Selenium (IV) and Sulphur (VI) as Elements Modifying Plant Quality: Content of Selenium and Sulphur Forms in Wheat
Marzena S. Brodowska, Magdalena Kurzyna-Szklarek, Mirosław Wyszkowski

TL;DR
This study shows how adding selenium and sulfur to wheat can improve its nutritional quality without causing toxicity.
Contribution
The study identifies optimal selenium doses and application timing for biofortifying wheat without affecting sulfur levels.
Findings
Selenium fertilization increased selenium content in wheat grain without causing toxicity.
Sulfur application increased total, sulfate, and organic sulfur in wheat grain and straw.
Optimal selenium dose for biofortification is 20 mg Se ha−1 applied during stem elongation.
Abstract
In order to achieve high-quality yields, it is essential to provide plants with the necessary nutrients, including selenium (Se) and sulphur (S), to meet their nutritional requirements. This study aimed to determine the effect of selenium (0, 10 and 20 g Se ha−1) and the date of its application (in the tillering phase and in the stem elongation phase) and sulphur application (0, 15 and 30 kg S ha−1) on the content of selenium and various forms of sulphur (total sulphur, sulphate sulphur and organic sulphur) and the N:S ratio in winter spelt wheat and winter common wheat. The research hypothesis assumed that different doses of selenium and sulphur and the timing of their application would have a beneficial effect on the Se and S content in the grain and straw of spelt wheat and common wheat. Selenium fertilisation significantly increased the content of this element in the grain of spelt…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
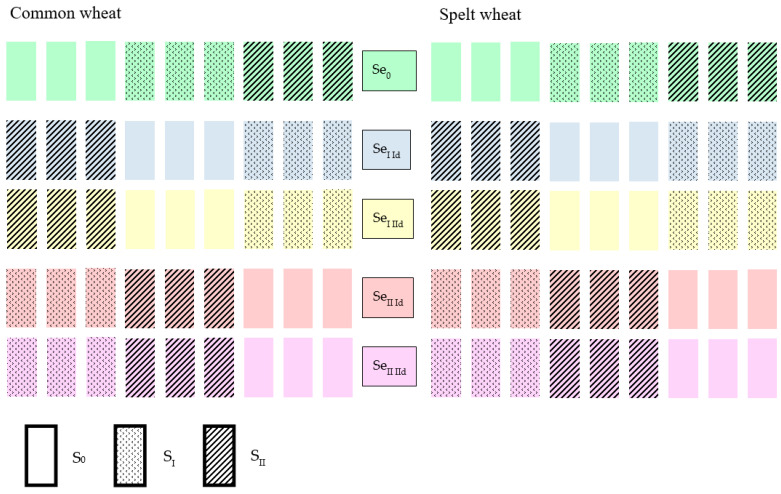 Figure 1
Figure 1- —University of Life Sciences in Lublin, Faculty of Agrobioengineering, Department of Agricultural and Environmental Chemistry
- —Minister of Science
- —University of Warmia and Mazury in Olsztyn, Faculty of Agriculture and Forestry, Department of Agricultural and Environmental Chemistry
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSelenium in Biological Systems · Nitrogen and Sulfur Effects on Brassica · Silicon Effects in Agriculture
1. Introduction
Since the 1960s, selenium (Se) fertilisation of crops has attracted the interest of researchers around the world [1,2]. Most of the experiments conducted involved the use of various selenium salts in the form of soil fertilisers or in combination with phosphorus and nitrogen fertilisers [3,4]. The method of application, but also the fertilisation strategy, are considered key issues in selenium fertilisation, as the responses of crops depend on the fertiliser dose, Se speciation, and the method and timing of application [5,6]. Knowledge of the impact of fertiliser type, strategy and timing allows the development of sustainable fertilisation strategies targeted at site-specific causes of Se deficiency [7]. It is believed that crops respond more strongly to fertilisers containing selenate (VI) compared to fertilisers with selenite (IV), mainly due to its greater solubility and availability in plants [5]. The Se level in plants increases quite rapidly after fertilisation, but also decreases when the Se level in the soil solution is reduced. Its half-life is estimated to be 21 to 80 days in grassland ecosystems [8,9], but other researchers have observed a positive response in crops even 3 years after application [2]. Foliar application is considered to be almost twice as effective as granular soil fertilisers or seed enrichment treatments [6,10].
The average Se level in wheat in the United Kingdom and Scandinavian countries ranges from 7 to 22 μg kg^−1^ d.m. [11], and the nutritional value considered adequate for animal and human health is approximately 50 to 100 μg kg^−1^ [12]. Therefore, an increase in Se levels in crops of approximately 50% as a result of optimal fertilisation is considered sufficient to prevent Se deficiency in humans and animals. Assuming a 10% recovery of selenium from fertilisers [13], a wheat yield of approximately 7 tonnes ha^−1^ and the fact that only about 50% of the added selenium ends up in the grains, the recommended selenium dose is approximately 4 to 13 g Se ha^−1^. This is considered sufficient to achieve the required nutritional value. This estimated fertiliser dose is also within the typical fertiliser doses recommended by Finnish and Canadian guidelines [14,15,16,17]. Other authors have reported slightly higher recovery rates, ranging from 14 to 18% in cereals [18], 8 to 32% in wheat [5] and 5 to 25% in cereal crops [19]. In contrast, the results of Tolu et al. [9] and Guo et al. [3] suggest that even more than 90% of the Se applied is unavailable and cannot be taken up by crops. This low level of Se recovery when applied foliar can be partly explained by the fact that it is usually applied during the growing season before intensive leaf development, resulting in a combination of Se applied to the leaves and to the soil. Nevertheless, foliar application of Se is considered to be on average 8 times more efficient than soil application, which is why foliar application is preferred over soil application [6,19].
