Transcription, Alternative Splicing, and Post-Translational Regulation of CaLOXs in the Dynamic Regulation of Jasmonate Levels in Wounded Pepper Leaves
Juliette T. Keith, Yinting Chen, Jennifer Gabriel, Nicole M. van Dam, Jacqueline C. Bede

TL;DR
The study explores how jasmonate levels in pepper leaves change after wounding, focusing on gene expression, splicing, and enzyme regulation.
Contribution
The paper reveals how CaLOX gene variants and splicing events dynamically regulate jasmonate levels in response to plant wounding.
Findings
CaLOX2 shows rapid but short-lived wound-induced expression due to a phosphosite Ser residue.
CaLOX7 has higher wound-induced expression at 6 h due to a non-phosphorylatable Ala residue.
Exon 4 retention in CaLOX8 transcripts may reduce enzyme activity by blocking the active site.
Abstract
In response to stresses, jasmonates increase rapidly, leading to plant resistance against necrotrophic pathogens and chewing insect herbivores. Jasmonate biosynthesis is regulated at many levels, including transcriptionally, through alternative splicing, and the phosphorylation of the 13S-lipoxygenase (LOX) that catalyzes an early step in jasmonate biosynthesis. In pepper, transcriptomic analysis of a foliar wounding time course was conducted to deepen our understanding of these regulatory mechanisms. All four CaLOXs are constitutively expressed. CaLOX2, which encodes an enzyme with a Ser in a predicted regulatory phosphosite, shows a rapid but short-lived increase in wound-induced expression. In contrast, CaLOX7, which encodes a protein with a non-phosphorylatable Ala at the phosphosite, shows higher wound-induced expression at 6 h. As well, at this timepoint, there is a predicted…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
