Enhancing Cryopreservation Efficiency in Populus davidiana × P. tremuloides Shoot Tips: Optimization of Vitrification Protocols and Mechanistic Insights into Flavonoid-Mediated Stress Adaptation
Panke Yang, Zelin Li, Yu Qi, Yuandong Ma, Chunming Li, Maolan Liu, Wenjun Ma, Hui Bai, Huanzhen Liu

TL;DR
This study improves cryopreservation in hybrid poplar by optimizing protocols and uncovering how flavonoids help plants survive freezing stress.
Contribution
The study introduces optimized cryopreservation protocols and reveals the role of flavonoid biosynthesis in stress adaptation during cryopreservation.
Findings
Optimized vitrification protocols achieved 85.91% and 79.70% regeneration rates with genetic stability.
Flavonoid biosynthesis pathway was robustly activated during osmotic dehydration, with upregulated PAL and CHS genes and quercetin accumulation.
Stress adaptation involves a coordinated defense network linking flavonoid biosynthesis, hormone signaling, and antioxidant systems.
Abstract
Cryopreservation is vital for conserving the elite germplasm of the hybrid poplar Populus davidiana × P. tremuloides, which is difficult to propagate conventionally. This study established optimized vitrification and encapsulation–vitrification protocols, achieving high regeneration rates of 85.91% and 79.70%, respectively, with confirmed genetic stability. The process induced oxidative stress, altering markers (MDA, H2O2) and antioxidant enzyme activities (SOD, POD, CAT). Integrated transcriptomic and metabolomic analysis of key steps—preculture/loading (DLA) and osmotic dehydration (DLB)—revealed extensive stress-responsive reprogramming. A central finding was the robust activation of the flavonoid biosynthesis pathway during DLB, marked by upregulation of key genes (PAL, CHS) and accumulation of flavonols (e.g., quercetin). This response was linked to hormone signaling and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
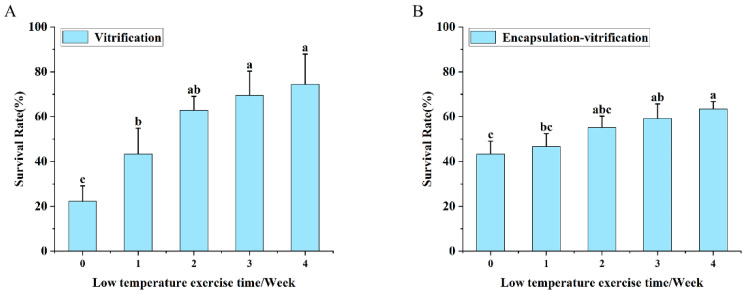 Figure 1
Figure 1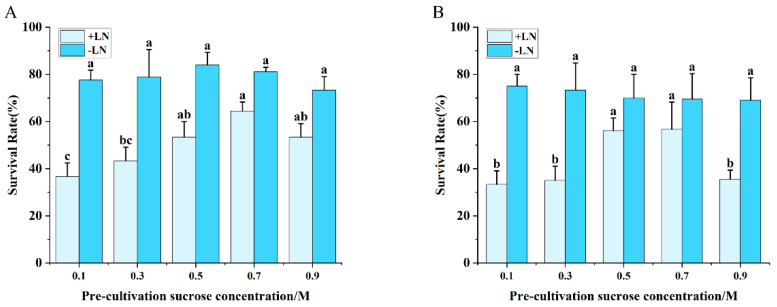 Figure 2
Figure 2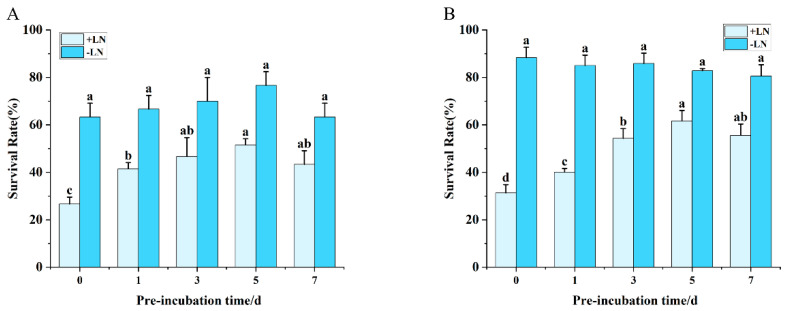 Figure 3
Figure 3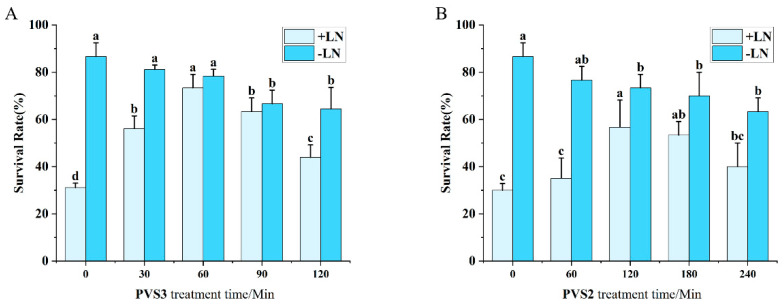 Figure 4
Figure 4 Figure 5
Figure 5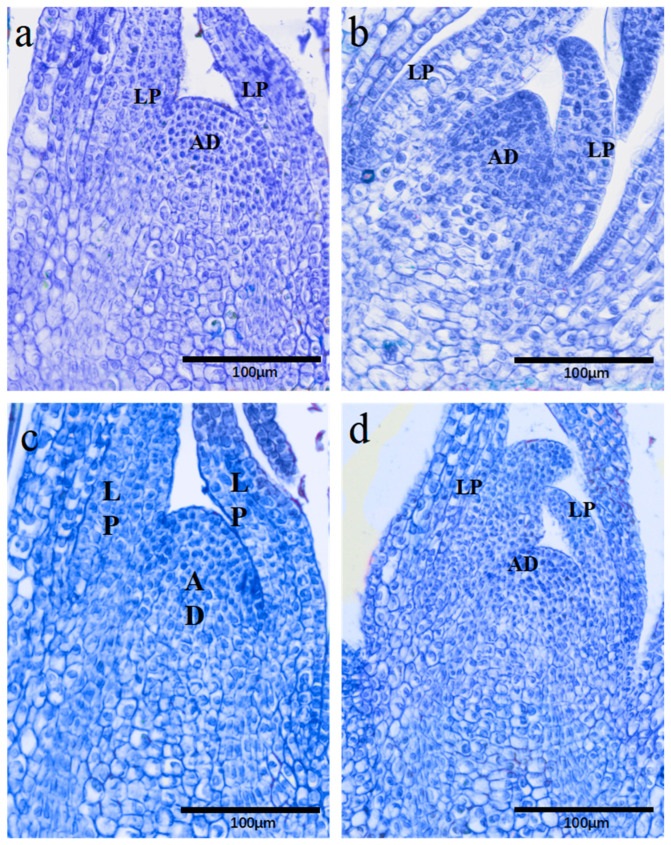 Figure 6
Figure 6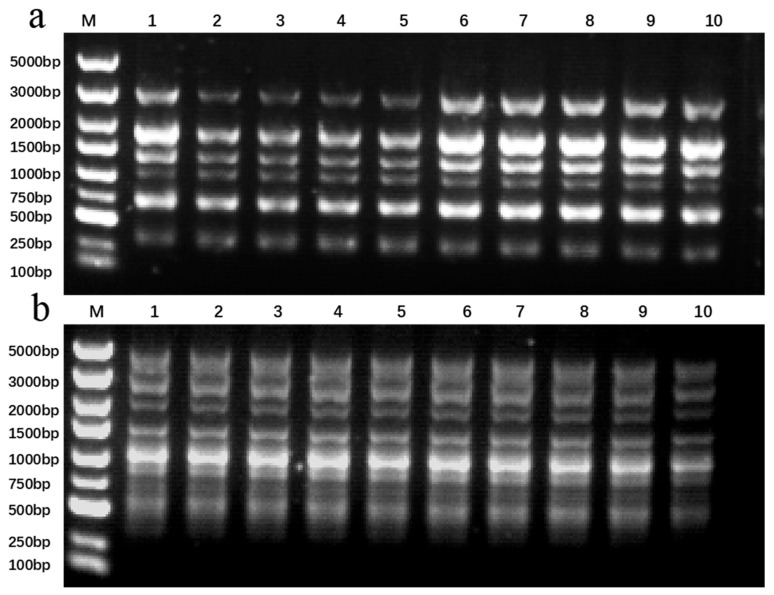 Figure 7
Figure 7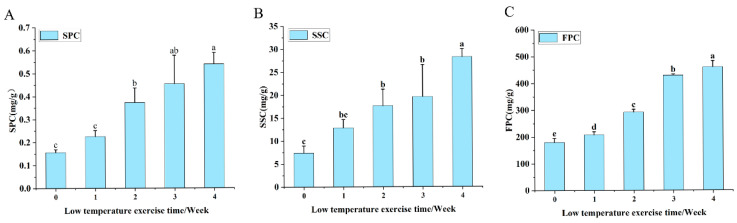 Figure 8
Figure 8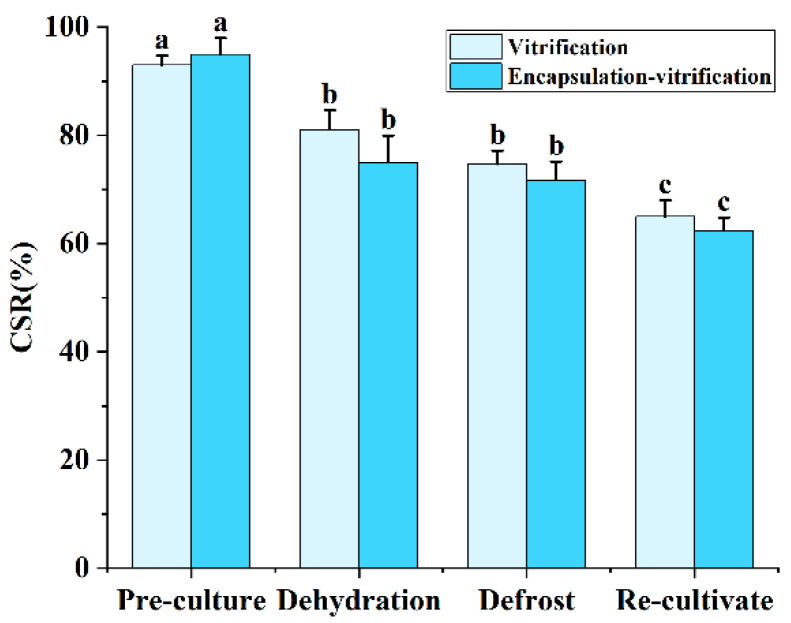 Figure 9
Figure 9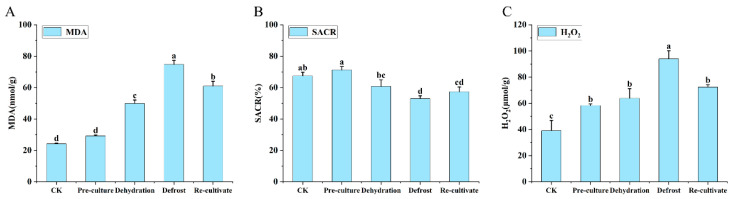 Figure 10
Figure 10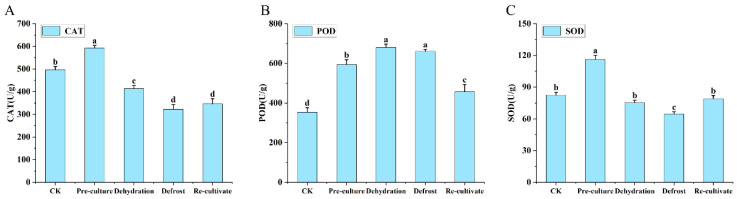 Figure 11
Figure 11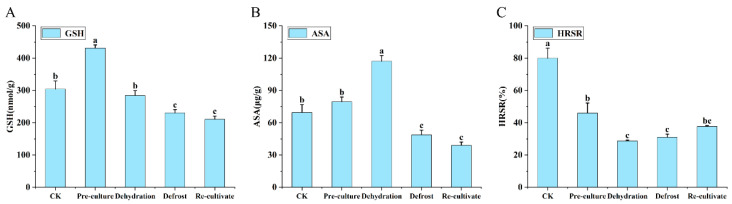 Figure 12
Figure 12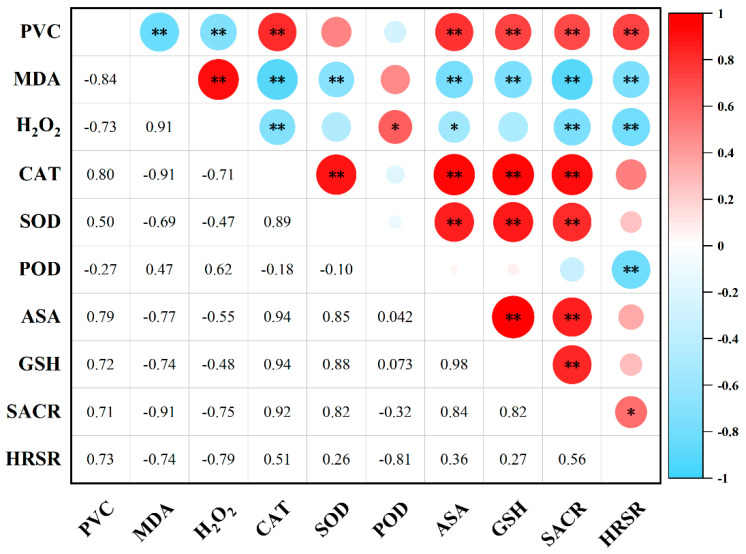 Figure 13
Figure 13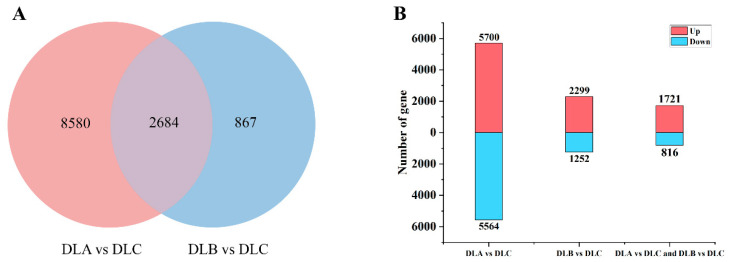 Figure 14
Figure 14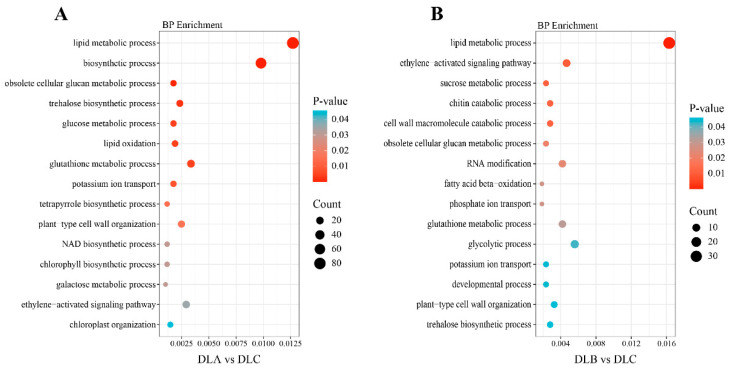 Figure 15
Figure 15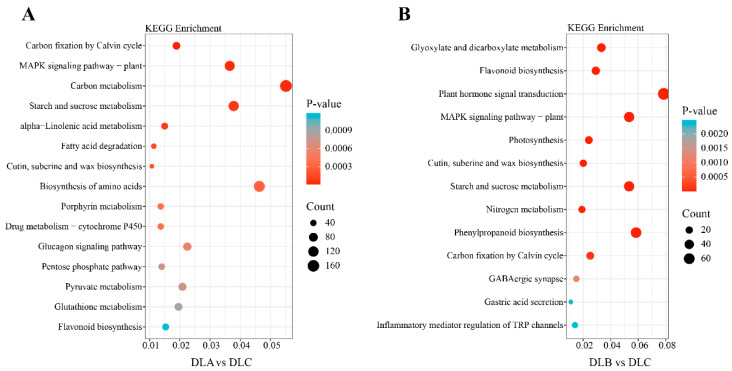 Figure 16
Figure 16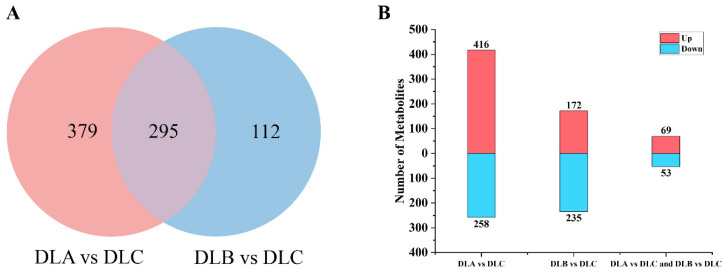 Figure 17
Figure 17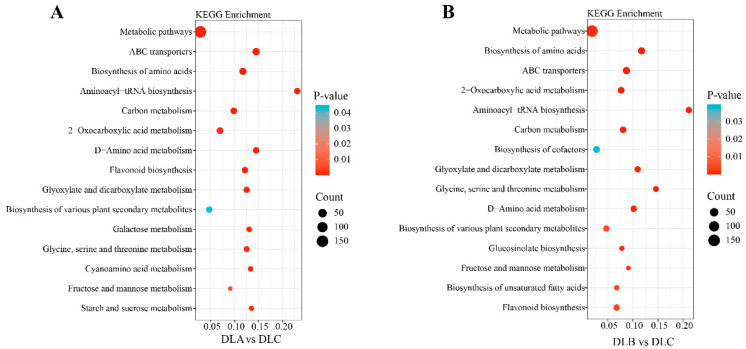 Figure 18
Figure 18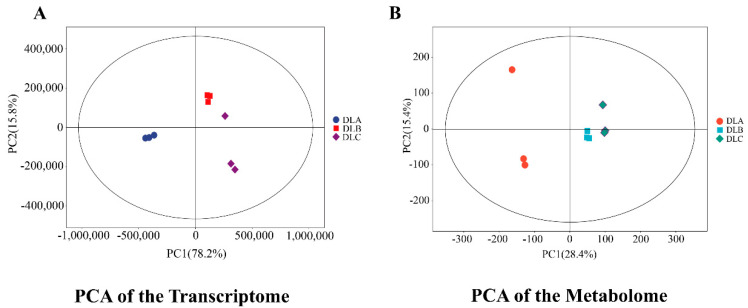 Figure 19
Figure 19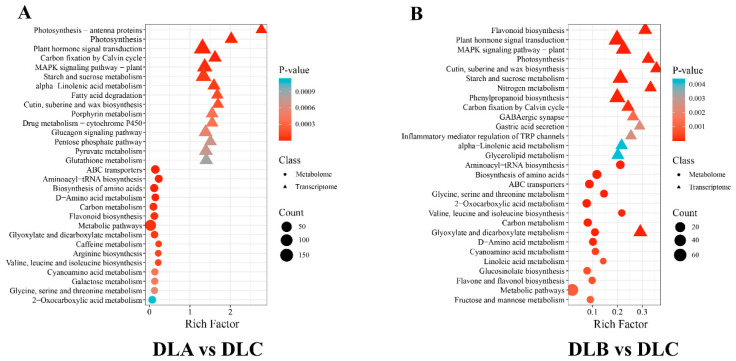 Figure 20
Figure 20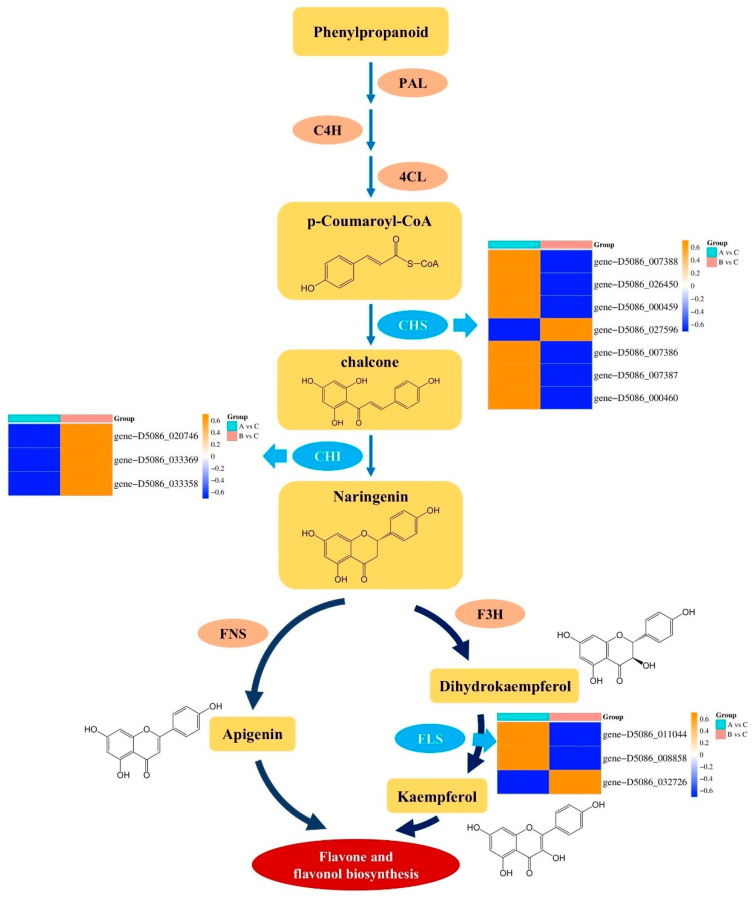 Figure 21
