The Role of Vitamin D in Autoimmune Diseases
Federica Vincenzi, Carlo Smirne, Stelvio Tonello, Pier Paolo Sainaghi

TL;DR
This review explores how vitamin D influences immune responses and its potential role in managing autoimmune diseases like multiple sclerosis and rheumatoid arthritis.
Contribution
The paper provides a comprehensive overview of vitamin D's molecular mechanisms in immune regulation and its implications for autoimmune diseases.
Findings
Vitamin D deficiency is linked to the pathogenesis of autoimmune diseases such as multiple sclerosis and type 1 diabetes.
Vitamin D promotes tolerogenic immune responses and suppresses inflammation through its receptor interaction.
Supplementation may help reduce disease activity in immune-mediated disorders.
Abstract
Vitamin D is a steroid hormone whose relevant immunomodulatory role has been widely described. Therefore, its contribution to the pathogenesis of immune-mediated diseases is an important and ongoing matter of research. Specifically, the active form of vitamin D, i.e., 1,25-dihydroxyvitamin D, through the interaction with its receptor, exerts different activities on the innate and adaptive immune system, among which are suppression of inflammation and promotion of tolerogenic responses. Indeed, vitamin D insufficiency/deficiency has been related to the pathogenesis and/or disease activity of several autoimmune diseases, including, amongst others, multiple sclerosis, rheumatoid arthritis, systemic lupus erythematosus, and type 1 diabetes mellitus. Based on these premises, in this review, we will describe the main molecular mechanisms modulated by vitamin D in the regulation of immune…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
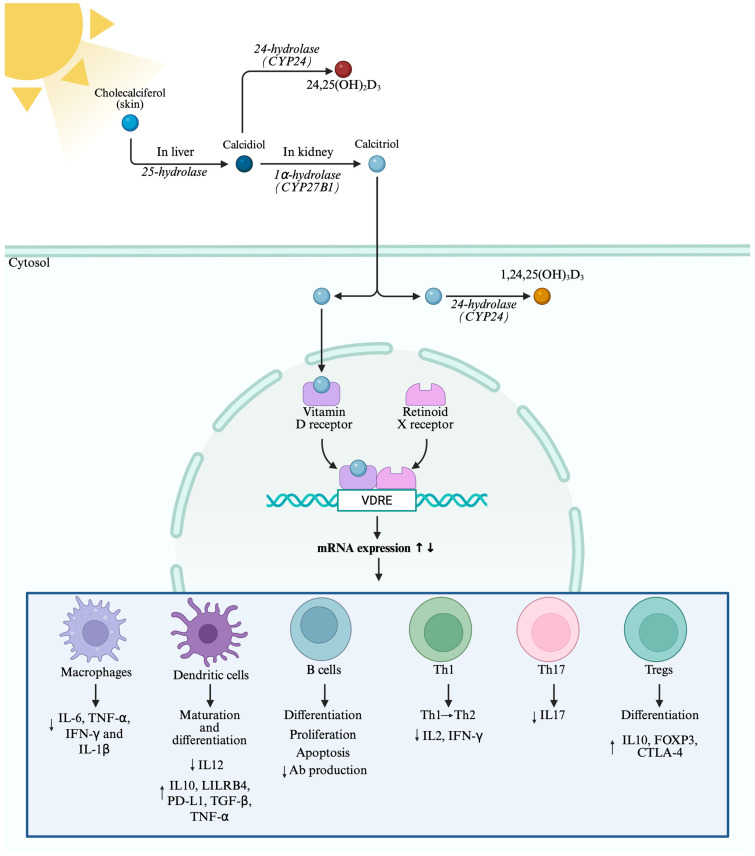 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVitamin D Research Studies · Thyroid Disorders and Treatments · Diabetes and associated disorders
1. Introduction
Vitamin D is a fat-soluble vitamin mainly involved in bone remodeling, through the regulation of calcium reabsorption from bone and intestine. Due to its structure, vitamin D acts as a hormone, influencing cellular differentiation and proliferation, immune system regulation, neural function, and cardiovascular health [1]. Despite its classical function, increasing evidence has been gathered on the immunomodulatory effects of vitamin D on both innate and adaptive immune systems. This knowledge arises from studies performed in 1903 by Dr. Nils Filsen, who cured the epidermal form of tuberculosis (lupus vulgaris) using light irradiation. The following findings regarding the synthesis of vitamin D in the skin after ultraviolet (UV) light exposure opened a window to the possible immunomodulatory role of vitamin D, particularly in the treatment of mycobacterial infections, such as tuberculosis and leprosy [2]. Further studies showed that both vitamin D receptor (VDR) and metabolizing enzymes are expressed in a plethora of immune cells, including dendritic cells, lymphocytes, monocytes, and macrophages. These findings highlight the capacity of immune cells to synthetize vitamin D, producing both VDR-mediated paracrine or intracrine responses [2]. In light of the current knowledge about vitamin D’s immunomodulatory role, researchers focused on the possible contribution of vitamin D in autoimmune diseases and immune-related disorders. Indeed, the association between vitamin D deficiency and an increased risk of autoimmune diseases has been described [3]. Several immune-mediated diseases have been the topic of research, including rheumatoid arthritis (RA), systemic lupus erythematosus (SLE), multiple sclerosis (MS), and type 1 diabetes mellitus (T1D), which will be the focus of this review. However, it is essential to give a brief overview of the relationship between vitamin D and less studied diseases including psoriasis, inflammatory bowel disease (IBD), autoimmune thyroiditis, and celiac disease.
