A Fig). Isolate 261-1b had 30–32 SNPs compared to the other isolates, which differed amongst themselves by only 0–2 SNPs. Clustering did not show any clear correlation with time or treatment stage. Few rearrangements were identified among the IncFII plasmids (S2B Fig), and these were not specifically associated with the AMR genes present in these plasmids (S2C Fig). One IncFII plasmid carrying blaCTX-M-55 was identified in E. coli from the same sources; however, it was structurally different and unrelated to the IncFII/blaCTX-M-15 plasmids from Klebsiella spp. (S3 Fig and Fig 3).
IncI1 plasmids were also compared using core SNP analysis (62 genes; 0–649 SNPs in pairwise comparisons) and Mauve alignments (Fig 5). The two plasmids from K. pneumoniae clustered with two E. coli plasmids and all four were IncI1/ST3 plasmids carrying blaCTX-M-1. These IncI1-CTX-M-1 plasmids were not identical, but very similar both in their core gene sequences with 0 SNPs between the K. pneumoniae and 10 SNPs between the E. coli (Fig 5). Three of these were recovered from the same farm at different time periods but in samples from the same treatment stage (K. pneumoniae 269-2e, 269-3b and E. coli 254-3a). They were more similar to one another (0–3 SNPs) than to the plasmid recovered from an isolate from the other farm (10 SNPs). The IncI1-CTX-M-1 plasmid cluster was clearly distinct from the more diverse IncI-CMY-2 and IncI1-CTX-M-14 plasmids (Fig 5). Between K. pneumoniae and E. coli there were 3–593 SNPs.
Comparison of IncI1 plasmids (n = 13) from both K. pneumoniae and E. coli carrying blaCTX-M or blaCMY. A) Maximum likelihood-based on SNPs among 55,250 nucleotides among core plasmid loci and B) Mauve whole genome alignments of the whole plasmids.
Plasmid mobility
Both IncI1 and IncFII plasmids carrying blaCTX-M-1 and blaCTX-M-15, respectively, were transferred when using K. pneumoniae strains as donors and an E. coli recipient, with higher conjugation efficiency at 37˚C for all three donors compared to 30˚C (Table 5). Among these three donors, IncI1 plasmids transferred at approximately 100x higher rates than the IncFII plasmid. In contrast, when E. coli donors were used with a K. pneumoniae recipient, no transfer of blaCTX-M plasmids was detected in either broth or surface mating. Conjugation of IncY plasmids was also confirmed between E. coli strains, as well as from Klebsiella to E. coli, however transfer was found to occur at low frequency (2.67x10^-6^) and was only confirmed for surface mating at 37 ˚C.
Table 5: Transfer frequencies of blaCTX-M plasmids from K. pneumoniae donor strains to E. coli CV601gfpKanRRifR recipient strain using broth mating.
Discussion
ESC-resistance is a One Health issue, yet very little is known about the epidemiology of ESC-R K. pneumoniae in animals in Canada, despite their importance in both animal and human clinical infections [10,52]. There is potential for transmission of ESC-resistant strains and their AMR determinants between humans and animals, through close proximity or consumption of contaminated food via crop amendment or meat products [16,53–56]. Therefore, this study sought to further characterize ESC-R K. pneumoniae isolates from dairy manure in terms of their strain diversity, antimicrobial resistance, and potential contribution to the transmission dynamics of ESC-R plasmids in manure.
A large proportion (86.5%) of Klebsiella pneumoniae isolates carrying blaCMY or blaCTX-M were MDR, regardless of animal source. Similarly, in our parallel study, there were 79% MDR isolates among ESC-resistant E. coli from the same manure samples [23]. The majority of resistances observed in dairy manure isolates were to TET, STR, SXT and SUL, which are some of the most commonly prescribed antimicrobials in veterinary medicine, including for dairy cattle [57]. This followed again the same trend as in E. coli [23]. Twenty-six percent of the Klebsiella spp. from dairy cattle showed an intermediate susceptibility to CIP (1% in E. coli), but full resistances to CIP, CHL, KAN and GEN were low and in similar ranges in Klebsiella spp. and E. coli [23]. All ESC-R Klebsiella spp. isolates were susceptible to ETP and no resistances were identified to other critically important antimicrobials for human medicine.