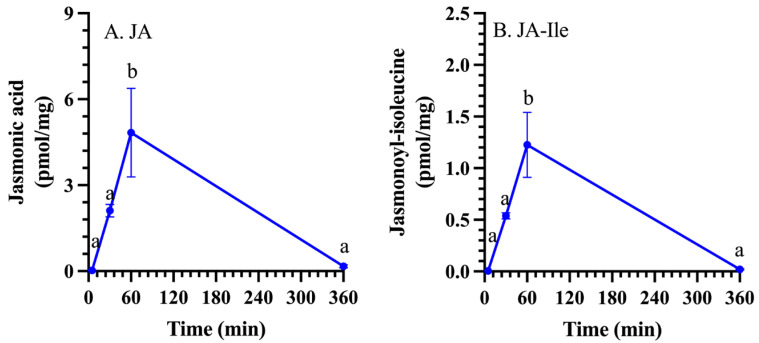 Figure 1
Figure 1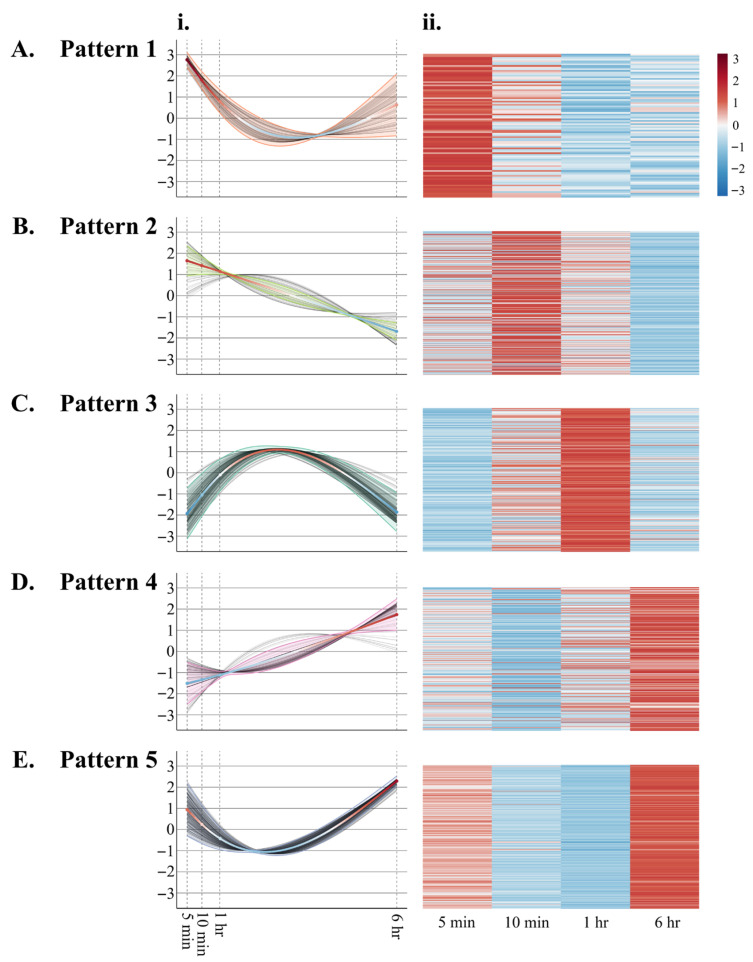 Figure 2
Figure 2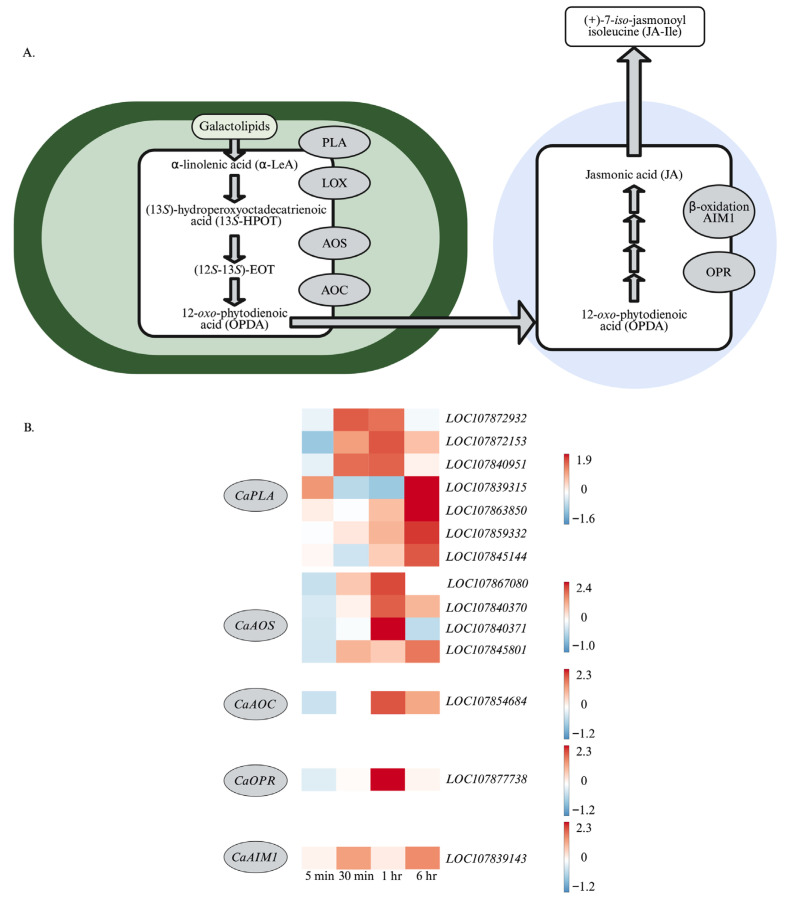 Figure 3
Figure 3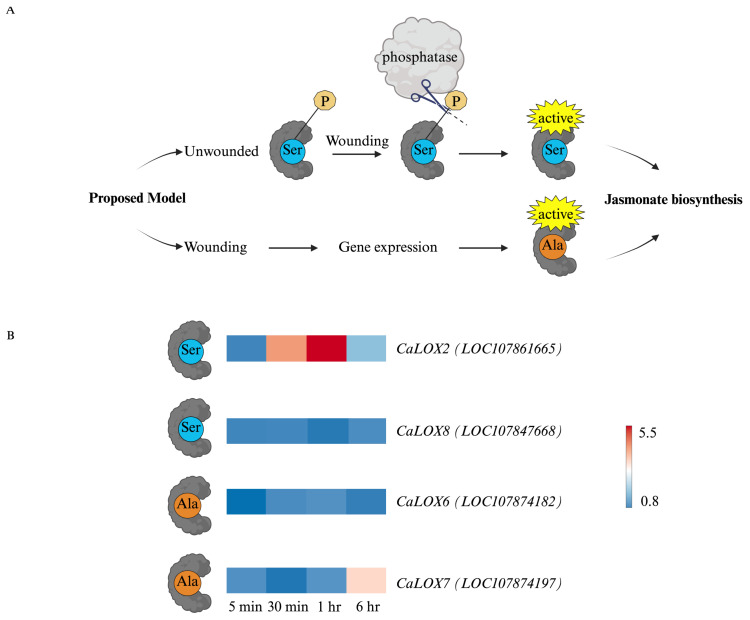 Figure 4
Figure 4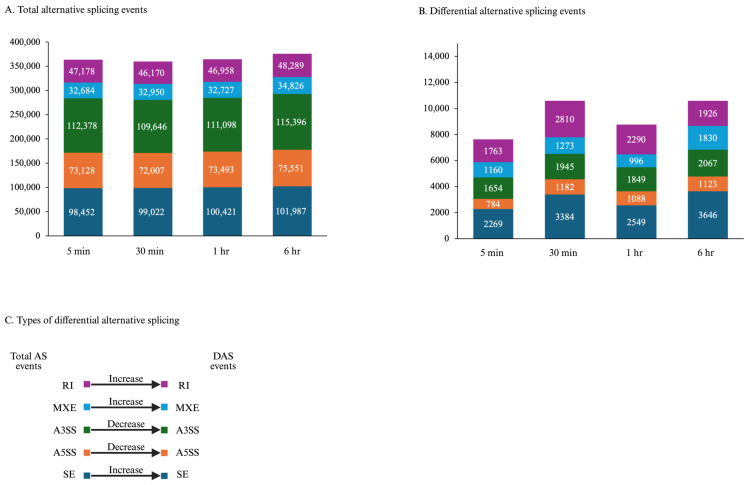 Figure 5
Figure 5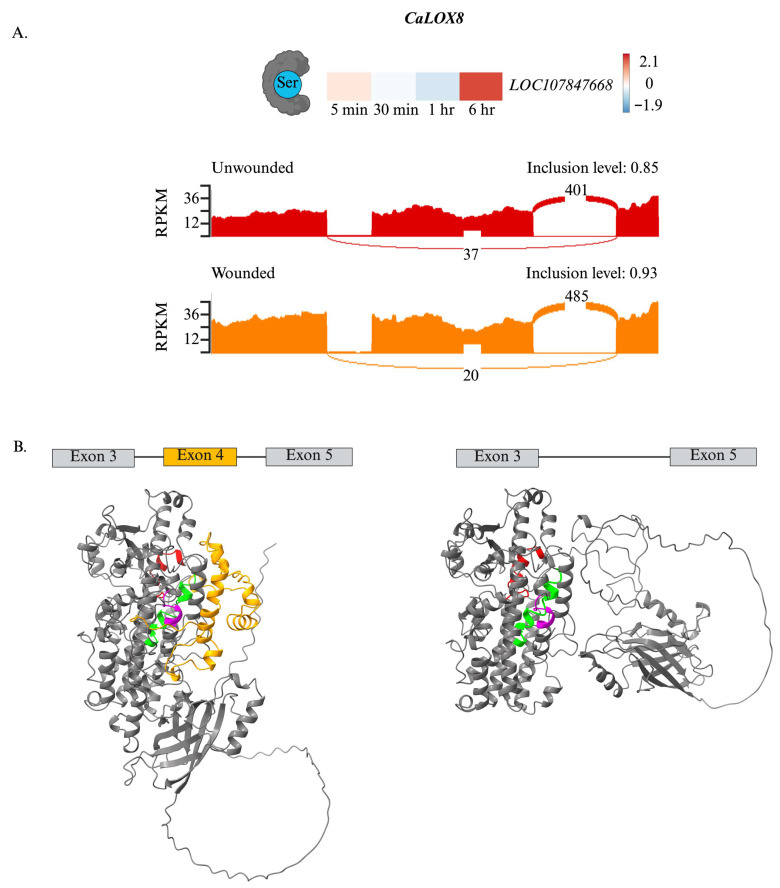 Figure 6
Figure 6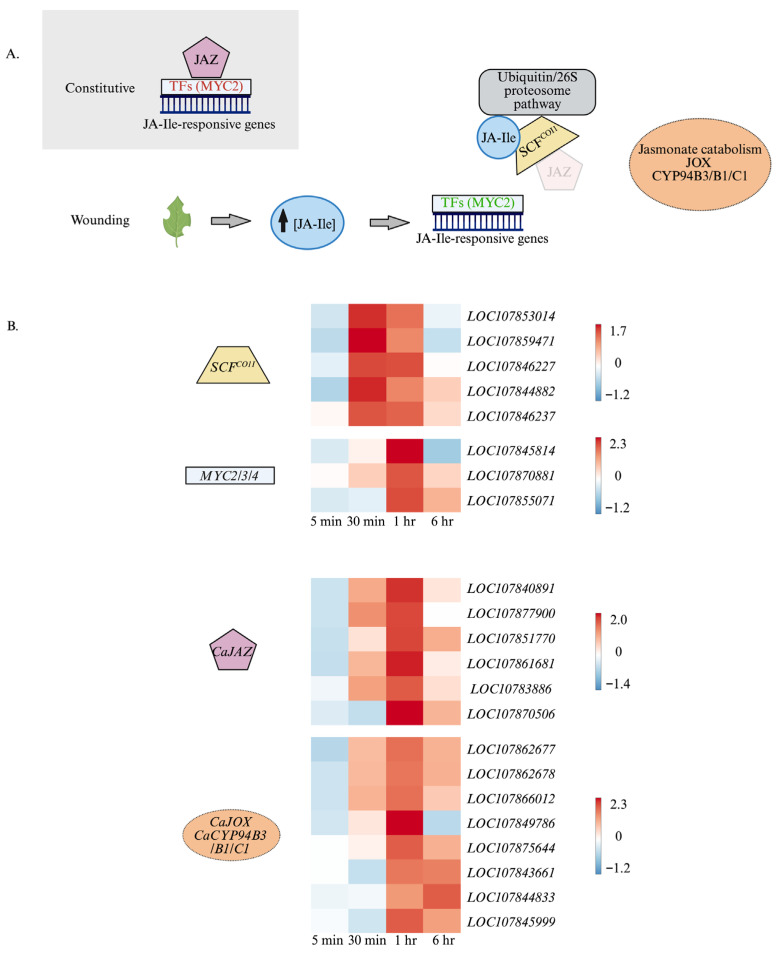 Figure 7
Figure 7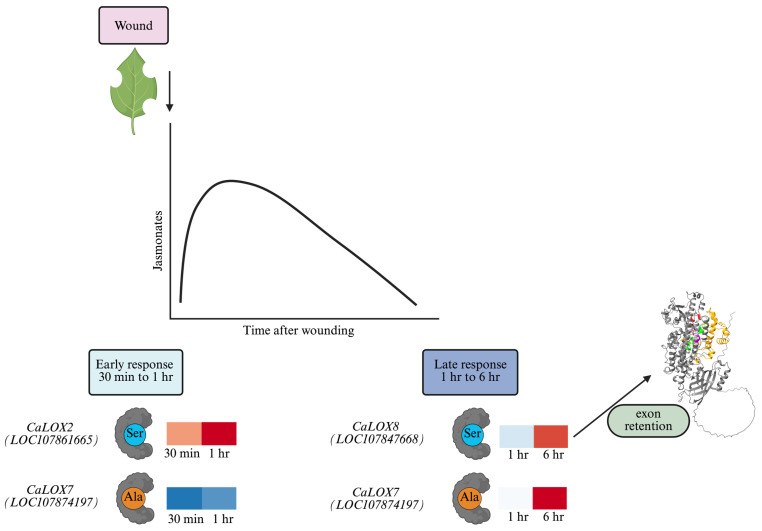 Figure 8
Figure 8- —German Research Foundation
- —Natural Science and Engineering Research Council (NSERC) Discovery
- —Fonds de recherche du Québec—nature et technologies (FRQNT) center SÈVE
- —Walter M. Stewart Postgraduate Scholarship in Agriculture
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect-Plant Interactions and Control · Plant-Microbe Interactions and Immunity · Plant Parasitism and Resistance
1. Introduction
Jasmonates are a family of oxylipin signaling molecules that regulate plant development processes, including flowering time and senescence [1,2,3], and stress responses [4,5,6]. In response to mechanical damage, such as that incurred by chewing insect herbivory, jasmonate signaling gives rise to approximately 95% of the wound-induced protein synthesis that leads to the production of protective plant specialized metabolites and morphological barriers [7,8]. Given the importance of this pathway in plant resistance to pathogens and insect herbivores, understanding its regulation is key and may lead to novel strategies to enhance crop productivity [9].
