Characterization of PDAT Genes in Oat (Avena sativa L.) and the Role of AsPDAT-5C in Lipid Biosynthesis and Abiotic Stress Response
Yan Sun, Jinzhou Yang, Ruirui Hu, Chen Li, Qian Yang, Xiping Sun, Zhiwei Zhang, Runzhi Li, Jinai Xue

TL;DR
This study explores PDAT genes in oat, focusing on AsPDAT-5C's role in lipid production and stress tolerance.
Contribution
The study identifies and functionally characterizes AsPDAT-5C in oat for lipid biosynthesis and abiotic stress response.
Findings
AsPDAT-5C complements a TAG-deficient yeast mutant, confirming its enzymatic activity.
Overexpression of AsPDAT-5C increases lipid content in plants without affecting growth.
AsPDAT-5C enhances tolerance to cold and phosphorus-deficiency stresses.
Abstract
Phospholipid:Diacylglycerol Acyltransferase (PDAT) catalyzes the final step of the acyl-CoA-independent triacylglycerol (TAG) biosynthesis pathway and plays an important role in lipid metabolism and abiotic stress responses in plants. Oat (Avena sativa L.) possesses the highest lipid content among cereal crops, yet the functions of PDAT genes in this species remain largely unexplored. In this study, we identified and characterized three AsPDAT genes in oat, which form a homeologous triplet evenly distributed across the three subgenomes and show high conservation in sequence and gene structure. Phylogenetic analysis indicated a clear divergence between monocot and dicot PDATs. Expression profiling revealed that the three AsPDAT genes share similar organ-specific and stress-responsive expression patterns, suggesting functional conservation following polyploidization, with AsPDAT-5C…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