The majority of isolates from this study were from a single farm and there were ESC-resistant isolates recovered at every manure treatment stage. In contrast, recovery on the other farms was sporadic and no ESC-resistant K. pneumoniae were recovered at all from two of the six farms investigated [22]. This suggests that contrary to the situation with E. coli, K. pneumoniae carrying blaCTX-M may not be endemic to all farms or are more challenging to detect using our current methods. Overall, due to the low recovery rate of K. pneumoniae and K. quasipneumoniae isolates, it is difficult to assess the effect manure treatments may have on their prevalence and more extensive quantitative analyses would be needed for this.
In keeping with the high K. pneumoniae strain diversity observed in dairy manure (30 STs), only two STs were found on more than one farm, ST3369 and ST2159. Both were found on the largest farm investigated (farm seven) and one additional farm. Additionally, ST7679 and ST1966 were recovered at different treatment stages, ranging from raw to environmentally applied manure and at different sampling times. In both cases, isolates were closely related but not identical suggesting that these strains may remain in the system over extended periods of time. Further studies would be needed to assess whether these strains may possess adaptive features for the manure environment. In addition, eleven K. quasipneumoniae isolates were recovered from farm seven at three different treatment stages, with the majority found in the raw manure. This may indicate recurrent entrance into the system and limited success at surviving the conditions encountered during manure treatment. Comparing the dairy manure strains with other animal sources, there was extensive strain and plasmid diversity within animal sources and no overlaps at the clonal level between different animal sources in this study. K. pneumoniae strains frequently associated with human infections globally include ST11, ST13, ST14, ST15, ST37, ST147, ST258, ST307, ST405 or ST512 [1,58–62]. Of these, only two were identified in our study (ST37 (raw manure) and ST405 (anaerobically digested manure)). ST37 is suspected to have spread globally through importation and trade and is considered a highly virulent lineage [63]. The low frequency of these K. pneumoniae strains in dairy cattle manure is reassuring but requires future surveillance.
Contrary to the situation in E. coli [23] only a small number of the ESC-resistant isolates carried a blaCMY gene and most had a blaCTX-M gene. Three blaCTX-M variants were identified and all belonged to enzyme group one [64]. The most common variant was blaCTX-M-15, which is known to be widespread globally in K. pneumoniae from both human and veterinary clinical infections [65–69]. Similar to what has been observed in E. coli from the same manure samples, the blaCTX-M-15 was often carried on the same plasmid as blaTEM-1 [23,49]. The overwhelming prevalence of blaCTX-M-15 and the infrequent recovery of blaCTX-M-1 and blaCTX-M-32 suggests a low diversity of blaCTX-M variants in K. pneumoniae from dairy manure. This seems to contrast with the situation in E. coli in which seven blaCTX-M variants were identified in our parallel study of these same samples, and blaCTX-M-15 was found in only 55% of cases [23]. K. pneumoniae intrinsically carry blaSHV variants, which are typically the non-ESBL variants blaSHV-1 and blaSHV-11, [70,71]. However, in rare cases they can carry blaSHV-27, which is variant documented to have an ESBL phenotype. Four isolates of two unrelated STs from farm seven were identified carrying blaSHV-27 in this study, though the ESBL phenotype of this variant could not be confirmed due to co-occurrence with CTX-M genes. The prevalence of ESBL SHV variants appears to be low currently, but monitoring may be warranted.
The majority of blaCTX-M genes were carried on plasmids. Only two isolates from dairy manure and the two dog isolates carried their blaCTX-M-15 on the chromosome. In each case, the chromosomal integration was likely due to mobilization through the action of ISEcp1. This insertion sequence is frequently associated with the mobilization of blaCTX-M-15 [72] and our findings are therefore not entirely surprising.
