Silencing of the Mycorrhiza-Inducible Phosphate Transporter TaPT3-2D in Wheat Enhances Pathogen Susceptibility and Impairs Arbuscular Mycorrhizal Symbiosis
Yi Zhang, Danfeng Wang, Yuchen Ma, Xueqing Wang, Kedong Xu, Xiaoli Li, Xinxin Shangguan, Haohao Cao, Guozhang Kang, Chengwei Li

TL;DR
A wheat phosphate transporter, TaPT3-2D, is essential for nutrient uptake, mycorrhizal symbiosis, and resistance to fungal diseases.
Contribution
TaPT3-2D is identified as a molecular link connecting phosphate homeostasis, symbiotic signaling, and disease resistance in wheat.
Findings
TaPT3-2D expression is strongly induced by mycorrhizal colonization and fungal infections.
Silencing TaPT3-2D reduces phosphate uptake, mycorrhizal colonization, and increases fungal susceptibility.
TaPT3-2D partially rescues defective phosphate uptake in mutant yeast, confirming its transporter function.
Abstract
The interplay between phosphate (Pi) signaling and defense pathways is crucial for plant fitness, yet its molecular basis, particularly in wheat, remains poorly understood. Here, we functionally characterized the plasma membrane-localized high-affinity phosphate transporter TaPT3-2D and demonstrated its essential roles in Pi uptake, arbuscular mycorrhizal (AM) symbiosis, and fungal disease resistance. Quantitative analyses showed that TaPT3-2D expression was strongly induced by AM colonization (165-fold increase) and by infection with Bipolaris sorokiniana (54-fold increase) and Gaeumannomyces tritici (15-fold increase). In contrast, virus-induced gene silencing (VIGS) of TaPT3-2D reduced Pi uptake and mycorrhizal colonization. Moreover, TaPT3-2D-silenced plants exhibited increased susceptibility to biotrophic, hemibiotrophic, and necrotrophic fungi, accompanied by reduced expression of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
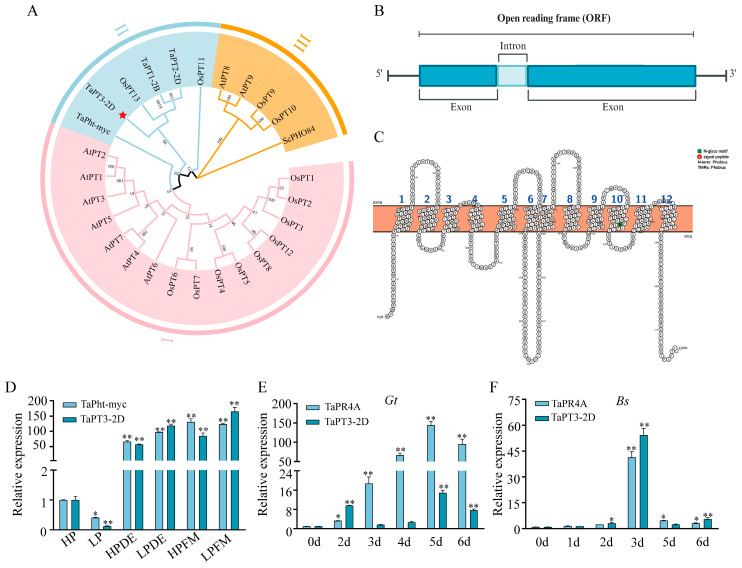 Figure 1
Figure 1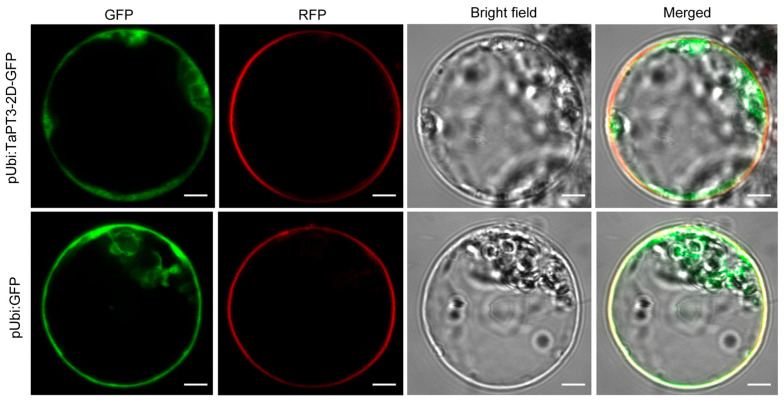 Figure 2
Figure 2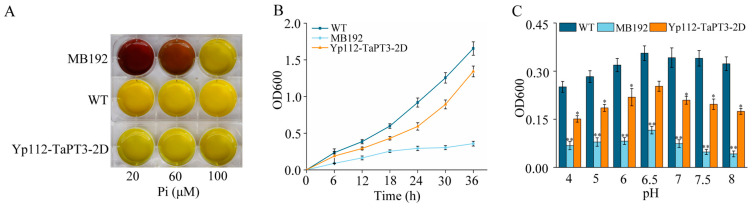 Figure 3
Figure 3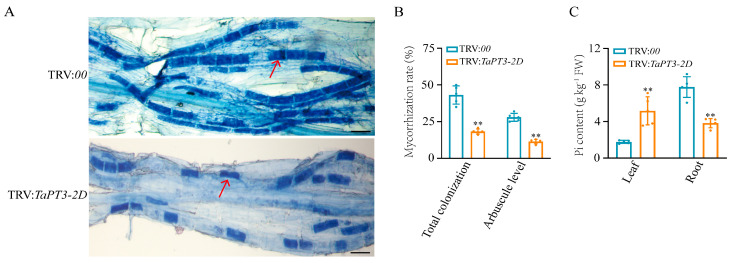 Figure 4