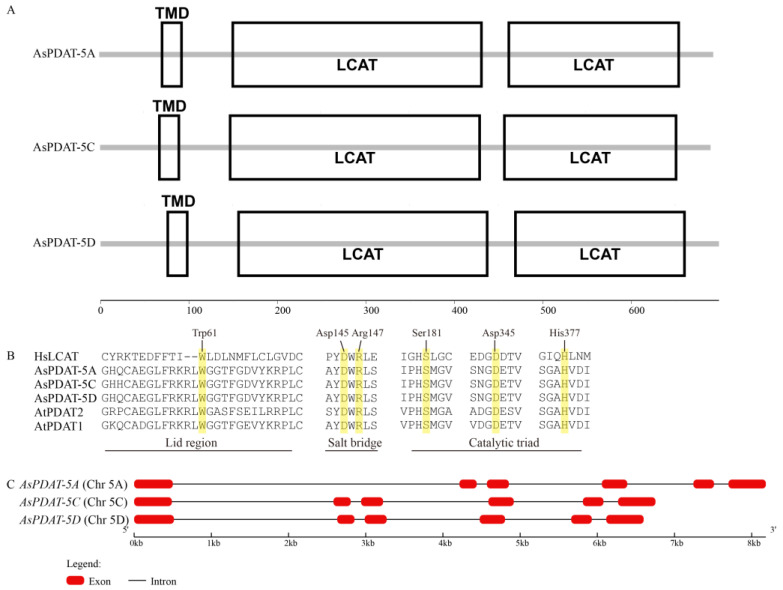 Figure 1
Figure 1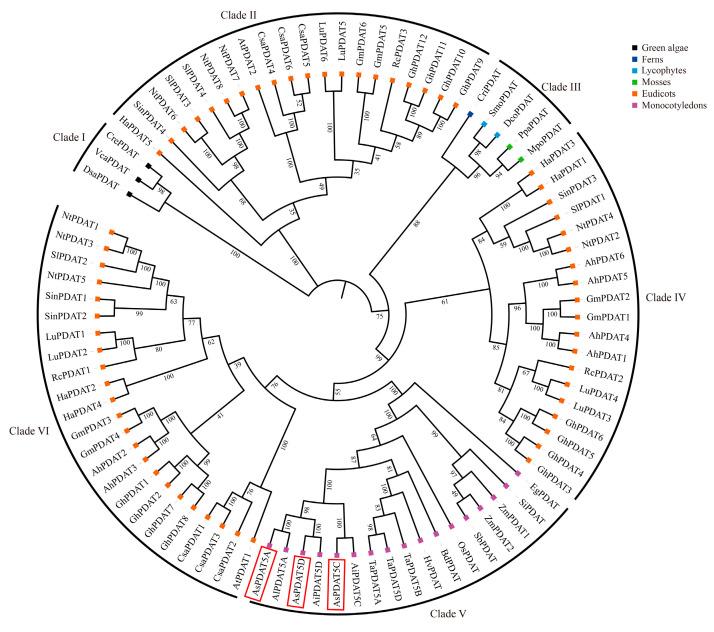 Figure 2
Figure 2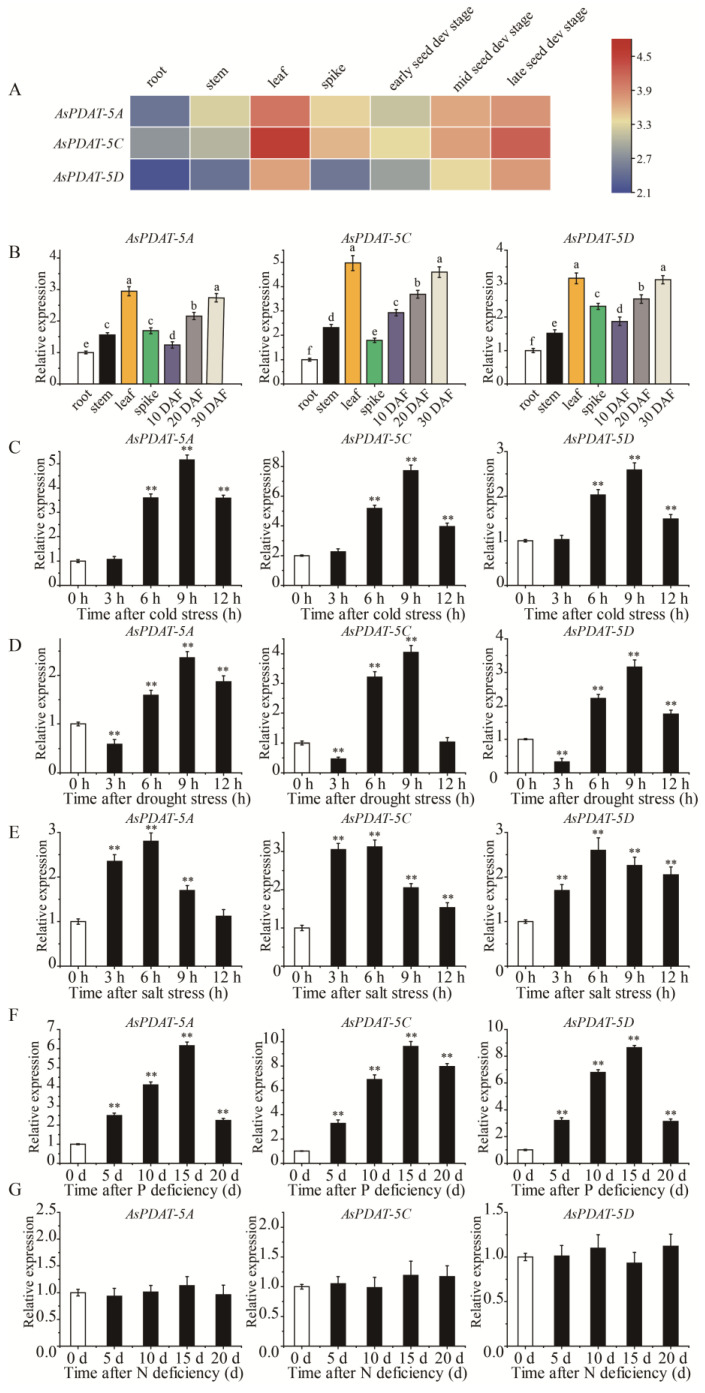 Figure 3
Figure 3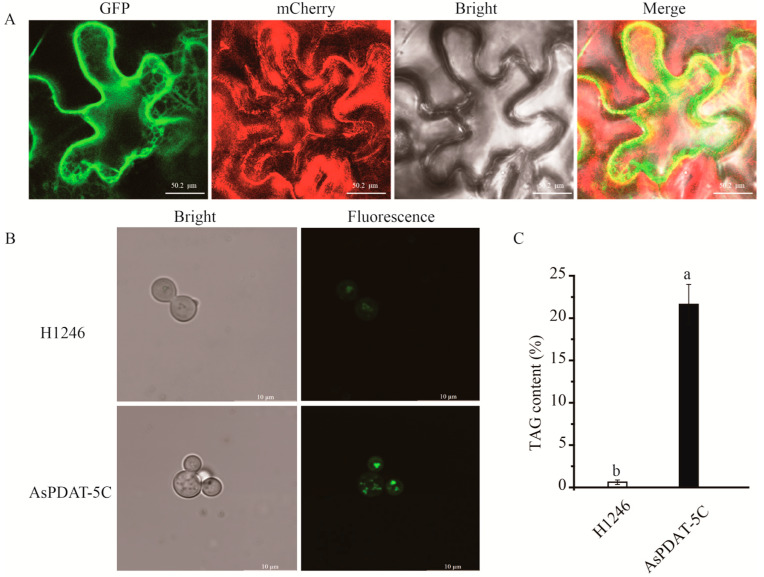 Figure 4
Figure 4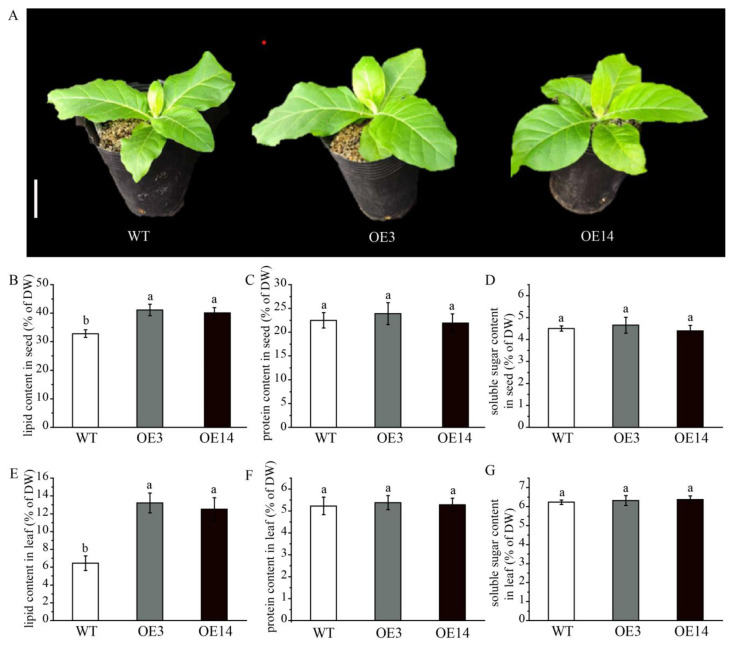 Figure 5
Figure 5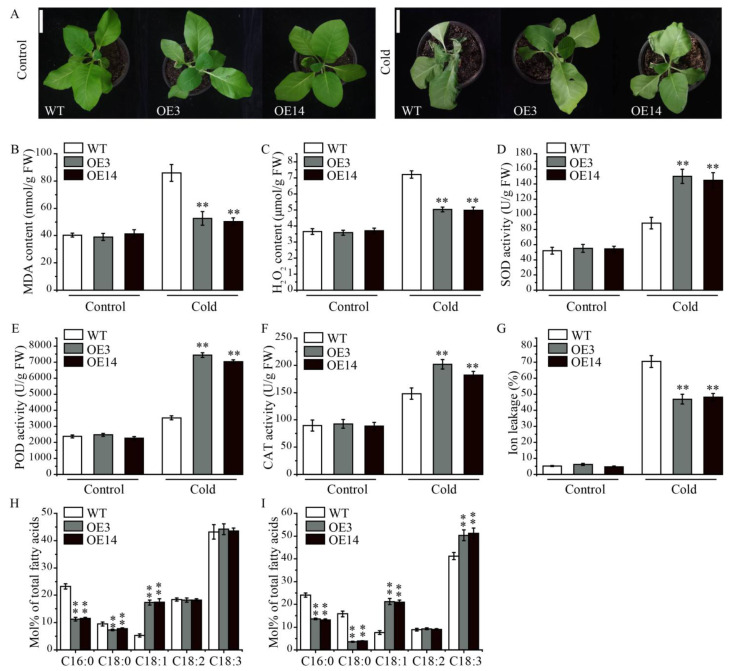 Figure 6
Figure 6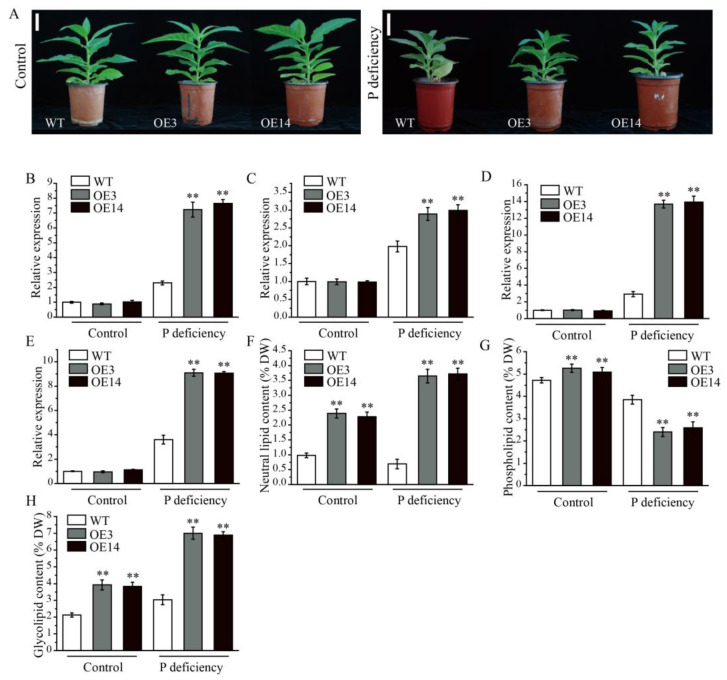 Figure 7
Figure 7- —Postdoctoral Research Start Project of Shanxi Agricultural University
- —Introduction of Talent Research Start-up Fund by Shanxi Agricultural University
- —National Key Research and Development Program of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLipid metabolism and biosynthesis · Plant Surface Properties and Treatments · Peroxisome Proliferator-Activated Receptors
1. Introduction
Plant lipids are an essential nutritional resource for humans and animals and a vital renewable feedstock for biodiesel and industrial applications. [1]. Triacylglycerol (TAG), the predominant storage lipid in mature seeds, is the final metabolic product of fatty acid biosynthesis [1]. The last committed step of TAG assembly is primarily catalyzed by two classes of acyltransferases: Diacylglycerol Acyltransferase (DGAT) and Phospholipid:Diacylglycerol Acyltransferase (PDAT), which utilize acyl-CoA and phospholipids as acyl donors, respectively [2].