Figure 21- —National Key Research and Development Program of China during the 14th Five-Year Plan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant tissue culture and regeneration · Seed Germination and Physiology · Plant Molecular Biology Research
1. Introduction
Populus spp. serve as economically valuable timber species and ecologically important protective trees. The sustainable preservation of their superior germplasm resources constitutes a fundamental prerequisite for the advancement of forestry [1]. The hybrid Populus davidiana × P. tremuloides, combining the genetic backgrounds of Chinese and American aspen, displays excellent adaptability and fast growth, indicating high breeding potential. Nevertheless, the commercial utilization and preservation of its superior genotypes are critically limited by the inherently low rooting efficiency during cutting-based vegetative propagation [2]. In this context, cryopreservation represents a pivotal strategy to overcome this bottleneck, enabling the long-term and stable conservation of germplasm resources.
In plant cryopreservation, vitrification-based methods—specifically the vitrification method and the encapsulation–vitrification method—have been established as key techniques for achieving high survival and regeneration rates [3]. The vitrification method involves pretreating plant shoot tips with a specific vitrification solution followed by rapid cooling to induce a vitrified state, thereby enabling long-term genetic storage [4]. The encapsulation–vitrification method combines vitrification with encapsulation; before loading, shoot tips are mixed with sodium alginate solution and embedded, after which the beads are permeated with vitrification solution prior to rapid freezing [5]. To improve permeation of cryoprotectants across the cell membrane and to enhance tolerance to the dehydration stress imposed by vitrification solutions, a pretreatment step known as “loading” is commonly applied [6]. This process reduces tissue water content, mitigates injury from abrupt osmotic shifts, and increases intracellular levels of protective compounds [7]. In some cases, however, loading alone is insufficient to confer full tolerance, making preculture necessary before proceeding [8]. Osmotic dehydration represents the critical phase for achieving glass transition [9]. By using high-concentration vitrification solutions such as PVS3, the cytoplasm is further concentrated to a critical threshold, markedly raising intracellular viscosity and suppressing ice nucleation. This allows cells to form a stable glassy state rather than lethal ice crystals during freezing [10]. Finally, ultra-rapid freezing in liquid nitrogen instantly solidifies the dehydrated cytoplasm into an amorphous glass [11], thereby completely circumventing the structural damage caused by ice crystallization. Thus, the efficacy of both cryopreservation techniques relies on three tightly linked core steps: preculture/loading, osmotic dehydration, and liquid-nitrogen freezing [12].