Figure 4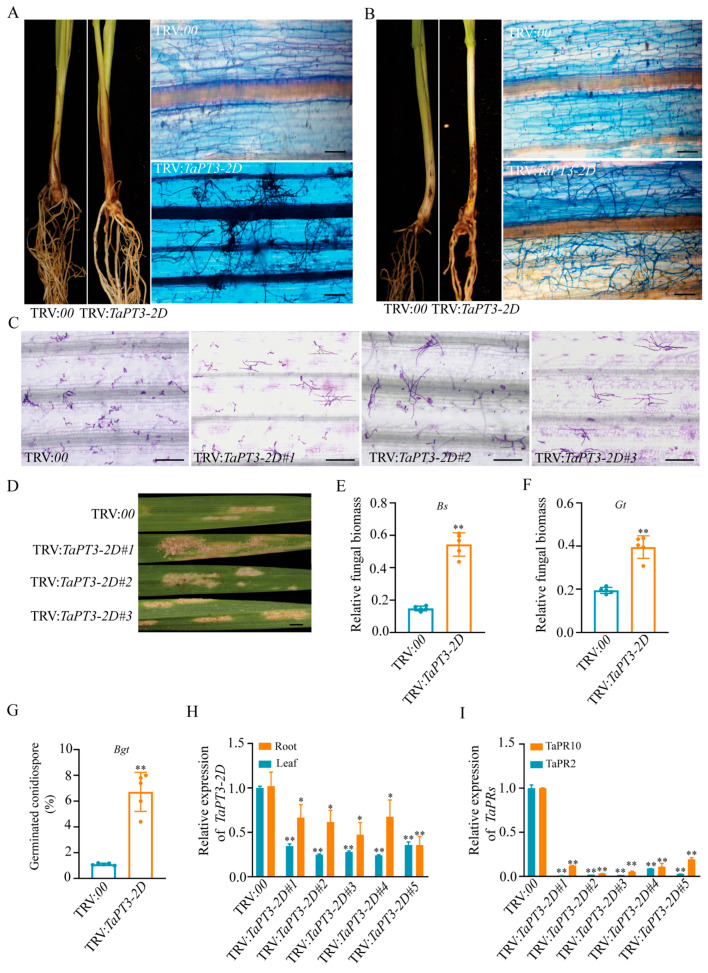 Figure 5
Figure 5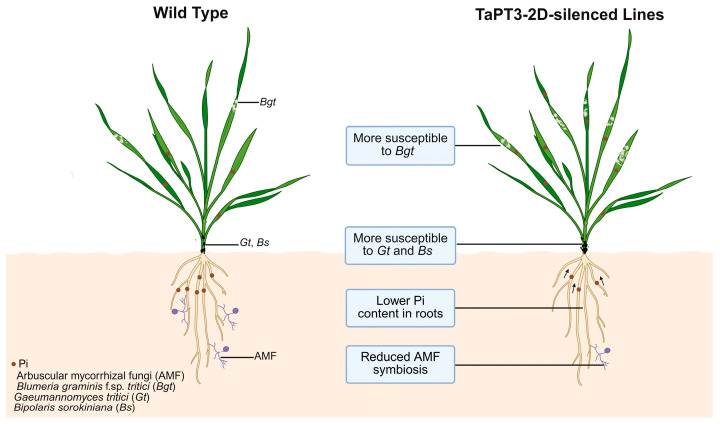 Figure 6
Figure 6- —National Natural Science Foundation of China
- —Department of Science and Technology Planning Project of Henan Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant nutrient uptake and metabolism · Mycorrhizal Fungi and Plant Interactions · Soil Carbon and Nitrogen Dynamics
1. Introduction
As an essential macronutrient, inorganic phosphate (Pi) availability frequently limits the productivity of major cereal crops such as wheat, posing a significant challenge to global food security [1]. This limitation arises from its rapid fixation into insoluble complexes within the soil matrix, inherently low bioavailability, and spatially heterogeneous distribution in natural soil environments [2]. In response, plants have evolved a range of adaptive strategies to optimize Pi acquisition. These include (i) remodeling root system architecture to exploit heterogeneous soil Pi distributions; (ii) secreting acid phosphatases and phytases to hydrolyze organic phosphate esters and mobilize fixed Pi; (iii) establishing arbuscular mycorrhizal (AM) symbioses to enhance Pi absorption via fungal hyphae; and (iv) inducing high-affinity phosphate transporters to facilitate Pi uptake under low-Pi conditions [2].
AM symbioses are evolutionarily conserved mutualistic relationships between Glomeromycotina fungi and more than 70% of extant vascular plant species [3,4]. Functional AM symbioses require host plants to allocate substantial amounts of photosynthetically derived carbon in the form of hexoses and C16:0 β-monoacylglycerol to intraradical fungal structures, sustaining fungal growth and development [5]. Reciprocally, AM fungi efficiently forage for and redistribute water and inorganic nutrients (e.g., Pi and nitrate) through extensive extraradical hyphal networks. These resources are delivered to host roots across symbiotic interfaces via specialized membrane-embedded transporter complexes [5].