Fertilisation with macronutrients can affect the Se content in crops [2]. This is related to the competition between SO_4_^2−^ and PO_4_^3−^ ions and Se in terms of uptake by plants [20,21]. In the case of selenium fertilisers, a strong tendency to reduce the response of crops to Se fertilisation with increasing doses of S and P has been observed. The use of selenite (IV) fertilisers weakens this tendency. The observed reduction in Se uptake due to sulphates is explained by ion competition for transport pathways in plant roots [22]. The reducing effect of sulphate ions has been reported in previous studies [23,24,25,26].
Selenium and sulphur (S) are chemically very similar, which affects their uptake and metabolism in plants. Both elements occur in the soil mainly in the form of anions—selenates and sulphates—and compete for the same transport systems in the roots [27,28]. Selenium uptake occurs mainly through sulphate transporters in the plasma membrane of roots, and its further conversion follows the sulphur metabolic pathway [29]. High sulphur availability in the rhizosphere limits selenate uptake due to competitive antagonism at the transporter level [30,31]. On the other hand, sulphur deficiency may increase selenium absorption, as plants under sulphur deficiency conditions intensify the expression of sulphate transporters, which promotes the influx of selenates [30]. This phenomenon is observed, among others, in Arabidopsis thaliana, where an increase in sulphate concentration in the substrate reduces Se accumulation in shoots, while improving biomass growth [30]. After entering the plant, selenium enters the sulphur assimilat—ion pathway, where enzymes such as ATP-sulfurylase and APS reductase act on both sulphates and selenates [32]. This biochemical convergence allows selenium to replace sulphur in amino acids, forming selenocysteine and selenomethionine, which are incorporated into proteins [28]. However, such substitution often disrupts protein functions, which at high Se concentrations leads to toxicity [27].
Recent studies confirm that sulphur fertilisation strategies significantly affect the effects of selenium biofortification. For example, adequate sulphur supply reduces Se accumulation in wheat grain, while sulphur deficiency increases selenium uptake but may limit plant growth [33,34]. This indicates the need to balance S and Se supply in order to achieve optimal crop quality and health benefits.
Selenium and sulphur play an irreplaceable role in wheat metabolism, improving physiological processes, protein synthesis and stress resistance. Adequate selenium fertilisation enhances the activity of antioxidant enzymes, accelerates root development and increases the accumulation of selenium in organic forms—mainly selenomethionine—in the grain [35]. Sulphur, on the other hand, is crucial for the synthesis of sulphur amino acids and proteins, enhancing photosynthetic efficiency and supporting yield growth [36,37]. Selenium deficiency can lead to reduced resistance to environmental stress, weakened antioxidant protection and an increased risk of nutritional deficiencies in consumers. On the other hand, excess selenium can be toxic, but intelligent dosing—according to current research—allows this risk to be avoided [38]. Sulfur deficiency results in reduced protein content and nitrogen metabolism disorders, while adequate supply promotes synergistic interactions with nitrogen and selenium, while supporting the efficient use of nutrients [37,39]. As a result, the optimal use of selenium and sulphur as part of integrated fertilisation contributes to improved plant growth, grain quality and nutritional value.
Research indicates clear differences between spelt wheat (Triticum spelta L.) and common wheat (Triticum aestivum L.) in terms of metabolism and micronutrient uptake, which affects the effectiveness of biofortification strategies. Field experiments have shown that spelt wheat achieves higher selenium concentrations in the grain, which correlates with morphological characteristics such as greater plant height, indicating different accumulation capacities compared to common wheat [40]. In addition, sulphur-selenium application increased the copper and zinc content mainly in spelt straw, confirming its greater susceptibility to the synergistic effect of micronutrients [41]. In turn, common wheat reacts more strongly to the availability of protein nitrogen, especially when interacting with selenium and sulphur, as confirmed by data showing a significant increase in mineral nitrogen in straw after fertilisation [37]. Additionally, genetic studies highlight differences between species—spelt shows better selenium accumulation potential in traditional varieties, which is a promising target for selection and breeding focused on nutritional value [40]. The conclusion is that metabolic and adaptive differences between spelt and common wheat should be taken into account when designing fertilisation and biofortification programmes, which will enable optimised growth support, stimulation of the accumulation of valuable components and increased nutritional value of crops.
