Anti-Obesity and Diuretic Effects of Immature Watermelon Rind Extract in HFD-Induced Obese Mice
Yun-seong Lee, Ji yong Kim, Sunju So, Bo-Young Lee

TL;DR
This study shows that immature watermelon rind extract can reduce obesity and improve metabolism in mice fed a high-fat diet.
Contribution
The novel finding is that immature watermelon rind extract significantly reduces obesity and improves metabolic markers in HFD-induced obese mice.
Findings
WM and WMS reduced body weight, liver weight, and epididymal fat mass in obese mice.
Treatments lowered serum cholesterol and triglycerides and increased serum sodium levels.
Histological analysis showed reduced NASH scores and smaller adipocytes in treated mice.
Abstract
Background/Objectives: Immature watermelon (WM) rind contains higher levels of citrulline and potassium than mature fruit and may exert diuretic and metabolic benefits. This study aimed to evaluate the anti-obesity and diuretic effects of WM and salt-treated watermelon rind extract (WMS) in high-fat diet (HFD)-induced obese mice, focusing on changes in lipid metabolism, sodium handling, and tissue-level alterations. Methods: Citrulline concentrations in WM and WMS were quantified using high-performance liquid chromatography with ultraviolet detection (HPLC-UV). Four-week-old male C57BL/6 mice were fed an HFD for 6 weeks and subsequently administered WM (380 mg/kg) or WMS (380 mg/kg) orally for an additional 6 weeks. Body weight, food intake, organ and fat-pad weights, serum biochemical markers, and sodium (Na+) levels were measured. Histopathological analyses of liver and epididymal…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1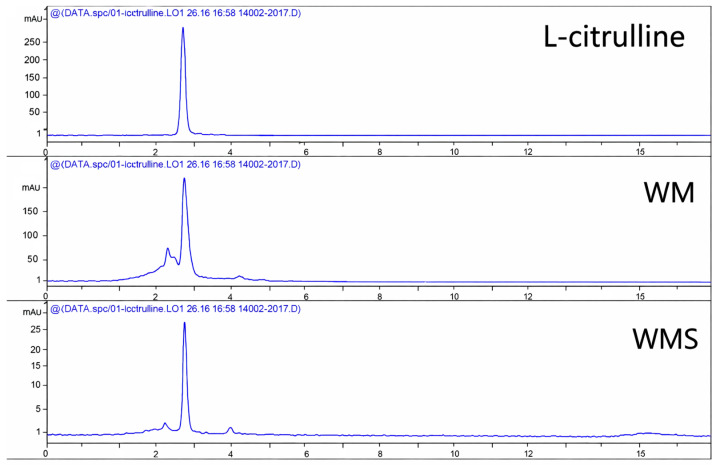 Figure 2
Figure 2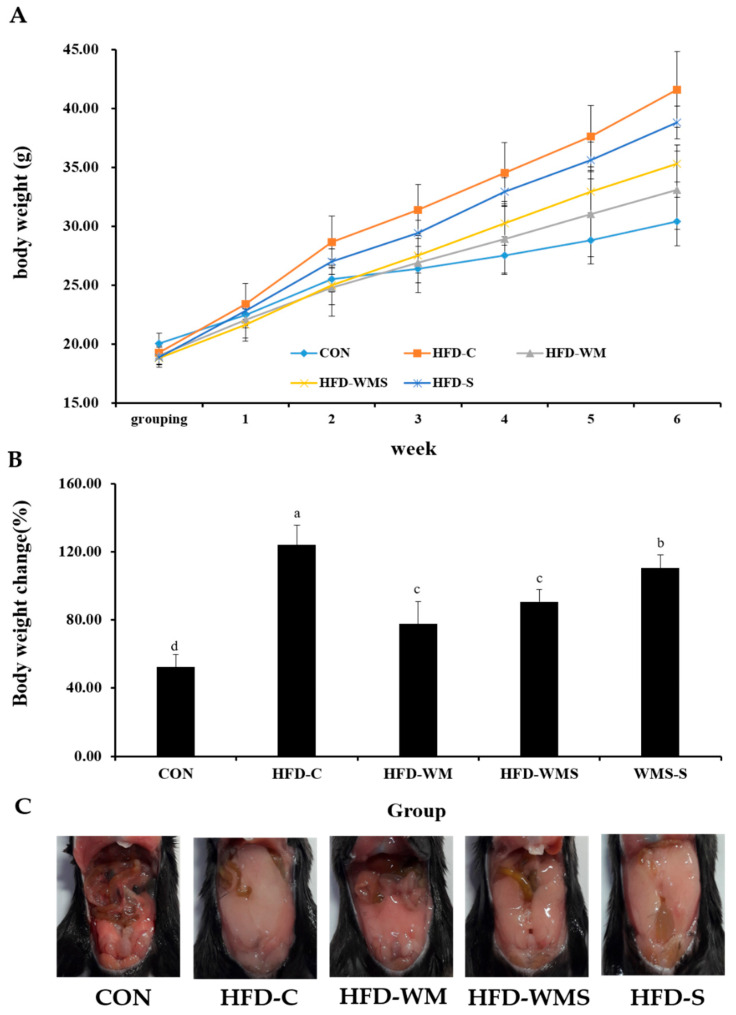 Figure 3
Figure 3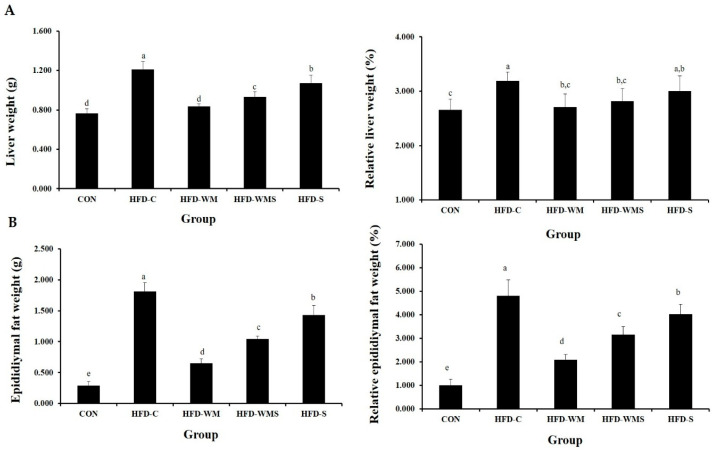 Figure 4
Figure 4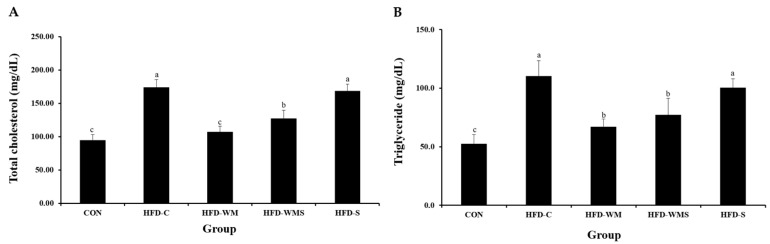 Figure 5
Figure 5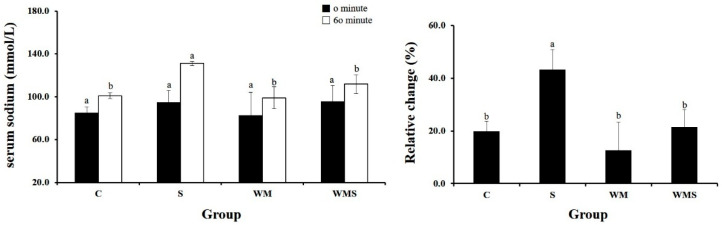 Figure 6
Figure 6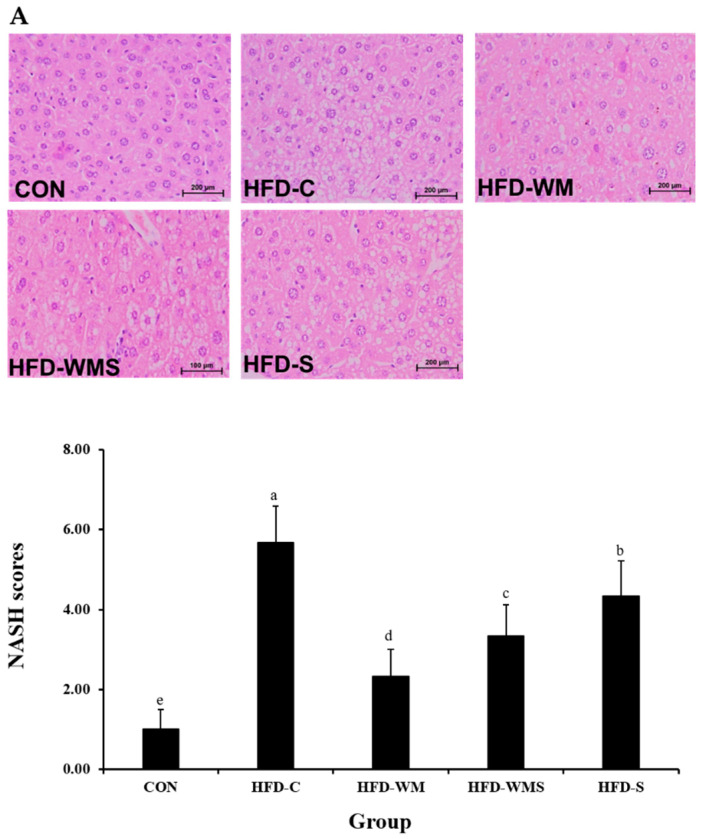 Figure 7
Figure 7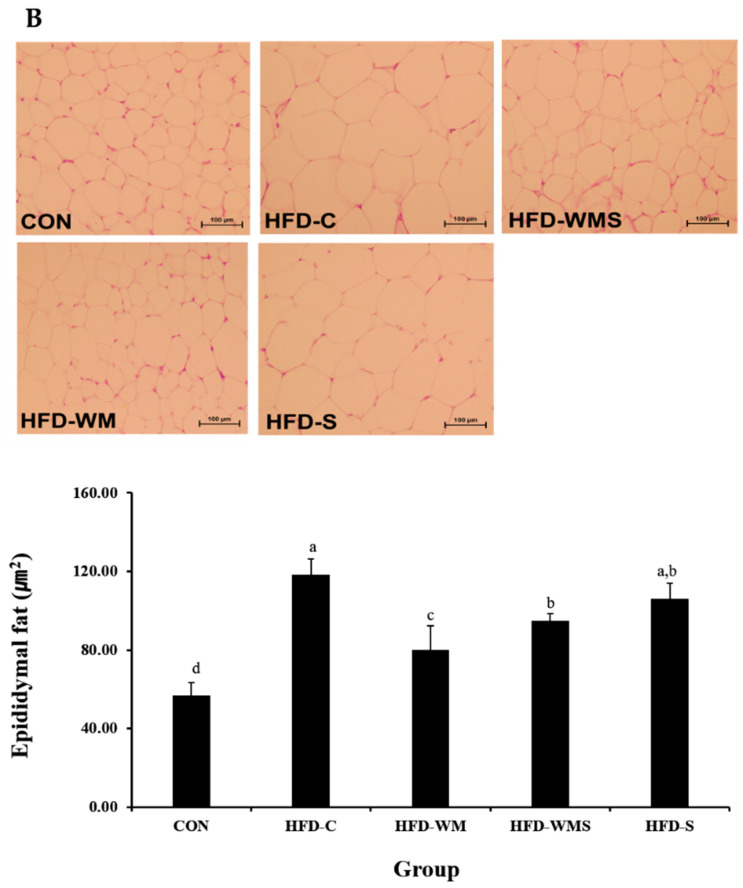 Figure 8
Figure 8- —Wonkwang University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAdvances in Cucurbitaceae Research · Pharmacology and Obesity Treatment · Phytochemicals and Antioxidant Activities
1. Introduction