Following host root penetration, intraradical hyphae traverse outer cortical cell layers and undergo extensive dichotomous branching to form specialized arbuscules within inner cortical cells [3]. These ephemeral branched structures establish dedicated symbiotic interfaces that facilitate bidirectional molecular signaling during symbiosis establishment and high-efficiency nutrient exchange during the mature arbuscular stage [4]. During arbuscule development, a plant-derived peri-arbuscular membrane (PAM) forms concentrically around the branching fungal hyphae, establishing a physical barrier that segregates the fungal symbiont from the host cytosol and concurrently functions as a dedicated symbiotic resource exchange interface [6]. The PAM is enriched in high-affinity proton-coupled phosphate transporters (predominantly PHT1-family proteins), which are strictly required for unidirectional Pi transfer from the fungus to the plant host [6,7].
The concerted transcriptional induction of multiple phosphate transporters constitutes a core adaptive response to Pi starvation in higher plants [8]. These plasma membrane-localized high-affinity transporters are rapidly upregulated under low-Pi conditions and serve as the primary mediators of Pi acquisition and redistribution, thereby maintaining systemic Pi homeostasis [8,9]. PHT1 orthologs have been identified in numerous plant species, including nine in Arabidopsis thaliana and 13 in Oryza sativa [10,11]. In A. thaliana, AtPT1, AtPT4, AtPT5, AtPT8, and AtPT9 perform distinct roles in Pi absorption and long-distance transport [12,13,14,15]. In rice, OsPT2, OsPT6, OsPT9, and OsPT10 predominantly mediate high-affinity Pi uptake from the rhizosphere during Pi deprivation [16,17]. Notably, the expression of OsPT9 and OsPT10 appears to be largely independent of Pi availability, enabling sustained Pi uptake under Pi-replete conditions and contributing to Pi homeostasis [16]. The allohexaploid bread wheat (Triticum aestivum L.) genome consists of three homoeologous subgenomes (A, B, and D) and therefore contains many homoeologous gene copies [18]. Consistent with this genomic complexity, wheat harbors an expanded phosphate transporter gene family comprising 35 identified members [19]. Among these, overexpression of TaPHT1;4 and TaPT2 significantly enhances rhizosphere Pi acquisition efficiency and promotes biomass accumulation [20,21]. In addition, TaPHT1.9-4B coordinates Pi uptake, long-distance Pi translocation, and growth under Pi starvation [22].
The essentiality of phosphorus for plant growth is undisputed. Accordingly, Pi uptake is tightly regulated to maintain optimal cellular Pi status while balancing colonization by beneficial microbes with constitutive defense readiness and inducible immune responses against fungal pathogens [23,24,25,26]. The acquisition of adequate Pi not only supports normal growth and development but also enhances pathogen defense capacity [23]. For example, foliar application of 50 mM K_2_HPO_4_ to rice increased grain yield by up to 32% while reducing blast disease (Magnaporthe oryzae) incidence by up to 42% [24]. However, plant defense responses under different Pi regimes are highly variable and depend on genetic background, as well as crosstalk between Pi availability and key defense hormone pathways, including salicylic acid (SA), jasmonic acid (JA), and ethylene signaling [25,26,27,28]. In rice, both excessive Pi fertilization and Pi overaccumulation repress the expression of defense-related genes, thereby compromising resistance to M. oryzae compared with sufficient- or low-Pi conditions [25]. In contrast, in A. thaliana, elevated Pi levels are associated with increased resistance to necrotrophic (Plectosphaerella cucumerina) and hemibiotrophic (Colletotrichum higginsianum) fungal pathogens [26,27]. Finally, phosphorus deficiency can suppress plant immunity, potentially favoring associations with plant growth-promoting rhizobacteria (PGPR) that enhance Pi acquisition efficiency and/or alleviate Pi stress [28]. Together, these findings highlight the importance of tightly regulated Pi uptake in maintaining cellular Pi homeostasis while balancing microbial interactions with effective defense against fungal pathogens.
Although the phosphate transport functions of PHT1 family members are well characterized, their direct involvement in immune signaling, especially within symbioses, remains poorly understood. We previously found that six TaPHT1 genes, including the newly identified TaPT3-2D, are constitutively upregulated in AM-colonized wheat roots regardless of external Pi status [19]. This Pi-independent expression suggests that TaPT3-2D regulation may be linked to symbiotic or defense signaling rather than nutrient sensing alone, raising the possibility that it integrates multiple biotic signals. To test this hypothesis, we conducted a functional analysis of TaPT3-2D, focusing on its subcellular localization, biochemical activity as a phosphate transporter, and physiological roles in plant-microbe interactions and Pi homeostasis. Compared with wild-type (WT) plants, TaPT3-2D-silenced wheat exhibited increased susceptibility to biotrophic, hemibiotrophic, and necrotrophic fungal pathogens; impaired AM symbiosis; and reduced Pi accumulation. Together, these results indicate that the mycorrhiza-inducible phosphate transporter TaPT3-2D regulates wheat immunity, AM symbiosis, and Pi uptake.
2. Results
2.1. TaPT3-2D Is an AM-Inducible Phosphate Transporter Gene
We previously performed a genome-wide characterization of the wheat PHT1 gene family and identified TaPT3-2D as a novel AM-inducible gene. Sequence analysis in Chinese Spring wheat revealed that TaPT3-2D maps to the chromosomal locus TraesCS2D02G045700. In addition, the two homologous genes TraesCS2B02G059100 and TraesCS2D02G045600 were identified and designated TaPT1-2B and TaPT2-2D, respectively [19]. Notably, the coding sequences (CDS) of these three homologous genes (TaPT1-2B, TaPT2-2D, and TaPT3-2D) share 80.6% nucleotide sequence identity (Figure S1).