The majority of plasmids carrying blaCTX-M-15 were replicon type IncF and very diverse (Fig 3). This is similar to what is found in E. coli from the same manure samples, with highly diverse IncF plasmids carrying blaCTX-M-15 and blaCTX-M-55 [23]. Very similar IncFII plasmids were found in multiple genetically unrelated Klebsiella spp. strains in one of the farms investigated (Table 4; Fig S2), suggesting that these plasmids may be highly mobile within Klebsiella strains in the farm and manure treatment environments. In contrast, only one IncFII plasmid (carrying blaCTX-M-55) was identified in E. coli from the same dairy manure samples [23]. It was structurally different than the IncFII plasmids carrying blaCTX-M-15 in Klebsiella spp. This may indicate differences in the dynamics among IncF subtypes for Klebsiella spp. in general, or specifically for Klebsiella strains within manure. E. coli and Klebsiella spp. commonly carry different IncFII replicon alleles [73], which may be a factor in the dissemination dynamics observed.
Only two genetically related but not identical isolates from the same farm (Fig 1) were found to carry blaCTX-M-1. It was located on an IncI1/ST3 “epidemic” plasmid [74] and a similar plasmid was also detected in E. coli from the same manure samples [23]. The IncI1 plasmid tested transferred at rates hundreds to thousands time higher than the IncFII plasmids. This plasmid may have recently been introduced to the dairy cattle environment in Canada or Ontario, as suggested by its low frequency in both K. pneumoniae and E. coli. Its high level of transferability and common presence in other commodities such as poultry in Canada [46] suggest a need for further monitoring of its spread in dairy cattle and manure. Conjugation rates for both IncI1 and IncFII plasmids from K. pneumoniae to E. coli were higher at increased mesophilic temperatures, 37˚C, similar to those used on the dairy farms during anaerobic digestion. Conjugative transfer was successfully achieved between K. pneumoniae and E. coli, but not between E. coli and K. pneumoniae for any of the plasmid types tested. However, only one K. pneumoniae strain was tested as a recipient and the lack of transfer may be related to characteristics specific to this strain rather than to a general inability to transfer from E. coli to K. pneumoniae.
The high similarity of IncY replicons supported the transmission of blaCTX-M-15 plasmids between E. coli, K. pneumoniae and K. quasipneumoniae., which was further corroborated by our conjugation experiments demonstrating transfer from K. pneumoniae to E. coli. Due to the recent description of K. quasipneumoniae, little is known about its role in ESC-resistance and transmission [75]. However, K. quasipneumonaie carrying blaCTX-M-2 among other AMR genes, has been recovered in human sewage in Japan, and was capable of transferring blaCTX-M-harbouring IncA/C plasmids to E. coli [76]. Our findings may indicate that K. quasipneumoniae is also involved in the persistence and transmission of ESC-resistance plasmids in the manure environment. This information further stresses the diversity of potential hosts for resistance plasmids. The occurrence of conjugative IncY plasmids carrying multiple resistance genes highlights the need for more research into the role of phage-plasmids and similar MGEs in the evolution of antimicrobial resistance [51,77].
Conclusions
Although diverse strains were identified, ESC-resistant K. pneumoniae were not recovered from every farm, and only sporadically in the positive ones. Thus, these bacteria may play a small role in the broad dissemination of ESC-resistance. However, under specific circumstances, as found in one of the farms under investigation from which the majority of isolates originated, K. pneumoniae could act as reservoirs of transmission within local bacterial populations. The successful transfer of blaCTX-M plasmids between K. pneumoniae and E. coli as well as the demonstration in silico of plasmid similarities between K. pneumoniae and K. quasipneumoniae further supports the need to investigate not only E. coli, but also other members of the Enterobacterales, in attempts to better understand the dynamics and epidemiology of ESC-resistance in manure.