Obesity is classified as a serious medical condition, as it induces various complications such as diabetes, hypertension, cardiovascular diseases, cerebrovascular diseases, arthritis, breast cancer, and colorectal cancer [1]. According to the World Health Organization (WHO), approximately 4 million deaths annually are attributed to complications caused by overweight and obesity. The prevalence of overweight and obesity quadrupled between 1975 and 2016, and it is projected that by 2030, 42% of U.S. adults will be obese. Annual healthcare expenditure for obese individuals has also been steadily increasing [2,3]. Obesity results from an imbalance between energy intake and expenditure, with excess energy stored as fat in adipocytes [4]. Visceral fat accumulation is considered a major contributor to inflammatory diseases and is closely associated with increased blood LDL cholesterol, triglycerides, and reduced HDL cholesterol, thereby heightening insulin resistance and the risk of cardiovascular diseases [5]. Abdominal obesity, particularly subcutaneous fat accumulation, is reported to lead to metabolic disorders, such as non-insulin-dependent diabetes mellitus, hypertension, and dyslipidemia, increasing mortality rates [6]. Obesity can also result from genetic predisposition, endocrine disorders, medication use, lack of exercise, and eating disorders triggered by psychological stress, all of which pose significant social issues [7]. Excessive sodium intake is strongly linked to hypertension, stroke, cardiovascular diseases, and kidney disorders, as supported by numerous studies [8]. Increasing evidence demonstrates that prolonged high sodium intake is associated with adipocyte hypertrophy and increased white adipose tissue mass, even independent of greater caloric intake [9,10]. The 2010 Korea National Health and Nutrition Examination Survey reported that high sodium intake is a characteristic dietary issue among diabetes patients. The average daily sodium intake for Korean adults was 4878 mg, while that for diabetic individuals was 5939 mg [11,12]. Koreans consume excessive sodium compared to other countries, mainly due to dietary habits involving pickled foods like kimchi, soups, stews, and processed foods such as instant noodles [13]. While exercise and dietary modification are fundamental to preventing and treating obesity, lifestyle changes are challenging. Although pharmacological treatments have been proposed as alternatives, they often yield limited efficacy or cause adverse effects [14,15]. Consequently, research on bioactive compounds derived from natural materials with minimal side effects is gaining traction. Watermelon (Citrullus lanatus), a member of the Cucurbitaceae family, is a popular