Phylogenetic analysis incorporating previously characterized rice PHTs (Table S1) demonstrated that TaPT1-2B, TaPT2-2D, TaPT3-2D, TaPht-myc (a mycorrhizal-specific PHT1 protein), and two rice PHT1 members (OsPT11 and OsPT13) cluster within the same phylogenetic group (Figure 1A). The AM-specific or AM-inducible transporters OsPT11 and OsPT13 are known to mediate the development of AM symbioses and Pi acquisition [29]. Notably, TaPT3-2D and OsPT13 represent a distinct subcluster.
TaPT1-2B, TaPT2-2D, and TaPT3-2D share high protein sequence similarity (76.89%) (Figure S2) and exhibit significantly greater homology with OsPT13 (72.64–81.94%) than with OsPT11 (60.38–62.83%) (Figure S3, Table S2). The TaPT3-2D gene contains one intron, and its 1566 bp CDS encodes a 521 aa protein (Figure 1B). The intron-exon boundary conforms to the canonical GT-AG splicing rule, as confirmed by reference annotation in WheatOmics using JBrowse. In silico analysis predicted that TaPT3-2D contains 12 transmembrane (TM) domains (Figure 1C).
2.2. Expression Profiling of TaPT3-2D and Its Homologous Genes
Transcript abundance of TaPT3-2D was markedly upregulated upon colonization by two AM fungi, with the strongest induction observed under low-Pi conditions in plants colonized by Diversispora epigaea and Funneliformis mosseae. This expression pattern supports the classification of TaPT3-2D as an AM-inducible phosphate transporter. Under low-Pi conditions, TaPT3-2D expression in D. epigaea- and F. mosseae-colonized plants significantly exceeded that of TaPht-myc, the first identified mycorrhizal-specific PHT1 gene in wheat (Figure 1D). In contrast, expression of TaPT1-2B and TaPT2-2D was significantly repressed by AM colonization, particularly in plants colonized by D. epigaea (Figure S4A).
To examine whether the expression of these three TaPHT genes is also affected by pathogen infection, wheat roots were inoculated with Bipolaris sorokiniana (Bs) and Gaeumannomyces tritici (Gt). As a well-established marker of pathogen challenge, TaPR4A exhibits diagnostic expression patterns during infection. Following Gt inoculation, TaPR4A transcription increased progressively and peaked at 5 days post inoculation (DPI) before declining modestly at 6 DPI. Expression of TaPT2-2D and TaPT3-2D was induced at 2 DPI, peaked at 5 DPI, and decreased by 6 DPI, whereas TaPT1-2B was significantly induced only at 2 DPI (Figure 1E and Figure S4B). Following Bs infection, TaPR4A and TaPT3-2D were significantly upregulated at 3 DPI and declined thereafter (Figure S4C). In contrast, TaPT1-2B and TaPT2-2D exhibited strong induction at 5 and 6 DPI. Given the observed involvement of TaPT3-2D in both AM symbiosis and pathogenic fungal infection, we next sought to elucidate its biological function.
2.3. Subcellular Localization of TaPT3-2D
Given that phosphate transporters are typically localized to the plasma membrane, we assessed the subcellular localization of TaPT3-2D by transiently expressing a C-terminal GFP fusion under the control of the ubiquitin promoter in rice protoplasts. Consistent with the known properties of PHT1 proteins, TaPT3-2D-GFP fluorescence was observed exclusively at the plasma membrane, contrasting with the diffuse cytosolic and nuclear distribution of free GFP controls (Figure 2). These results therefore validate TaPT3-2D as a plasma membrane-localized phosphate transporter.
2.4. Complementation of TaPT3-2D in Yeast
The phosphate transporter activity of TaPT3-2D was evaluated using the yeast mutant MB192 transformed with p112A1NE-TaPT3-2D. Acid phosphatase activity staining revealed dose-dependent complementation at 20 and 60 μM Pi, suggesting that TaPT3-2D mediates Pi transport (Figure 3A). Growth kinetics in low-Pi YNB medium (60 µM) positioned TaPT3-2D-complemented MB192 cells between WT and mutant phenotypes, supporting partial functional recovery (Figure 3B). pH profiling identified pH 6.5 as optimal for TaPT3-2D-dependent Pi uptake, with complementation efficiency declining under both acidic and alkaline extremes (Figure 3C).
2.5. TaPT3-2D Is Required for AM Symbiosis and Pi Uptake
To characterize the role of TaPT3-2D in AM symbiosis, TaPT3-2D knockdown lines and control plants were inoculated with F. mosseae. Both the total colonization rate and arbuscule abundance were significantly lower in TaPT3-2D knockdown lines than in control plants. Mature arbuscules were observed in both control and virus-induced gene silencing (VIGS) plants, with no obvious differences in arbuscule morphology observed (Figure 4A,B). To further examine the role of TaPT3-2D in Pi transport, Pi concentrations were measured in roots and shoots. TaPT3-2D-silenced plants exhibited significantly lower Pi concentrations in roots, accompanied by increased Pi concentrations in shoots, suggesting that TaPT3-2D contributes to root-to-shoot Pi transport (Figure 4C).
2.6. Silencing of TaPT3-2D Increases Susceptibility to Fungal Pathogens
Given the strong induction of TaPT3-2D in response to pathogenic fungal infection, tobacco rattle virus (TRV)-based VIGS-mediated silencing of TaPT3-2D was performed to verify its role in wheat defense against pathogens. Following inoculation with the root pathogens Gt and Bs at 16 days after silencing, TaPT3-2D-VIGS lines exhibited accelerated disease progression. These plants showed significantly increased susceptibility to Gt at 21 DPI and to Bs at 40 DPI compared with control plants (Figure 5A,B). Consistently, both pathogens accumulated significantly more biomass in TaPT3-2D-silenced lines than in WT controls (Figure 5E,F).