Low levels of vitamin D have been reported in patients with psoriasis, a known chronic inflammatory autoimmune disease affecting the skin [4]. Moreover, due to their anti-inflammatory properties, vitamin D and derivatives seem to be effective adjuvant treatments for psoriasis, both orally and topically administered [5]. Another immune-mediated condition is IBD, characterized by chronic intestinal inflammation. This persistent inflammation leads to malabsorption, which can contribute to increased risk of vitamin D deficiency. Moreover, vitamin D/VDR signaling can modulate intestinal epithelium permeability, the release of antimicrobial peptides, and mucus secretion. Therefore, its supplementation may help restore mucosal integrity and reduce inflammation [6]. Celiac disease is another autoimmune intestinal disorder triggered by gluten intake, resulting in chronic inflammation and villous atrophy [7]. In celiac disease, intestinal barrier integrity is altered, and, as for IBD, vitamin D deficiency may be caused by malabsorption. Even if vitamin D may help reduce inflammation, no conclusive results have been gathered regarding the effects of its supplementation in the context of celiac disease [8]. An association between celiac disease and autoimmune thyroiditis, including Hashimoto’s thyroiditis and Graves’ disease, has been reported. This could be due to the diversity of the gut microbiome and nutrient deficiency, including vitamin D, which plays an important role in regulating gut, thyroid, and immune system function [7]. Interestingly, vitamin D dampens excessive inflammation and T cell activation; therefore, it may contribute to reducing tissue damage in autoimmune thyroiditis [9].
In our review, we will explore the mechanisms by which vitamin D regulates both the innate and adaptive immune systems and the relationship between vitamin D and immune-related disorders, focusing on RA, MS, SLE, and T1D.
2. Vitamin D Metabolism and Function
Despite its name, vitamin D is a hormone precursor which can be endogenously synthetized after UV-B exposure or can be introduced with the diet. Vitamin D encompasses vitamin D_2_ (ergocalciferol), derived from plants, and vitamin D_3_ (cholecalciferol), which is synthetized in animals [10]. Upon UV-B exposure, the 7-dehydrocholesterol in the skin is photoconverted into cholecalciferol, which binds to the Vitamin D Binding Protein (VDBP), and it is transported to the liver. Cholecalciferol is the biologically inactive form of vitamin D. Therefore, it is subjected to a series of modifications to convert it to calcitriol, also known as 1,25-dihydroxycholecalciferol, the active form of vitamin D, as shown in Figure 1. In the liver, a plethora of enzymes belonging to the cytochrome P450 (CYP) class, including CYP27A1 and CYP2R1, hydroxylate cholecalciferol in position C25, producing calcidiol, also known as 25-hydroxycholecalciferol [11]. This metabolite is the most abundant form in the bloodstream, and, due to its long half-life, it is the best indicator of vitamin D status. Indicatively, serum levels of calcidiol should be greater than 30 ng/mL (75 nmol/L) but not exceeding 150 ng/mL (374 nmol/L). Moreover, levels between 21–29 ng/mL (52.5–72.5 nmol/L) indicate vitamin D insufficiency, while deficiency can be suggested with calcidiol levels lower than 20 ng/mL (50 nmol/L) [12,13]. However, no definitive consensus has been reached regarding the normal circulating levels of vitamin D.