Although the vitrification technique is well-established, its crucial pretreatment phase—spanning from loading with low-concentration protectants to exposure to the high-concentration PVS2 solution—concurrently primes the tissue for vitrification yet progressively subjects it to physiological stress [13]. The preculture/loading step, conducted in a low-toxicity environment to equilibrate osmotic pressure, induces mild water and preliminary oxidative stresses, thereby priming the material for subsequent PVS2 exposure [14]. The PVS2 treatment itself induces severe cellular dehydration via its hypertonic environment [15], while its chemical constituents directly disrupt metabolic homeostasis. This synergy disrupts reactive oxygen species (ROS) metabolism, leading to oxidative damage manifestations such as membrane lipid peroxidation, protein denaturation, and nucleic acid impairment [16].
To mitigate the aforementioned stresses, plant cells maintain intracellular redox homeostasis through an orchestrated coordination of enzymatic antioxidants [17] (e.g., SOD, CAT, POD, APX) and non-enzymatic antioxidants [18] (e.g., GSH, AsA) [19]. The efficacy of this integrated antioxidant system is a decisive factor for cellular viability under such stressful conditions [20]. Consequently, the physiological state and survival potential of cells prior to cryogenic exposure are fundamentally governed by their comprehensive resilience and adaptive capacity to the multi-step pretreatment regimen [21]. Presently, however, a systematic analysis is lacking regarding how the antioxidant system regulates the dynamic equilibrium between cumulative damage and stress adaptation throughout the continuous loading-to-osmotic treatment process.
Therefore, using shoot tips of Populus davidiana × P. tremuloides as experimental material in a vitrification-based cryopreservation system, this study systematically compared the survival and regeneration performance of materials after each procedural step, along with conducting related physiological and biochemical assays. Transcriptomic and metabolomic analyses were conducted on three critical stages—preculture/loading (DLA), osmotic dehydration (DLB), and liquid nitrogen freezing (DLC)—to comprehensively elucidate the dynamic interaction networks underlying cellular damage and protection, spanning from stress acclimation through deep freezing. The findings will not only clarify the mechanisms of stress-induced cross-tolerance during early procedural stages but also provide theoretical support for optimizing poplar germplasm cryopreservation protocols, while advancing the understanding of plant adaptive responses under combined-stress conditions.
2. Results
2.1. Effect of Cold Acclimation Duration on Shoot Tip Cryopreservation in Populus davidiana × P. tremuloides
Under the vitrification (Figure 1A) and encapsulation–vitrification (Figure 1B) cryopreservation methods, the survival rate of shoot tips exhibited a significant increase with prolonged cold acclimation duration. The highest survival rates, reaching 74.44% and 63.43%, respectively, were achieved after a 4-week acclimation period.
2.2. Effect of Preculture on Shoot Tip Survival Following Cryopreservation
In both cryopreservation methods, shoot tip survival exhibited a concentration-dependent increase with elevated sucrose levels in the preculture medium. The vitrification method (Figure 2A) attained a maximal survival rate of 64.00% at 0.7 mol/L sucrose, whereas the encapsulation–vitrification method (Figure 2B) achieved its peak survival of 56.11% at 0.5 mol/L.