Because the obligate biotroph Blumeria graminis f.sp. tritici (Bgt) employs a life strategy distinct from that of Gt and Bs, we further examined resistance to Bgt in TaPT3-2D-silenced wheat. The fourth leaves of TaPT3-2D-silenced and control (TRV:00) plants were harvested at 16 DPI and inoculated with Bgt conidiospores. Colonization efficiency at 60 h post-infection (HPI) was significantly higher in silenced lines compared to vector controls (Figure 5C–G). Macroscopic examination revealed extensive mycelial colonization of TaPT3-2D-silenced leaves (Figure 5D), consistent with microscopic analyses. Collectively, these results demonstrate that suppression of TaPT3-2D compromises wheat resistance to Bgt infection.
qRT-PCR validation confirmed efficient silencing of TaPT3-2D, with significantly reduced transcript abundance observed in both the roots and shoots of silenced plants compared to WT controls (Figure 5H). Concomitant with enhanced disease susceptibility, TaPT3-2D-silenced plants also exhibited marked downregulation of the defense-related genes TaPR2 and TaPR10 (Figure 5I). Together, these findings suggest that silencing of TaPT3-2D increases susceptibility to pathogens and impairs AM symbiosis and Pi uptake (Figure 6).
3. Discussion
3.1. TaPT3-2D Is a Novel Mycorrhizal-Induced PHT1 Similarly to OsPT13
Phylogenetic analysis revealed that TaPT3-2D and its two homologs form a monophyletic clade with three previously characterized AM-specific or AM-inducible phosphate transporters: OsPT13, OsPT11, and TaPht-myc (Figure 1A). Among these, TaPT3-2D exhibits the highest protein sequence similarity with OsPT13 (72.6%), followed by TaPht-myc (66.1%) and OsPT11 (62.3%) (Table S2). Notably, TaPT3-2D shares significantly higher sequence similarity with OsPT13 than with TaPht-myc, suggesting that TaPT3-2D is a functional ortholog of OsPT13 with a likely conserved role in mycorrhizal Pi transport.
Topological modeling further indicated that TaPT3-2D and OsPT13 both contain 12 TM domains, including a prominent hydrophilic loop linking TM6 and TM7, while both termini localize to the cytosol (Figure 1C) [29]. Previous studies have shown that OsPT13 is evolutionarily conserved exclusively in monocots, whereas OsPT11 exhibits cross-clade conservation across both monocots and dicots. Consistent with its mycorrhizal specificity, OsPT13 is upregulated 14-fold in Glomus intraradices-inoculated roots [30]. In comparison, TaPT3-2D expression increased 57- to 165-fold in wheat roots colonized by F. mosseae or D. epigaea across different Pi regimes. The exceptionally strong induction of TaPT3-2D (>100-fold) by these AM fungi highlights its central role in AM-mediated Pi uptake in wheat. In contrast, neither TaPT1-2B nor TaPT2-2D was significantly induced by AM colonization, indicating functional divergence among these homologous genes (Figure S4A). Guided by these findings, we next sought to further characterize the role of TaPT3-2D in Pi acquisition and AM symbiosis.
3.2. TaPT3-2D Is a High-Affinity Phosphate Transporter Integral to Pi Assimilation and AM Symbiosis
To verify the role of TaPT3-2D in Pi transport, we employed the high-affinity phosphate transporter-deficient yeast strain MB192 (Δpho84), a well-established system for functional validation of plant and fungal phosphate transporters [22,31,32,33]. Functional complementation with TaPT3-2D partially restored MB192 growth under low-Pi conditions (Figure 3), confirming its activity as a high-affinity phosphate transporter. The incomplete recovery of growth likely reflects limitations inherent to heterologous expression systems, including differences in transcriptional regulation, protein folding, and post-translational modification between plants and yeast [34].
VIGS provides a rapid and efficient approach for transient gene knockdown and functional analysis in plants, and TRV-based systems have been successfully applied in both monocots and dicots [35,36,37]. In TaPT3-2D-silenced wheat, Pi concentrations were significantly reduced in roots but increased in shoots, suggesting that TaPT3-2D contributes to root-to-shoot Pi transport. Several non-mutually exclusive mechanisms may explain this phenotype. First, silencing TaPT3-2D may induce a systemic Pi starvation response, leading to compensatory upregulation of other high-affinity Pi transporters (e.g., other TaPHT1 members) or genes involved in phloem loading. Such responses could prioritize Pi allocation to shoots to sustain growth despite impaired root Pi acquisition or symplastic transport. Second, reduced AM colonization (Figure 4A,B) likely decreases Pi sink strength in roots, resulting in a greater proportion of acquired Pi being translocated to shoots. Finally, TaPT3-2D may play a direct role in Pi loading or unloading within roots, thereby modulating root-to-shoot translocation efficiency. Further studies examining the expression of other phosphate transporters in the TaPT3-2D-silenced background will be required to distinguish among these possibilities.
In addition to altered Pi distribution, TaPT3-2D-silenced plants exhibited significantly reduced total AM colonization rates and arbuscule abundance. Although these results demonstrate that TaPT3-2D is required for normal AM symbiosis, the functional consequences of this morphological impairment on nutrient exchange remain to be determined. Future studies using radioisotope tracers (e.g., ^33^P) will be essential to directly quantify mycorrhiza-mediated Pi uptake in TaPT3-2D-deficient plants. In parallel, analyses of carbon allocation, including the expression of symbiosis-associated sugar transporters and carbon flux to the fungal partner, will provide a more comprehensive assessment of symbiotic efficiency regulated by TaPT3-2D.