summer fruit with high water content. Beyond its consumption as a fresh fruit, watermelon has also been utilized in medicinal applications [16]. It is rich in minerals such as potassium, phosphorus, magnesium, calcium, sodium, and iron, as well as free sugars (fructose, glucose, sucrose) and organic acids (citric, malic, succinic, and fumaric acids) [17]. Watermelon contains citrulline, an amino acid that aids urea synthesis, promoting diuresis and exhibiting potential effects against edema, nephritis, cystitis, urethritis, hypertension, inflammation, and fever [17]. Previous domestic research on Citrullus lanatus (watermelon) has investigated a diverse range of applications and bioactivities. Studies have characterized its volatile aromatic compounds [18], developed functional beverages utilizing natural pigment extracts derived from watermelon flesh [19], and explored its potential as a raw material in the production of fermented alcoholic beverages [20]. Additional investigations have examined juice concentration techniques employing reverse osmosis [21] and analyzed the spatial distribution of sugar content across different anatomical sections of watermelon and melon fruits [22]. The edible flesh of watermelon has been reported to exhibit antibacterial, depigmenting (whitening), and anti-inflammatory activities [23], whereas the rind demonstrates notable antioxidant and depigmenting properties. Moreover, watermelon seeds have been shown to possess antibacterial, depigmenting, and anti-inflammatory effects [24,25]. Recent murine and human studies have advanced the understanding of watermelon species’ anti-obesity potential. For instance, Kang et al. (2024) demonstrated that Citrullus mucosospermus extract attenuated weight gain in high-fat diet (HFD)-induced mice [26]. Furthermore, Daughtry et al. (2023) reported that the consumption of blended watermelon led to reductions in BMI and body fat among children with overweight/obesity [27]. Meanwhile, watermelon rind has been shown to contain high levels of L-citrulline and potassium, which may enhance sodium excretion and modulate lipid metabolism [28,29]. Recent studies have highlighted the growing importance of valorizing agricultural by-products by extracting functional compounds from plant materials traditionally discarded as waste [30,31]. Immature watermelons removed during pre-harvest pruning represent a largely unexplored biomass resource, consisting of a pale, undifferentiated mesocarp without red edible flesh.