The survival rate of shoot tips during preculture exhibited a distinct bell-shaped response over time (Figure 3). For both cryopreservation techniques, survival peaked on the 5th day of preculture, reaching 51.52% with the vitrification method (Figure 3A) and 61.67% with the encapsulation–vitrification method (Figure 3B), after which a progressive decline was observed. No significant differences were detected among the parallel controls (−LN) across treatments.
This pattern can be attributed to the dual role of preculture. While optimal conditions enhance desiccation tolerance, prolonged exposure or excessive sucrose concentrations exacerbate osmotic stress, becoming detrimental to survival. Therefore, identifying the critical balance in sucrose concentration and preculture duration is essential to moderately reduce tissue water content, bolster cryotolerance, and ultimately maximize post-thaw regeneration.
2.3. Effect of Vitrification Solution and Exposure Duration on Shoot Tip Survival During Cryopreservation
Exposure to vitrification solutions is a critical protective step prior to cryopreservation, serving to reduce intracellular free water content. In this study, PVS2 and PVS3 were evaluated for their effects on shoot tip survival. Under the vitrification protocol (Figure 4A), survival exhibited a biphasic effect in response to PVS3 exposure time, peaking at 73.33% after 60 min before declining. In contrast, the encapsulation–vitrification method (Figure 4B) achieved optimal survival (56.67%) following 120 min of PVS2 treatment. In non-frozen controls (–LN), prolonged exposure to both solutions progressively reduced survival, consistent with cumulative osmotic and chemical injury. Notably, PVS2 induced a more pronounced toxicity compared to PVS3, as reflected by the steeper decline in control survival over time.
2.4. Establishment of a Shoot Tip Cryopreservation System for Populus davidiana × P. tremuloides
Based on the optimized parameters established in the preceding experiments, standardized protocols for the cryopreservation of in vitro shoot tips of Populus davidiana × P. tremuloides were developed (Figure 5).
Vitrification-based cryopreservation: Shoot tips (2 mm in length) excised from 4-week cold-acclimated plantlets (Figure 5a) were precultured in 0.7 mol/L sucrose medium at 4 °C for 5 days in darkness (Figure 5b). Subsequently, samples were subjected to loading treatment at room temperature for 20 min, followed by dehydration with PVS3 for 60 min (Figure 5d), and then plunged into liquid nitrogen for 1 h (Figure 5f). For rewarming, samples were immersed in a 40 °C water bath for 1 min, drained, and rinsed with recovery medium for 20 min. Tissues were then incubated in darkness for 7 days before transfer to standard light conditions.
Encapsulation–vitrification cryopreservation: Shoot tips precultured in 0.5 mol/L sucrose medium at 4 °C for 5 days in darkness were encapsulated in alginate/CaCl_2_ beads (Figure 5c). After loading for 20 min, samples were treated with PVS2 on ice for 120 min (Figure 5e), transferred to liquid nitrogen for 1 h (Figure 5f), rewarmed in a 40 °C water bath for 3 min, and rinsed with loading solution for 20 min before being returned to culture conditions identical to those used in the vitrification protocol.
Viable shoot tips exhibited dark pink pigmentation (Figure 5h), whereas non-viable ones turned brown (Figure 5g). After 4 weeks of culture, regenerated plantlets were obtained (Figure 5i). The optimized vitrification and encapsulation–vitrification protocols yielded regeneration rates of 85.91% and 79.70%, respectively.
2.5. Histological Analysis of Cell Survival in Cryopreserved Populus davidiana × P. tremuloides Shoot Tips
Histological analysis of cryopreserved shoot tips revealed that surviving cells were predominantly located in the apical meristem (layers 1–6) and the primordia of the first three leaves. These cells exhibited intensely stained cytoplasm and well-preserved cellular integrity (Figure 6a). In contrast, non-frozen control shoot tips directly immersed in liquid nitrogen (negative control) displayed lightly stained cells with ruptured membranes and severe structural disruption (Figure 6b). Shoot tips processed with both cryopreservation methods showed cytological features comparable to those of viable cells, including densely stained cytoplasm, intact membranes, and preserved nuclear structure, with no evident cryo-damage (Figure 6c,d).
2.6. Assessment of Genetic Stability in Regenerated Plants Following Cryopreservation
Plantlets regenerated following cryopreservation exhibited comparable performance to the non-cryopreserved controls in terms of rooting and vegetative growth (Table 1), indicating no phenotypic alterations induced by the cryopreservation process. Genetic stability was further confirmed by ISSR analysis, in which eight primers generated 50 reproducible bands (4–9 bands per primer; mean: 6.25). Both cryopreservation methods produced monomorphic banding patterns identical to the control across all samples (Figure 7; Table 2), demonstrating the preservation of genetic integrity in Populus davidiana × P. tremuloides after cryopreservation.
2.7. Physiological and Biochemical Analysis of the Cryopreservation Process of Stem Tips of Populus davidiana × P. tremuloides
During the cold acclimation period, the contents of soluble proteins, soluble sugars, and free proline in Populus davidiana × P. tremuloides progressively increased with prolonged treatment duration (Figure 8). Relative to non-acclimated controls, the levels of these osmolytes showed respective increases of 70.37% (Figure 8A), 73.82% (Figure 8B), and 60.95% (Figure 8C). The accumulation of these osmoregulatory substances is associated with enhanced tolerance to low-temperature stress, thereby contributing to improved shoot tip survival following cryopreservation.
Longitudinal assessment of shoot tip cell viability throughout the cryopreservation process is summarized in Figure 9. Initial pretreatment resulted in high cell survival rates (93–95%) without evident injury. Following dehydration, viability declined to 81% in the vitrification group and 75% in the encapsulation–vitrification group. This divergence is attributable to the composition of PVS3, which contains only sucrose and glycerol without DMSO, thus exerting relatively lower cytotoxicity. A further decrease in survival was observed after liquid nitrogen exposure, though not significantly different from dehydration levels. After recovery culture, final survival rates reached 65% for vitrification and 62% for encapsulation–vitrification.
Throughout the cryopreservation process (Figure 10), malondialdehyde (MDA) concentration exhibited an initial increase followed by a decline, with all treatment groups maintaining levels significantly higher than those of the control (Figure 10A). The superoxide anion scavenging rate reached its maximum after preculture, decreased markedly during dehydration and thawing, and subsequently recovered gradually throughout the re-culture period (Figure 10B). Hydrogen peroxide (H_2_O_2_) concentration showed a pattern of rapid increase followed by a sharp decrease, with pronounced fluctuations observed specifically during the dehydration and thawing stages (Figure 10C).
Throughout the cryopreservation process (Figure 11), the three antioxidant enzymes exhibited divergent response patterns. CAT activity increased by 19.4% after pretreatment, declined to its nadir during thawing, and subsequently recovered slightly (Figure 11A). POD activity peaked following dehydration but decreased to 456 U/g after thawing, suggesting its role in early stress response and subsequent limitation due to cellular injury (Figure 11B). SOD activity displayed an overall decline initially, reaching the lowest level at thawing, followed by gradual recovery during re-culture (Figure 11C).
During cryopreservation (Figure 12), both reduced glutathione (GSH) and ascorbic acid (ASA) exhibited a biphasic pattern, characterized by an initial increase followed by a subsequent decrease. GSH concentration peaked during the preculture stage and progressively declined as the protocol advanced (Figure 12A). Similarly, ASA reached its maximum level after preculture, followed by a gradual reduction (Figure 12B). In contrast, the hydroxyl radical scavenging rate was highest in the control group, decreased significantly after preculture, reached its nadir during dehydration and thawing, and rebounded gradually upon re-culture (Figure 12C).
2.8. Correlation Analysis of Various Physiological Indicators During Cryopreservation
A significant dynamic correlation between oxidative damage and antioxidant defense was observed in Populus davidiana × P. tremuloides shoot tip cells during cryopreservation (Figure 13). Malondialdehyde (MDA) content showed significant negative correlations with catalase (CAT) and superoxide dismutase (SOD) activities, as well as with ascorbic acid (ASA) and reduced glutathione (GSH) levels (p < 0.01), indicating that severe oxidative damage suppresses antioxidant capacity. Hydrogen peroxide (H_2_O_2_) content was positively correlated with certain antioxidant components, potentially reflecting its role as a signaling molecule in early stress response activation. Strong positive correlation was observed between CAT and SOD activities (p < 0.01), demonstrating their synergistic action in reactive oxygen species elimination. Furthermore, ASA and GSH contents showed coordinated trends and were positively correlated with both superoxide anion and hydroxyl radical scavenging rates, highlighting the crucial role of non-enzymatic antioxidants in oxidative stress mitigation. Most importantly, post-thaw survival rate exhibited strong positive correlations with key antioxidant indicators including CAT, SOD, ASA, and GSH (p < 0.01), confirming that enhanced antioxidant defense capacity represents a key mechanism for improving cryopreservation efficiency.