Although TRV-mediated VIGS enabled efficient initial functional characterization of TaPT3-2D in wheat, definitive validation of its biological role will require stable genetic lines. Future studies employing CRISPR/Cas9-mediated knockout and overexpression approaches will be essential to exclude potential confounding effects associated with transient silencing and to facilitate long-term investigations of Pi homeostasis, AM symbiosis, and immune responses.
3.3. TaPT3-2D Is Involved in the Regulation of Wheat Innate Immunity
TaPT3-2D is a member of the PHT1 family, one of the most extensively characterized phosphate transporter families in wheat. As expected, the majority of research on PHT1 genes has focused on the roles of these genes in Pi uptake [20,21,22,38], whereas their functions in mycorrhizal symbiosis and plant immunity remain underexplored. In contrast, members of the PHT3 and PHT4 subfamilies have been more extensively examined for their roles in disease resistance [39,40,41]. For example, silencing of phosphate transporter AtPHT4;1 enhances susceptibility to the virulent pathogen Pseudomonas syringae in A. thaliana [40]. Independent studies have established AtPHT4;1 as a dual-function node coordinating pathogen defense and circadian regulation, and its expression is directly controlled by the core clock component CCA1 [42]. Similarly, silencing of the mitochondrial phosphate transporter TaMPT (a PHT3 family member) increases susceptibility to Fusarium head blight in wheat [39]. Consistent with these findings, we found that silencing TaPT3-2D increases wheat susceptibility to biotrophic, hemibiotrophic, and saprophytic fungi.
Notably, members of the same phosphate transporter family can exert contrasting effects on disease resistance. Although AtPHT4;6 and AtPHT4;1 both belong to the PHT4 family, silencing AtPHT4;6 significantly enhances resistance to virulent P. syringae [41], underscoring functional divergence within this gene family. Similarly, atpht1;4 mutants exhibit decreased susceptibility to the soil-borne bacterial pathogen Ralstonia solanacearum [43], while overexpression of the PHT1 family gene OsPT8 suppresses pattern-triggered immunity and attenuates resistance to rice blast (M. oryzae) and bacterial blight (Xanthomonas oryzae pv. oryzae) [44]. Collectively, although the effects of Pi transporters on disease susceptibility vary across pathosystems, these findings converge to support a functional link between PHT-mediated Pi uptake and plant immunity.
MicroRNAs (miRNAs) are also key regulators of Pi homeostasis, particularly miR399 and miR827. Overexpression of miR399 in A. thaliana and rice leads to Pi accumulation in leaves and increased susceptibility to M. oryzae [25,27]. Consistent with these reports, TaPT3-2D-silenced wheat exhibited elevated leaf Pi levels, enhanced pathogen susceptibility, and reduced AM symbiosis (Figure 4 and Figure 5). However, the mechanism by which plants integrate Pi status and microbial signals to coordinate defense and symbiosis remains elusive. Elucidating the molecular function of TaPT3-2D in Pi homeostasis and plant-microbe interactions will require protein–protein interaction analyses using yeast two-hybrid (Y2H), glutathione-S-transferase (GST) pull-down, bimolecular fluorescence complementation (BiFC), and firefly luciferase complementation imaging assays both in vivo and in vitro.
Although our data establish a link between TaPT3-2D, Pi homeostasis, AM symbiosis, and disease susceptibility, the underlying causal pathway remains to be resolved. The enhanced pathogen susceptibility observed in TaPT3-2D-silenced wheat may result from (i) direct disruption of Pi-dependent immune signaling, (ii) indirect effects caused by impaired AM symbiosis and the loss of symbiosis-associated defense benefits, or (iii) an independent, as-yet-unidentified role of TaPT3-2D in immune regulation. Future studies employing Pi supplementation assays in TaPT3-2D mutant backgrounds will be critical for distinguishing among these possibilities and determining whether altered immune responses primarily reflect changes in local Pi status or compromised symbiotic interactions.
Finally, although our results indicate that TaPT3-2D is required for full induction of defense-related genes such as TaPR2 and TaPR10, the upstream signaling pathways affected by TaPT3-2D silencing remain unknown. Future work should systematically characterize the dynamics of key defense hormones, including SA, JA, and ethylene, and evaluate the expression of core pattern-triggered immunity (PTI) components in this genetic background. Such analyses will be essential to determine whether TaPT3-2D modulates immunity through specific hormonal pathways or by broadly influencing early immune perception and signaling, thereby defining a more precise molecular connection between phosphate transport and immune regulation.