Recent studies indicate the growing importance of selenium and sulphur in agriculture, both in the context of biofortification and improving plant resistance to environmental stress. The use of selenium in appropriate doses and at appropriate times increases its accumulation in cereal grains while minimising the risk of toxicity [36]. Increasing attention is being paid to innovative forms of application, such as selenium nanoparticles, which have multidirectional effects, including improved antioxidant properties and reduced heavy metal bioaccumulation [42,43]. At the same time, research on molecular mechanisms confirms the role of selenium in regulating secondary metabolism and alleviating metal stress [38,39]. In the case of sulphur, its presence in the plant growth environment supports photosynthesis, sulphur amino acid synthesis and improves crop quality, which, in combination with selenium, creates synergistic biofortification strategies [44]. In the face of global selenium and sulphur deficiencies in soils, the integration of fertilisation with these elements appears to be a key element of sustainable agriculture, supporting food security and human health.
The research hypothesis was that varying doses of selenium and sulphur and the timing of the application of the former would have a beneficial effect on the Se and S various forms (total-S, sulphate S and organic S) content in the grain and straw of spelt wheat and common wheat. With this in mind, studies were conducted to determine the effect of selenium and the date of its application and sulphur on the content of selenium and various forms of sulphur (total sulphur, sulphate sulphur and organic sulphur) and the N:S ratio in winter spelt wheat and winter common wheat. The analysis of different sulphur fractions—total, sulphate and organic—is crucial for understanding the role of this element in plant metabolism. Sulphates are a form available to plants and are the main substrate for the synthesis of sulphur amino acids, such as cysteine and methionine, which affect protein metabolism and immune mechanisms [45]. The permanent organic form of sulphur, bound to organic matter, acts as a buffer and constitutes a susceptibility reserve, gradually mineralised by microorganisms, which helps maintain the balance of sulphates in the soil [46]. Studies also confirm that the sulphur content in the soil affects microbial diversity and activity, which determines the dynamics of transformations and the availability of sulphur to plants [47]. As a result, a comprehensive analysis of sulphur fractions enables precise fertiliser dosing and optimisation of growth conditions, which translates into an effective and sustainable fertilisation strategy.
2. Results and Discussion
2.1. Selenium of Wheat
The selenium content in spelt and common wheat grains (Table 1a,b) was consistent with the data in the literature [34,48].
However, in most cases, the control plants (without selenium fertilisation) had a lower content than 0.1 mg kg^−1^, which is considered optimal for nutritional purposes [40]. The selenium content in cereal grains was also studied by Wang et al. [49], who showed that the average content of this element was below 0.1 mg kg^−1^, while the average selenium content in spring wheat grain was 0.066 mg kg^−1^. In Slovakia, the average selenium content in wheat was even lower, at 0.029 mg kg^−1^ d.m. [50].
Selenium fertilisation resulted in an average 4-fold (lower dose) and more than 5-fold (higher dose) increase in its content in common wheat grain, as well as in a 3-fold and in a 4-fold increase in Se content in spelt wheat grain, compared to objects without selenium in the fertiliser dose (Table 1b). Selenium fertilisation at a later date resulted in a twofold increase in Se content in common wheat and spelt wheat grain. Sulphur fertilisation reduced the selenium content of common wheat grain by an average of 17% and 25%, respectively.
The selenium content in spelt and common wheat straw (Table 2a,b) was very low. No significant changes in its content were observed under the influence of the experimental factors used.
In studies by Manojlović et al. [51], selenium fertilisation was associated with a two- to five-fold increase in its content in winter wheat grain. Foliar application of selenium also resulted in a significant increase in its content in maize and bean seeds [52]. Ramkissoon et al. [18] and Wang et al. [53] report an increase in selenium concentration in wheat grain to 0.350 mg kg^−1^ after applying a dose of 30 g ha^−1^. Radawiec et al. [48] and Ducsay et al. [50,54] reported an increase in selenium content to 0.445 mg kg^−1^ after applying a dose of 10 g ha^−1^ of selenium. Wang et al. [55] found that foliar fertilisation with sodium selenite (IV) at a dose of 6 and 12 g Se ha^−1^ is associated with an increase in Se content in winter wheat grain from 0.042 to 0.067 mg kg^−1^ and from 0.065 to 0.180 mg kg^−1^ d.m., respectively. Research by Klikocka et al. [56] and Yeasmin et al. [21] reports that fertilisation with sulphur at a dose of 50 kg ha^−1^ results in an increase in selenium content in spring wheat grain.
It is believed that selenium doses ranging from 10 to 20 g ha^−1^ Se allow biofortification goals to be achieved [50,51,52,57]. Studies evaluating the effect of Se application at doses ranging from 0 to 25 g ha^−1^ showed an increase in Se content in plants [58,59]. In our own studies, doses consistent with those mentioned above were used, but they did not result in an excessive increase in selenium content in wheat grains compared to the above studies. The target desired range of selenium content in cereal grains after biofortification, at which there is no risk of toxic effects, is 0.1 to 1.0 mg kg^−1^ [60,61,62]. The studies conducted allowed us to obtain selenium content in grain within the above range.