Calcidiol is further converted in the proximal tubule of the kidney through the enzyme 1α-hydroxylase (CYP27B1) to calcitriol, the hormonally active form. Studies also reported the presence of CYP27B1 in extrarenal sites, including placenta, bone, and various immune cells, underlying their capacity to produce calcitriol for intracrine and paracrine effects [14]. Calcitriol is then transported using VDBP in the target organs, including kidneys, gut, and bone, where it can exert its genomic and non-genomic functions. Genomic functions are mediated by the binding of calcitriol to cytosolic VDR in the target cell. This interaction induces VDR phosphorylation and heterodimerization with retinoid-X-receptor (RXR). The calcitriol–VDR-RXR complex translocates into the nucleus and binds to vitamin D response elements (VDREs), inducing either activation or repression of genes regulating different biological activities. This process is also mediated by the interaction of VDR with transcriptional coactivators or corepressors [15,16]. Through VDR, calcitriol can activate a negative feedback loop, promoting the expression of the gene encoding for CYP24. This enzyme has 24-hydroxylase activity, responsible for the hydroxylation in position C24 of both calcidiol and calcitriol, producing inactive metabolites and preventing toxic accumulation [17]. Moreover, calcitriol can downregulate the expression of both the CYP27B1 gene in the kidney and of the parathyroid hormone (PTH) gene. This negative feedback decreases the synthesis of both calcitriol and PTH, which modulates calcium homeostasis and positively regulates CYP27B1 [18,19]. In addition to its genomic effects, calcitriol can also bind to membrane VDRs, activating a series of non-genomic pathways, including calcium and mitogen-activated protein kinase (MAPK) signaling [15]. These effects are mediated by VDR’s interaction with other molecules, including Inhibitor Of Nuclear Factor Kappa B Kinase Subunit Beta (IKKβ), a regulator of the nuclear factor kappa B (NF-κB) pathway, signal transducer and activator of transcription (STAT)1, c-Jun, and β-catenin. Through these pathways, calcitriol can modulate the innate and adaptive immune system and antiviral response as well as cell survival [20]. Currently, it is known that VDR is widely expressed in several tissues, including prostate, breast, brain, colon, and immune cells. Referring to the latter, VDR has been found to be highly expressed in T lymphocytes, dendritic cells, monocytes, and macrophages, underlying the variety of physiological actions potentially exerted by this molecule [21].
In the following sections, we will describe the effects of vitamin D on innate and adaptive immunity. It is essential to set out that in the existing literature, the term “vitamin D” is frequently used without explicit specification of the molecular form administered. For clarity and consistency, in this review, the term “vitamin D” will be used to denote studies in which the specific vitamin D metabolite or analogue is not explicitly stated, unless otherwise indicated.
3. Vitamin D and Innate Immunity
The innate immune system serves as the body’s first line of defense, mounting a non-specific response against invading pathogens. It is characterized by complement activation and antibacterial response as well as antigen presentation to adaptive immune cells. In this context, several studies have highlighted the involvement of vitamin D in the regulation of the innate immune system, depicting its possible contribution to pathogen clearance as well as to the interplay between innate and adaptive immunity [22]. Figure 1 summarizes the effects of vitamin D in both innate and adaptive immune responses.
3.1. Macrophages
Monocytes and macrophages, key players in the innate immune system, are also targets of vitamin D, which modulates their antimicrobial and anti-inflammatory responses. These cells can recognize pathogen-associated molecular patterns (PAMPs) on the surface of pathogens, producing cytokines and exerting a phagocytic activity, contributing to the resolution of the infection through the interaction of PAMPs with toll-like receptors (TLRs) on monocytes and macrophages [23]. Upon TLR activation, a cascade of intracellular events leads to the synthesis of the active vitamin D metabolite, calcitriol, within macrophages. Indeed, studies performed on human macrophages treated with Mycobacterium tuberculosis have shown that TLR, mainly TLR1 and TLR2, interaction with PAMPs induces the activation of CYP27B1 and VDR, suggesting an intracrine synthesis of calcitriol and the subsequent signaling activated by the binding to VDR [24]. Therefore, particularly in granulomatous disorders such as tuberculosis, sarcoidosis, and some lymphomas, the excessive production of calcitriol by macrophages stimulates intestinal calcium absorption, leading to hypercalcemia and hypercalciuria [25]. This effect is exacerbated by the presence in macrophages of a non-functional variant of 24-hydroxylase, which prevents the negative regulation of vitamin D production [26].