PDATs are structurally conserved and share high sequence similarity with mammalian lecithin:cholesterol acyltransferase (LCAT) family members [3]. As previous studies have shown, PDAT genes often exist as multiple copies in plant genomes [4]. The first PDAT gene was cloned from yeast based on sequence homology to human LCAT [5], and PDAT was subsequently identified in the model plant Arabidopsis [6,7] as well as in several oilseed crops and oleaginous microalgae [8,9,10]. So far, genetic modification of PDAT genes has been considered a promising strategy for engineering lipid production. Overexpression and knockout of yeast PDAT (LRO1) supported its indispensable contribution to lipid accumulation at the exponential growth stage, while DGAT2 (DGA1) contributed predominantly during the stationary growth phase [11]. In contrast to the situation in yeast, however, AtPDAT1 activity encoded by At5g13640 was not a major determining factor for seed lipid accumulation, but had overlapping function with AtDGAT1 in normal pollen and seed development in Arabidopsis [12]. Notably, AtPDAT1 was highly expressed not only in seeds but also in vegetative organs, and it has been shown to be crucial for lipid biosynthesis in fast-growing plant organs [13]. Furthermore, heterologous overexpression of SsPDAT1 from Sapium sebiferum increased total lipid content in Brassica napus seeds [14], while overexpression of an endogenous PDAT gene in B. napus resulted in a more modest increase of 1.86–2.77% [15]. In Xanthoceras sorbifolium, expression analyses showed that the XsPDAT1 gene was induced under conditions that positively correlated with lipid accumulation, implicating its involvement in regulating lipid biosynthesis [16]. Although similar responses of PDAT genes in boosting lipid synthesis have also been reported in microalgae, they function primarily under unfavorable environmental conditions [10,17].
Recent studies have also proposed the involvement of PDAT genes in plant tolerance to various abiotic stresses [3]. For example, the overexpression of AtPDAT1 led to substantially increased thermotolerance, and this response was notably compromised in atpdat1 mutants [18]. A recent study further indicated that AtPDAT1 was critical for maintaining membrane homeostasis under cold stress by remodeling lipid metabolism [19]. Beyond temperature stress, PDATs were also thought to be key regulators in response to other environmental perturbations, such as salt, drought, light, and wounding, as suggested by stress-inducible expression patterns of CsPDATs in Camelina sativa and OepPDATs in olive and the association with lipid metabolism [20,21]. Nevertheless, PDAT genes remain functionally characterized in only a limited number of species, and the mechanisms underlying PDAT-mediated stress tolerance are still poorly understood.
Oat (Avena sativa L., 2n = 6x = 42, AACCDD), belonging to the genus Avena within the grass family Poaceae, is the seventh most important cereal crop in terms of worldwide production, and widely used as an energy and nutrient source for human food and animal feed [22]. It exhibits strong environmental adaptability and accumulates abundant metabolites, including soluble dietary fiber, polyunsaturated fatty acids, vitamins, alkaloids, polyphenols and minerals, which determine the nutritional and health benefits of oat [22]. Particularly, oat seed lipid content is the highest (up to 18%) within the cereal species, and the accumulated lipid is predominantly (up to 90%) confined to the starchy endosperm, which has attracted the attention of researchers for the development of high-oil oat varieties as a sustainable oil production [23,24]. Also, several gene expression studies of developing oat seeds have been conducted, which have led to the identification of a number of promising targets for lipid improvement in oat [25,26]. However, to date, genes contributing to lipid accumulation in oat endosperm have not been functionally characterized.
In this study, we systematically investigated the PDAT family in oat, which has not been previously described. The oat genome contained three homologous copies of AsPDAT genes caused by polyploidization, which tend to possess functional redundancy because of sequence similarity relationships and similar expression patterns. Overexpression of AsPDAT-5C in tobacco significantly increased lipid levels in both seed and leaf organs. We further characterized the role of AsPDAT-5C in regulating abiotic stress resistance, including cold and phosphorus deficiency. Collectively, the identification of AsPDAT-5C provides a promising genetic resource for enhancing lipid yield. Our work further reveals the role of AsPDAT-5C in plant adaptation to abiotic stresses by demonstrating its impact on lipid metabolism.
2. Results
2.1. Genome-Wide Identification and Sequence Analysis of PDATs in Oat
To identify PDAT members in oat, the protein sequences Arabidopsis PDAT (AtPDAT1/At5g13640 and AtPDAT2/At3g44830) were used as queries for BLAST search against the oat genome. It is well known that cultivated oat is a hexaploid species with most genes present as triplicate homoeologs, which are derived from hybridization between its diploid and tetraploid progenitor species. Accordingly, we identified three putative PDAT members forming a collinear homeologous set located on chromosomes 5A, 5C, and 5D, which were designated AsPDAT-5A, AsPDAT-5C, and AsPDAT-5D based on their chromosomal origins. The deduced amino acid sequences of the three AsPDATs shared high sequence similarity (96–98%). EMBOSS Needle pairwise alignment showed that AsPDAT-5A, AsPDAT-5C, and AsPDAT-5D share 73.2%, 77.2%, and 76.9% amino acid identity with AtPDAT1, respectively, but less than 60% identity with AtPDAT2. We also computed basic physicochemical properties for each protein, including amino acid number, molecular weight (MW), theoretical isoelectric point (pI), and aliphatic index (Table 1). Subcellular localization predictions indicated that all three AsPDATs are likely endoplasmic reticulum (ER)-localized proteins.
All three AsPDAT homoeologs were predicted to contain an N-terminal transmembrane domain (TMD) composed of 21 amino-acid residues and a conserved lecithin:cholesterol acyltransferase (LCAT) domain (Figure 1A). The LCAT domain is characteristic of lipid-metabolizing enzymes such as mammalian LCAT and plant PDAT. Multiple sequence alignment of oat AsPDATs, Arabidopsis AtPDATs, and human HsLCAT showed that PDATs shared the functionally important features of LCAT enzyme, including Trp-61 within the ‘lid’ region, a salt bridge and the catalytic Ser-Asp-His triad (Figure 1B). It is worth mentioning that the residues surrounding these conserved sites diverge considerably between plant PDATs and human LCAT (Figure 1B), which may contribute to the formation of novel structures and functions of plant PDAT enzymes during evolution. Based on the well-assembled reference genome, we found that all three AsPDAT genes had similar exon-intron organization, each consisting of six exons and five introns (Figure 1C).