After wounding or recognition of a necrotrophic pathogen, jasmonate biosynthesis starts in the chloroplast, where α-linolenic acid, a substrate of lipoxygenases (LOXs), is released from the galactolipids of the chloroplast membrane by a galactolipase [10]. The fatty acid undergoes oxidation, catalyzed by a member of the 13S-LOX class of lipoxygenases, producing (13S)-hydroperoxy-octadecatrienoic acid (13-HPOT) [11]. At this point in the pathway, different oxylipins may be produced. The reaction that converts 13-HPOT to (12S-13S)-epoxy-hydroperoxy-octadecatrienoic acid ((12S-13S)-EOT) is catalyzed by allene oxide synthase (AOS) and commits the pathway towards jasmonate biosynthesis [12]. (12S-13S)-EOT is reconfigured by allene oxide cyclase (AOC) into the more stable (9S,13S)-12-oxo-phytodienoic acid (OPDA), a bioactive jasmonate [13,14,15]. OPDA is transported from the chloroplast into the cytosol and then into the peroxisome [16,17]. In this organelle, jasmonate biosynthesis continues with the reduction of OPDA by OPDA reductase 3 (OPR3) [18,19,20]. After esterification to CoA [21], OPC-8 undergoes β-oxidation by the fatty acid β-oxidation machinery composed of acyl-CoA oxidase (ACX), multifunctional protein (MFP), and 3-ketoacyl CoA thiolase (KAT) [22,23,24,25,26,27]. The product, jasmonoyl-CoA, is transformed into jasmonic acid (JA), which moves into the cytosol [28,29,30]. There, it may be converted into its biologically active form, jasmonoyl-isoleucine (JA-Ile), by jasmonate resistant 1 (JAR1) [31].
After entering the nucleus [32], JA-Ile binds to the Skp1/Cullin1/F-box protein coronatine-insensitive 1 (SCF^CO1^) complex, bridging the ubiquitin E3 ligase with its target repressor jasmonate ZIM-domain (JAZ) proteins, leading to its degradation through the 26S-proteasome, thus freeing MYC2, MYC3, or MYC4 transcription factors from repression and allowing JA-Ile-responsive gene expression to proceed [32,33,34,35,36]. Targets of these transcription factors include genes that encode enzymes in the jasmonate biosynthetic pathway, leading to feedforward regulation [37].
In unwounded leaves, the basal JA level in Arabidopsis thaliana (arabidopsis) is less than 30 pmol/gm fresh mass [38]. After wounding, JA levels increase within seconds after mechanical damage, reaching levels that are over 10-fold higher than those of unwounded leaves within 2 min. This response is too rapid to be attributed to the increase in the expression of genes encoding enzymes in the jasmonate biosynthesis pathway. This indicates that the pathway must be constitutively present and activated upon damage [10,39,40]. Thivierge et al. [41] found that AtLOX2, which catalyzes an early step in jasmonate biosynthesis, is constitutively phosphorylated on Serine^600^ (Ser^600^) and dephosphorylated in response to wounding. Using phosphovariants, in vitro studies confirmed that the unphosphorylatable AtLOX2 had over a 100 times increase in V_max_ compared to the phosphomimic, which likely reflects blocking of the substrate from the catalytic site [42]. Therefore, wounding stress is proposed to lead to the dephosphorylation of AtLOX2, contributing to the rapid increase in JA-Ile biosynthesis.
In arabidopsis AtLOX2, the phosphosite, where the Ser^600^ is shown in red, is flanked by the amino acid sequence ARQSLVNG [42]. The sequence ARQSLVNG is relatively conserved among plant 13S-LOXs; however, there are variations at the phosphosite Ser, where Ala or Ile occupy this position, generating non-phosphorylatable variants. We chose pepper (Capsicum annuum) as our study organism since pepper has four 13S-LOXs [43]; CaLOX2 and CaLOX8 have a Ser at the phosphosite, whereas CaLOX6 and CaLOX7 have an unphosphorylatable Ala at this site, suggesting that these proteins may be regulated at the post-translational or transcriptional levels, respectively (Note: Pepper LOXs have been color-coded; those with an Ala at the phosphosite (CaLOX6 and CaLOX7) in orange and those with a Ser at this site (CaLOX2 and CaLOX8) in blue).
Alternative splicing (AS) also plays an important role in post-transcriptional regulation of stress-related gene expression in plants [44]. In tea, Camellia sinensis, six out of eleven LOXs have splice variants [45]. The splice variants of CsLOX3 and CsLOX1 contained premature stop codons, suggesting they are targeted for nonsense-mediated decay, while the splice variants of CsLOX2, CsLOX5, CsLOX9, and CsLOX10 are proposed to encode truncated proteins without specific functional domains. For example, AS of CsLOX5 and CsLOX10 removes the oxygen-binding domain of these enzymes, producing truncated proteins without an oxygenase function. These nonactive splice variants are proposed to interfere with functional LOXs by competing for transcriptional machinery or for substrates at the protein level. The splice variants in tea do not appear to affect the predicted phosphosite based on the AtLOX2 amino acid sequence.
As previously mentioned, JAZ proteins are negative regulators of jasmonoyl-isoleucine-responsive gene expression [33,34,35]. As a means of negative feedback regulation, genes encoding these proteins are induced in response to wounding [8]. Splice variants of JAZ proteins desensitize the plant to jasmonate signaling [46,47]. These splice isoforms are able to bind to the MYC transcription factors; however, they lack the C-terminal jas motif and, thus, cannot be targeted for degradation, even in the presence of high JA-Ile levels.