In this context, the present study investigates the biological activity of extracts derived from whole immature watermelons to enhance resource utilization and reduce agricultural waste. Specifically, we examined immature watermelons culled during production and analyzed the sodium content of their extracts. The purpose of this study was to evaluate whether a mildly salt-treated immature watermelon rind preparation naturally rich in potassium and citrulline could serve as a potassium- and citrulline-enhanced salty food model designed as a partial alternative to conventional salt. Potassium promotes natriuresis by stimulating Na^+^/K^+^-ATPase-dependent renal sodium transport [32], whereas citrulline improves renal microcirculation and enhances nitric oxide-mediated natriuretic responses [33]. Accordingly, sodium ingested within this matrix is expected to be more readily excreted rather than retained, thereby reducing the metabolic burden typically associated with salt intake [34]. In addition, citrulline has been reported to improve mitochondrial function, promote fatty-acid oxidation, and support thermogenic activity [35,36], suggesting potential anti-obesity effects beyond sodium handling. Ultimately, immature watermelon rind extract and its salt-treated may alleviate obesity and metabolic dysfunction induced by a high-fat diet. This research therefore aims to establish immature watermelon rind as a value-added functional material.
2. Materials and Methods
2.1. Experimental Materials
Immature watermelons used in this study were purchased from a local farm in Jeollabuk-do, Korea. The watermelons were sourced from the ‘Urikkul’ cultivar grown in Jeongeup, Iksan, Gimje, and Gochang within Jeonbuk State, South Korea. To ensure consistency in sugar content and moisture levels during processing, we standardized the immature watermelons to a horizontal diameter of 10 ± 1 cm and a vertical diameter of 11 ± 1 cm. Immature watermelons were used at a developmental stage prior to red flesh formation (Figure 1A). At this stage, the mesocarp and epicarp are not yet differentiated; therefore, the whole fruit was sliced and dried as a single integrated material. A total of 30 kg of immature watermelon was washed, thinly sliced, and dried to obtain 2.4 kg of dried material. The dried slices were placed in an extraction vessel, and distilled water five times the weight of the material was added. Hot water extraction was conducted at 100 °C for 3 h and repeated twice. The combined extract was filtered and adjusted to a final concentration of 2–5 Brix. For the preparation of the immature watermelon extract salt (WMS), 1 kg of Korean solar salt (produced in Shinan, Jeollanam-do, Republic of Korea) was gradually added to the immature watermelon extract (WM) while boiling at 100 °C, stirring continuously until fully dissolved. The resulting brine was filtered through a fine mesh (2 mm × 2 mm) to remove impurities and then heated at 200 °C until the salt crystallized. The salt crystals were naturally dried in the shade for over three days and then dried at 60 °C for 4 h using a dryer. After grinding the dried crystals, they were filtered through a fine mesh (2 mm × 2 mm) and further dried at 80 °C for approximately 4 h. The final product, immature watermelon extract salt (WMS), was obtained by re-filtering through the same mesh (Figure 1B).
2.2. Citrulline Analysis
Comparing the citrulline content of WM and WMS, component analysis was conducted using high-performance liquid chromatography (HPLC). A standard citrulline solution was prepared at 1000 ppm and serially diluted to concentrations of 0, 20, 40, 60, and 80 µg/mL. Each concentration (10 µL) was injected into the HPLC, and a calibration curve was generated at 207 nm. The chromatographic separation was performed using a YMC-Triart C18 column (250 × 4.6 mm I.D., S-5 μm, 12 nm). The mobile phase consisted of 3 mM phosphoric acid and acetonitrile in a 70:30 ratio, with a flow rate of 0.7 mL/min at room temperature. The calibration curve is represented by the equation
2.3. Experimental Animals
A total of 25 male C57BL/6 mice (4 weeks old) were purchased from Samtako (Osan, Republic of Korea) and used in this study. The animals were randomly allocated into five experimental groups (n = 5 per group). The average body weight of the mice was 19.29 ± 0.84 g. Obesity was induced by feeding the mice a high-fat diet (HFD, 60% kcal fat; Samtako, Osan, Republic of Korea), and each extract was suspended in distilled water and administered once daily by oral gavage according to individual body weight to ensure complete dose delivery. The high-fat diet feeding period lasted six weeks to induce obesity, after which all treatments (WM, WMS, refined salt, or water) were administered daily for another six weeks concurrently with HFD feeding. Each group consisted of five mice, which represents a minimal sample size commonly used in rodent obesity studies under the 3Rs principle. The animals were housed under controlled conditions (12 h light/dark cycle, 23 ± 2 °C, 50 ± 10% humidity). The high-fat diet was purchased from Samtako (Osan, Republic of Korea) and its composition is shown in Table 1. All animal procedures were conducted in accordance with the Guide for Animal Experimentation of Wonkwang University and approved by the Institutional Animal Care and Use Committee (IACUC) of Wonkwang University (Approval No. WKU24-44). No predefined inclusion or exclusion criteria were applied during the experiment. No animals or data points were excluded from the analysis, and no attrition occurred during the study. The investigators were not blinded to group allocation during animal allocation, conduct of the experiment, outcome assessment, or data analysis.
2.4. Experimental Groups
The mice were randomly divided into five groups as follows: normal control group (CON, fed standard diet and distilled water; DW), high-fat diet control group (HFD-C, fed 60% HFD and DW), HFD with immature watermelon extract 380 mg/kg group (HFD-WM), HFD with salt-treated immature watermelon extract 380 mg/kg group (HFD-WMS), and HFD with refined salt 380 mg/kg group (HFD-S). Each experimental group consisted of five mice, which represents the minimal sample size required to achieve statistical reliability while adhering to the 3Rs principle, as supported by previous rodent studies (Table 2). The oral dose of 380 mg/kg sodium was selected based on human high-salt intake after allometric scaling. Although recommended sodium intake is <2 g/day, actual consumption often reaches 3.0–4.3 g/day (≈50–70 mg/kg in a 60-kg adult) [37]. Using the standard body-surface-area Km conversion factor for rats (≈6.2), this corresponds to ~310–360 mg/kg in rats [38]. Accordingly, 380 mg/kg was chosen as a physiologically relevant high-sodium load that challenges Na^+^/K^+^ homeostasis while remaining within the safe and commonly used dose range for nutritional and toxicological studies. WM and WMS extracts were prepared as freeze-dried powders and reconstituted in distilled water according to body weight extrapolation. The preparations were administered once daily by oral gavage at a fixed volume of 300 µL per mouse (approximately 10 mL/kg), corresponding to a final dose of 380 mg/kg/day, which is consistent with established dosing ranges for phytochemical preparations. All treatments were delivered as aqueous suspensions to ensure accurate and consistent dosing, independent of drinking-water variability. Water intake was recorded separately and expressed as mL/mouse/day.