2.2. Phylogenetic Analysis of PDATs in Oat
A phylogenetic tree constructed from representative eudicots, monocots, mosses, ferns, lycophytes, and green algae showed that PDATs could be separated into six distinct clades, each of which was further subdivided into two or more subclades (Figure 2). Algal PDATs were phylogenetically separated from land plant PDATs and grouped at the basal position of the tree. Clade III included PDAT orthologs from mosses, ferns and lycophytes, whereas Clade V was exclusive to monocots. Eudicot PDATs did not cluster together and were distributed across three different clades: clades II, IV, and VI, with each clade consisting of members from a mixture of species. Interestingly, clade VI within eudicots was phylogenetically closer to the monocot clade than to other eudicot PDATs. It was further found that PDAT copy numbers varied widely across different plant species. In summary, green algae, mosses, ferns, and lycophytes generally contained a single PDAT in each species. Within monocots, most (sub)genomes also possessed one single PDAT gene, with the only exception being maize, which is a diploid species but contains two PDAT copies. In contrast, PDAT genes have expanded notably in eudicots. As a hexaploid plant, the oat genome contained three AsPDAT homeologs, corresponding to the sum found in its ancestors, namely, one in diploid Avena longiglumis and two in tetraploid Avena insularis. The phylogenetic reconstruction indicated that oat AsPDAT genes originated from its ancestral lineages (Figure 2).
2.3. Expression Analysis of AsPDAT Genes in Oat
As the first step to explore the potential functions of AsPDAT genes in oat, we examined their expression across different organs and under diverse environmental conditions. We firstly utilized the publicly available transcriptome data from oat to detect the expression patterns of AsPDAT genes in root, stem, leaf, spike, and seeds at three developmental stages. Fragments per kilobase of transcript per million mapped reads (FPKM) values showed that all three AsPDAT homeologs shared similar spatial expression patterns, with detectable transcripts in all organs and comparatively higher expression in leaves and during late seed development (Figure 3A). Among them, AsPDAT-5C was more highly expressed than the other two AsPDAT genes (Figure 3A). Additionally, we performed quantitative real-time PCR (qRT-PCR) to validate these patterns. Consistent with the transcriptome data, all three AsPDAT genes were preferentially expressed in leaves and developing seeds (Figure 3B). Interestingly, each gene displayed coordinated transcriptional changes under several abiotic stresses (Figure 3C–G). Under cold stress, the expression levels of AsPDAT genes were basically unaffected during the first 3 h, then sharply increased with the peak at 9 h, and then declined (Figure 3C). We also confirmed that AsPDAT genes were transcriptionally induced at 5 d after phosphorus (P) deficiency, with maximal levels at 15 days followed by a decrease (Figure 3F). In addition, AsPDAT genes were significantly altered under drought (Figure 3D) and saline (Figure 3E) conditions, but not under nitrogen (N) deficiency (Figure 3G). Collectively, these findings indicate that the three AsPDAT genes may function similarly in response to multiple stresses. Notably, AsPDAT-5C was the most strongly induced under both cold and P-deficient conditions (Figure 3C,F), underscoring its potential role in adaptation to these specific stresses.
2.4. AsPDAT-5C Is Localized to Endoplasmic Reticulum and Exhibits Acyltransferase Activity
To determine the subcellular localization of AsPDAT-5C, the 35S::AsPDAT-5C-GFP fusion construct was transiently expressed in Nicotiana benthamiana leaf epidermal cells via Agrobacterium-mediated transformation. Confocal microscopy revealed that the green fluorescence signal from the AsPDAT-5C-GFP fusion protein extensively overlapped with the signal from the co-expressed endoplasmic reticulum (ER) marker mCherry-HDEL (Figure 4A), confirming the ER localization of AsPDAT-5C.
To investigate whether AsPDAT-5C possesses phospholipid:diacylglycerol acyltransferase (PDAT) activity, its full-length coding sequence was cloned into the yeast expression vector pYES2.0 and expressed in the TAG-deficient Saccharomyces cerevisiae quadruple mutant H1246 under the control of a galactose-inducible promoter. Lipid accumulation was visualized using the neutral lipid-specific fluorescent dye BODIPY 505/515. Following galactose induction, H1246 cells expressing AsPDAT-5C displayed intense BODIPY fluorescence, indicative of neutral lipid synthesis (Figure 4B). In contrast, control cells transformed with the empty vector showed negligible fluorescence (Figure 4B). Consistent with this phenotypic complementation, quantitative analysis confirmed a significant increase in TAG content in the AsPDAT-5C-expressing yeast cells (Figure 4C). Collectively, these results demonstrate that AsPDAT-5C is an ER-localized protein that functions as a PDAT, capable of restoring TAG biosynthesis in the lipid-deficient yeast mutant H1246.
2.5. Overexpressing AsPDAT-5C Increased Total Lipid Content in Tobacco (Nicotiana tabacum)
To examine the role of AsPDAT-5C gene in lipid biosynthesis, we stably introduced a constitutive 35S::AsPDAT-5C construct into model plant tobacco (N. tabacum) via Agrobacterium-mediated leaf disc assay. From the resulting transgenic lines, two independent lines (OE3 and OE14) showing the highest AsPDAT-5C transcript levels were selected for further analysis (Supplementary Figure S1). Under pot-grown conditions, the overexpression lines showed no obvious phenotypic differences from wild-type (WT) plants (Figure 5A). In WT tobacco controls, the total seed lipid content was 32.8% of dry weight (DW), while overexpression of AsPDAT-5C led to a significant increase in total seed lipid content to 41.1% in OE3 and 40.08% in OE14 (Figure 5B). A similar increase was also observed in transgenic tobacco leaves, where lipid content rose from 6.46% DW in WT to 13.2% in OE3 and 12.5% in OE14 (Figure 5E). These results demonstrate that AsPDAT-5C enhances lipid accumulation in both seed and vegetative organs. In contrast, protein and soluble sugar levels in leaves and seeds of the overexpression lines remained largely unchanged relative to WT (Figure 5C,D,F,G), indicating that the increased lipid content did not come at the expense of these major metabolites.
2.6. Overexpression of AsPDAT-5C Enhanced Cold Stress Tolerance
Because the expression of AsPDAT-5C was stress-inducible, we are interested in exploring whether AsPDAT-5C was involved in regulating cold tolerance. Following 3 d of cold exposure, WT seedlings displayed more severe wilting, curling and necrosis compared to the transgenic tobacco lines (Figure 6A). In addition, the MDA content and H_2_O_2_ accumulation were significantly lower in AsPDAT-5C overexpressed lines compared to those of the WT (Figure 6B,C), while the enhanced activities of antioxidant enzymes such as superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT) were observed in the seedlings of overexpression plants (Figure 6D–F). Transgenic lines also displayed enhanced cold-tolerance phenotype with reduced ion leakage relative to WT after cold treatment (Figure 6G). To provide a more comprehensive analysis, we conducted measurements on the leaf fatty acid composition. Under normal conditions, overexpression of AsPDAT-5C showed a reduction in proportions of C16:0 and C18:0 and an increase in C18:1 (Figure 6H). After cold treatment, transgenic leaves contained significantly less C16:0 and C18:0 and more of the unsaturated fatty acids C18:1 and C18:3 than WT leaves (Figure 6I). These shifts toward a more unsaturated fatty acid profile are consistent with a role for AsPDAT-5C in modulating membrane lipid composition to enhance cold tolerance.