Conversely to renal enzymes, which are regulated by PTH, the activation of vitamin D-metabolizing enzymes in extrarenal sites is mediated by the circulating levels of calcidiol, the most abundant form in the bloodstream. Indeed, CYP27B1 in monocyte/macrophages is regulated by both calcidiol and different types of cytokines, such as interferon (IFN)-γ, interleukin (IL)-1, and tumor necrosis factor (TNF)-α [26]. Experiments performed on peripheral blood mononuclear cells (PBMCs) of healthy individuals have shown that IFN-γ and lipopolysaccharide (LPS) stimulation enhance calcitriol synthesis, demonstrating that in vitro differentiation of immature monocytes in macrophages is associated with increased production of calcitriol [27]. Reinforcing these findings, Stoffels et al. established that the promoter of the CYP27B1 gene harbors binding sites for inflammation-responsive transcription factors, evidencing the contribution of inflammatory pathways to the enzyme’s regulation [28]. Furthermore, they established that the activity of 1α-hydroxylase is blocked by inhibitors of MAPK, Janus kinase (JAK), and NF-κB, suggesting the contribution of these pathways in the regulation of CYP27B1 activity and therefore in calcitriol synthesis [28].
Beyond its differentiating properties, vitamin D exerts potent anti-inflammatory effects, primarily by modulating NF-κB signaling and MAPK pathways. Studies on murine RAW 264.7 cells reported that calcitriol attenuates LPS-induced NF-κB gene enhancer B cells inhibitor alpha (IκBα) phosphorylation, leading to the inhibition of NF-κB. This results in the transcriptional downregulation of microRNA (miR)-155, a critical modulator of innate immunity, which in turn increases suppressor of cytokine signaling (SOCS) translation, thereby inhibiting LPS-mediated production of pro-inflammatory cytokines such as IL-6, TNF-α, IFN-γ, and IL-1β [29]. These findings have been confirmed by Cohen-Lahav et al., who reported that calcitriol can also increase the levels of IκBα, thus reducing the production of pro-inflammatory stimuli [30]. The synthesis of inflammatory cytokines following LPS stimulation is also mediated by the members of the MAPK family, comprising extracellular-signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and p38. In this regard, in vitro studies showed that pretreatment of human PBMCs with physiological concentrations of calcidiol, between 30–50 ng/mL, and of calcitriol, between 1–10 nM, inhibit LPS-induced p38 activation. Inhibition of p38 significantly decreases both IL-6 and TNF-α mRNA expression [31].
3.2. Dendritic Cell Modulation
Dendritic cells (DCs) are antigen-presenting cells responsible for the regulation of both innate and adaptive immune response. DCs are an heterogeneous population that can be divided into myeloid-derived (mDCs) and lymphoid-derived DCs (plasmacytoid, pDCs) according to the expression of cluster differentiation (CD)11c. mDCs share several features with monocytes, including the expression of CD33, CD13, and CD11c, along with low levels of CD123, and they are major producers of IL-12. In contrast, pDCs resemble plasma cells, characterized by high expression of CD4, CD62L, and CD123, and they mainly produce IFN-α.
Upon antigen exposure, mature DCs increase their antigen processing ability and display major histocompatibility complex (MHC)-I and MHC-II on their surface for antigen presentation as well as costimulatory molecules for T cell activation, including CD80 and CD86 [32,33]. Studies have identified similarity between DCs and macrophages in the expression pattern of CYP27B1 and VDR. However, mature DCs present elevated levels of CYP27B1 and reduced levels of VDR, while immature DCs highly express VDR. According to these findings, it can be suggested that calcitriol produced by mature DCs does not exert an intracrine effect but acts on immature DCs, allowing some DCs to mature but preventing an exaggerated response and possible pathological manifestations [34].