Furthermore, when comparing the results of our own research to studies reporting high selenium supply without toxic effects and at the same time ensuring the desired selenium content in the blood [63], they are similar. The highest selenium content in spelt wheat grain, 0.38 mg kg^−1^ Se, was found for the total combination of the highest sulphur dose and the highest selenium dose in spelt wheat grain applied on the second date, while in common wheat, the highest content was 0.30 mg kg^−1^ and was found in the combination of the highest dose of selenium applied in the second period without sulphur fertilisation. The results obtained demonstrate the effective implementation of agrotechnical biofortification aimed at increasing the selenium content in the grain of test plants, but without toxic effects. In straw, however, no positive effect of selenium fertilisation on its content was observed, therefore straw should not be considered a source of selenium for the biofortification of animals. The research conducted may help fertiliser manufacturers select the best fertilisers and Se levels to ensure the safe inclusion of Se in the food chain in the future, thereby improving Se intake by humans and animals [64].
According to Ducsay et al. [50], the best time to introduce selenium into wheat is during the tillering (BBCH 20–29) and stem elongation (BBCH 30–39) stages. Zhang et al. [36]. According to their research, the best method of fertilising wheat to enrich it with selenium is foliar or soil fertilisation around the BBCH 31 growth stage. Our own research is consistent with these theories, as in both test plants, higher selenium content was found in plants fertilised with Se during the stem elongation stage (BBCH 31–34).
2.2. Total Sulphur of Wheat
Wheat is classified as a plant with relatively low sulphur requirements, but a deficiency of this element can result in poorer nitrogen management and reduced grain quality [65,66]. The literature contains reports of no difference in sulphur content between grain and straw [67], higher total sulphur content in straw than in grain [68], and those in which the sulphur content in straw is lower than in grain [69]. The results of our own research on the sulphur content in both spelt and common wheat grain and straw [Table 3a,b] are consistent with the research of other authors [34].
The application of a lower dose of selenium resulted in an average 7% increase in total sulphur content, while a higher dose resulted in a 10% decrease in spelt wheat grain [Table 3b]. The application of both lower and higher doses of selenium to the common wheat growth environment was associated with an increase in total sulphur content in the grain by an average of 16–17%. Later selenium fertilisation resulted in an 8% decrease in total sulphur content in spelt wheat grain. The total sulphur content in spelt wheat grain increased by an average of 5% as a result of applying a lower dose of sulphur to the plant growth environment. An increase in the total sulphur content in common wheat grain, compared to control objects without sulphur in fertilisers, was observed in plants fertilised with a lower dose of sulphur.
The effect of applying selenium and sulphur on the total sulphur content of wheat straw is illustrated in Table 4a,b. Selenium fertilisation in most cases resulted in a decrease in total sulphur content in spelt wheat straw by an average of 5% and 15%, respectively [Table 4b]. The application of selenium at a lower dose was associated with a 24% decrease in total sulphur content in common wheat straw, and at a higher dose with a 20% increase. Fertilisation with a higher dose of sulphur resulted in an increase in total sulphur content in spelt wheat straw by an average of 17% and in common wheat straw by 26%.
Studies by Hrivna et al. [68] and Liu et al. [66] confirm an increase in total sulphur content in winter wheat grain as a result of sulphur fertilisation. In studies by Rodrigo et al. [70], the total sulphur content in barley did not change under the influence of selenium fertilisation, even at a dose of 500 g Se ha^−1^. Similarly, in the studies by Yeasmin et al. [21], the content of the elements studied, including sulphur, was not sensitive to selenium fertilisation. However, in the studies by Silva et al. [71] and Saeed et al. [33], the sulphur content decreased in the test plants under the influence of selenium fertilisation.
2.3. Sulphate Sulphur of Wheat
The sulphate sulphur content in both the grain (Table 5a,b) and straw (Table 6a,b) of spelt wheat and common wheat was at the level most commonly found in the literature [58,72].
Fertilisation with selenium at a lower dose was associated with an average 6% increase in sulphate sulphur content only in common wheat grain (Table 5b). Fertilisation with a lower sulphur dose resulted in an average 26% increase in sulphate sulphur content, and a higher dose resulted in a 56% increase in spelt wheat grain. Sulphur fertilisation was associated with a significant reduction in the sulphate form of sulphur in common wheat grain by 19% and 13%, respectively.
The application of selenium to the growth environment of common wheat was associated with a decrease in sulphate sulphur content by an average of 21% and 11% in straw, respectively (Table 6b). Sulphur fertilisation resulted in an increase in sulphate sulphur content in spelt and common wheat straw in most cases. The application of a higher dose of S was associated with an average increase of 29% in spelt wheat straw and 18% in common wheat straw.
According to Schulte and Kelling [73], the sulphur content in plants is significantly influenced by the abundance of this element in the soil. Research by Pokhrel et al. [74] confirms an increase in sulphate sulphur content in rapeseed under the influence of sulphur fertilisation. Gil-Díaz et al. [75] noted this relationship in spring barley biomass. Studies by Qiu et al. [76] and Lima Gomes et al. [77] report an increase in sulphate sulphur content in the plants studied—in barley and tomatoes, respectively—under the influence of selenium fertilisation.