2.7. Overexpression of AsPDAT-5C Increased Tolerance to P Deficiency
To further investigate the stress-related function of AsPDAT-5C, the WT and overexpression lines were subjected to P-deficient conditions. After 15 days of P deficiency, transgenic plants exhibited better growth performance, while WT plants showed pronounced growth retardation and leaf shrinkage (Figure 7A). To explore the molecular mechanism underlying this enhanced tolerance, we selected four well-established low P-responsive marker genes, including NtPHT1;1 (phosphate transporter, GenBank accession number: AF156696), NtPHO1 (phosphate transporter, GenBank accession number: 107814602), NtPHR1 (phosphate starvation response regulator, GenBank accession number: 107783857), and NtMGD2 (monogalactosyldiacylglycerol synthase 2, GenBank accession number: AB047476), and analyzed their expression profiles in leaves of WT and overexpression plants under both control and P-deficient conditions. Under normal P supply, transcript levels of these genes were comparable between WT and overexpression lines. Upon P stress, however, all four genes were significantly induced in both WT and transgenic plants, with their expression consistently and markedly higher in the overexpression lines compared to WT (Figure 7B–E), suggesting that the overexpression of the AsPDAT-5C gene, which improved P deficiency tolerance, may be associated with the expression of these stress-responsive marker genes. Consistent with the role of PDAT in lipid metabolism, lipid profiling revealed that under P deficiency, transgenic leaves accumulated less phospholipid but more neutral lipid and galactolipid compared to WT leaves (Figure 7F–H). These results suggest that AsPDAT-5C enhances P deficiency tolerance by remodeling membrane lipid composition, specifically by reducing phospholipid demand and promoting the synthesis of non-phosphorus-containing neutral and galactolipids.
3. Discussion
As a widely used feedstock in both food and non-food applications in modern society, enhancing the accumulation of storage lipid (predominantly triacylglycerol, TAG) in plants is of great interest in molecular breeding of oil crops, which has encouraged the identification of gene targets involved in TAG biosynthesis in plants [27]. PDAT is a strong candidate enzyme responsible for the control of TAG biosynthesis in an acyl-CoA-independent manner. In contrast to DGAT, which catalyzes the final step of the acyl-CoA-dependent TAG biosynthesis and has been extensively studied in oilseeds, the biological functions of PDAT remain less clear [3]. Within cereal crops, oat is unique for its unusually high lipid-accumulating capacity in seed, marking its potential as an alternative oil crop worldwide [23]. Motivated by the likely role of PDAT in TAG accumulation, we conducted a comprehensive analysis of the PDAT gene family in oat.
Genome-wide identification revealed a homeologous triplet of PDAT genes in oat, and this balanced configuration may have contributed to the stability of TAG metabolism during oat evolution. Taking advantage of publicly available sequenced genomes, we further demonstrated that PDATs are broadly existent in the Viridiplantae (Figure 2), consistent with their indispensable roles in primary metabolism. The topology of the phylogenetic tree allowed us to classify the PDAT proteins into six clades, with most clades having high statistical support (bootstrap > 80%; Figure 2). Within these clades, most internal branches also showed strong support, indicating stable sub-structuring. Interestingly, the large differences in the number of PDAT genes in different plant lineages were observed. The eudicot-specific PDAT gene expansion indicated no direct link between the number of these genes and their genome size. Phylogenetic analysis revealed that algal PDATs formed a distinct clade from land plants (Figure 2), suggesting that algal PDATs may possess divergent enzymatic functions. This hypothesis was supported by the structural differences between algal and land plant PDATs [10]. For instance, the Chlamydomonas PDAT has been reported to exhibit previously unrecognized transacylation and lipase activities using galactolipids and neutral lipids as substrates [10]. Previous studies have also observed the existence of multiple PDAT gene copies in different eudicots [4,28], but not all encoded PDATs are functional TAG-synthesizing enzymes, such as AtPDAT2, LuPDAT5, LuPDAT6, and RcPDAT3 [6,29,30]. Interestingly, all these PDATs fall within the clade II in our tree (Figure 2). In addition to the possible nonfunctionalization of clade II PDATs, we speculate that these PDATs might have algal-like lipase activities due to their phylogenetic proximity to algal sequences (Figure 2), but more direct enzyme activity assays are needed to investigate this hypothesis. The PDAT members from the three Avena species clustered together, and phylogenetic analysis of oat, wheat, barley, all belonging to the Pooideae, share a closer evolutionary relationship (Figure 2). Additionally, our phylogeny are consistent with previous inference that the A and D subgenomes of hexaploid oat are more closely related to each other than to the C subgenome [31].
Although the involvement of PDAT in enhancing lipid production has been reported in several oil crops, the activities of AsPDATs in oat remained unclear. In this study, the expression levels of AsPDAT genes increased gradually and peaked during the late stage of seed development (Figure 3A,B), suggesting that AsPDATs may promote seed lipid accumulation in a developmentally dependent way, which was consistent with the transcript profiles in Arabidopsis [12], flax [29] and Sapium sebiferum [14]. Our investigation further suggested that the three AsPDAT homeologous genes may be functionally redundant due to their similar expression patterns (Figure 3) and high sequence similarities (96–98%). The minimal variation in physicochemical properties further indicated functional conservation and redundancy within AsPDAT proteins. Notably, the AsPDAT-5C gene constantly exhibited the highest expression patterns (Figure 3), suggesting that it may be the major PDAT isoform in oat. Heterologous expression in the mutant yeast H1246 and subcellular localization assay suggested that AsPDAT-5C contributes to ER-localized PDAT activity for TAG accumulation in oat (Figure 4). Previous research has indicated that neither mutation nor overexpression of AtPDAT1 had overt impact on Arabidopsis seed lipid phenotype [6,7], but silencing of AtPDAT1 led to a 63% reduction in total lipid content in the Arabidopsis dgat1 mutant background [12], demonstrating that AtPDAT1 is a key determinant of seed lipid biosynthesis in the absence of DGAT1 activity. However, our data showed that ectopic expression of AsPDAT-5C in tobacco could give a 30% increase in seed lipid content (Figure 5B). This is consistent with the genetic manipulations of PDAT genes that are expected to elevate seed lipid biosynthesis, including overexpression of CsaPDAT1 in Camelina sativa [20] and overexpression of SsPDAT1 in Brassica napus [14]. In this study, AsPDATs genes exhibited the highest expression level in oat leaves not in seeds, reflecting their fundamental roles in association with lipid accumulation in vegetative organs (Figure 3A,B). Indeed, overexpression of AsPDAT-5C also significantly increased lipid levels in tobacco leaves (Figure 5E). Supporting this result, expression of AtPDAT1 in Arabidopsis caused a large increase in its leaf lipid content [13]. Although lipid concentrations are usually low in vegetative organs, bioengineering efforts to bolster lipid production in leafy biomass crops are a promising sustainable source of renewable feedstocks [32]. From a biotechnological point of view, this clarification of the role of AsPDAT-5C may provide a desirable target for future genetic engineering to enhance lipid accumulation in both seed and leaf organs in biomass crops.