2.9. Transcriptomics Analysis
2.9.1. Screening of Differentially Expressed Genes
Transcriptome analysis was performed on shoot tips of Populus davidiana × P. tremuloides subjected to three critical cryopreservation steps: preculture/loading (DLA), osmotic dehydration (DLB), and liquid nitrogen freezing (DLC) (Figure 14A,B). Under the threshold of|log_2_Fold Change| > 1 and PADJ < 0.05, 2684 common differentially expressed genes (DEGs) were identified. Considerable numbers of unique DEGs were detected among treatment groups, with the highest number specific to the DLC group, suggesting that osmotic dehydration exerted the most pronounced effect on transcriptional regulation. Statistical analysis of DEG numbers in paired comparisons (DLA vs. DLC, DLB vs. DLC) revealed a higher proportion of upregulated genes in the DLB vs. DLC group (Figure 14B), implying that osmotic dehydration may activate multiple stress-responsive pathways. In contrast, the DLC group, representing the liquid nitrogen freezing step, displayed relatively moderate transcriptomic alterations, indicating limited additional impact of freezing itself on gene expression. Both DLA and DLB treatments likely elicited extensive transcriptional reprogramming, presumably through physical stresses such as dehydration and osmotic perturbation. Among the identified DEGs, a substantial number are hypothesized to be associated with water stress, oxidative response, and cell wall remodeling, reflecting the adaptive mechanisms of Populus davidiana × P. tremuloides shoot tips during vitrification-based cryopreservation.
2.9.2. GO and KEGG Enrichment Analysis of Differentially Expressed Genes (DEGs)
GO Enrichment Analysis of DEGs
To elucidate the functional impacts of different treatments on shoot tip development in Populus davidiana × P. tremuloides, Gene Ontology (GO) enrichment analysis was performed on key differentially expressed genes (DEGs) from DLA vs. DLC and DLB vs. DLC comparisons. Enrichment covered three major GO categories—biological process (BP), cellular component (CC), and molecular function (MF)—with emphasis on BP terms. Significantly enriched terms were visualized via bubble plot (Figure 15).
DEGs from both groups were significantly enriched in metabolic, stress-responsive, and cellular organization processes. DLA predominantly activated osmotic regulation, energy metabolism, and antioxidant defense mechanisms. Enriched processes such as trehalose biosynthesis and glutathione metabolism align with plant strategies for accumulating compatible solutes and scavenging reactive oxygen species under osmotic stress. Activation of potassium ion transport and cell wall organization reflects requirements for ionic homeostasis and structural remodeling. Furthermore, enrichment in chlorophyll and tetrapyrrole biosynthesis suggests that loading influenced photosynthetic metabolism, potentially as an adaptive preculture response prior to subsequent stress.
Compared to DLA, DLB significantly enriched catabolic and cell wall remodeling processes, including chitin and cell wall macromolecule decomposition, indicative of cell wall degradation and restructuring under severe dehydration. Enrichment of fatty acid β-oxidation and glycolysis revealed a shift toward energy-generating catabolism to support stress responses. Concurrently, persistent enrichment in phosphate and potassium ion transport underscored the importance of ion homeostasis in dehydrating environments.
Notably, both comparisons showed enrichment in ethylene signaling and glutathione metabolism, identifying these as core mechanisms in stress adaptation. Enrichment of developmental processes further implied long-term effects of dehydration on growth and differentiation, consistent with plants modulating developmental programs under stress.
KEGG Pathway Enrichment Analysis of Differentially Expressed Genes
To systematically elucidate the biological pathways underlying transcriptomic differences between DLA vs. DLC and DLB vs. DLC, KEGG pathway enrichment analysis was performed on the differentially expressed genes. The results revealed significant enrichment of “Plant hormone signal transduction”, “Phenylpropanoid biosynthesis”, and “Starch and sucrose metabolism” pathways in both comparison groups (Figure 16). In terms of signaling and stress response, enrichment of the “MAPK signaling pathway-plant” and drug metabolism-cytochrome P450 pathways indicated that DLA may represent a physiological state involving active responses to environmental cues.
In the DLA vs. DLC comparison (Figure 16A), significant enrichment was observed in the flavonoid biosynthesis pathway and pathways related to multiple plant secondary metabolite biosynthesis, suggesting that loading treatment activated secondary metabolic systems in Populus davidiana × P. tremuloides shoot tips. The accumulation of flavonoids, as important antioxidants, may represent a protective strategy against osmotic stress.
In the DLB vs. DLC comparison (Figure 16B), enriched pathways partially overlapped with the DLA group but displayed distinct characteristics. A prominent feature of DLB was the strong enrichment of plant hormone signaling pathways, indicating central regulatory roles of hormonal regulation in physiological responses to osmotic dehydration. Similarly to DLA, enrichment of flavonoid biosynthesis and phenylpropanoid biosynthesis pathways demonstrated that DLB also activated secondary metabolism as a stress-coping mechanism.
2.10. Metabolome Analysis
2.10.1. Screening of Differentially Metabolites
To investigate the differences in metabolic responses between the two groups, we conducted metabolic profiling analysis on each of them and visualized the results (Figure 17). Through the Venn diagram (Figure 17A), it was found that 674 different metabolites were identified in DLA vs. DLC, among which 416 were upregulated and 258 were downregulated; while in DLB vs. DLC, 407 different metabolites were identified, with 172 upregulated and 235 downregulated. In the DLA vs. DLC experiment, the number of upregulated metabolites was significantly higher than that of downregulated metabolites, suggesting that the physiological state of DLA might be more inclined to activate or accumulate a series of metabolites. In sharp contrast, in the DLB vs. DLC experiment, the number of downregulated metabolites far exceeded that of upregulated metabolites. This result indicates that the treatment of DLB might have dominated a metabolic state characterized by inhibition of metabolite consumption or synthesis.
2.10.2. KEGG Pathway Enrichment Profiling of Metabolomes
KEGG pathway enrichment analysis was performed to elucidate the biological pathways associated with differential metabolites in DLA and DLB comparisons. The results revealed substantial similarity in core metabolic pathway regulation between both treatments. Significantly enriched pathways common to DLA and DLB included Metabolic pathways, Biosynthesis of amino acids, ABC transporters, Carbon metabolism, and 2-Oxocarboxylic acid metabolism. This enrichment pattern indicates that extensive remodeling of primary metabolism—particularly the synthesis and transformation of fundamental biomolecules such as amino acids and organic acids—represents a core metabolic feature shared by both treatments. The enrichment of ABC transporters suggests a critical role of transmembrane transport, potentially reflecting enhanced compartmentalization or excretion of toxic compounds under stress conditions.
In the DLA vs. DLC comparison (Figure 18A), specific pathway enrichment was observed for Flavonoid biosynthesis and Galactose metabolism. The activation of flavonoid biosynthesis, associated with plant antioxidant responses, aligns with transcriptomic findings indicating that loading treatment preferentially induces stress-adaptive mechanisms.
In contrast, the DLB vs. DLC comparison (Figure 18B) exhibited distinct enrichment in the Biosynthesis of cofactors, Biosynthesis of unsaturated fatty acids, and Glucosinolate biosynthesis. These results suggest that DLB employs a more refined metabolic strategy focused on maintaining redox homeostasis, modulating membrane lipid composition, and mobilizing specialized plant defense systems, reflecting a targeted adaptive response to osmotic dehydration.
2.11. Transcription and Metabolome Combined Analysis
2.11.1. Principal Component Analysis
To comprehensively evaluate the overall variations and grouping situations of DLA, DLB and DLC samples at the transcriptomic and metabolomic levels, we conducted principal component analysis (PCA) on the gene expression data and metabolite profile data, respectively. The results are shown in Figure 19.
The PCA of the transcriptome (Figure 19A) revealed that the variance contribution rates of the first principal component (PC1) and the second principal component (PC2) were 78.2% and 15.8%, respectively, and they collectively explained up to 94.0% of the total variation. Such a high degree of explanation indicates that PC1 and PC2 can adequately represent the overall characteristics of the dataset. More importantly, the three groups of samples, DLA, DLB, and DLC, showed clear and distinct clustering in the two-dimensional space. The PCA analysis of the metabolome (Figure 19B) showed that the variance contribution rates of PC1 and PC2 were 28.4% and 15.4% respectively, and the cumulative explanation rate (43.8%) was lower than that of the transcriptome. This reflects the complexity of the metabolite spectrum, with its variation driven by more dimensions. Nevertheless, we still observed a clear trend of sample separation. DLC was clearly distinguished from DLA and DLB along the PC1 axis, indicating that both treatments significantly altered the overall metabolite composition. Notably, although the separation degree between DLB and DLA in spatial distribution was not as obvious as that in the transcriptome, it could still be effectively distinguished. This indicates that although DLA and DLB caused different metabolic changes in direction, there might be overlap or intermediate states in some core metabolites, which is consistent with the results we observed in the differential metabolite Venn diagram.