Indeed, it has been reported that calcitriol interferes with DC differentiation and maturation, in particular acting on mDCs, inducing a tolerogenic state (tDCs). This phenotype is characterized by expression of CD11c and downregulation of CD40, CD80, CD86, CD1a, and MHC-II [35]. An in vitro study of Penna et al. on human-derived DCs demonstrated that calcitriol, beyond inhibiting differentiation and maturation of immunogenic DCs, is very effective in reducing the secretion of IL-12, enhancing IL-10 production, and promoting DC apoptosis, thus resulting in T cell hyporesponsiveness [36]. Moreover, calcitriol upregulates the expression of T-cell-inhibitory molecules on tDCs, including leukocyte immunoglobulin-like receptor subfamily B member 4 (LILRB4) and Programmed Cell Death 1 Ligand (PD-L1), induces IL-10, transforming growth factor β (TGF-β), and TNF-α production, and downregulates NF-κB expression. These mechanisms result in decreased T cell activation and a shift in T cell polarization from a T helper (Th)1 and Th17 response to a regulatory T cells (Tregs) response [33]. In line with these affirmations, vitamin D may indirectly regulate the adaptive immune system, particularly T cell differentiation, through the modulation of DC differentiation, maturation, and apoptosis.
4. Vitamin D and Adaptive Immunity
Besides the indirect regulation of T cells mediated by DCs, the presence of VDR and CYP27B1 in B and T lymphocytes suggests that vitamin D can directly modulate these cells. Moreover, studies have reported that B and T cell activation leads to an increase in the expression of both VDR and CYP27B1. However, unlike the effects on innate immunity, vitamin D generally suppresses adaptive immunity [37].
4.1. B Cells
Only a limited number of studies have investigated the relationship between vitamin D and B cells. However, the expression of VDR- and vitamin D-metabolizing enzymes in B cells, like in other immune cells, has been reported [38]. Studies have shown that calcitriol modulates B cell production of IL-10. Heine et al. describe that the calcitriol–VDR interaction with VDREs in the promoter of IL-10 upregulates its expression in human CD40/IL-4-activated B cells [39]. Evidence suggests that vitamin D regulates B cell processes, including differentiation, proliferation, and apoptosis, and suppresses antibody production [40,41]. These effects may be relevant in the pathogenesis of inflammatory and autoimmune diseases.
4.2. T Cells
T cells can be subdivided into distinct functional subsets. CD4^+^ Th cells support B cell antibody (Ab) production and other cells in pathogen killing. CD4^+^ Th cells can differentiate into Th1 cells, mediators of response to viruses and intracellular infections, Th2 cells, which secrete IL-4, IL-5, and IL-13 and contribute to response to parasitic infections, B cell activation, and Ab production. Th17 cells act on extracellular pathogens and produce IL-17, Granulocyte–Macrophage Colony-Stimulating Factor (GM-CSF), and IL-22. Moreover, the presence of Th9, Th22, and T follicular helper cells has been reported. Another subset of T cells comprises CD8^+^ cytotoxic T cells, which are responsible for apoptosis of viral- or bacterial-infected cells. Persistent T cell activation and failure of regulatory control can then cause immune-mediated diseases. To avoid this uncontrolled T cell activation, Tregs, which are primary mediators of peripheral tolerance, can, in turn, exert a protective role by suppressing T cell responses [42].
Vitamin D reduces the production of pro-inflammatory cytokines by Th1 and Th17 cells, including IFN-γ, IL-2, and IL-17, while promoting the induction of IL-10-producing Tregs expressing forkhead box P3 (FOXP3) and Cytotoxic T-Lymphocyte Antigen 4 (CTLA-4) [43]. Vitamin D can modulate intracellular pathways mediating IL-2 production, leading to reduced synthesis. Specifically, calcitriol suppresses Nuclear Factor Of Activated T Cells (NFAT)/Activator Protein-1 (AP-1) protein complex formation, implicated in IL-2 expression. The subsequent association of the ligand–VDR-RXR complex to the NFAT binding site on IL-2 promoter inhibits its expression. The reduced IL-2 production prevents activation and proliferation of T cells [44]. Moreover, by inhibiting IFN-γ, vitamin D reduces antigen presentation and T cell recruitment. Th1 cells play a major role in graft rejection and autoimmune diseases, while Th2 cells act as regulatory cells. The inhibition of IL-2 and IFN-γ synthesis mediated by calcitriol induces a switch from a Th1 to Th2 phenotype, thus reducing the risk of autoimmunity and graft rejection [44,45].