Although functional redundancy between DGAT and PDAT in lipid synthesis was proposed over two decades ago, the presence of varied expression indicates the crucial role of PDAT in the plant vegetative stages. Previously, metabolic and biochemical profiles have revealed enhanced lipid synthesis in plant vegetative organs under a variety of environmental stresses, confirming the role of lipid metabolism in stress tolerance improvement [33]. However, it is unknown whether PDAT-mediated lipid mechanism is a controlled process required to tolerate abiotic stresses. Through gene expression analysis, we determined that AsPDAT genes were significantly induced under multiple unfavorable conditions (Figure 3C–F). Enhanced cold tolerance has been previously proposed in AtPDAT1 overexpression in Arabidopsis plants [34,35]. Similarly, our transgenic tobacco plants overexpressing AsPDAT-5C also performed better under cold conditions (Figure 6). When suffering from changes in temperatures, plants have evolved a series of sophisticated mechanisms to cope with cold stress, one adaptive strategy by which plants respond to low temperature change is to increase fatty acid unsaturation levels to maintain membrane integrity and fluidity, which is beneficial for improving photosynthetic efficiency [36]. Indeed, our studies found that unsaturated fatty acids were increased in the overexpression plants compared to that of wild type at low temperatures (Figure 6I). This observation is in accordance with both the function of PDAT as an enzyme having a substrate preference for unsaturated fatty acids and the function of the ER as the cellular site for membrane lipid biosynthesis [37]. In addition, cold stress could cause oxidative damage due to excess ROS accumulation. Our study revealed that overexpression plants exhibited enhanced peroxidase activities (POD, SOD and CAT) and lower levels of MDA and H_2_O_2_ (Figure 6). Thus, we proposed that AsPDAT-5C-mediated cold tolerance could also be partially explained by a stronger ROS scavenging capacity.
Previous observations strongly suggested that in addition to its common role in lipid synthesis, PDAT may contribute to adaptation to abiotic stresses in plants [3]. However, the role of PDAT in nutrient deficiency is poorly understood. In accordance with the altered expression pattern (Figure 3F), our data revealed a novel positive effect of AsPDAT-5C on low-P stress (Figure 7). Compared to the lipid composition in WT leaves, overexpression of AsPDAT-5C facilitated the degradation of phospholipids during P deficiency (Figure 7G), which likely resulted from its phospholipase activity using phospholipids as substrates. Phospholipids constitute about 1/3 of organic P in plant organs, and degradation of phospholipids to release P for other critical cellular processes is an effective strategy to cope with P-limited environments in plants [37,38]. As expected, the elevated accumulation of non-P neutral lipids was due to the acyltransferase activity of AsPDAT-5C. As a major class of neutral lipids, the importance of TAG metabolism in P stress response has begun to be appreciated [39,40], but the specific enzymes and underlying mechanisms have not been investigated. The present study provides evidence that AsPDAT-5C mediates TAG biosynthesis, but it also displays actions on phospholipid hydrolysis; thus, we cannot rule out the possibility that other genes for TAG production, like DGAT, help plants adapt to low-P environments. Glycolipids, another important class of non-P-containing lipids, are mainly utilized to functionally compensate for the lack of phospholipids and thus hold prime importance under P deficiency [41]. As reflected by the upregulation of glycolipids under low-P conditions (Figure 7H), our results suggest that oat AsPDAT-5C may cooperate with other genes involved in glycolipid biosynthesis to maintain the balance of lipid metabolism and enhance P utilization efficiency in plants under low-P conditions. This assumption is supported by the observation that tobacco galactolipids (MGDGs) biosynthetic gene, NtMGD2, a P-deficiency-responsive gene, showed induced expression in AsPDAT-5C OE plants. Therefore, the enhanced stress tolerance in AsPDAT-5C overexpression lines may not be solely attributable to this single gene but could result from its ability to orchestrate or synergize with a broader transcriptional and metabolic reprogramming involving these other critical genes.
4. Materials and Methods
4.1. Sequence Acquisition and Bioinformatics Analysis of PDAT Family Genes
Genomic files of oat (A. sativa), A. longiglumis, and A. insularis were downloaded from the OatOmics (http://www.oatomics.com, accessed on 10 August 2024). The genomic sequences of other species were downloaded from the public database Phytozome (https://phytozome-next.jgi.doe.gov/, accessed on 10 August 2024). The amino acid sequences of Arabidopsis AtPDAT proteins retrieved from TAIR (https://www.arabidopsis.org/, accessed on 10 August 2024) were used as queries to identify candidate PDAT orthologs in all listed plant species (Supplementary Table S1) by the BLAST approach. The presence of a transmembrane region and conserved LCAT signature in all candidate sequences was determined using InterPro (https://www.ebi.ac.uk/interpro/, accessed on 10 August 2024) and SMART database (http://smart.embl-heidelberg.de/, accessed on 10 August 2024). Multiple sequence alignments were performed using the MAFFT versions 7.526. The sequence identity was determined using EMBOSS Needle (https://www.ebi.ac.uk/jdispatcher/psa/emboss_needle, accessed on 10 August 2024). The phylogenetic tree was constructed using the Maximum Likelihood (ML) method with the Jones-Taylor-Thornton model, nearest-neighbour interchange, uniform rates and 1000 bootstrap replicates in MEGA11.0 software [42]. Visualization of the tree was performed using iTOL (https://itol.embl.de/, accessed on 10 August 2024). The physical and chemical properties of AsPDAT proteins in oat, including molecular weight, instability index, hydrophobicity, and isoelectric point, were predicted by using the ExPASy server (https://www.expasy.org/, accessed on 10 August 2024). Gene structure annotation files of AsPDAT genes retrieved from OatOmics were used to visualize the diagram of exon/intron distribution in Gene Structure Display Server (http://gsds.cbi.pku.edu.cn/, accessed on 10 August 2024). Subcellular localization was predicted using the Cell-PLoc 2.0 package (http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/, accessed on 10 August 2024).
4.2. Plant Materials and Growth Conditions
All experiments were carried out in the spring of 2023 at the experimental station of Shanxi Agricultural University, Shanxi, China. The seeds of oat cultivar CEav5651 were obtained from the agricultural genetic resources center, Shanxi Agricultural University. Oat seeds were planted in a 3:1 (w/w) mixture of soil and sand and placed in a greenhouse with 16 h light/8 h dark and 80% relative humidity at 22 °C. The photoperiod of 16 h per day was maintained under natural light supplemented with white fluorescent tube at an approximate photon flux intensity of 220 μmol·m^−2^·s^−1^. To perform organ-specific expression profiling of AsPDAT genes, different oat organs were collected from roots, stems, leaves, and spikes at the heading stage. The seed samples were collected at three development stages (10, 20, and 30 days after flowering, DAF). These samples were immediately snap-frozen in liquid nitrogen and stored at −80 °C for future use. Nicotiana benthamiana and Nicotiana tabacum employed in this study were grown in a manual climatic box (RDN-560C-4, Ningbo, China) under a photoperiod of 16 h light and 8 h darkness, photon flux intensity of 250 µmol∙m^−2^∙s^−1^, constant temperature of 25 °C, and relative humidity of 70%. A hydroponic cultivation experiment was performed to conduct stress treatment in oat. Oat seeds were firstly germinated on filter paper soaked with distilled water and incubated in the dark for 96 h at 24 °C. Later, the germinated seeds were transplanted into hydroponic buckets containing Half-strength Hoagland’s modified nutrient solution (pH 6.0) and placed in a greenhouse with 16 h light/8 h dark and 80% relative humidity at 22 °C. The photoperiod of 16 h per day was maintained under natural light supplemented with white fluorescent tube at an approximate photon flux intensity of 220 μmol·m^−2^·s^−1^. Half-strength Hoagland’s modified nutrient solution contains 2.5 mM KNO_3_, 0.5 mM NH_4_NO_3_, 0.5 mM KH_2_PO_4_, 1 mM MgSO_4_, 0.15 mM FeEDTA, 5 μM KI, 0.1 mM H_3_BO_3_, 0.15 mM MnSO_4_, 0.05 mM ZnSO_4_, 0.1 μM CoCl_2_, 20 mM Na_2_MoO_4_, 0.01 μM CuSO_4_, and 3 mM Ca(NO_3_)2. When oat seedlings had grown for four weeks, diverse stress treatments were initiated, including salt stress (200 mM NaCl), drought (18% PEG6000, w/v, −0.75 MPa), and cold (4 °C). Additionally, for phosphorus (P) deficiency treatment, oat seedlings were transferred to a P-deficient solution, which was identical to the half-strength Hoagland’s modified solution except that 0.5 mM KH_2_PO_4_ was replaced by 0.5 mM KCl. For nitrogen (N) deprivation treatment, oat seedlings were transferred to N-deficient solution where KNO_3_ (2.5 mM), NH_4_NO_3_ (0.5 mM), and Ca(NO_3_)2 (3 mM) in the half-strength Hoagland’s modified nutrient medium were replaced with 2.5 mM KCl, 0.5 mM NH_4_Cl, and 3 mM CaCl_2_, respectively. For wild-type and transgenic tobacco (N. benthamiana) lines, plants were initially grown in vermiculite and irrigated weekly with half-strength Hoagland’s modified nutrient solution. The 40-day-old tobacco seedlings were prepared and treated at 4 °C for cold stress, with normal growth conditions as the control. Leaf samples were collected at 72 h for further use. The treatment conditions of P deprivation and N deprivation were irrigated with P-deficient half-strength Hoagland’s modified nutrient solution, with normal growth conditions as the control. Leaf samples were collected at 15 day for further use. Unless otherwise specified, all general chemicals and reagents were obtained from Beijing Solarbio Science & Technology (Solarbio, Beijing, China).