4. Materials and Methods
4.1. Plant Materials and Fungal Inoculation
The Chinese winter wheat cultivar ‘Zhoumai 26’ was used for all experiments. Seedlings were transplanted into pots containing a sterilized mixture of soil and river sand at a 1:2 (v/v) ratio. To establish AM symbioses, autoclaved sand-based inocula containing 60 g of F. mosseae (Nicol. & Gerd, BGC XZ02A, isolated from the rhizosphere of Dangxiong, Tibet, China) or D. epigaea (BGC NM04B, formerly Glomus versiforme) were applied to 10 seedlings per treatment. Control plants received an equivalent amount of autoclaved inoculum. Both experimental and control plants were irrigated biweekly with half-strength Hoagland’s nutrient solution amended with either 5 μM (low-Pi) or 500 μM (high-Pi) KH_2_PO_4_. The experiment consisted of six treatment groups: high-Pi control (HP, 500 µM KH_2_PO_4_), low-Pi control (LP, 5 µM KH_2_PO_4_), high-Pi inoculated with D. epigaea (HPDE), low-Pi inoculated with D. epigaea (LPDE), high-Pi inoculated with F. mosseae (HPFM), and low-Pi inoculated with F. mosseae (LPFM). Tissue samples were collected at 42 DPI, immediately snap-frozen in liquid nitrogen, and stored at −80 °C for subsequent RNA extraction. The hemibiotrophic root rot pathogen Bs, provided by Henan Agricultural University, was cultured on potato dextrose agar (PDA) at 20 °C for 15 days. Ten-day-old seedlings were inoculated by applying Bs spore suspensions (4 × 10^4^ spores mL^−1^) to stem bases according to a previously described protocol [45]. Root samples were collected at 0, 1, 2, 3, 5, and 6 DPI for RNA extraction. A separate set of seedlings was inoculated with the necrotrophic pathogen Gt (formerly Gaeumannomyces graminis var. tritici), provided by Henan Academy of Agricultural Science, according to a previously described protocol [46]. Root tissues were collected at 0, 2, 3, 4, 5, and 6 DPI for RNA extraction. Pathogen-inoculated plants were maintained in a growth chamber at 70% relative humidity and 22 °C during the light period (16 h) and 15 °C during the dark period (8 h). The biotrophic fungal pathogen Bgt was maintained on ‘Aikang 58’ wheat at 22 °C during the light period (16 h) and 15 °C during the dark period (8 h). The pathogenesis-related 4 family gene TaPR4A, which is strongly induced by multiple fungal pathogens, was utilized as a molecular marker for pathogenicity [47].
Experiments involving AM colonization and pathogen challenge were conducted independently, and samples from each treatment were processed and analyzed separately to avoid confounding effects. Each treatment consisted of 10 individual plants, and all experiments were performed with three biological replicates.
4.2. Subcellular Localization of TaPT3-2D in Rice Protoplasts
The full-length TaPT3-2D CDS was cloned using KAPA HiFi HotStart ReadyMix (Roche, Basel, Switzerland) and inserted downstream of the GFP gene in the pUbi-GFP-GW vector via Gateway recombination, generating the TaPT3-2D-GFP fusion vector under the transcriptional control of the ubiquitin promoter. The recombinant construct was verified by Sanger sequencing and subsequently introduced into Agrobacterium tumefaciens strain GV3101. The empty GFP vector was transformed in parallel as a control. Rice protoplasts were isolated from 10-day-old rice seedlings, and protoplast transformation was performed using a previously published protocol [48]. The TaPT3-2D-GFP fusion construct or GFP control was introduced into protoplasts, which were then incubated in the dark at 28 °C for 16 h. Subcellular localization of the expressed proteins was examined using a confocal laser scanning microscope (Stellaris 8, Leica, Wetzlar, Germany), with FM4-64 staining employed as a plasma membrane marker. The green channel was excited at 488 nm with emission collected between 500–547 nm; the red channel was excited at 587 nm via a white-light laser line with emission collected between 600–632 nm. Images were assembled for presentation in Adobe Illustrator 2022.
4.3. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
Total RNA was isolated using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer’s protocol. Genomic DNA contamination was removed by treatment with RNase-free DNase I (Takara Bio Inc., Kusatsu, Japan). Total RNA was reverse transcribed to synthesize cDNA using the PrimeScript RT Perfect Real Time Kit (Takara Bio Inc., Kusatsu, Japan). qRT-PCR was performed on a CFX^96^ Touch Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA) using SYBR^®^ Premix Ex Taq™ (Takara Bio Inc., Kusatsu, Japan). Relative gene expression levels were quantified using the 2^−ΔΔCt^ method [49]. Each sample was analyzed with three independent biological replicates and three technical repeats.
4.4. Functional Analysis of TaPT3-2D in Yeast
To ascertain the biological function of TaPT3-2D, the Pi uptake-deficient yeast mutant MB192 was transfected with the p112A1NE expression vector harboring TaPT3-2D, following a previously described method [50]. Briefly, the TaPT3-2D CDS was subcloned into the p112A1NE vector to generate the recombinant TaPT3-2D-p112A1NE construct, which was subsequently introduced into MB192 yeast. Next, TaPT3-2D-p112A1NE-harboring MB192 cells, WT cells, and untransformed MB192 cells were cultured in yeast nitrogen base (YNB) liquid medium. Upon reaching the logarithmic growth phase, cells were harvested by centrifugation and washed twice with Pi-free YNB. Next, the three yeast strains were separately inoculated into YNB liquid medium supplemented with 20, 60, or 100 µM KH_2_PO_4_ and incubated at 30 °C for 10 h. Bromcresol purple was employed as a pH indicator, with a medium color transition from brown-red to yellow signaling acidification as a correlate of yeast growth. For quantitative growth analysis, MB192, WT, and Yp112-TaPT3-2D yeast cells were cultured in YNB liquid medium supplemented with 60 μM KH_2_PO_4_. Cell cultures were harvested at six time points (6, 12, 18, 24, 30, and 36 h) to quantify the optical density at 600 nm (OD_600_). To examine the impact of pH on Pi uptake, each yeast strain was cultured in YNB liquid medium supplemented with 60 μM KH_2_PO_4_ and adjusted to different pH values (4, 5, 6, 6.5, 7, 7.5, and 8). Following a 24 h incubation period, OD_600_ was measured to assess cell growth. Each assay was performed with three independent biological replicates per strain.