Supporting information
S1 TableTemporary multi-locus sequence typing identifications for K. pneumoniae isolates with novel alleles or novel allelic configurations.(DOCX)
S2 TableMulti-locus sequence typing (MLST) identifications for K. quasipneumoniae isolates based on the MLST scheme for K. pneumoniae to enable strain differentiation.(DOCX)
S1 FigPhylogenetic maximum likelihood SNP analysis (A) on IncY-blaCTX-M-15 plasmids using core genes (*n *= 73) with mauve alignments (B) and annotated plasmid segment carrying AMR genes (C).Plasmids were from K. pneumoniae and K. quasipneumoniae recovered from dairy manure. Plasmid names are colour coded (A) based on manure process in which they were recovered, raw manure (red), digestate with solids (green), digestate without solids (orange), dewatered (blue). All plasmids are harboured in isolates from farm seven, except for 246-3a.(DOCX)
S2 FigPhylogenetic maximum likelihood SNP analysis (A) on IncFII-blaCTX-M-15 plasmids using core genes (n = 121) with mauve alignments (B) and annotated plasmid segment carrying AMR genes (C).All plasmids are harboured in K. pneumoniae recovered from dairy manure on farm seven. Nodes are colour coded (A) based on manure process in which they were recovered, raw manure (red), digestate with solids (orange), digestate without solids (green), dewatered (blue) and heat-treated compost (purple).(DOCX)
S3 FigComparison of IncFII plasmids from one E. coli (259-1a) and K. pneumoniae (n = 10) recovered from dairy manure using mauve alignments.(DOCX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1De Oliveira DMP, Forde BM, Kidd TJ, Harris PNA, Schembri MA, Beatson SA, et al. Antimicrobial resistance in ESKAPE pathogens. Clin Microbiol Rev. 2020;33(3):e 00181-19. doi: 10.1128/CMR.00181-19 32404435 PMC 7227449 · doi ↗ · pubmed ↗
- 2Meatherall BL, Gregson D, Ross T, Pitout JDD, Laupland KB. Incidence, risk factors, and outcomes of Klebsiella pneumoniae bacteremia. Am J Med. 2009;122(9):866–73. doi: 10.1016/j.amjmed.2009.03.034 19699383 · doi ↗ · pubmed ↗
- 3Vading M, Nauclér P, Kalin M, Giske CG. Invasive infection caused by Klebsiella pneumoniae is a disease affecting patients with high comorbidity and associated with high long-term mortality. P Lo S ONE. 2018;13(4):e 0195258. doi: 10.1371/journal.pone.0195258 PMC 588918329624618 · doi ↗ · pubmed ↗
- 4Zhanel GG, Adam HJ, Baxter MR, Fuller J, Nichol KA, Denisuik AJ, et al. 42936 pathogens from Canadian hospitals: 10 years of results (2007-16) from the CANWARD surveillance study. J Antimicrob Chemother. 2019;74(Suppl 4):iv 5–21. doi: 10.1093/jac/dkz 283 31505641 · doi ↗ · pubmed ↗
- 5MasséJ, Dufour S, Archambault M. Characterization of Klebsiella isolates obtained from clinical mastitis cases in dairy cattle. J Dairy Sci. 2020;103(4):3392–400. doi: 10.3168/jds.2019-17324 32089315 · doi ↗ · pubmed ↗
- 6Pepin-Puget L, El Garch F, Bertrand X, Valot B, Hocquet D. Genome analysis of enterobacteriaceae with non-wild type susceptibility to third-generation cephalosporins recovered from diseased dogs and cats in Europe. Vet Microbiol. 2020;242:108601. doi: 10.1016/j.vetmic.2020.108601 32122604 · doi ↗ · pubmed ↗
- 7Vo ATT, van Duijkeren E, Fluit AC, Gaastra W. Characteristics of extended-spectrum cephalosporin-resistant Escherichia coli and Klebsiella pneumoniae isolates from horses. Vet Microbiol. 2007;124(3–4):248–55. doi: 10.1016/j.vetmic.2007.04.027 17521833 · doi ↗ · pubmed ↗
- 8Jensen AN, Storm C, Forslund A, Baggesen DL, Dalsgaard A. Escherichia coli contamination of lettuce grown in soils amended with animal slurry. J Food Prot. 2013;76(7):1137–44. doi: 10.4315/0362-028X.JFP-13-011 23834787 · doi ↗ · pubmed ↗