4.3. RNA Isolation and Gene Expression Analysis
Total RNA of the above-mentioned samples was isolated using the RNA Extraction Kit (TIANGEN, Beijing, China). The Prime Script RT Reagent Kit (Takara Bio, Dalian, China) was used to synthesize cDNA and SYBR Premix Ex Taq II (Takara Bio, Dalian, China) was used for quantitative real-time PCR (qRT-PCR) analysis on a CFX96 Real-Time PCR Detection System (BIO-RAD, Hercules, CA, USA). The AsActin gene (GenBank accession number: KP257585.1) was chosen as an internal control and the 2^−ΔΔCT^ method was used to quantify the relative expression of AsPDAT genes in oat. NtTubulin (GenBank accession number: N181029A17) was used as an internal control to investigate gene expression in transgenic tobacco plants overexpressing AsPDAT-5C and WT plants. Each sample was assayed with three biological replicates. Primers used were shown in Supplementary Table S2. The transcriptome data used in this study were obtained from Data Center’s Genome Sequence Archive (http://bigd.big.ac.cn/, accessed on 25 November 2025) under accession numbers SAMN19582573 and SAMN19582574. Raw transcriptome data were first subjected to quality control using FastQC v0.12.1 (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/, accessed on 25 November 2025). Subsequently, the clean reads were aligned to the reference genome with HISAT2 v2.2.1 (https://daehwankimlab.github.io/hisat2/, accessed on 25 November 2025). Read counts for each gene were generated with HTSeq-count, and the final gene expression levels were calculated as fragments per kilobase of transcript per million mapped reads (FPKM) values using R (https://www.r-project.org/, accessed on 25 November 2025). This established workflow ensures the accuracy and reproducibility of the results.
4.4. Subcellular Localization and AsPDAT-5C Expression in Yeast Mutant H1246
To determine the subcellular localization of AsPDAT-5C, its coding sequence (CDS) was amplified and cloned into the plant expression vector pCAMBIA1300-35S-GFP, generating a C-terminal fusion construct designated pCAMBIA1300-35S::AsPDAT-5C-GFP. For precise organelle localization, the endoplasmic reticulum (ER) marker construct pCAMBIA1300-35S::ER-mCherry-HDEL (Coolaber Biological Technology, Beijing, China) was used. Each construct was then individually transformed into Agrobacterium tumefaciens strain GV3101 via the freeze-thaw method to generate two distinct bacterial strains. For infiltration, Agrobacterium suspensions harboring the respective constructs were mixed in equal volumes (based on OD600) and co-infiltrated into the leaves from four-week-old Nicotiana benthamiana plants. After 48 h of incubation, epidermal cells were imaged using a confocal laser-scanning microscope (Leica TCS SP8, Leica Microsystems, Wetzlar, Germany). The co-localization of the AsPDAT-5C-GFP fusion signal with the ER-mCherry marker was analyzed to confirm ER localization.
For functional complementation assays, the CDS of AsPDAT-5C was subcloned into the galactose-inducible yeast expression vector pYES2.0, yielding the construct pYES2.0-AsPDAT-5C. This construct was transformed into the TAG-deficient Saccharomyces cerevisiae H1246 quadruple mutant using the polyethylene glycol/lithium acetate method [43]. Tragansformants were selected on synthetic complete (SC) medium lacking uracil (SC-Ura). For induction, single colonies were inoculated into liquid SC-Ura medium containing 2% (w/v) galactose and cultured at 30 °C with shaking at 180 rpm for 72 h in the dark using a shaker incubator (YIHENG Technology, Shanghai, China). To visualize neutral lipid accumulation, induced yeast cells were stained with the lipid droplet-specific fluorescent dye BODIPY 505/515 and observed under the confocal microscope (Leica TCS SP8, Leica Microsystems, Wetzlar, Germany). Total TAG was extracted from the yeast cells and quantified using a thin-layer chromatography (TLC)-based densitometry method, as previously described [43]. All experiments were performed with three biological replicates. Primers used for vector construction are listed in Supplementary Table S2.
4.5. Vector Construction and Transgenic Plant Generation
The amplified AsPDAT-5C coding sequence was cloned into the pCAMBIA1303 vector driven by the 35S promoter for constitutive overexpression. The final construct (pCAMBIA1303-AsPDAT-5C) was introduced into tobacco (N. tobaccum) WT for stable genetic transformation using the Agrobacterium (GV3101)-mediated method as described previously [44]. The AsPDAT-5C overexpressed tobacco plants were obtained and characterized by reverse transcription PCR (RT-PCR). Two independent T2 transgenic lines showing higher expression levels of AsPDAT-5C were used in the experiments. Wild type (WT) tobacco plants were used as control. The primer sequences are listed in Supplementary Table S2.
4.6. Total Lipid, Soluble Sugar and Protein Measurement
Total lipid content was determined following a previously established protocol [44]. Briefly, freeze-dried mature tobacco leaf and seed samples harvested from WT and AsPDAT-5C-overexpressing lines were ground into powder. Then, 0.05 g powder was weighed and dissolved in a 7.5 mL of methanol/chloroform (2:1, v/v) mixture. After overnight incubation at 37 °C, the mixture was centrifuged at 5000 rpm for 8 min. The supernatant was added with 5 mL of chloroform and 9 mL of a 1% NaCl (w/v). After centrifugation at 8000 rpm for 10 min, the extract was separated into two distinct phases: an upper aqueous phase and a lower organic phase containing the total lipid. The upper aqueous phase was discarded. The lower lipid-containing organic phase was transferred into a pre-weighed glass vial (weight recorded as W1). The solvent was completely evaporated under a gentle stream of nitrogen gas. The vial with the extracted lipid was then reweighed (W2). Total lipid content was determined using the following formula: total lipid content (%) = [(W2 − W1)/0.05] × 100. Following the instructions of the reagent kit, soluble sugar and protein contents from freeze-dried mature tobacco leaf and seed samples were determined using the Plant Soluble Sugar Content Assay Kit (Solarbio, Beijing, China) and Bradford Protein Assay Kit (GenStar, Beijing, China), respectively. All subsequent content calculations are expressed on a dry weight basis.