2.5. Measurement of Body Weight and Food Intake
Body weight was measured weekly at a fixed time (10:00 AM) using a digital balance (IB-3100, Innotem, Yangju, Republic of Korea). Food and water intake was recorded twice weekly at consistent times. The food efficiency ratio (FER) was calculated as the weight gain divided by total food intake during the experimental period.
2.6. Organ Weight Measurement
Following blood collection, the liver, epididymal fat, and retroperitoneal fat were excised and weighed. The absolute and relative organ weights were calculated. Relative organ weight (%) was calculated using the formula (organ weight/body weight) × 100.
2.7. Serum Lipid Profile and Sodium Ion Concentration
After 6 weeks, blood was collected from the retro-orbital vein under isoflurane anesthesia following a 12 h fast and centrifuged at 3000 rpm for 20 min. Serum levels of total cholesterol (TC), triglycerides (TG), high-density lipoprotein cholesterol (HDL-C), and low-density lipoprotein cholesterol (LDL-C) were measured using an enzymatic colorimetric assay (Get Kit, Roche, Mannheim, Germany) on a Modular Analytics device (PE, Roche, Germany). Additional parameters were analyzed using an ion-sensitive electrode analyzer.
2.8. Histopathological Analysis
At the end of the experiment, the animals were euthanized using an approved method in accordance with institutional guidelines to ensure death and minimize pain and distress. The liver and adipose tissues were excised, rinsed with saline, and blotted dry. Liver tissues were fixed in 10% neutral buffered formalin, embedded in paraffin using standard procedures, sectioned at 4 μm thickness, and stained with hematoxylin and eosin (H&E). Histopathological observations were conducted at 200× magnification using an Olympus BX50 optical microscope (Olympus, Tokyo, Japan). Nonalcoholic steatohepatitis (NASH) scoring was based on the presence of macrovesicular steatosis, hepatocellular ballooning, and lobular inflammation, mainly observed in zone 3 of the liver. A NASH score ≥ 5 was considered diagnostic of NASH, whereas a score ≤ 2 was classified as non-NASH [39].
2.9. Statistical Analysis
All experimental data are presented as mean ± standard deviation (SD). Statistical significance between the groups was assessed using one-way ANOVA, followed by Duncan’s post hoc test using SPSS v.12. A p-value < 0.05 was considered statistically significant. Prior to statistical analysis, the data were assessed for normality and homogeneity of variance. All data met the assumptions required for parametric statistical analysis.
3. Results
3.1. Citrulline Analysis in WM and WMS
Citrulline content in WM and WMS samples was analyzed, with detection occurring at a retention time of 3.029 min. The citrulline concentrations were found to be 7.7 mg/g in WM and 3.1 mg/g in WMS. The results of the analysis of WM and WMS, with a visual representation, are provided in Figure 2.
3.2. Change in Body Weight
In a 6-week high-fat diet (HFD) to induce obesity, the administration of the respective test substances led to notable changes in body weight among the experimental groups. No expected or unexpected adverse events were observed during the experimental period. As shown in Figure 3A, both the HFD-WM and HFD-WMS groups exhibited a significant reduction in body weight compared to the HFD control group (HFD-C). By the end of the experiment, the weight gain percentage was 52.4% in the CON group and 124.2% in the HFD-C group, whereas the HFD-WM and HFD-WMS groups showed significantly attenuated weight gains of 79.5% and 88.8%, respectively (Figure 3B). Post-mortem gross morphological observations are shown in Figure 3C. Notable abdominal fat accumulation was observed in all groups except the CON group. However, the extent of fat deposition was visibly reduced in both the HFD-WM and HFD-WMS groups relative to the HFD-C group.
3.3. Body Weight Gain, Food Intake, and Food Efficiency Ratio
During the 6-week dietary intervention, data on body weight progression, weight gain, food consumption, and food efficiency ratio were collected (Table 3). Obesity was operationally defined as an increase of ≥20% body weight relative to the standard diet control and a significant increase in epididymal white adipose tissue weight at 6 weeks. In our study, body weight gain was markedly increased in the HFD-C group, with a total 6-week weight gain of 22.35 g, corresponding to a 115.7% increase compared with CON (10.36 g). WM supplementation reduced weight gain to 14.09 g, representing a 37.0% attenuation relative to HFD-C, while WMS reduced gain to 16.51 g (−26.1%). The positive-control group (HFD-S) gained 19.94 g, showing a 10.8% reduction compared with HFD-C. Despite these findings, we recognize that 6 weeks of HFD may produce an early obesogenic phenotype rather than full established obesity. The food efficiency ratio remained consistent across all groups except for the normal control group, with a value of 0.02. The slight, non-significant daily reduction in feed intake observed in the WM and WMS groups may accumulate over time, leading to mild calorie restriction or altered palatability, which could partially explain the observed anti-obesity effect.