2.11.2. KEGG Analysis for Jointly Regulating the Transcriptome and Metabolome
To systematically clarify the regulatory relationship between gene expression and metabolite accumulation, we conducted a combined KEGG pathway enrichment analysis of transcriptomics and metabolomics for DLA vs. DLC and DLB vs. DLC. This analysis integrated the changes in mRNA and metabolites at two levels, thereby enabling a more comprehensive revelation of the biological pathways that are simultaneously activated or inhibited under specific physiological conditions (Figure 20). The joint analysis revealed significant transcriptional and metabolic co-enrichment in multiple core metabolic and signaling pathways between DLA and DLC (Figure 20A). Firstly, in terms of energy and carbon assimilation, pathways such as Photosynthesis—antenna proteins, Photosynthesis, and Carbon fixation by Calvin cycle were strongly activated at the transcriptome level, while pathways such as Carbon metabolism and Pyruvate metabolism were simultaneously enriched at both the transcriptional and metabolic levels. This indicates that DLA has achieved coordinated enhancement from gene expression to metabolite levels in terms of light energy capture, carbon fixation, and downstream energy metabolism. Secondly, in terms of stress response and secondary metabolism, the MAPK signaling pathway in the transcriptome, glutathione metabolism, and the flavonoid biosynthesis pathway in the metabolome were all significantly enriched, forming a complete response chain from signal perception, antioxidation to defense substance synthesis. At the same time, the pathways of cutin, subcine and wax biosynthesis, and fatty acid degradation were enriched, consistent with the physiological needs of DLA in cell barrier reconstruction and lipid mobilization. This multi-omics co-enrichment pattern clearly depicts that DLA is in a high-energy-consuming and highly stressful physiological state.
Compared with DLA, the combined enriched pathway spectrum of DLB vs. DLC shows different regulatory focuses (Figure 20B). DLB also exhibits activation at the transcriptional level of “photosynthesis” and “Calvin cycle”, but its unique enriched pathways are more focused on the transformation and transportation of substances. For instance, the enrichment of Nitrogen metabolism suggests that DLB adjusts the assimilation and utilization of nitrogen, while the enrichment of Phenylpropanoid biosynthesis and Flavone and flavonoid biosynthesis indicates that the secondary metabolic flow is more specifically directed towards the synthesis of defense compounds. Particularly noteworthy is that Plant hormone signal transduction is significantly enriched in the transcriptome data of DLB, which is consistent with our previous findings in transcriptome analysis, once again emphasizing the core role of hormone-mediated precise regulation in the response mechanism of DLB. At the same time, the continuous enrichment of ABC transporters in the metabolomics data suggests that the transmembrane transport process is crucial for maintaining the metabolic homeostasis of DLB.
By comparing the combined enrichment pathways of DLA and DLB, it can be observed that the transcription and metabolic profiles of DLA jointly point to a pattern of acute stress and energy mobilization. Its regulatory network extensively activates multiple links ranging from signal transduction, photosynthesis to antioxidant and lipid metabolism. The regulation of DLB, on the other hand, appears more restrained and specific. Its core lies in strengthening photosynthesis while precisely regulating nitrogen metabolism, the synthesis and transportation of specific secondary metabolites through hormone signals, in order to achieve physiological balance and adaptation.
2.11.3. Characterization of Key Enzymatic Reactions and Transcriptional Regulation in the Flavonoid Biosynthesis Pathway
Flavonoids serve as crucial secondary metabolites enabling plants to withstand both biotic and abiotic stresses. Elucidating the molecular regulatory mechanisms underlying their biosynthetic pathway is essential for understanding plant stress resistance and secondary metabolic phenotypes [22]. In this study, through integrated transcriptomic and metabolomic analyses, we reconstructed the biosynthetic pathway from phenylpropanoids to flavonoids and flavonols (Figure 21). Furthermore, by correlating the expression patterns of differentially expressed genes, we clarified the functional roles of key enzymatic genes in the regulation of this pathway.
Phenylpropanoids serve as the initial substrates for flavonoid biosynthesis [23]. Through sequential catalysis by phenylalanine ammonia-lyase (PAL), cinnamate 4-hydroxylase (C4H), and 4-coumarate-CoA ligase (4CL), p-coumaroyl-CoA is generated [24]. This segment represents a critical branch of the phenylpropanoid pathway, supplying the direct precursor for flavonoid formation. Both enzymatic activity and gene expression levels of these enzymes fundamentally govern carbon flux allocation into flavonoid synthesis [25]. Under the catalysis of chalcone synthase (CHS), p-coumaroyl-CoA is converted into chalcone, which is subsequently isomerized to naringenin—the core flavonoid precursor [26]. The heatmap depicting CHS-associated genes (right panel) reveals that genes such as D5086_007388 and D5086_026450 exhibit significant differential expression in both DLA vs. DLC and DLB vs. DLC comparisons (expression change range: −0.6 to 0.6), indicating that transcriptional regulation of these CHS genes may directly modulate chalcone synthesis efficiency. Similarly, differential expression patterns of chalcone isomerase (CHI)-related genes (left panel, e.g., D5086_020746 and D5086_033369) further demonstrate that heterogeneity in CHI gene expression likely regulates the conversion of chalcone to naringenin, thereby influencing downstream flavonoid branching pathways.
Naringenin serves as a metabolic branch point leading to distinct flavonoid classes through two enzymatic routes [27]: (1) catalysis by flavone synthase (FNS) yielding apigenin (flavones); (2) conversion into dihydrokaempferol by flavanone 3-hydroxylase (F3H), followed by formation of kaempferol (flavonols) via flavonol synthase (FLS). The heatmap of FLS-associated genes (e.g., D5086_011044, D5086_008858) reveals distinct expression patterns across experimental groups, indicating that transcriptional regulation of FLS plays a critical role in flavonol biosynthesis. In contrast, FNS activity determines the production efficiency of flavones such as luteolin. This differential regulation at the branch point may shift the metabolic flux between flavones and flavonols, ultimately influencing stress-related phenotypes—including antioxidant capacity and UV protection—in plants [28].
3. Discussion
Cryopreservation serves as a critical method for the long-term and secure preservation of germplasm resources. It effectively maintains genetic stability and has been widely applied to various materials, including seeds and shoot tips. Furthermore, it demonstrates broad prospects in fields such as the conservation of transgenic materials, virus-free seedling production, and ecological restoration [29]. The success of cryopreservation hinges on whether plant materials can survive and recover after undergoing a series of intense stresses (sudden osmotic changes, dehydration, cold shock). Multi-omics data from this study clearly indicate that shoot-tip cells do not passively endure damage but instead actively initiate a multi-layered, highly coordinated adaptive program. This program begins with the rapid perception and transduction of stress signals. At the DLA stage, transcriptomic data show significant enrichment of the “MAPK signaling pathway—plant” pathway. This pathway serves as a central hub for converting diverse environmental stimuli into intracellular signals in plants [30] and has been demonstrated in species like Arabidopsis to be involved in responses to cold, drought, salt, and oxidative stress [31,32,33]. Its early activation indicates that the mild stress induced by DLA treatment is perceived by the cells, triggering downstream reprogramming of gene expression. At the DLB stage, the signaling network becomes more complex, with one of the most prominent features being the strong enrichment of the “Plant hormone signal transduction” pathway in the transcriptome. The data suggest that when confronted with stress induced by PVS2/PVS3 solutions, shoot-tip cells may precisely coordinate physiological responses by integrating multiple hormone signals [31,34]. This hormone-mediated precise regulation, differing from the more “generalized” MAPK signal initiation seen at the DLA stage, likely represents an optimized strategy for coping with extreme and sustained stress.
Driven by this signaling network, cells activate a series of programs to maintain internal homeostasis. Among these, maintaining redox balance is crucial. Physiological data from this study confirm the occurrence of oxidative stress (increases in MDA and H_2_O_2_), while multi-omics data reveal the comprehensive mobilization of the antioxidant defense system. This includes not only enzymatic systems like SOD, POD, and CAT but also non-enzymatic systems. Notably, the “Glutathione metabolism” pathway is enriched in the transcriptome at both DLA and DLB stages. Glutathione (GSH), as a core antioxidant and redox buffer, plays a key role in oxidative stress tolerance in Arabidopsis [31,35]. Its upregulation, occurring alongside the activation of the flavonoid biosynthesis pathway, helps build a multi-layered ROS scavenging system. Beyond countering oxidative damage, cells must directly address dehydration and cold challenges. Physiological data show that cold acclimation significantly increased the content of soluble sugars, soluble proteins, and free proline, consistent with reports of enhanced cold hardiness in many woody plants [36]. Transcriptomic data further support this adaptive process at the gene expression level, with the activation of pathways such as “Starch and sucrose metabolism” and “Amino acid biosynthesis.” The accumulation of these compatible solutes directly stabilizes cellular structures [37,38]. Additionally, the remodeling of energy metabolism is evident. The co-enrichment of pathways like “Carbon fixation” and “Carbon metabolism” at the DLA stage suggests that cells may maintain or enhance basic energy production to supply raw materials and energy for subsequent high-energy-demanding stress responses, such as the synthesis of large quantities of defensive compounds [39]. The enrichment of “Nitrogen metabolism” at the DLB stage indicates an adjustment in nitrogen assimilation and recycling under severe dehydration to support the synthesis of essential substances.