4.7. Fatty Acid Profile and Lipid Composition Analysis
Preparation of fatty acid methyl esters (FAMEs) was carried out using a previously established protocol [43]. Heptadecanoic acid (C17:0) was used as an internal standard during lipid extraction. The fatty acid profiles of each sample were analyzed on an Agilent 7890B gas chromatograph (GC) equipped with a 5975B mass selective detector and a G3903-63011 capillary column (Agilent Technologies, Santa Clara, CA, USA). Relative quantification of individual fatty acids in samples was achieved by normalizing the retention time and peak area of the internal standard (C17:0). Lipid component separation was achieved using the Cleanert Silica solid phase extraction (SPE) Cartridge (Agela, Tianjin, China), similar to the method described previously [39]. Briefly, the neutral lipids, glycolipids, and phospholipids were separated by stepwise gradient elution from chloroform, acetone, and methanol, respectively. The different lipid components were quantified by weighing after evaporation under nitrogen.
4.8. Analysis of Oxidative Stress Parameters
The malondialdehyde (MDA) content was quantified using the thiobarbituric acid (TBA) method as described by Heath and Packer [45]. Briefly, 0.1 g fresh leaf samples was homogenized in an extraction solution containing 10% (w/v) trichloroacetic acid (TCA). The homogenate was then mixed with 5% (w/v) TBA solution and incubated in a boiling water bath for 30 min. After cooling, the mixture was centrifuged at 12,000× g for 30 min. The absorbance of the supernatant was measured at 450 nm, 532 nm, and 600 nm for the calculation of MDA content. The hydrogen peroxide (H_2_O_2_) content was determined spectrophotometrically following the protocol of Velikova et al. [46]. Briefly, 0.1 g fresh leaf samples were homogenized in ice-cold 0.1% (w/v) trichloroacetic acid and centrifuged at 12,000× g for 20 min at 4 °C. The supernatant was incubated with a reaction buffer consisting of 10 mM potassium phosphate buffer (pH 7.0) and 1 mM potassium iodide (KI). The absorbance of the reaction mixture at 390 nm was measured to determine H_2_O_2_ content. The enzyme activities of superoxide dismutase (SOD, EC 1.15.1.1, Cat: A001-3-2), peroxidase (POD, EC 1.11.1.7, Cat: A084-3-1), and catalase (CAT, EC 1.11.1.6, Cat: A007-1-1) in tobacco seedling leaves were measured using the corresponding assay kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer’s instructions. For the ion leakage assay, leaves of tobacco seedlings were collected in 15 mL tubes containing 10 mL of ddH_2_O. The initial conductivity of the solution was measured and designated as S0. The tubes were then shaken at room temperature for 30 min, after which the conductivity was measured and recorded as S1. Subsequently, the samples were boiled for 30 min, cooled to room temperature with shaking, and the final conductivity (S2) was measured. Ion leakage was calculated according to the formula Ion leakage (%) = [(S1 − S0)/(S2 − S0)] × 100.
5. Conclusions
This study presented the first genome-wide identification of the PDAT gene family in hexaploid cultivated oat, a cereal renowned for its high seed lipid content. We systematically identified three homologous AsPDAT genes derived from polyploidization, which were distributed across subgenomes and exhibited high conservation in sequence and structure. Phylogenetic and expression profiling revealed that these homologs likely retain similar functions, with AsPDAT-5C showing consistently higher transcript levels under both developmental and abiotic stress conditions. Functional characterization confirmed that AsPDAT-5C encodes an endoplasmic reticulum-localized enzyme with authentic PDAT activity, capable of complementing TAG biosynthesis in yeast. Crucially, stable overexpression of AsPDAT-5C in tobacco significantly enhanced lipid accumulation in both leaves and seeds and substantially improved plant tolerance to cold and P deficiency stresses. Mechanistic investigations indicated that this multi-stress resilience is mediated through AsPDAT-5C-driven lipid remodeling, including increased fatty acid unsaturation for membrane integrity under cold and a shift from phospholipids to non-phosphorus lipids for phosphate homeostasis under P limitation. Our results provide foundational insights into the AsPDAT gene family and establish AsPDAT-5C as a key regulator integrating lipid metabolism with environmental adaptation. This work not only offers a valuable genetic resource for understanding lipid dynamics in polyploid cereals but also highlights AsPDAT-5C as a promising biotechnological target for the dual improvement of lipid yield and abiotic stress resilience in crops.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yang Y. Benning C. Functions of triacylglycerols during plant development and stress Curr. Opin. Biotechnol.20184919119810.1016/j.copbio.2017.09.00328987914 · doi ↗ · pubmed ↗
- 2Xu Y. Caldo K.M.P. Pal-Nath D. Ozga J. Lemieux M.J. Weselake R.J. Chen G. Properties and Biotechnological Applications of Acyl-Co A:diacylglycerol Acyltransferase and Phospholipid:diacylglycerol Acyltransferase from Terrestrial Plants and Microalgae Lipids 20185366368810.1002/lipd.1208130252128 · doi ↗ · pubmed ↗
- 3Sah S.K. Fan J. Blanford J. Shanklin J. Xu C. Physiological Functions of Phospholipid:Diacylglycerol Acyltransferases Plant Cell Physiol.20246586387110.1093/pcp/pcad 10637702708 · doi ↗ · pubmed ↗
- 4Pan X. Peng F.Y. Weselake R.J. Genome-wide analysis of phospholipid: Diacylglycerol acyltransferase (PDAT) genes in plants reveals the eudicot-wide PDAT gene expansion and altered selective pressures acting on the core eudicot PDAT paralogs Plant Physiol.201516788790410.1104/pp.114.25365825585619 PMC 4348769 · doi ↗ · pubmed ↗
- 5Dahlqvist A. Stahl U. Lenman M. Banas A. Lee M. Sandager L. Ronne H. Stymne S. Phospholipid:diacylglycerol acyltransferase: An enzyme that catalyzes the acyl-Co A-independent formation of triacylglycerol in yeast and plants Proc. Natl. Acad. Sci. USA 2000976487649210.1073/pnas.12006729710829075 PMC 18631 · doi ↗ · pubmed ↗
- 6Stahl U. Carlsson A.S. Lenman M. Dahlqvist A. Huang B. Banas W. Banas A. Stymne S. Cloning and functional characterization of a phospholipid:diacylglycerol acyltransferase from Arabidopsis Plant Physiol.20041351324133510.1104/pp.104.04435415247387 PMC 519051 · doi ↗ · pubmed ↗
- 7Mhaske V. Beldjilali K. Ohlrogge J. Pollard M. Isolation and characterization of an Arabidopsis thaliana knockout line for phospholipid: Diacylglycerol transacylase gene (At 5g 13640)Plant Physiol. Biochem.20054341341710.1016/j.plaphy.2005.01.01315907694 · doi ↗ · pubmed ↗
- 8Li R. Yu K. Hildebrand D.F. DGAT 1, DGAT 2 and PDAT expression in seeds and other organs of epoxy and hydroxy fatty acid accumulating plants Lipids 20104514515710.1007/s 11745-010-3385-420101470 · doi ↗ · pubmed ↗