The above highlights that there are possible multiple levels of regulation that shape jasmonate dynamics upon wounding; however, the underlying mechanisms are not fully understood. In this study, we are interested in the transcriptional expression of genes encoding jasmonate biosynthetic enzymes, the expression pattern of LOX genes that encode either a Ser or Ala at the regulatory phosphosite in the mature protein, and alternative splicing events of LOX and JAZ genes. Thus, we conducted a time course of wound responses in pepper leaves to deepen our understanding of the regulation of the jasmonate pathway.
2. Results
2.1. Phytohormone Levels in Wounded Pepper Leaves
Foliar JA and JA-Ile levels rose sharply in the first hour after wounding and then returned to basal levels at 6 h post-damage (Figure 1, Supplemental Table S1).
2.2. Wound-Induced Gene Expression in Pepper Leaves
After processing and removal of poor-quality reads, an average of 103.97 million 100 bp reads per sample was obtained with a Pfred score of 39 or above (Supplemental Table S2). The high-quality reads aligned with the NCBI C. annuum genome assembly UCD10Xv1.1 (48,341 annotated genes) with an average mapping efficiency of 91.6% uniquely mapped reads. One thousand three hundred and forty-five (1345) genes, representing 5.3% of the total expressed genes, were wound-responsive at one or more timepoints over the experimental time course; of these, 1141 genes (84.8%) and 205 genes (15.2%) were upregulated or downregulated, respectively, in wounded leaves (Supplemental Figure S1).
Wound-responsive genes clustered into 5 main patterns (Figure 2, Supplemental Table S3): Pattern 1: Genes that show a repressed gene expression after wounding; Pattern 2: Genes that show highest expression at 30 min post-wounding; Pattern 3: Genes that show highest expression 1 h post-wounding; Pattern 4: Genes that show highest expression 6 h post-wounding; and Pattern 5: Genes that show dynamic expression initially decreasing but then showing the highest expression at 6 h post-wounding.
2.3. Wound-Induced Expression of Genes Encoding Enzymes in Jasmonate Biosynthesis in Pepper Leaves
Within 30 min after wounding, early and sustained expression of genes encoding early enzymes in the jasmonate biosynthetic enzymes, including chloroplastic phospholipases (phospholipase A1 (LOC107872932), CaDAD1 (LOC107872153), CaPLIP2 (LOC107840951)) and allene oxide synthase1 (CaAOS1 (LOC107867080)) (Figure 3). At one hour post-wounding, genes encoding CaAOS3 (LOC107840370, LOC107840371), allene oxide cyclase (CaAOC (LOC107854684)), and 12-oxo-phytodienoic acid reductase3 (CaOPR3 (LOC107877738)) are strongly expressed. Later wound-induced jasmonate biosynthetic genes include CaAOS3 (LOC107845801), peroxisomal CaAIM1 (LOC107839143), and chloroplastic phospholipases (LOC107839315, LOC107863850, LOC107859332), including CaFAR2 (LOC107845144).
Focusing on pepper LOXs, we were interested in the expression of genes that encoded proteins with a Ser or an Ala at the phosphosite, predicting that those with a Ser would be constitutively expressed or immediately after wounding, whereas those with an Ala would be expressed later [41,42]. All CaLOXs were constitutively expressed. CaLOX2, which encodes an enzyme with a Ser at the phosphosite, shows rapid but short-lived expression (Figure 4). In contrast, CaLOX7, which encodes an enzyme with Ala at the phosphosite, is highly expressed 6 h after wounding.
2.4. Alternative Splicing of 13S-CaLOXs in Wounded Pepper Leaves
In addition to transcriptional regulation, genes may undergo alternative splicing, potentially resulting in novel proteins or nonfunctional proteins. In undamaged leaves, about 350,000 splicing events are predicted, with the majority being exon skipping (SE) or alternative 3′-splice sites, with little change across timepoints (Figure 5). There is a 2–3% change in splicing events (differential AS (DAS)) in wounded pepper leaves. In these DAS events, an increase in SE, intron retention (RI), and mutually exclusive exons (MXE) with reduced splicing of 3′- or 5′-sites (A3SS and A5SS, respectively).
As with tea CsLOXs [45], CaLOXs also may undergo AS (Figure 6). Notably, a change in CaLOX8 splicing is predicted 6 h post-wounding. In contrast to undamaged plants, there is an 8% predicted higher retention of exon 4, which is the one preceding the exon containing the substrate-binding domain (exon 5), in wounded pepper leaves. Through protein modeling, retention of this exon may affect protein structure, potentially shielding access of the substrate to the active site, negatively affecting enzyme activity (Figure 6B).
2.5. Wound-Induced Expression of Genes Encoding Proteins Involved in Jasmonate Signaling in Pepper Leaves
As nuclear JA-Ile levels increase, JA-Ile bridges the SCF^COI1^ complex to JAZ proteins, resulting in their ubiquitination and degradation by the 26S-proteasome, releasing MCY2, MYC3, and MYC4 transcription factors from repression [33,34,35,36]. Within the first hour after wounding, transcript levels of components of the SCF^COI1^ complex increase, including those encoding F-box proteins (LOC107853014, LOC107859471) and E3-ubiquitin ligases (CaPUB22 (LOC107846227), CaPUB23 (LOC107844882, LOC107846237)). One hour after damage, wound-induced CaMYC2 (LOC107845814, LOC107870881) and CaMYC3 (LOC107855071) expression is observed (Figure 7A). A number of WRKY transcription factors, including CaWRKY17 (LOC107854937), CaWRKY22 (LOC107839265), CaWRKY40 (LOC107861978, LOC107876015), and CaWRKY53 (LOC107839290), are induced at early and mid-timepoints in wounded leaves.
2.6. Wound-Induced Expression of Genes Encoding Enzymes in Specialized Metabolism in Pepper Leaves: Non-Volatile and Volatile Compounds
Upregulation of transcripts encoding enzymes involved in non-volatile specialized metabolism was observed in pepper leaves 6 h after wounding. Wound-induced genes include enzymes involved in the biosynthesis of the tropane alkaloid calystegine (tropinone reductase (LOC107850005, LOC107867623)), triterpenoid and steroid biosynthesis (squalene epoxidase (LOC107840150)), β-amyrin synthase (LOC107841772), β-amyrin 28-monooxygenase (LOC107845824, LOC107859002), β-amyrin 11-oxidase (LOC107853307), and potentially involved in terpenoid indole alkaloid biosynthesis (7-deoxyloganetin glucosyltransferases (LOC107840056, LOC107867483) and stemmadenine O-acetyltransferase (LOC107873928)).
Early polyphenolic-related genes upregulated within the first hour after wounding include the transcription factor CaMYB15 (LOC107852599), flavonoid biosynthesis-related genes (trans-cinnamate: CoA ligase (LOC107859652), flavonol 4′sulfotransferase (LOC107845791), anthocyanidin 3-O-glucosyltransferase 2 (LOC107860695), and agmatine hydroxycinnamoyl transferase 1 (LOC107848097)), and the lignin biosynthetic genes (caffeoyl shikimate esterase (LOC107848284), caffeoyl-CoA O-methyltransferases (LOC107859652, LOC107860279), and the peroxidase (LOC17866097) [49,50,51,52,53,54,55]. Later wound-induced transcript expression includes genes that encode enzymes in flavonoid biosynthesis (flavonoid 3′-hydroxylase CYP75B137 (LOC107867152, LOC107867141) and vestitone reductase (LOC107845154)) and lignin biosynthesis (peroxidases (LOC107840027, LOC107867619, LOC107842218, LOC107843551, LOC107860325, LOC107874093, LOC107863246, LOC107866096, LOC107866094, LOC107860938), dirigent protein 22 (LOC107849702), laccases (LOC107872619, LOC107874615)) [56,57,58].