4.5. VIGS Analysis of TaPT3-2D in Wheat
To validate the role of TaPT3-2D in AM symbiosis and Pi transport, gene silencing was performed using a TRV-based VIGS system. A highly conserved 300 bp segment of TaPT3-2D was chosen as the target region and amplified from ‘Zhoumai 26’ wheat. Subsequently, the In-Fusion HD Cloning Plus system was employed to perform recombination cloning, enabling the insertion of the TaPT3-2D-carrying pYL156 plasmid into the TRV vector to generate the TRV:TaPT3-2D construct. The empty pYL156 vector was similarly introduced into the TRV genome to generate the negative control TRV:00. To generate gene-silenced seedlings, ‘Zhoumai 26’ seeds were subjected to a modified whole-plant VIGS protocol [51], with unsilenced controls harboring the TRV:00 construct. At 16 DPI, total RNA was isolated from the roots and leaves of both VIGS-silenced and unsilenced plants to analyze TaPT3-2D expression.
4.6. Functional Verification of TaPT3-2D-Silenced Plants in Response to Fungal Infection
To verify the role of TaPT3-2D in the wheat response to fungal infection, 30 VIGS-silenced wheat seedlings were individually inoculated with either biotrophic symbiotic AM fungi, the hemi-biotrophic pathogen Bs, the necrotrophic pathogen Gt, or the biotrophic pathogen Bgt. VIGS-silenced and TRV:00 control plants were inoculated with F. mosseae. At 42 DPI, freshly harvested AM-colonized roots were collected and stained with trypan blue following a previously described protocol [52]. The frequencies of total and AM colonization were quantified for 30 root samples by magnifying intersections under a BX61 fluorescence microscope equipped with a 20× objective lens. Microscopic images of pathogens were similarly visualized using the same microscope configuration.
Pathogen inoculation procedures were performed as described in Section 4.1. For pathogen biomass quantification, qPCR was conducted using primers specific to the 18S rRNA genes of Gt (accession number: FJ771002) and Bs (accession number: KM066949). TaActin gene (accession number: KC775780) was utilized as the internal reference gene. Primers used for qRT-PCR are provided in Table S3. Finally, the total Pi concentration was quantified in fresh leaf and root samples from TaPT3-2D-silenced and control plants [53].
4.7. Statistical Analysis
Statistical analyses were performed using one-way analysis of variance (ANOVA) to evaluate differences in relative gene expression among treatments and time points, as well as phenotypic variation among genotypes. When ANOVA indicated significant effects (p < 0.05), Duncan’s multiple range test was used for post hoc comparisons. Least significant difference (LSD) values were calculated at the 95% and 99% confidence levels to support the analysis. Significant differences are indicated in the figures, and corresponding LSD values are provided in the figure legends. All statistical analyses were conducted using SPSS version 23.
5. Conclusions
In this study, we functionally characterized TaPT3-2D, an AM-inducible phosphate transporter in wheat. Heterologous expression in Δpho84 mutant MB192 yeast rescued Pi uptake deficiency, suggesting that TaPT3-2D functions as a phosphate transporter. VIGS-mediated suppression of TaPT3-2D attenuated pathogen resistance, reduced AM symbiosis, and decreased the Pi content in roots. Collectively, these findings demonstrate that TaPT3-2D serves as a key integrator of phosphate nutrition, symbiotic signaling, and disease resistance in wheat.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gao Z.Y. Cao H.B. Huang M. Bao M. Qiu W.H. Liu J.S. Winter wheat yield and soil critical phosphorus value response to yearly rainfall and P fertilization on the Loess Plateau of China Field Crops Res.202329610892110.1016/j.fcr.2023.108921 · doi ↗
- 2Kumar R. Ngangkham U. Abdullah S.N.A. Editorial: Phosphorus starvation in plants Front. Plant Sci.202314121143910.3389/fpls.2023.121143937304720 PMC 10250812 · doi ↗ · pubmed ↗
- 3Genre A. Lanfranco L. Perotto S. Bonfante P. Unique and common traits in mycorrhizal symbioses Nat. Rev. Microbiol.20201864966010.1038/s 41579-020-0402-332694620 · doi ↗ · pubmed ↗
- 4Rich M.K. Vigneron N. Libourel C. Keller J. Xue L. Hajheidari M. Radhakrishnan G.V. Le Ru A. Diop S.I. Potente G. Lipid exchanges drove the evolution of mutualism during plant terrestrialization Science 202137286486810.1126/science.abg 092934016782 · doi ↗ · pubmed ↗
- 5Jiang Y. Wang W. Xie Q. Liu N. Liu L. Wang D. Zhang X. Yang C. Chen X. Tang D. Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi Science 20173561172117510.1126/science.aam 997028596307 · doi ↗ · pubmed ↗
- 6Oldroyd G.E.D. Leyser O. A plant’s diet, surviving in a variable nutrient environment Science 2020368 eaba 019610.1126/science.aba 019632241923 · doi ↗ · pubmed ↗
- 7Shi J. Wang X. Wang E. Mycorrhizal symbiosis in plant growth and stress adaptation: From genes to ecosystems Annu. Rev. Plant Biol.20237456960710.1146/annurev-arplant-061722-09034236854473 · doi ↗ · pubmed ↗
- 8Gu M. Chen A. Sun S. Xu G. Complex regulation of plant phosphate transporters and the gap between molecular mechanisms and practical application: What is missing?Mol. Plant 2016939641610.1016/j.molp.2015.12.01226714050 · doi ↗ · pubmed ↗