3.4. Liver Weight and Adipose Tissue Weight
Liver weights, both absolute and relative (per 100 g body weight), are presented in Figure 4A. HFD feeding markedly increased liver and visceral fat weights compared with the CON group. WM supplementation significantly attenuated these effects. Liver weight was reduced by 31.1% in the HFD-WM group and 23.0% in the HFD-WMS group relative to HFD-C. Epididymal fat mass, which increased more than six-fold in HFD-C compared with CON, was reduced by 64.2% and 42.5% in the HFD-WM and HFD-WMS groups, respectively (Figure 4B). Relative fat mass normalized to body weight also showed consistent reductions, indicating that WM effectively suppresses high-fat diet-induced visceral adiposity.
3.5. Triglycerides and Total Cholesterol
Serum levels of triglycerides and total cholesterol, the key biomarkers of dyslipidemia and cardiovascular risk, were evaluated (Figure 5A,B). Both parameters were significantly decreased in the HFD-WM and HFD-WMS groups compared to the HFD-C group, implying improved lipid metabolism. In addition to total cholesterol (TC) and triglycerides (TG), detailed values of HDL-C and LDL-C were analyzed. HDL-C levels were significantly elevated in the HFD-C group (128.72 mg/dL) compared with the CON group (75.88 mg/dL, +69.6%). Treatment with WM and WMS markedly reduced HDL-C to 79.76 mg/dL (−38.1%) and 92.76 mg/dL (−27.9%), respectively. LDL-C was calculated using the Friedewald formula. The HFD-C group exhibited a sharp increase (23.20 mg/dL) compared with the CON group (8.40 mg/dL, +176%). WM supplementation lowered LDL-C to 14.00 mg/dL (−39.7%), while WMS reduced it to 19.20 mg/dL (−17.2%) relative to HFD-C. These results confirm that WM supplementation effectively improves HFD-induced dyslipidemia.
3.6. Serum Sodium Ion Concentration
Serum electrolyte concentrations were measured before and 60 min after the administration of the respective samples. As shown in Table 4, the WM group exhibited a 43.2 ± 7.6% increase in serum sodium (Na^+^) levels compared to the S group. In contrast, the WMS group showed a significantly lower increase of 21.5 ± 6.6% (Figure 6). The normal-diet control (CON) group was not included in this subsection because the purpose of Section 3.6 was to evaluate treatment-dependent differences within the HFD-derived experimental groups (HFD-C, HFD-WM, HFD-WMS, and HFD-S), rather than to compare these values with baseline physiological conditions. Baseline comparisons between CON and HFD animals—including body weight, hepatic morphology, adipose histology, and lipid profiles—are already presented in Section 3.1, Section 3.2, Section 3.3, Section 3.4 and Section 3.5. Therefore, the analytical framework in Section 3.6 intentionally used the HFD-control (HFD-C) group as the reference to determine the extent to which each intervention modulated HFD-induced metabolic alteration.
3.7. Histological Effects of WM on Liver and Epididymal Adipose Tissue in High-Fat Diet-Induced Obese Mice
Liver tissue evaluation through H&E staining and optical microscopy examination is presented in Figure 7A. In the CON group, assessment of steatosis grade, lobular inflammation, and hepatocellular ballooning revealed minimal fat accumulation, with no observable inflammation or ballooning. Hepatocytes maintained distinct round nuclei and relatively uniform spacing. HFD-C mice exhibited severe hepatic steatosis, inflammation, and ballooning, resulting in a 5.7-fold increase in NASH compared with the CON group. WM markedly improved these pathological features. The HFD-WM group showed 55.6% and 40.0% reductions in steatosis and lobular inflammation scores, respectively, with complete elimination of ballooning (100% reduction). NASH was reduced by 58.8% compared with HFD-C. The HFD-WMS group also demonstrated substantial improvements, with a 41.2% decrease in NASH. Epididymal adipocyte hypertrophy, which more than doubled in HFD-C mice, was reduced by WM 39.6% and WMS 22.8% (Figure 7B). These results indicate that WM effectively alleviates both hepatic injury and adipocyte enlargement induced by high-fat diet feeding.