2.4. Organic Sulphur of Wheat
Organic Sulphur content of Wheat was presented in Table 7a,b (grain) and 8a,b (straw). Selenium fertilisation was associated with a significant increase in organic sulphur content of 11% was observed in spelt wheat grain under the influence of fertilisation with a lower dose (Table 7b). Selenium fertilisation caused an increase in the organic sulphur content in common wheat grain, with the difference that this reaction was observed after the application of both doses (an average increase of 23% and 33%, respectively). The second application of selenium was associated with a 10% decrease in the organic sulphur content of spelt wheat grain. An average 14% decrease in organic sulphur content in spelt wheat grain was observed after applying a higher dose of sulphur, compared to plots without sulphur in fertilisers. Fertilisation with a lower dose of sulphur resulted in a significant and clear increase in organic sulphur content (by 25%) in common wheat grain.
A significant increase in organic sulphur content in spelt wheat grain under the influence of a lower selenium dose may be related to the induction of sulphur metabolic pathways by selenium, due to their chemical similarity [5,78]. In the case of common wheat, an increase in organic sulphur content was observed after the application of both selenium doses, indicating a stronger interaction between these elements in this species [21]. The reduction in organic sulphur content in spelt wheat grain associated with the delay in selenium application may be due to a shift in the phase of activity of the enzymes responsible for the synthesis of sulphur amino acids [64].
The lack of increase in organic sulphur content in spelt wheat grain under the influence of higher sulphur fertilisation is consistent with observations that excessive sulphur doses may not bring the expected effects in terms of protein quality improvement and may even lead to a decrease in the content of sulphur compounds in the grain [68,79]. This may be due to disturbances in the metabolism of sulphur amino acids when there is an excess of this element [66]. On the other hand, the effect of a lower sulphur dose, manifested by a significant increase in the organic sulphur content in common wheat grain, confirms the importance of optimising fertiliser doses in order to avoid antagonism [65].
A higher dose of sulphur resulted in an average 14% increase, and a higher dose of selenium resulted in a 23% decrease in the organic sulphur content of spelt wheat straw (Table 8b). Sulphur fertilisation caused an average 29% and 57% increase in the organic sulphur content of common wheat straw, respectively. Fertilisation with a higher dose of selenium and a delay in its application resulted in an increase in the organic sulphur content of common wheat straw.
In spelt wheat straw, a higher dose of sulphur resulted in an increase in organic sulphur content, while a higher dose of selenium resulted in a decrease. This confirms the hypothesis of competition between these elements in transport pathways [21,34]. In common wheat, sulphur fertilisation caused a marked increase in the organic sulphur content of straw, and delaying the application of selenium further increased this effect, which may be related to a longer period of accumulation of sulphur compounds in vegetative tissues [34].
2.5. N:S Ratio of Wheat
The N:S ratio (total nitrogen content to total sulphur content) in the generative organs of crops is an indicator that is very often used to diagnose sulphur deficiencies in plants [80,81]. The accumulation of non-protein forms of nitrogen in plants (amides, amino acids and nitrates) is promoted by an increase in the N:S ratio. On the other hand, a decrease in this ratio leads to the accumulation of inorganic sulphur compounds [82]. Various theories regarding the optimal value of this ratio can be found in the literature. Sedlár et al. [83] report that in climatic conditions, wheat with the best technological value is obtained with an N:S ratio ranging from 9.5–10:1 to 17:1. The highest cereal yields are obtained when the ratio is 15:1 [84], while the critical value of the ratio is 17:1 [85]. In our own research, the values of the discussed indicator were in most cases within the given ranges (Table 9a,b and Table 10a,b).
Fertilisation with a lower dose of selenium resulted in an average 9% reduction in the C:N ratio, while a higher dose resulted in a 6% increase in spelt wheat grain (Table 9b). Selenium fertilisation was associated with a narrowing of the N:S ratio by 13% and 16%, respectively, in common wheat grain. A higher average N:S ratio in spelt wheat grain was obtained in plots fertilised with selenium on the second application date. In spelt wheat grain, fertilisation with a higher dose of sulphur led to an average 5% reduction in the C:N ratio in most cases.
The application of selenium resulted in an increase in the N:S ratio in spelt wheat straw by an average of 6 and 10%, respectively (Table 10b). Fertilisation with selenium and timing of its application did not clearly differentiate the N:S ratio values in common wheat straw. Fertilisation with a higher dose of sulphur resulted in a 10% reduction in the N:S ratio in spelt wheat straw. The presence of both lower and higher doses of sulphur in the plant growth environment was associated with a reduction in the N:S ratio by an average of 11% and 31%, respectively, in common wheat straw.