This study observes significant activation of the flavonoid biosynthesis pathway, evidenced by multi-layered data from transcription (upregulation of key genes like PAL, CHS, FLS) to metabolism (accumulation of flavonols like quercetin). The potent antioxidant activity of flavonoids allows them to effectively quench excess ROS and protect cellular structures [40]. However, when viewed in the context of broader data, flavonoid accumulation should not be seen as an isolated event. Firstly, it is an output branch of the larger “Phenylpropanoid biosynthesis” network [41,42], whose upstream activation provides common precursors for various compounds, including flavonoids and lignin [25,39]. Secondly, flavonoid synthesis is precisely regulated by upstream signals. Its co-enrichment with MAPK and glutathione metabolism at the DLA stage resembles a rapid response under a “stress alarm,” while at the DLB stage, its synthesis is integrated into a more finely tuned network centered on “Plant hormone signal transduction.” In Arabidopsis, hormones like JA and ABA have been shown to specifically regulate different flavonoid branches [43,44,45]. This may explain why the metabolic profile at DLB is more focused compared to DLA—hormonal signals may direct carbon flux toward the most urgently needed specific flavonoid subtypes or other phenylpropanoids at that time [46,47].
Furthermore, metabolomic data hint at other potential protective mechanisms. For instance, the upregulation of genes related to “Cutin, suberin, and wax biosynthesis” may be associated with the reinforcement of the cell wall and outer layers of the plasma membrane to reduce water loss [48]. The consistent enrichment of “ABC transporters” may be involved in expelling harmful substances from cells or locating protective compounds to specific sites, which is crucial for maintaining cytoplasmic homeostasis [49].
In summary, at the DLA stage, cells perceive the initial stress, sound an alarm via pathways like MAPK, rapidly enhance basic metabolism and antioxidant capacity, and initiate the large-scale synthesis of defensive substances, including flavonoids, trading “resource consumption” for immediate protection in preparation for impending severe stress [50]. At the DLB stage, facing intense dehydration, cells activate a precise regulatory network centered on hormonal signaling [51]. The focus of the response shifts toward a more refined “balance-reconstruction,” including the hormone-signal-mediated fine-tuning of specific metabolic pathways, adjustments to primary metabolism like nitrogen metabolism, and enhanced transmembrane transport of substances to maintain homeostasis [52,53].
4. Materials and Methods
4.1. Plant Material
The Populus davidiana × P. tremuloides Z9-1 mountain ash is a clonal variety bred through crossbreeding between a superior tree of mountain ash from Weihe Mountain in Heilongjiang Province and an excellent clonal variety of American mountain ash. It is based on its tissue culture seedlings, which are induced to form sterile seedlings using differentiation medium, then transferred to the rooting medium to induce rooting. The propagation cycle is 4 weeks. The temperature in the cultivation room is 25 ± 2 °C, and the photosynthetic effective radiation is 50 μmol·m^−2^·s^−1^, with 16 h of light per day. The formulation of the culture medium and reagents used in the experiment is shown in Table 3, with a pH of 5.8.
4.2. Identification of Critical Parameters in the Cryopreservation System for Shoot Tips of Populus davidiana × P. tremuloides
Experimental optimization of cryopreservation protocols was performed using a one-factor-at-a-time approach. Each experimental group comprised 10–15 shoot tips, with three biological replicates per treatment.
The optimized vitrification-based cryopreservation protocol for Populus davidiana × P. tremuloides shoot tips was conducted as follows: shoot tips (approx. 2.0 mm in length) were excised and precultured on MS medium (pH 5.8) supplemented with 0.5 mol/L sucrose at 4 °C for 3 days. Subsequently, samples were loaded at room temperature for 20 min, treated with PVS3 solution for 60 min, and immersed in liquid nitrogen for 1 h. For recovery, shoot tips were thawed in a 40 °C water bath for 1 min, rinsed twice with loading solution for 20 min, transferred to regeneration medium, and maintained in darkness for 7 days before being returned to standard light conditions.
For the encapsulation–vitrification method, an additional encapsulation step was introduced following preculture: shoot tips were suspended in sodium alginate solution and cross-linked by dripping with CaCl_2_ solution for 20 min under static conditions to form encapsulation beads. The beads were then subjected to loading at room temperature for 20 min, treated with PVS2 solution on ice for 60 min, and cryopreserved in liquid nitrogen for 1 h. Thawing was performed in a 40 °C water bath for 3 min. After unloading, shoot tips were transferred to recovery medium and maintained in darkness for 7 days before being returned to normal light conditions.
Building upon the two cryopreservation protocols described above, a series of single-factor experiments were conducted to systematically optimize the preservation procedure. The screening methodology was adapted from established protocols by Ma [54]. (1) Cold acclimation duration: Second-generation sterile plantlets were cold-acclimated at 4 °C for 0, 1, 2, 3, and 4 weeks. Non-acclimated seedlings served as the control to assess the effect of acclimation period on survival rate. (2) Sucrose preculture concentration: Shoot tips from 2-week cold-acclimated plantlets were inoculated onto MS medium (pH 5.8) supplemented with 0.1, 0.3, 0.5, 0.7, or 0.9 mol/L sucrose and precultured at 4 °C for 3 days. Survival rates were compared among concentrations. (3) Sucrose preculture duration: Shoot tips from 2-week cold-acclimated plantlets were precultured in MS medium with 0.5 mol/L sucrose at 4 °C for 1, 3, 5, 7, or 9 days to evaluate the influence of preculture time on survival. (4) Vitrification solution exposure time: For the vitrification method, shoot tips from 2-week cold-acclimated plantlets were treated with PVS3 for 0, 30, 60, 90, or 120 min. For the encapsulation–vitrification method, samples were treated with PVS2 on ice for 0, 60, 120, 180, or 240 min. Effects of dehydration duration on survival were compared accordingly.
4.3. Assessment of Post-Cryopreservation Shoot Tip Survival and Regeneration
Determination of Survival Rate in Cryopreserved Shoot Tips via TTC Staining [54]: The treated shoot tips were immersed in a 2,3,5-triphenyltetrazolium chloride (TTC) solution at a concentration of 4 g/L and incubated in a 37 °C water bath in darkness for 30 min. After staining, the samples were examined under a stereomicroscope. Shoot tips exhibiting red coloration in the apical meristem were recorded as viable, while those without coloration were considered non-viable. The survival rate was calculated as the percentage of viable samples relative to the total number of treated samples.
The regeneration ability assessment was carried out by removing the cells, inoculating the stem tips onto the recovery medium, and then culturing in the dark for 7 days. After that, the medium was switched to normal light conditions. The regeneration was considered successful when the stem tips turned green and new leaves emerged. The regeneration rate was calculated as the percentage of regenerated stem tips out of the total number of inoculated stem tips.
4.4. Histological Analysis of Shoot Tip Cell Survival Following Cryopreservation
Histological examination was performed on shoot tips of Populus davidiana × P. tremuloides following cryopreservation. Fresh, untreated shoot tips served as the positive control, whereas those directly plunged into liquid nitrogen without cryoprotection were designated as the negative control. The experimental groups comprised shoot tips that had undergone a 3-day recovery culture after treatment with the two cryopreservation protocols. The samples were immersed in formalin-acetic acid-alcohol fixative (FAA, 70%) for 30 min, followed by stepwise dehydration in a graded ethanol series (50%, 70%, 80%, 85%, and 95% anhydrous ethanol) with each step lasting 1 h at low temperature. After dehydration, the samples were rinsed twice in xylene (1.5 h each). Subsequently, the samples were infiltrated with paraffin and stored at 4 °C overnight; this infiltration process continued for 5 days, with the paraffin solution being replaced twice daily. The samples were then embedded and solidified in embedding cassettes on a cooling stage. The paraffin-embedded blocks were sectioned into slices of 10–14 μm thickness using a rotary microtome (Leica, RM2245), stained with 0.1% toluidine blue, and finally examined and photographed under a microscope for histological observation [55].
4.5. Assessment of Genetic Stability
Genetic Stability Analysis Using ISSR Markers: Tissue-cultured plantlets prior to cryopreservation treatment were used as the control. Genomic DNA was extracted from leaves of the tissue-cultured plantlets using the cetyltrimethylammonium bromide (CTAB) method. Eight primers were selected from an initial set of 100 through preliminary screening for detection. Polymerase chain reaction (PCR) amplification was performed on the samples. The reaction mixture (30 µL total volume) contained 5 µL of 2× Rapid Taq Master Mix, 2 µL of template DNA, 2 µL of primer, and 21 µL of ultrapure water. The PCR protocol was set as follows: initial denaturation at 95 °C for 3 min, 35 cycles of denaturation at 95 °C for 15 s, primer annealing for 15 s, and extension at 72 °C for 1 min, followed by a final extension at 72 °C for 5 min. The amplification products were separated by agarose gel electrophoresis and visualized using a gel imaging system, and clearly distinguishable bands were counted for analysis.
4.6. Analysis of Physiological and Biochemical Parameters in Populus davidiana × P. tremuloides Shoot Tips
Shoot tips from Populus davidiana × P. tremuloides were subjected to various cryo-preservation procedures via the vitrification method and used as experimental material. According to the method of Zhu [56], the changes in soluble sugar and soluble protein contents in Populus davidiana × P. tremuloides were determined. Reagents from Suzhou Greis Biotechnology Co., Ltd. were used to separately measure the contents of malondialdehyde (MDA, G0109W), hydrogen peroxide (H_2_O_2_, G0112W), catalase (CAT, G0105W), peroxidase (POD, G0107W), superoxide dismutase (SOD, G0101W), ascorbic acid (ASA, G0201W), reduced glutathione (GSH, G0206W), the mass fraction of free proline, and the scavenging ability of superoxide anion and hydroxyl radical.