4. Discussion
In this study, we investigated the anti-obesity effects and sodium-excreting action of immature watermelon salt, prepared by adding salt to an extract of immature watermelon, when orally administered to high-fat diet (HFD)-induced obese animal models. Obesity is a rapidly increasing chronic disease worldwide and is a major risk factor for cardiovascular disease, type 2 diabetes, and metabolic syndrome [40]. As such, there is growing interest in the development of safe and effective strategies for weight management [41]. Watermelon contains citrulline, which is known to have diuretic effects, and is rich in potassium, which facilitates sodium excretion [22]. Recent studies have focused on strategies to suppress obesity by preventing intracellular fat accumulation or stimulating the breakdown of stored fat [42]. L-citrulline, a non-essential amino acid, has drawn attention for its potential to enhance exercise performance and improve metabolism, thereby exerting anti-obesity effects [43]. The citrulline content of the immature watermelon extract (WM) and immature watermelon salt (WMS) used in this study was analyzed using HPLC, and the levels were found to be 7.7 mg/g and 3.1 mg/g, respectively. The HPLC–UV method used for citrulline quantification follows previously validated protocols demonstrating high linearity, repeatability, and recovery (RSD < 2%, recovery 95–103%) under identical chromatographic conditions [44,45]. Although full validation was not performed in our laboratory, our analytical parameters were consistent with these validated methods, supporting the accuracy of citrulline determination in WM and WMS. According to Rimando et al. [46], the citrulline content in watermelon rind with different flesh colors ranged from 0.8 to 1.50 mg/g, indicating that the immature watermelon used in this experiment contained a higher amount of citrulline. L-citrulline, abundantly present in watermelon rind, has been shown to increase mitochondrial fatty acid oxidation and up-regulate UCP1 expression in adipose tissue, thereby elevating energy expenditure [47]. Furthermore, watermelon rind extract may exert anti-inflammatory effects, improving adipose tissue homeostasis via increased adiponectin and reduced pro-inflammatory cytokines [48]. Indeed, recent studies in obese models have demonstrated that watermelon extract supplementation reduces inflammatory markers and improves lipid metabolism [49]. Although this study did not assess these mechanistic pathways directly, these observations provide a biological rationale for the observed reductions in fat mass and histological improvements in the liver. Future work should include gene- and protein-level assays to elucidate these mechanisms. Furthermore, Takeda et al. (2016) [50] reported in an animal study that citrulline supplementation improved mitochondrial function and promoted fatty acid oxidation. This suggests that citrulline may optimize cellular energy production processes, enhancing the use of fat as a primary energy source. In our experiment, the measurement of food intake in HFD-induced obese animals over a 6-week period showed that although the HFD-C and HFD-S groups did not exhibit significant weight reduction, the HFD-WM and HFD-WMS groups demonstrated a significant decrease in body weight. The slight reduction in feed intake observed in the WM and WMS groups, although not statistically significant per day, may cumulatively contribute to reduced caloric intake over the experimental period, thereby enhancing the anti-obesity effect. Also, the comparable efficacy between WM and WMS may result from matrix components (fibers, phenolics, minerals) influencing L-citrulline bioavailability and NO-related natriuresis. Overall, the main mechanistic focus was placed on the Na^+^/K^+^-ATPase-dependent regulation of fluid and energy homeostasis [51], whereas dietary salt and the citrulline contained in the immature watermelon rind extract were considered auxiliary modulators and markers that may act synergistically along this axis. In other words, the extract was not intended to isolate the effect of citrulline alone but rather to evaluate how a potassium- and citrulline-rich matrix could support Na^+^/K^+^-ATPase-mediated sodium handling, mitochondrial function, and energy expenditure, thereby exerting anti-obesity effects through an integrated and complementary mechanism. Future work will include an L-citrulline control and molecular assays. Upon necropsy, visible abdominal fat was observed in all groups except the control group (CON), with reduced fat accumulation observed in the HFD-WM and HFD-WMS groups compared to the HFD-C group. This finding is consistent with the study by Lee et al. [52], in which the administration of plant-based salt (Salicornia) significantly reduced body weight and weight gain in the HHS group compared to the HH control group. After six weeks of test substance administration and diet intervention, measurements of epididymal and retroperitoneal fat weights showed a reduction in retroperitoneal fat in the HFD-WM and HFD-WMS groups compared to the HFD-C group, and a significant decrease was observed relative to the CON group. Interestingly, several parameters (body weight, visceral fat mass, and NASH score) were slightly lower in the HFD-S group than in the HFD-C group, suggesting that high-salt supplementation alone may induce mild diuretic or osmotic effects [53]. However, WM and WMS supplementation produced more consistent and pronounced improvements than refined salt, indicating that rind-derived bioactive components, rather than sodium per se, contribute to the observed anti-obesity and hepatic benefits [54,55]. Although significant reductions in body and liver weights were observed, no OGTT or IPGTT was conducted. Consequently, this study cannot address whether WM or WMS improved glucose tolerance or insulin sensitivity. Future investigations should incorporate these metabolic tests to determine whether anti-obesity effects are accompanied by improved glucose regulation. Lipid metabolism is essential for energy storage and distribution, regulation of glucose metabolism, and maintenance of energy homeostasis [56]. Adipose tissue is heterogeneously distributed in multiple depots that differ in physiological function and metabolic risk. Visceral white adipose tissue (vWAT) is strongly linked to metabolic disease, whereas subcutaneous WAT (scWAT) is comparatively less harmful and may even exert protective effects [57,58]. Brown adipose tissue (BAT) dissipates energy via UCP1-mediated thermogenesis and is a recognized target for obesity intervention [59,60]. Notably, our study evaluated only liver and epididymal WAT (a rodent visceral depot) and did not examine scWAT or BAT. This limitation means that the metabolic impact of WM and WMS on other fat depots remains unknown. Future work should include assessments of scWAT, multiple visceral depots, and BAT to provide a more comprehensive understanding of the anti-obesity and metabolic effects of these extracts. Our findings align with evidence that habitual high sodium intake is associated with greater adiposity and central overweight/obesity in humans, and with emerging data linking high salt to visceral adipose