The results of our own research are consistent with the studies by Roa et al. [65] and Klikocka et al. [86], who report a narrowing of the N:S ratio as a result of the use of sulphur in fertilisation, which results in a more favourable nutritional value of plants. Arrigoni et al. [87], on the other hand, obtained the highest value of this parameter in wheat grain fertilised with nitrogen and sulphur. Despite the fact that wheat is not a plant with a high sulphur requirement, its deficiency can lead to reduced nitrogen utilisation and a narrowing of the N:S ratio [80]. The experiment by Ullah et al. [88], which examined different types of sulphur fertilisers, proved that elemental sulphur had the least favourable effect on the N:S ratio, while ammonium sulphate had the most positive effect. Chien et al. [89], Pimentel et al. [90] and LaBarge [72] also found a positive effect of ammonium sulphate on the total sulphur content in the tested plants, as well as on the N:S ratio.
The yield and quality of wheat useful parts, including the content of selenium and sulphur compounds, depend on optimal doses of mineral fertilisers [91,92], which should also be used in quantities that do not affect the environment [93]. The results of our own 3-year field research presented above, as well as the scientific discussion, show that the application of selenium and sulphur has a beneficial effect on the chemical composition of spelt wheat and common wheat. It seems necessary to conduct research with other plant species, adjusting doses to their nutritional and fertiliser requirements.
In summary, it can be concluded that the interactions between selenium and sulphur result from their chemical and biochemical similarities—both elements belong to the same group of the periodic table and use similar transport and assimilation pathways in plants. Selenium in the form of selenite (SeO_3_^2−^) is taken up by plants mainly through sulphate transporters, leading to enzymatic competition at the reduction stage in chloroplasts [38]. After entering the cell, selenium undergoes a reduction pathway analogous to sulphur, participating in the biosynthesis of sulphur amino acids such as selenocysteine and selenomethionine [45]. This competition explains why an increased supply of sulphur can limit the accumulation of selenium in grain, while not significantly affecting its content in straw—intensive protein synthesis occurs in grain, requiring sulphur amino acids, which promotes the incorporation of selenium into organic structures.
The different reactions of spelt and common wheat result from different mechanisms of nitrogen and sulphur metabolism regulation and differences in the expression of sulphate transporters and reductase enzymes [40]. Common wheat, characterised by higher protein synthesis intensity in the grain, has a greater ability to incorporate selenium into organic forms, which explains the higher increase in its concentration in the grain compared to spelt. In straw, on the other hand, structural rather than metabolic processes dominate, which is why Se–S interactions are less pronounced. The developmental phase is also of key importance—the application of selenium in the stem elongation phase (BBCH 31–34) coincides with the intense activity of sulphate-reducing enzymes, which promotes competition and the incorporation of selenium into organic metabolites [39].
Discrepancies in the observed effects of selenium and sulphur fertilisation may result from several biological and environmental factors. Firstly, the lack of clear changes in the sulphur fraction content when selenium is applied may be related to differences in uptake mechanisms—sulphates are actively taken up by specialised transporters, while selenite only partially competes with them, which limits the impact of selenium on the sulphur pool in the plant [45]. Secondly, the observed differences between grain and straw may result from the preferential targeting of sulphur amino acids and their selenium analogues to generative organs, where intensive protein synthesis determines greater selenium incorporation [38]. Structural processes dominate in straw, which are less dependent on sulphur and selenium metabolism, which explains the lack of clear interactions. The differences between spelt and common wheat may, in turn, be the result of different expression of sulphate transporters and reductase enzymes, as well as differences in the rate of translocation of components to the grain [40]. Alternatively, the effect of the timing of selenium application may be modulated by environmental conditions (humidity, temperature) that determine the activity of selenite and sulphate reducing enzymes, leading to variable biofortification efficiency [39].
3. Materials and Methods
3.1. Research Methodology
This research was conducted on the basis of a three-year field experiment carried out at the Experimental Station of the University of Life Sciences in Lublin, located in Czesławice (51°18′23′′ N, 22°16′02′′ E), Poland. The research was conducted between 2015 and 2018. The research was conducted on plots located in the same place on soil with the same properties. The experiment was set up on clay soil according to the IUSS Working Group WRB [94] classification, formed from loess deposits. The chemical composition of the soil was included in the first, already published article on this experiment [34]. The tested soil was characterised by a slightly acidic reaction (pH_KCl_ - 6.3) and a moderate mineral nitrogen content of 0.83 mg NH_4_-N kg^−1^ d.m. and 22.39 mg NO_3_-N kg^−1^ d.m. It was distinguished by a high concentration of phosphorus (105.5 mg P kg^−1^ d.m.) and magnesium (59.0 mg kg^−1^ d.m.), while the sulphur content in the form of sulphate was average and amounted to 10.4 mg SO_4_ kg^−1^ d.m. A similar average level was recorded for potassium, which amounted to 134.5 mg K kg^−1^ d.m. These properties indicate a soil rich in macroelements, with a simultaneous need to monitor the availability of sulphur in the context of plant biofortification. Two model plant species were used in the study: winter spelt wheat (Triticum spelta L.) Rokosz variety (Hodowla Roślin Strzelce Sp. z o.o., Strzelce, Poland) and winter common wheat (Triticum aestivum L.) Astoria variety (Poznańska Hodowla Roślin, Tulce, Poland).