A number of pepper genes that encode enzymes related to volatile biosynthesis are upregulated in response to wounding. Terpenoid genes are induced 1 h ((R)-linalool synthase (LOC107840096; monoterpenoid biosynthesis); (3S,6E)-nerolidol synthase (LOC107845646; sesquiterpenoid biosynthesis); geranylgeranyl diphosphate synthase (LOC107856323; diterpenoid biosynthesis)) and 6 h (1-deoxy-D-xylulose-5-phosphate synthase 2 (LOC107850768), 2C-methyl-D-erythritol 4-phosphate pathway; neomenthone dehydrogenase (LOC107839158, monoterpenoid biosynthesis); (3S,6E)-nerolidol synthase (LOC124892657, LOC107845646; sesquiterpenoid biosynthesis))) after foliar wounding. As well, a downregulation of eugenol synthase (LOC107870192, monoterpenoid biosynthesis) is also observed at 6 h post-damage. The wound-induced upregulation of acetyl-CoA benzyl alcohol acetyl transferases (1 h: LOC107866161; 6 h: LOC107879437) may result in an increase in volatile benzyl ester emissions [59]. The wound-induced expression of 9-divinyl ether synthase (30 min: LOC107840369) and acetyl-CoA benzyl alcohol acetyl transferases (1 h: LOC107866161; 6 h: LOC107879437) may result in an increase in volatile 9-divinyl ether and benzyl ester emissions [59,60].
2.7. Wound-Induced Expression of Classical Jasmonate Marker Genes in Pepper Leaves
Two groups of classic jasmonate-responsive gene markers [61,62,63], proteinase inhibitors (LOC107866028, LOC107864939, LOC107862777, LOC107866027, LOC124897129, LOC124897130, LOC107843348, LOC107879783, LOC107879784), and defensins (LOC107852231, LOC124890736, LOC107877537), are expressed late (6 h) after mechanical damage.
2.8. Wound-Induced Expression of Negative Regulators of Jasmonate Signaling
Pepper genes encoding negative regulators of jasmonate signaling are expressed as early as 30 min post-wounding. Genes encoding enzymes involved in jasmonate catabolism ultimately result in lower available JA-Ile. Cytochrome P_450_ enzymes B1/B3 and C1 are involved in the consecutive oxidation of JA-Ile to the 12-OH and 12-COOH, respectively [64,65,66]. Cytochrome P_450_ B1/B3 genes (LOC107862677, LOC107862678, LOC107866012) increase at 30 min, peaking at 1 h and returning to lower but still wound-induced levels at 6 h, whereas the genes that encode cytochrome P_450_ 94C1 are induced at 1 h (LOC107849786; LOC107875644) or 6 h (LOC107843661) post-damage. Genes encoding CaJOX4 (LOC107845999) and CaJOX3 (LOC107844833), enzymes that oxidize jasmonic acid [67], are expressed 1 and 6 h after wounding, respectively (Figure 7B). A late-expressed gene encodes methyl jasmonate esterase 1 (LOC107840334) that catalyzes the conversion of methyl jasmonate to jasmonic acid [68]. Even if JA-Ile levels increase, this increase can be muted by JAZ repressors that bind to MYC2, MYC3, and MYC4 transcription factors [33,34,35]. Expression of most wound-induced CaJAZ genes begins as early as 30 min with highest levels at 1 h and returning to basal levels at 6 h post-wounding (CaTIFY10A (LOC107840891, LOC107877900), CaTIFY10C (LOC107861681), CaTIFY5A (LOC107838868)), with the exception being CaTIFY10B (LOC107851770) and CaJAZ7 (LOC107870506) that are only wound-induced at 1 h [69] (Figure 7B).
In arabidopsis, splice variants of JAZ desensitize the plant to jasmonate signaling [46,47]. These variants lack the C-terminal jas motif; therefore, they can bind and repress the MYC transcription factors but cannot be targeted for degradation, and, thus, repression of jasmonate signaling is maintained regardless of high JA-Ile concentration. In wounded pepper leaves, over this time course, we did not observe alternative splicing of JAZ genes over the time course of our experiment.
3. Discussion
In response to foliar wounding, pepper JA and JA-Ile levels increased rapidly to peak around 1 h post-damage and then returned to basal levels (Figure 1). This dynamic fluctuation reflects the regulation of jasmonate biosynthesis and jasmonate signaling. In this study, we focused on understanding the potential role of transcription, alternative splicing, and, indirectly, post-translational phosphorylation through the presence of a phosphosite on the regulation of pepper 13S-LOXs that catalyze an early step in jasmonate biosynthesis (Figure 8).
3.1. Jasmonate Profile Reflects Genes Involved in Jasmonate Biosynthesis, Signaling and Repression
In general, a strong upregulation of genes that encode jasmonate biosynthetic enzymes, including phospholipases, CaLOX2, CaLOX7, CaAOS1, CaAOS3, CaAOC, CaOPR3, and CaAIM, is observed (Figure 3 and Figure 4). In arabidopsis, an enzyme early in jasmonate biosynthesis, AtLOX2, has been identified as being involved in the regulation of flux through this pathway. This enzyme is constitutively phosphorylated at Ser^600^ and dephosphorylated in response to wounding [41]. In vitro studies with phosphovariants show that AtLOX2 phosphomimics have basal activity, which increases 10-fold in the phosphonull mutants [42]. Together, this supports a model where enzymes for jasmonate biosynthesis are constitutively present and flux through the pathway reflects substrate availability and post-translational regulation, such as the phosphorylation of AtLOX2 [10,42]. Pepper has four 13S-LOXs that may be involved in jasmonate biosynthesis [43]. Of interest, some of these CaLOXs, CaLOX2 and CaLOX8, have a Ser at their homologous phosphosite, suggesting that they may be regulated through phosphorylation, whereas other CaLOXs, CaLOX6 and CaLOX7, have a non-phosphorylatable Ala at this site. We hypothesize that CaLOX2 and CaLOX8 are constitutively expressed and involved in early stress-associated jasmonate biosynthesis compared to CaLOX6 and CaLOX7, which may be involved in longer, more sustained jasmonate production. In support of this, CaLOX2, which encodes an enzyme with a Ser at the phosphosite, is expressed early after wounding, in contrast to CaLOX7, which encodes an enzyme with an Ala at the phosphosite, that is highly expressed later in the time course (Figure 4). A similar pattern was observed in a previous study by Sarde et al. [70] in response to thrip, Frankliniella occidentalis, infestation or JA application to pepper leaves; they observed early induced expression of CaLOX2 and the later strong expression of CaLOX7. They observed expression of CaLOX6 at 8- and 24 h post-JA treatment, which was longer than the time course conducted in our experiment. Ongoing studies generating transgenic plants with phosphovariants are being conducted to fully understand the role of LOX phosphophorylation at this Ser in arabidopsis and pepper.
In addition to transcriptional and post-translational regulation, genes are also regulated through AS [44]. At each timepoint, approximately 350,000 AS events were identified, with a 2–3% difference in AS events between unwounded and wounded pepper leaves. The dominant types of differential AS events were SE, MXE, and RI, which are consistent with a meta-analysis of stress-related splicing events in pepper [71] (Figure 5).
CaLOX8 is constitutively expressed in pepper leaves and not wound-induced (Figure 4); however, a differential AS exon skipping event was identified for this transcript. In wounded leaves at 6 h, there is a predicted increase in the retention of the 4th exon of CaLOX8, which is the exon preceding exon 5 that contains the substrate-binding domain (Figure 6). Modeling with AlphaFold3 suggests that this may block fatty acid substrate access to the active site, potentially negatively affecting enzyme activity, but this must be confirmed experimentally through enzyme assays with recombinant isozymes and validation of the different isozymes in planta (Figure 6B).