However, the modulatory effects of the hormonally active form calcitriol on Th2 differentiation and cytokine production have yielded contradictory findings. Indeed, some studies reported that calcitriol upregulates GATA binding protein 3 (GATA-3) and c-Maf expression, two Th2-specific transcription factors, therefore promoting IL-4 and IL-5 secretion [35,46]. On the contrary, Biswas et al. reported that calcitriol stimulation of murine Th2 cells led to decreased IL-4 and IL-13 levels, accompanied by increased production of IL-10 [47]. Moreover, they observed decreased expression of GATA-3, Growth Factor Independent 1 Transcriptional Repressor (Gfi1), and Interferon Regulatory Factor 4 (IRF4) in calcitriol-treated Th2 cells [47]. Additionally, calcitriol suppresses IFN-γ production in Th1-polarized murine cells and inhibits IL4 in Th2-polarized murine cells; however, it promotes IL-4 production in non-polarizing conditions [48].
Moreover, additional studies reported that calcitriol can regulate a wide range of genes involved in CD4^+^ T cell proliferation and apoptosis. In particular, calcitriol can upregulate the expressions of the genes encoding for CTLA-4, CD38, CYP24A1, the cytokines IL-10 and IL-6, and many transcription factors, including c-Jun, BTB Domain And CNC Homolog 1 (BACH), and STAT3. Interestingly, type 1 (IFN-γ) and type 17 (IL-17, IL-22, and IL-26) cytokine expression in Th cells is suppressed by calcitriol [49]. Given the effects of vitamin D on both innate and adaptive immunity, supplementation may improve immune function and modulate immune-mediated diseases. In the following sections, we will focus our attention on the current knowledge about vitamin D in autoimmune diseases.
5. Vitamin D in Autoimmune Diseases
Autoimmune diseases, including RA, MS, SLE, and T1D, are characterized by immune dysregulation. In particular, aberrant T cell reactivity and autoantibody production contribute to tissue damage. Vitamin D immunomodulatory effects have been a topic of research in the context of autoimmunity. In this setting, vitamin D deficiency has been reported in a range of autoimmune diseases and may impair immune tolerance, potentially contributing to disease onset [50,51]. Hence, the VITAL trial (NCT01169259) reported that 5-year supplementation with 2000 international units (IU; 1 μg vitamin D corresponds to 40 IU) [52] of vitamin D daily, with or without omega 3 fatty acids, reduces the incidence of autoimmune diseases [53]. In addition to the immunomodulatory role of vitamin D, emerging evidence has been gathered regarding the role of VDR single-nucleotide polymorphisms (SNPs) in the pathogenesis of immune-mediated disorders [54]. VDR polymorphisms may alter VDR function and signaling, thereby affecting the immunomodulatory role of vitamin D in the context of autoimmune diseases. The most relevant SNPs studied include BsmI (rs1544410, C>T), ApaI (rs7975232, A>C), TaqI (rs731236, T>C), and FokI + 30,920 (rs2228570, C>T), affecting VDR translation and mRNA stability. Moreover, studies report that the association between SNPs and several autoimmune diseases varies according to the ethnicity of the population [55]. TaqI polymorphism is characterized by a mutation in the transcription region of the VDR gene, which may result in post-transcriptional regulation alterations [56]. ApaI and BsmI polymorphisms are within intron 8, and they may affect VDR function and signaling through alternative splicing. FokI polymorphism is caused by a C-to-T transition in the start codon; therefore, it alters the structure of the final protein [57]. A TaqI heterozygous (CT vs. TT) model has been associated with MS risk in European and Asian populations [58], while the homozygote model (TT vs. CC) has been negatively associated with MS susceptibility in the Iranian population [59]. Additionally, despite no pooled association being described between TaqI SNP and RA risk, a protective role of TaqI has been reported in the African population [55]. TaqI has also been associated with reduced risk of autoimmune thyroid disease [60].
Regarding ApaI, a protective role against MS of ApaI recessive and homozygous model has been reported in Asian patients [58]. Interestingly, the ApaI homozygous model has been associated with increased risk of SLE and RA in the Italian population [61].
A BsmI heterozygous model has been reported to be associated with increased RA risk [62]. Moreover, an association between FokI polymorphism and increased RA risk in Europeans has been reported [63], while the homozygous model has been associated with decreased SLE risk in African and Asian populations [64]. As previously stated, these SNPs may alter VDR function and, therefore, the immunomodulatory role that can be exerted by vitamin D.
In the following sections, we will describe the immunomodulatory effects of vitamin D in the context of immune-mediated diseases, focusing on MS, RA, SLE, and T1D.