The experiment was designed as a three-factor system in a factorial design. The first factor included three levels of sulphur application: S_0_—0 kg S ha^−1^, S_1_—15 kg S ha^−1^ and S_2_—30 kg S ha^−1^. The second factor consisted of three levels of selenium supplementation: Se_0_—0 g Se ha^−1^, Se_1_—10 g Se ha^−1^ and Se_2_—20 g Se ha^−1^. The third factor referred to two selenium application dates, i.e., in the tillering phase (BBCH 22–24) and in the stem elongation phase (BBCH 31–34). Factors S and Se were introduced at three levels, while the application date was introduced at two levels, which enabled the analysis of interactions between the studied variables. The field experiment design is shown in Figure 1.
The plants were sown in the first ten days of October and harvested in the second ten days of August, at the stage of full physiological maturity. Sulphur was applied before sowing in the form of ammonium sulphate ((NH_4_)2_SO_4), while selenium was applied in the form of sodium selenite (IV) (Na_2_SeO_3_). Sodium selenite (IV) [Na_2_SeO_3_] is preferred in biofortification due to its high solubility, stability in the soil environment and easy reduction to organic forms in plants, which promotes the synthesis of selenomethionine and selenocysteine [38]. Ammonium sulphate [(NH_4_)2_SO_4] provides sulphur in the form of sulphate, which is a direct substrate for the biosynthesis of sulphur amino acids, while also introducing ammonium nitrogen, supporting protein metabolism [47]. Both forms are consistent with the principles of green chemistry—they are characterised by high efficiency of ingredient use, low mobility in the environment and minimal risk of ecotoxicity when dosed correctly [46]. Nitrogen was applied at an initial dose of 20 kg N ha^−1^ before sowing in the form of ammonium nitrate (NH_4_NO_3_), and then supplemented in spring to a total dose of 80 kg N ha^−1^: 40 kg N ha^−1^ in the tillering phase (BBCH 22–24), taking into account nitrogen from ammonium sulphate, and 20 kg N ha^−1^ in the stem elongation phase (BBCH 31–34). Phosphorus and potassium were applied before sowing in amounts of 60 kg P_2_O_5_ ha^−1^ and 80 kg K_2_O ha^−1^, respectively. Meteorological data and used pesticides during the growing season were characterised in a previously published article on this experiment [84].
After harvesting, representative samples of plant material (grain and straw) were taken for laboratory analysis in accordance with the applicable analytical procedures. Yield of plants and content of total, mineral and organic nitrogen are given in a previously published papers [34,37].
3.2. Analytical and Statistical Methods
During harvesting, plant material covering an area of 0.25 m^2^ was collected from each experimental unit for laboratory analysis. After separating the grain and straw fractions, the samples were dried at 60 °C until a constant weight was achieved, and then ground in an electric grinder to obtain a homogeneous homogenate. The resulting material was used as a substrate for determining the content of selected elements. The total sulphur content (S_tot_) was determined by the nephelometric method [95], after prior digestion of the samples with fuming nitric acid (V) (HNO_3_). The sulphate sulphur (SO_4_^2−^-S) content was also determined nephelometrically, after extraction of plant material with a 2% acetic acid (CH_3_COOH) solution, according to the methodology of Kurmanbayeva et al. [96]. Additionally, the crushed samples (both grain and straw) were digested in concentrated nitric acid (V), and the selenium (Se) content was determined in the resulting solution using inductively coupled plasma mass spectrometry (ICP-MS) [97], which ensures high sensitivity and precision of elemental analysis. In ICP-MS analysis, the determination range for selenium was 0.005–5 mg·L^−1^, with a limit of detection (LOD) of 0.001 mg·L^−1^. The limits of quantification (LOQ) correspond to the lower limits of the calibration ranges presented above. To ensure the reliability of the results, certified reference materials NCS ZC 73030 (Chinese National Analysis Centre for Iron and Steel, Beijing, China), quality control through blanks, internal standards, multi-point calibration and sensitivity drift checks using control samples were used. In colorimetric and nephelometric methods, calibration curves based on 5–7 points with linearity assessment (R^2^ ≥ 0.99) were prepared, and 3 analytical replicates were performed for each sample to assess precision. Such procedures ensure compliance with validation requirements and minimise the risk of systematic and random errors.
The collected empirical data were subjected to statistical analysis using three-factor analysis of variance (ANOVA), performed in the Statistica computing environment [98,99]. In order to identify significant differences between the mean values of the studied characteristics, Tukey’s multiple post hoc comparison test (HSD) was used, with a significance level of p = 0.05 [98]. Based on the test results, the least significant difference (LSD) was determined and statistically homogeneous groups of objects were identified [99]. The values presented in the tables for the interactions of factors A × B × C, which are marked with the same letter, do not show any significant statistical differences. This confirms that they belong to the same homogeneous group within used comparative test. The consistency of the letter designations indicates that the hypothesis of equality of means in the analysed factor combinations is not rejected, in accordance with the criteria adopted in the analysis of variance and post-hoc procedures (Tukey’s test), which allows them to be interpreted as statistically equivalent in the context of the studied feature. The use of the LSD (Least Significant Difference) test in parallel with Tukey’s HSD test stems from the need to balance sensitivity and Type I error control. The LSD test is more liberal and allows for the detection of subtle differences between means, which is useful in preliminary analysis, while Tukey HSD is more conservative and controls for cumulative error in multiple comparisons, ensuring greater reliability of results. In research practice, LSD can indicate potential differences, and Tukey HSD can confirm them, which is particularly important in agronomic studies with multiple factor combinations [100]. Data from individual years were accepted for statistical calculations as repetitions, i.e., statistical calculations were performed for the averages from the years.