In support of jasmonate signaling, CaMYC2 (LOC107845814, LOC107870881), CaMYC3 (LOC107855071), and components of the CaSCF^COI1^ complex (E3-ubiquitin ligases (CaPUB22 (LOC107846227), CaPUB23 (LOC107844882, LOC107846237); F-box proteins (LOC107853014, LOC107859471)) are upregulated in wounded pepper leaves (Figure 7A) [33,34,35]. Also, genes encoding a number of CaWRKY transcription factors known to be involved in jasmonate signaling are wound-induced. AtWRKY17 is involved in jasmonate × salicylic acid (SA) crosstalk and decreases SA signaling by positively regulating Pseudomonas syringae-induced jasmonate accumulation in arabidopsis [72,73]. AtWRKY40 supports jasmonate signaling by repressing the expression of SA-responsive genes as well as JAZ repressors during infection by the powdery mildew fungus Golovinomyces orontii [74]. AtWRKY53 leads to the expression of AtWRKY22 that negatively affects both SA- and jasmonate-responsive gene expression [75]. Of interest, CaWRKY53 (LOC107839290) is highly expressed at 30 min post-wounding. In arabidopsis, AtWRKY53 interferes with jasmonate signaling by negatively regulating AtLOX3 and AtLOX4 expression (Jiao et al., 2022) [76].
Contributing to the dynamic nature of jasmonate signaling, expression of genes that encode proteins that attenuate jasmonate-signaling is observed at 6 h post-wounding (Figure 2 and Figure 7B). In arabidopsis, cytochrome P_450_ 94B1/B3 catalyzes the oxidation of JA-Ile to 12OH-JA-Ile [64,66]. This is followed by the action of cytochrome P_450_ 94C1 that further oxidizes 12OH-JA-Ile to 12COOH-JA-Ile. Pepper cytochrome P_450_ 94B1/B3 genes are expressed early, peaking at 1 h post-wounding, compared to cytochrome P_450_ 94C1. In arabidopsis, AtCYP94C1 was also induced by wounding and treatment with methyl jasmonate [77].
Jasmonate oxidases (JOXs) catalyze the oxidation of jasmonic acid, removing its availability to be conjugated to isoleucine by JAR1 [31,67,78]. Pepper CaJOX4 and CaJOX3 genes are wound-induced at 1 h, with CaJOX3 expression increasing at 6 h after damage (Figure 7B). Methyl jasmonate esterase I (LOC107840334) is induced 6 h post-wounding and encodes an enzyme that catalyzes the demethylation of methyl jasmonate to jasmonic acid [68]. The expression of genes that encode JAZ proteins, which bind to MYC2, MYC4, and MYC6 transcription factors and repress jasmonate-responsive gene expression, is observed within 30 to 60 min of wounding and decreases to basal levels at 6 h (Figure 7B). Expression of CaJAZ genes was also observed when leaves were treated with jasmonic acid [79]. In arabidopsis, alternative splicing of JAZ transcripts produces proteins without a C-terminus jas motif, thereby reducing their degradation when JA-Ile levels are high and contributing to sustained negative regulation of jasmonate signaling [46,47]. In wounded pepper leaves, we did not observe alternative splicing of JAZ genes over the time course of our experiment.
3.2. Dynamic Jasmonate Levels Are Reflected in Specialized Metabolism Gene Expression
The fluctuations of JA-Ile levels in wounded pepper leaves translate into transcriptional reprogramming and upregulation of genes involved in specialized metabolism. Genes that encoded enzymes involved in non-volatile specialized metabolism biosynthesis were induced in pepper leaves 6 h post-damage. These genes encode enzymes in the general triterpenoid/steroidal pathway, including squalene epoxidase (LOC107840150), which catalyzes the oxidation of squalene to 2,3-oxidosqualene, and β-amyrin synthase (LOC107841772), which converts 2,3-oxidosqualene to β-amyrin [80,81]. In Glycyrrhiza glabra (licorice), β-amyrin is oxidized to 11-oxo-β-amyrin through the action of β-amyrin 11-oxidase [82]. This precursor can take the path to oleanane-type triterpenoid saponins through the action of β-amyrin 2,8-monooxygenase (LOC107845824, LOC107859002, LOC107859002) [80].
Even though pepper is not thought to produce terpenoid indole alkaloids, 7-deoxyloganetin glucosyltransferases (GGTs) (LOC107840056, LOC107867483) and stemmadenine O-acetyltransferase (LOC107873928) were wound-induced. In the Madagascar periwinkle, Catharanthus roseus, 7-deoxyloganetin glucosyltransferase catalyzes the conversion of the iridoid 7-deoxyloganetic acid to 7-deoxyloganic acid, which is enzymatically converted to secologanin through a number of steps [83]. A number of CaGGT genes are known to be encoded in the C. annuum genome [84]. In C. roseus, later in the biosynthetic pathway leading to the indole alkaloid catharanthine, stemmadenine O-acetyltransferase catalyzes the acetylation of stemmadine to stemmadenine acetate [85]. The presence of related enzymes in peppers suggests that similar indole-type alkaloids may be present in pepper leaves.
Tropinone reductase (LOC107850005, LOC107867623) reduces tropinone to pseudotropine in the biosynthesis of the insecticidal tropane alkaloids calystegine [86,87].
Phenylpropanoid compounds are important defensive specialized metabolites, including lignins that are part of structural barriers and flavonoids that confer resistance against insect pests and pathogens [88,89]. Early expression of the gene that encodes the transcription factor CaMYB15 (LOC107852599) positively regulates the expression of genes involved in phenylpropanoid biosynthesis [90,91]. The lignin biosynthesis-related biosynthetic genes caffeoyl shikimate esterase (LOC107848284) and caffeoyl-CoA O-methyltransferase (LOC107859652, LOC107860279) were expressed early after wounding, whereas genes encoding peroxidases, laccases, and dirigent proteins that may be involved in lignin biosynthesis are expressed at 6 h after wounding [49,50,53,56,58,92,93,94]. Flavonoid genes upregulated in response to wounding included flavonol 4′sulfotransferase (LOC107845791), flavonoid 3′-hydroxylase (LOC107867152, LOC107867141), and anthocyanidin 3-O-glucosyltransferase 2 (LOC107860695) [51,54,55,94,95]. Strong upregulation of the genes encoding 2-hydroxyisoflavanone dehydratase (LOC107856532) and vestitone reductase (LOC107856532) is noted 6 h post-wounding. In legumes, 2-hydroxyisoflavanone dehydratase catalyzes the conversion of flavonones to isoflavones. Farther down the isoflavonoid pathway, vestitone reductase catalyzes the NADPH-dependent reduction in (3R)-vestitone to 7,2′-dihydroxy-4′-methyoxylisoflavanol that then can be enzymatically converted into phytoalexins such as medicarpin, vestitol, and sativan [96,97,98,99]. The gene encoding vestitone reductase is strongly upregulated in the roots of resistant pepper infected with the oomycete Phytophthora capsica [57]. The gene encoding agmatine hydroxycinnamoyl transferase 1 (LOC107848097, OC107846882), which is involved in hydroxycinnamic acid aldehyde biosynthesis, is induced in wounded pepper leaves. In many plant species, agmatine hydroxycinnamic acid amides have been shown to play a role in plant defense against pathogens and insect herbivores [100,101,102,103,104,105].