dysfunction [55,61]. Hyperlipidemia, characterized by elevated levels of cholesterol, triglycerides, phospholipids, and free fatty acids in the blood, is a major cause of atherosclerosis, which underlies angina, stroke, aneurysms, and hypertension [62]. In this study, the levels of triglycerides and total cholesterol, key indicators of this condition, were lower in the HFD-WM and HFD-WMS groups compared to the HFD-C and HFD-S groups. Figueroa et al. (2019) [63] demonstrated that watermelon juice supplementation in postmenopausal women reduced arterial stiffness and improved vascular function. Improved vascular health enhances systemic circulation, promoting metabolic activity, and potentially supporting long-term weight management. In our results, serum sodium (Na^+^) levels were elevated by 43.2 ± 7.6% in the WM and WMS groups compared to the S group; however, the increase in the WMS group was significantly lower at 21.5 ± 6.6%. The diuretic and natriuretic potential of WM and WMS is supported by evidence that potassium and citrulline enhance renal sodium excretion through NO-mediated vasodilation, increased renal blood flow, and modulation of Na^+^/K^+^-ATPase-dependent tubular sodium transport. Although urinary Na^+^ and K^+^ excretion was not measured, the serum sodium profile observed in this study represents a complementary physiological indicator, as stabilization or reduction of serum sodium during sodium-containing extract supplementation suggests improved renal sodium handling and reduced extracellular fluid expansion. These findings are consistent with the known natriuretic mechanisms of potassium- and citrulline-rich foods. Also, their study showed that a low-sodium diet decreased serum triglyceride levels to 73.8 ± 12.5 mg/dL compared to 89.4 ± 27.3 mg/dL in the high-sodium group, which aligns with our findings regarding WMS administration. High sodium intake has been reported to increase plasma leptin levels and activate enzymes involved in lipogenesis, leading to adipocyte hypertrophy and visceral fat accumulation. These changes contribute to insulin resistance in adipose tissue, increasing the risk of type 2 diabetes and cardiovascular diseases [9,64,65]. Collins et al. (2017) [66] found that citrulline from watermelon extracts exhibited anti-inflammatory effects, contributing to improved metabolic function. Reducing inflammation is known to promote the secretion of beneficial adipokines such as adiponectin, thereby enhancing lipid metabolism. The present study examined a 6-week treatment period, which is relatively short for assessing long-term safety and efficacy. Thus, the durability of the anti-obesity effects of WM and WMS and their chronic safety profile remain unknown. Future research should include prolonged consumption trials, chronic toxicity assessments, and long-term follow-up to determine whether the benefits are sustained and safe. Recent systematic reviews of plant-derived anti-obesity agents underscore the necessity of such longer-term evaluations [67]. Currently, FDA-approved anti-obesity drugs include lipase inhibitors such as Orlistat and serotonin-norepinephrine reuptake inhibitors like Sibutramine. However, these drugs are associated with adverse effects, including steatorrhea, nausea, vomiting, headaches, constipation, insomnia, and gastrointestinal disturbances [68]. The results of this experiment are expected to reflect the citrulline and diuretic effects in WM and WMS, which contain acid salts. Although WM contained more than twice the citrulline content of WMS, their bioactivities were comparable. Although intestinal flora and metabolic signaling pathways were not directly assessed, the existing literature suggests mechanistic pathways consistent with our findings. Potassium-rich extracts may enhance renal Na^+^ excretion via Na^+^/K^+^-ATPase modulation, while citrulline stimulates NO-mediated mitochondrial fatty acid oxidation and increases ATP turnover. Additionally, plant-derived amino acids can influence gut microbial composition and reduce metabolic inflammation [69]. These mechanisms collectively provide theoretical support for the observed reductions in adiposity and lipid profiles. Future investigations will include an L-citrulline control group and examine gene/protein targets involved in citrulline pathways to better elucidate the mechanism. Despite the significant metabolic improvements observed in this study, several limitations should be acknowledged. First, the present findings are based on a high-fat diet-induced obesity model in mice, which, although widely used, does not fully recapitulate the complexity and heterogeneity of human obesity. Species-specific differences in metabolism, gut microbiota composition, sodium and potassium handling, and nitric oxide-related signaling pathways may influence the physiological responses to watermelon-derived extracts. Therefore, caution should be exercised when extrapolating these results directly to human physiology. Further studies, including well-designed clinical trials, are warranted to validate the translational relevance of the observed anti-obesity and metabolic effects in humans.
5. Conclusions
This study demonstrated that immature watermelon extract (WM) and watermelon salt (WMS), when administered to HFD-induced obese models, significantly reduced body weight, abdominal fat accumulation, retroperitoneal fat weight, and blood lipid levels, including triglycerides and total cholesterol. The increased sodium excretion observed in the WM and WMS groups, likely due to their citrulline and potassium content, may contribute to improved lipid metabolism. These results suggest that WM and WMS have potential as natural anti-obesity agents by promoting fat reduction and metabolic health.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mokdad A.H. Bowman B.A. Ford E.S. Vinicor F. Marks J.S. Koplan J.P. The continuing epidemics of obesity and diabetes in the United States JAMA 20012861195120010.1001/jama.286.10.119511559264 · doi ↗ · pubmed ↗
- 2World Health Organization The Statistics of Obesity Available online: https://www.who.int/health-topics/obesity#tab=tab_1(accessed on 20 May 2023)
- 3Cardel M.I. Atkinson M.A. Taveras E.M. Holm J.C. Kelly A.S. Obesity treatment among adolescents: A review of current evidence and future directions JAMA Pediatr.202017460961710.1001/jamapediatrics.2020.008532202626 PMC 7483247 · doi ↗ · pubmed ↗
- 4Bray G.A. Medical consequences of obesity J. Clin. Endocrinol. Metab.2004892583258910.1210/jc.2004-053515181027 · doi ↗ · pubmed ↗
- 5Sharma A.M. Adipose tissue: A mediator of cardiovascular risk Int. J. Obes.200245710.1038/sj.ijo.080221012457291 · doi ↗ · pubmed ↗
- 6Björntorp P. The associations between obesity, adipose tissue distribution and disease Acta Med. Scand. Suppl.198872312113410.1111/j.0954-6820.1987.tb 05935.x 3293356 · doi ↗ · pubmed ↗
- 7Na S.Y. Myung S.J. Obesity and colorectal cancer Korean J. Gastroenterol.201259162610.4166/kjg.2012.59.1.1622289950 · doi ↗ · pubmed ↗
- 8Kyung M.G. Lim J.Y. Lee K.S. Jung S.W. Choe K.B. Yang C.K. Kim Y.R. Effects of short-term supplementation of erythritol-salt on urinary electrolyte excretion in rats J. Nutr. Health 2014479910510.4163/jnh.2014.47.2.99 · doi ↗