4. Conclusions
Selenium fertilisation significantly increased the content of this element in spelt wheat and common wheat grains—fourfold and fivefold, respectively, compared to the control. This effect was further modulated by the timing of selenium application to the plant growth environment, indicating the importance of the developmental stage in the biofortification process. On the other hand, the selenium dose and the timing of its application did not show a clear effect on the content of both analysed forms of sulphur, which suggests the complexity of the interaction between these elements. It is important to note that the fertilisation levels did not lead to selenium content considered toxic to humans and animals, confirming the safety of the proposed strategy.
The presence of sulphur in the growing environment of spelt wheat and common wheat was associated with an increase in the total sulphur and sulphate sulphur content in the grain, and especially in the straw. Fertilisation with a higher dose of sulphur increased the total sulphur, sulphate sulphur and organic sulphur content in spelt wheat straw by an average of 17%, 29% and 23%, respectively, and in common wheat straw by 26%, 18% and 57%, respectively. At the same time, the presence of sulphur in the plant growth environment did not affect the selenium content in the grain, which indicates a lack of direct competition in the mechanisms of uptake of these elements under the tested conditions. The results suggest that the optimal dose of selenium for biofortification purposes is 20 mg Se ha^−1^ on clay soil, applied during the stem elongation stage (BBCH 31–34).
Biofortification of wheat through integrated selenium and sulphur fertilisation is an effective agronomic strategy aimed at increasing the selenium content in grain, which in turn may contribute to reducing selenium deficiencies in the human diet. This method is in line with the concept of improving food quality at source, enabling the supply of selenium in an organic form, characterised by high bioavailability and safety for the consumer. In the face of global selenium deficits resulting from its low concentration in the soils of many regions of the world, cereal biofortification appears to be a solution of strategic nutritional and health importance, supporting the prevention of diseases associated with selenium deficiency.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Malagoli M. Schiavon M. Dall’Acqua S. Pilon-Smits E.A.H. Effects of Selenium Biofortification on Crop Nutritional Quality Front. Plant Sci.2015628010.3389/fpls.2015.0028025954299 PMC 4404738 · doi ↗ · pubmed ↗
- 2Zhou X. Yang J. Kronzucker H.J. Shi W. Selenium Biofortification and Interaction with Other Elements in Plants: A Review Front. Plant Sci.20201158642110.3389/fpls.2020.58642133224171 PMC 7674621 · doi ↗ · pubmed ↗
- 3Guo Q. Ye J. Zeng J. Chen L. Korpelainen H. Li C. Selenium Species Transforming along Soil–Plant Continuum and Their Beneficial Roles for Horticultural Crops Hortic. Res.202310 uhac 27010.1093/hr/uhac 27036789256 PMC 9923214 · doi ↗ · pubmed ↗
- 4Liao Q. Xing Y. Li A.-M. Liang P.-X. Jiang Z.-P. Liu Y.-X. Huang D.-L. Enhancing Selenium Biofortification: Strategies for Improving Soil-to-Plant Transfer Chem. Biol. Technol. Agric.20241114810.1186/s 40538-024-00672-z · doi ↗
- 5Wang M. Zhou F. Cheng N. Chen P. Ma Y. Zhai H. Qi M. Liu N. Liu Y. Meng L. Soil and Foliar Selenium Application: Impact on Accumulation, Speciation, and Bioaccessibility of Selenium in Wheat (Triticum aestivum L.)Front. Plant Sci.20221398862710.3389/fpls.2022.98862736186067 PMC 9516304 · doi ↗ · pubmed ↗
- 6Pei W. Dai M. Shi S. Zhang Y. Wu D. Qiao C. Sun Y. Wang J. Effects of Foliar Selenium Spraying on the Growth and Selenium Content and Morphology of Rice Front. Plant Sci.202516158715910.3389/fpls.2025.158715940353239 PMC 12061967 · doi ↗ · pubmed ↗
- 7Deng G. Fan Z. Wang Z. Peng M. Dynamic Role of Selenium in Soil–Plant–Microbe Systems: Mechanisms, Biofortification, and Environmental Remediation Plant Soil 20255151085110510.1007/s 11104-025-07661-7 · doi ↗
- 8Jiang Z. Wang Z. Zhao Y. Peng M. Unveiling the Vital Role of Soil Microorganisms in Selenium Cycling: A Review Front. Microbiol.202415144853910.3389/fmicb.2024.144853939323878 PMC 11422209 · doi ↗ · pubmed ↗