In response to foliar damage, the production and release of plant volatile compounds may convey the presence of insect herbivores to natural enemies [106,107]. Twenty-four hours after infestation by caterpillars of the beet armyworm, Spodoptera exigua, pepper volatile emissions increase 7-fold, and these volatiles include the monoterpene linalool and the sesquiterpene nerolidol [108]. Linalool was also a primary leaf volatile emitted in response to Tetranchus urticae, two-spotted spider mite, infestation [79]. In our study, genes, in general, terpenoid pathways (1-deoxy-D-xylulose-5-phosphate synthase 2 (LOC107850768), 2C-methyl-D-erythritol 4-phosphate pathway), as well as later terpenoid biosynthetic enzymes ((R)-linalool synthase (LOC107840096; monoterpenoid biosynthesis), (3S,6E)-nerolidol synthase (LOC107845646, LOC124892657; sesquiterpenoid biosynthesis) that are involved in linalool and nerolidol biosynthesis are wound-induced. Six hours after foliar damage, expression of neomenthone dehydrogenase (LOC107839158, monoterpenoid biosynthesis), which catalyzes menthone biosynthesis, is observed, which may be involved in resistance to pathogens that potentially could invade the plant through the leaf wound [109]. Divinyl ether synthase genes are upregulated in response to infection of pepper leaves with the Obuda pepper virus (Tobamovirus) [110]. In tobacco, the divinyl ethers colneleic acid and colnelenic acid exhibit potent antimicrobial effects on the oomycete Phytophthora parasitica var. nicotianae [111].
In the late response after wounding, classic jasmonate-responsive genes encoding proteinase inhibitors (LOC107866028, LOC107864939, LOC10782777, LOC107866027, LOC124897129, LOC124879130, LOC107843348, LOC107879783, LOC107879784) and defensins (LOC107852231, LOC124890736, LOC107877537) are strongly upregulated [61,62,63].
3.3. Conclusions
In response to foliar wounding, jasmonate levels regulate plant defense responses. Therefore, understanding their regulation is key to enhancing plant resistance to pathogens and chewing insect herbivores. The dynamic levels of JA and JA-Ile reflect the transcriptional, alternative splicing, and post-translational regulation of CaLOXs (Figure 1, Figure 3, Figure 4, and Figure 8). All CaLOXs are constitutively expressed. In response to damage, CaLOX2, which has a Ser in its regulatory phosphosite, is expressed early after wounding of pepper leaves, supporting the possibility that this protein is regulated by phosphorylation and, in response to wound stress, a phosphatase is activated to dephosphorylate these enzymes, increasing flux into jasmonate biosynthesis. In contrast, wound-induced CaLOX7, which encodes an enzyme with Ala at the predicted phosphosite, is highly expressed at a later timepoint (6 h after wounding) that potentially could contribute to a more long-term sustained jasmonate biosynthesis. The alternative splicing pattern of CaLOX8 may also be affected by wounding; 6 h post-wounding, an 8% increase in the retention of exon 4 is predicted. Modeling suggests that retention of exon 4 may block access of the active site to the substrate, negatively affecting jasmonate biosynthesis (Figure 6). The rapid increase in JA and JA-Ile leads to the expression of genes encoding jasmonate biosynthetic and signaling proteins, including CaPLA1, CaLOX2, CaAOS, CaAOC, CaOPR2, and CaMYC2, 1 h post-wounding (Figure 3 and Figure 7A). At 6 h post-stress, expression of genes encoding JOX enzymes, which convert JA to the inactive 12-OH-JA, and JAZ repressors is observed, which contributes to the lower levels of JA and JA-Ile and wound-associated gene expression observed (Figure 1, Figure 2, and Figure 7B). Given the importance of jasmonates in plant resistance to necrotrophic pathogens and chewing insect herbivores [4,5,6], understanding the regulation of jasmonate biosynthesis is key to enhancing plant protection. These results clarify the possible regulation of LOXs at the transcriptional, splicing, and post-translational levels that lead to the dynamic, stress-associated jasmonate profiles.
4. Materials and Methods
4.1. Plant Maintenance
Three seeds (Capsicum annuum ’Mini Bell’ source: Richters (Goodwood, ON, Canada) were sown per pot (15.2 cm × 11.4 cm, d × h) containing Fafard Agro G6 potting mix (Scotts, Saint-Bonaventure, QC, Canada) and placed in a Conviron growth cabinet (Winnepeg, MB, Canada) at conditions set to emulate the average summer conditions in Montréal, Québec (day: 14 h at 25 °C; ramped to night over 2.5 h; night: 5 h at 22 °C, ramped to day over 2.5 h) with a light intensity of 250 µmoles/m^2^/s. At two weeks, seedlings were thinned to a single plant per pot. Four-week-old plants were used in experiments.
4.2. Wounding Experiment
Once the pepper plants had two sets of fully expanded leaves (~four weeks), a wounding time course was performed. Three days before the experiment, plastic shields were used to separate plant treatments (undamaged vs. damaged) to prevent volatile signaling. At noon, half of the plants were wounded multiple times using a hole punch to damage approximately 12% of the second set of fully unfurled leaves from the apex, avoiding the midvein. Damaged leaves from wounded plants or comparable leaves from undamaged plants were taken at 5 min, 30 min, 1 h, and 6 h after wounding and immediately frozen in liquid nitrogen and stored at −80 °C. These timepoints represent early and late times after wounding. Plants were not resampled; once leaves were removed, the plant was discarded. At each timepoint, leaves were taken for phytohormone and transcriptomic analyses. As well, unwounded and damaged plants were used to determine tissue losses due to wounding by comparing the dry weight (DW) of wounded and unwounded above-ground portions (Supplemental Table S4). The experiment was temporally repeated 4 times.
4.3. Jasmonate Quantification
Acidic jasmonates (OPDA, JA, JA-Ile) quantification was conducted based on Glauser et al. [112]. Pepper leaves were lyophilized, homogenized, and extracted in HPLC-grade methanol:water (70:30) containing a mixture of isotopically-labeled phytohormone internal standards (D6-JA and D6-JA-Ile (HPC Standards)). Following homogenization (Retsch Mill MM 400 (Düsseldorf, Germany)), 10 min, vibration 30 Hz), extracts were centrifuged (18,994× g, 20 min, 20 °C), and the supernatant was transferred to a new tube and evaporated to dryness in a speed-vacuum (Labconco, Kansas City, MI, USA) at room temperature. Using sonication, the pellet was resuspended in methanol/water (70:30) and recentrifuged (18,994× g, 5 min, room temperature) before separation by ultrahigh performance liquid chromatography (UPLC, Waters Acquity, Millford, MA, USA) coupled to a mass spectrometer (MS) (Bruker Elite EvoQ triple-quadrupole, Billerica, MA, USA).
Compounds were separated by UPLC chromatography on a Zorbax Eclipse XDB-C18 column (4.6 × 50 mm, 1.8 μm, Agilent, Santa Clara, MA, USA) maintained at a temperature of 42 °C. The mobile phase went from (A) 5% ACN, 0.1% formic acid to (B) 50% ACN, 0.1% formic acid to (C) 100% ACN, 0.1% formic acid using the following gradient: 30 s. of A, 10 s gradient of A to B, 90 s from B to C, where it was held for 1 min before returning to initial conditions over the next min. All solvents used were LC-MS grade. The flow rate was 400 μL/min, and the column was maintained at a temperature of 42 °C.
After separation, compounds were nebulized by electron spray ionization (negative mode) under the following conditions: capillary voltage 4500, cone 35 arbitrary units (a.u.)/350 °C, probe 60 a.u./475 °C, and nebulizer gas (N_2_) 60 a.u. Data processing was conducted using MS Data Review software (Bruker MS Workstation, vers. 8.2).