4.7. Detection of Relative Cell Survival Rate During Cryopreservation Process
The relative survival rate of cells was detected by following Wang’s [57] method: Materials were immersed in TTC staining solution and incubated overnight at 28 °C. The following day, samples were rinsed three times with distilled water and transferred into 95% anhydrous ethanol. The ethanol was boiled to extract the TTC-stained products until the shoot tips turned white. Subsequently, the volume was adjusted to 5 mL with 95% anhydrous ethanol. Absorbance was measured at 485 nm using a spectrophotometer. Untreated shoot tips served as the control. The ratio of absorbance of treated shoot tips to that of control shoot tips was taken as the proportion of viable cells.
4.8. Integrated Transcriptome and Metabolome Analysis of Populus davidiana × P. tremuloides Shoot Tips During Cryopreservation
4.8.1. Transcriptome Measurement
Following three different types of treatments (DLA, DLB and DLC), RNA was extracted from the shoot tips of poplars (Populus davidiana × P. tremuloides). Each sample had three biological replicates, and then nine cDNA libraries were constructed [58]. Total RNA was extracted from the shoot tip samples using the conventional CTAB method (Cyltrimethylammonium Bromide), as previously described [59]. The purity and concentration of RNA samples were determined using the Qubit Fluorometer and the Agilent 5400 Bioanalyzer to ensure their suitability for cDNA library construction. The qualified RNA was then used to construct cDNA libraries following the manufacturer’s instructions for the Illumina RNA-Seq Library Preparation Kit (Illumina, CA, USA), and the resulting libraries were subsequently sequenced. Qualified libraries were normalized based on effective concentration and combined proportionally to meet the desired sequencing depth. The final pool was subjected to PE150 (paired-end 150 bp) sequencing on an Illumina No-vaSeq system.
RNA-Seq data processing and quantification were performed using the RSEM (RNA-Seq by Expectation-Maximization) pipeline, which applies an expectation-maximization algorithm to accurately estimate transcript abundance. The reference genome sequence of Populus alba was retrieved from the NCBI database (https://www.ncbi.nlm.nih.gov, accessed on 14 February 2025), a comprehensive platform for plant genomic resources. Following quality control with fastp [60] to remove low-quality reads, the clean reads from each sample were subsequently aligned to the Populus alba reference genome. For comparative analysis, sequencing reads were aligned to the reference genome using STAR [61], followed by quantification of gene expression levels with RSEM. Differential expression analysis was performed on the different poplar samples using edge R 3.36.0 [62]. The significant p-values were adjusted for multiple testing using the BH (Benjamini–Hochberg) method. Genes with a false discovery rate (FDR) < 0.05 and an absolute log_2_(fold change) ≥ 1 were considered differentially expressed.
To perform functional enrichment analysis on the identified differentially expressed genes (DEGs), we used the ClusterProfiler software (Version 4.5) for both Gene Ontology (GO) and KEGG pathway analyses. Significant terms were defined as those with a BH adjusted p-value < 0.05. Additionally, enrichment analysis was corroborated using the DEGSeq R package (version 1.12.0) and Goseq (v1.46.0) packages under the same significance threshold.
4.8.2. Metabolome Measurement
The stem tips treated with liquid nitrogen were used as the control group (DLC), while the stem tips treated with preculture/loading (DLA) and osmotic dehydration (DLB) were used as the experimental groups. Each group had 3 replicates. Metabolite Extraction (the reagents and instruments used are listed in Table 4 and Table 5): (1) The samples were freeze-dried and ground into powder (60 Hz, 30 s). (2) Approximately 25 ± 1 mg of the lyophilized powder was accurately weighed into a centrifuge tube at low temperature, followed by addition of homogenization beads and 1000 μL of extraction solvent (methanol: acetonitrile: water = 2:2:1, v/v/v) containing isotope-labeled internal standards. (3) The mixture was vortexed for 30 s. (4) Homogenization was performed using a homogenizer (35 Hz, 4 min), followed by ultrasound treatment in an ice-water bath for 5 min. This cycle was repeated three times. (5) The samples were kept at −40 °C for 1 h. (6) Centrifugation was carried out at 4 °C and 12,000 rpm (centrifugal force 13,800× g, radius 8.6 cm) for 15 min. (7) The supernatant was filtered through a 0.22 μm microporous membrane, and the filtrate was collected for subsequent instrumental analysis.
Instrumental Analysis: Chromatographic separation was performed on a Vanquish ultra-high-performance liquid chromatography system (Thermo Fisher Scientific, 1290 Infinity II, USA) using a Phenomenex Kinetex C18 column (2.1 mm × 50 mm, 2.6 μm). The mobile phase consisted of (A) water containing 0.01% acetic acid and (B) isopropanol: acetonitrile (1:1, v/v). The sample tray temperature was maintained at 4 °C, and the injection volume was 2 μL. Mass spectrometry analysis was carried out on an Orbitrap Exploris 120 mass spectrometer operated under Xcalibur software (version 4.4, Thermo Fisher Scientific) for both full-scan and MS/MS data acquisition. The key parameters were set as follows: sheath gas flow rate, 50 Arb; auxiliary gas flow rate, 15 Arb; capillary temperature, 320 °C; full MS resolution, 60,000; MS/MS resolution, 15,000; collision energy, stepped NCE 20/30/40; spray voltage, 3.8 kV (positive ion mode) or −3.4 kV (negative ion mode).
The raw data were converted into mzXML format using ProteoWizard software (Version 3). Metabolite identification was subsequently performed with R packages against the BiotreeDB (V3.0, reference standard database) and BT-Plant (V1.1, plant-specific database) libraries [63], followed by visualization analysis.
4.9. Data Processing and Analysis
All data were recorded and organized using Excel 2024. One-way ANOVA tests and Pearson correlation analyses were conducted using SPSS 27.0. Graphs were created using Origin 2024Pro software.
5. Conclusions
In conclusion, this study, from the perspective of systems biology, demonstrates that the stem tips of Populus davidiana × P. tremuloides undergo a multi-level and finely regulated adaptive mechanism during the vitrification preservation process, especially when exposed to osmotic dehydration stress. The core of this mechanism involves rapidly transmitting stress information through hormone signaling, activating the antioxidant system to maintain oxidative balance, and massively reconstructing the secondary metabolic network (especially the flavonoid synthesis pathway) to provide efficient chemical protection. These findings not only deepen our understanding of the stress-resistant physiology of woody plant stem tips but also provide theoretical basis and practical directions for optimizing the cryopreservation preservation technology of forest tree germplasm. For example, adding an appropriate amount of flavonoid synthesis precursors in the pretreatment solution may enable “pre-adaptation” of the stem tips, enhancing their stress resistance, and thereby increasing the survival rate and regeneration rate after vitrification. Future research will focus on using gene editing or overexpression techniques to verify the specific functions of key genes (such as PAL, CHS) in improving the cryopreservation tolerance of poplar and other forest tree germplasm.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Huang M.B. Han S. Li M.M. Jiao Z.Y. Li Z. Wang Y.J. Liu C. Wang H.L. Yin W. Xia X. The Cysteine-Rich Peptide Pe GASA 15 from Populus Euphratica Enhances Drought and Salinity Tolerance in Poplar Ind. Crops Prod.202523712215010.1016/j.indcrop.2025.122150 · doi ↗
- 2Larsen S.U. Hestbjerg H. Jørgensen U. Kongsted A.G. Ensiling of Willow and Poplar Biomass Is Improved by Ensiling Additives Agriculture 202414185310.3390/agriculture 14101853 · doi ↗
- 3Ma X.Y. Blystad D.-R. Wang Q.C. Hamborg Z. Cryopreservation of Rubus Viruses in Raspberry Shoot Tips via Droplet-Vitrification: Assessment of Viral Preservation, Localization, and Post-Thaw Transmission Capacity Plant Methods 20252113710.1186/s 13007-025-01454-w 41139794 PMC 12557910 · doi ↗ · pubmed ↗
- 4Martín C. Nagel M. Ibáñez M.A. Senula A. Pirredda M. González-Benito M.E. Genetic and Epigenetic Fidelity of Garlic Cryopreserved Plant Material Compared to Field Collections Sci. Hortic.202534511413210.1016/j.scienta.2025.114132 · doi ↗
- 5Kulus D. Shoot Tip Cryopreservation of Lamprocapnos spectabilis (L.) Fukuhara Using Vitrification, Droplet-Vitrification and Encapsulation-Vitrification Techniques Cryobiology 20209728410.1016/j.cryobiol.2020.10.136 · doi ↗
- 6Sakai A. Engelmann F. Vitrification, Encapsulation-Vitrification and Droplet-Vitrification: A Review Cryo Letters 20072815117217898904 · pubmed ↗
- 7Langis R. Steponkus P.L. Cryopreservation of Rye Protoplasts by Vitrification Plant Physiol.19909266667110.1104/pp.92.3.66616667332 PMC 1062351 · doi ↗ · pubmed ↗
- 8Chang Y. Reed B.M. Preculture Conditions Influence Cold Hardiness and Regrowth of Pyrus Cordata Shoot Tips after Cryopreservation Hort Science 2001361329133310.21273/HORTSCI.36.7.1329 · doi ↗