Phytohormones were identified based on retention time and the transition m/z (Supplemental Table S5). Foliar phytohormone levels were calculated based on the peak area of the compound-of-interest normalized to the corresponding internal standard divided by the initial dry weight of leaf material.
Four samples were taken per treatment at each timepoint. However, in undamaged samples, JA levels are extremely low and occasionally below detection limits. Thus, a one-way analysis-of-variance (ANOVA) (factor: time) on wounded samples was performed with SPSS (version 29.0.2.0) software. Significant differences were determined by the Tukey honestly significant difference (HSD) post hoc test.
4.4. Transcriptomics: RNA-Seq
Plant tissue (~50 mg) was finely ground in liquid nitrogen using a sterile mortar and pestle and transferred to a pre-cooled Eppendorf tube. Total RNA was extracted using the RNeasy Plant Mini Kit (Qiagen (Hilden, Germany)). Once the liquid nitrogen had evaporated, guanidine thiocyanate (RLT) lysis buffer was added to the tissue and vortexed. The lysate was transferred into a QIAshredder column and centrifuged for two minutes (12,000× g). Ethanol (0.5 volume) was added, and the ethanol/lysate solution was centrifuged (10,000× g) for 45 s in an RNeasy mini spin column in a collection tube. After discarding the flow-through, the column was washed with RW1 and centrifuged (10,000× g, 45 s), this was followed by washing with RPE and centrifugation (10,000× g, 45 s and then 150 s). After each step, the flow-through was discarded. The RNeasy spin column was transferred into a new collection tube, and 30 µL of RNAse-free water was applied directly to the column’s membrane. A final 1 min centrifugation (≥8000× g) eluted the RNA.
RNA concentration and sample purity were determined using a Nanodrop spectrophotometer (ThermoFisher Scientific (Waltham, MA, USA)). To further confirm the quality of the RNA sample, it was separated on a bleach gel [113]. Total RNA was sent for library preparation and next-generation sequencing (100 paired-end reads) at McGill University and Genome Quebec Innovation Centre using the Illumina NovaSeq 6000 platform (San Diego, CA, USA). A polyA-enriched RNA library was used, and a consistent sequencing depth of ~100,000,000 reads was obtained. The Digital Research Alliance of Canada’s Narval Unix-based server was used to store data and to perform computations. After removal of the adaptor and quality control using FastQC [114], reads were aligned to the C. annuum reference genome UCD10Xv1.1 obtained from the National Center for Biotechnology Information (NCBI) using STAR version 2.7.11b [115] (Supplemental Tables S2 and S3A). The raw read data (FASTQ) from this study have been deposited in the NCBI Sequence Read Archive (Bioproject ID #PRJNA1348599).
Differential expression analysis and visualization (i.e., volcano, ridgeline, and heatmap plots) were performed with DESeq2 within the ExpressAnalyst platform [116,117]. DESeq2 was applied to identify wound-induced genes at each timepoint. Differentially expressed genes (DEGs) were filtered with a log fold-change (LogFC) cutoff of above 0.95 or below −0.95 and an adjusted p-value ≤ 0.05 (Supplemental Table S3B).
The expression pattern of DEGs across timepoints (5, 30, 60, and 360 min) was clustered using a smoothing spline clustering framework [118,119]. The raw count data of DEGs were normalized through DESeq2 variance stabilizing transformation [116]. For each gene, fitted cubic smoothing splines were subsequently standardized to z-scores to emphasize patterns over absolute expression. Candidate cluster numbers were evaluated using k-means, and the optimal k was selected according to the maximum average silhouette width.
To identify alternative splicing (AS) events and determine if they are differentially expressed across treatments, the rMATs tool was applied to the alignment files generated by STAR using the NovelSS feature [120]. To increase detection of novel splice junctions, the two-pass mode was applied, where the junctions detected in the first mapping pass are used as annotated junctions in the second pass. The software parameters were set to provide the alignments in both genomic and transcript coordinates, as well as quantification of the number of reads per gene. At each timepoint, wounded samples were compared to unwounded samples to identify differential AS events. The rMATS output was run through the rMATS to sashimiplot tool (version 3.0.0) to visualize differential AS events as sashimi plots [120]. To compare the CaLOX8 protein structure with or without exon 4, AlphaFold3 models were overlaid in ChimeraX [121,122].
4.5. Pepper Lipoxygenases
The pepper Ca13S-LOXs used in this study (CaLOX2 (LOC107861665), CaLOX6 (LOC107874182), CaLOX7 (LOC107874197), and CaLOX8 (LOC107847668)) follow the nomenclature of Sarde et al. [43], who identified these proteins through comparative proteomics, ensuring they had signature lipoxygenase and PLAT/LH2 domains and phylogenetic analyses. CaLOX2 (CAN.G862.55), CaLOX7 (CAN.G649.19), and CaLOX8 (CAN.G1931.1) were also identified as being 13S-lipoxygenases by Oliviera Camargo et al. [123]. CaLOX6 (partial sequence 13-LOXc) and CaLOX7 (13-LOXa) were identified as 13S-lipoxygenases by Juhasz et al. [124]. Throughout this paper, pepper CaLOXs have been color-coded; those with an Ala at the phosphosite (CaLOX6 and CaLOX7) are in orange, and those with a Ser (CaLOX2 and CaLOX8) are in blue.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yuan Z. Zhang D. Roles of jasmonate signalling in plant inflorescence and flower development Curr. Opin. Plant Biol.201527445110.1016/j.pbi.2015.05.02426125498 · doi ↗ · pubmed ↗
- 2Huang H. Liu B. Liu L. Song S. Jasmonate action in plant growth and development J. Exp. Bot.2017681349135910.1093/jxb/erw 49528158849 · doi ↗ · pubmed ↗
- 3Zhao X. Li N. Song Q. Li X. Meng H. Luo K. OPDAT 1, a plastid envelope protein involved in 12-oxo-phytodienoic acid export for jasmonic acid biosynthesis in Populus Tree Physiol.2021411714172810.1093/treephys/tpab 03733835169 · doi ↗ · pubmed ↗
- 4Koo A.J. Metabolism of the plant hormone jasmonate: A sentinel for tissue damage and master regulator of stress response Phytochem. Rev.201817518010.1007/s 11101-017-9510-8 · doi ↗
- 5Howe G.A. Major I.T. Koo A.J. Modularity in jasmonate signaling for multistress resilience Annu. Rev. Plant Biol.20186938741510.1146/annurev-arplant-042817-04004729539269 · doi ↗ · pubmed ↗
- 6Li C. Xu M. Cai X. Han Z. Si J. Chen D. Jasmonate signaling pathway modulates plant defense, growth, and their trade-offs Int. J. Mol. Sci.202223394510.3390/ijms 2307394535409303 PMC 8999811 · doi ↗ · pubmed ↗
- 7Gfeller A. Baerenfaller K. Loscos J. Chételat A. Baginsky S. Farmer E.E. Jasmonate controls polypeptide patterning in undamaged tissue in wounded Arabidopsis leaves Plant Physiol.20111561797180710.1104/pp.111.18100821693672 PMC 3149931 · doi ↗ · pubmed ↗
- 8Kaur D. Schedl A. Lafleur C. Martinez Henao J. van Dam N.M. Rivoal J. Bede J.C. Arabidopsis transcriptomics reveals the role of lipoxygenase 2 (At LOX 2) in wound-induced responses Int. J. Mol. Sci.202425589810.3390/ijms 2511589838892085 PMC 11173247 · doi ↗ · pubmed ↗