5.1. Multiple Sclerosis
MS is a progressive central nervous system (CNS) autoimmune disorder characterized by axonal demyelination, driven by self-reactive T and B cells activated in secondary lymphoid organs. After crossing the blood–brain barrier (BBB), these cells infiltrate the CNS, recognize self-antigens, and trigger an inflammatory response that leads to demyelination [65]. Several studies indicate that vitamin D deficiency is associated with MS progression, relapse, and severity, highlighting its contribution to disease pathogenesis [66,67]. Vitamin D effects in MS are attributed both to its immunomodulatory properties and its direct influence on CNS functions. Vitamin D functions as a neurosteroid, contributing to neuroplasticity and modulating immune activity within the brain [68]. Studies have shown accumulation of vitamin D in the nuclei of neurons and VDR expression in oligodendrocyte-like cells, microglia, and astrocytes, suggesting its role in the transcriptional activation of neuronal messengers. Microglia are immune cells within the CNS whose activation impacts neuron survival and has been linked with MS, schizophrenia, and other neurological disorders. In this framework, vitamin D confers neuroprotection by downregulating proinflammatory cytokines and limiting free radical release from microglia, thereby attenuating the immune response [69].
Indeed, Shirazi et al. reported that calcitriol, the active form of vitamin D, can reduce inflammation in experimental autoimmune encephalomyelitis (EAE), an animal model of MS [70]. In particular, calcitriol administration to EAE mice reduced the number of infiltrating CD11b^+^ and CD4^+^ cells as well as proinflammatory cytokine production, switching to an anti-inflammatory milieu characterized by secretion of IL-10 and IL-4. Furthermore, this study reported reduced CNS demyelination and amelioration of clinical symptoms [70]. Calcitriol supplementation also decreases the number of Th1- and Th17-infiltrating cells, key players of MS pathogenesis. Additionally, preservation of BBB permeability and integrity was observed in both the brain and spinal cord of EAE mice supplemented with calcitriol, potentially contributing to its protective effects against immune infiltration [71].
In light of the experimental evidence regarding the immunomodulatory effects of vitamin D in MS, several clinical trials evaluating the therapeutic benefit of vitamin D supplementation have been performed, as reported in Table 1.
The VIDAMS trial did not report a difference in the relapse rate between patients supplemented with low-dose (600 IU) or high-dose (5000 IU) vitamin D as an add-on to glatiramer acetate over 96 weeks [72]. In the EVIDIMIS trial, patients with relapsing-remitting MS (RRMS) or clinically isolated syndrome (CIS) were supplemented with 20,400 IU (high dose) or 400 IU (low dose) cholecalciferol, the inactive form of vitamin D, every other day for 18 months in addition to IFN β-1b. The primary endpoint consisted of the number of new T2-weighted (T2w) lesions on MRI, while among the secondary endpoints, relapse rate, disability progression, T2w lesion volume. and brain atrophy were evaluated. The low number of participants in this study did not allow for the detection of significant differences in the endpoint considered between the high- or low-dose supplementation [73,74]
However, promising results were reported in the D-Lay-MS randomized placebo-controlled trial, in which 303 untreated patients with CIS suggesting MS were randomized to the placebo group or monotherapy cholecalciferol group (100,000 IU) every 2 weeks for 24 months. Primary endpoints included the presence of disease activity, defined by MRI activity or occurrence of relapse, while secondary outcomes involve the presence of new or enlarged lesions, as well as Expanded Disability Status Scale (EDSS) and Fatigue Scale for Motor and Cognitive Functions (FSMC) scores until disease activity or at the end of follow-up. The results showed that cholecalciferol monotherapy reduces disease activity as well as the occurrence of new or enlarged lesions. However, in the two groups in the study, no significant differences in relapse and clinical scores were observed [75]. Despite no effects on the relapse rate being noticed, these results highlight the potential of high-dose vitamin D as a monotherapy in reducing disease activity and the necessity to study its implication as a supplementation to standard therapy. In this regard, in the CHOLINE study, patients with relapsing MS were randomized into either placebo or 100,000 IU cholecalciferol groups twice monthly for 96 months as an add-on to IFN β-1a (Rebif ^®^) treatment. This study evidenced a significant reduction in annualized relapse rate (ARR), T1 lesions, and EDSS score in patients who completed the study [76]. The protective effects of high-dose cholecalciferol supplementation to RRMS patients treated with IFN β-1a were confirmed by the SOLAR trial, which suggested potential benefits of vitamin D_3_ supplementation on MRI activity [77,78]. The SOLARIUM sub-study [79], enrolling 56 patients from the Netherlands cohort of the SOLAR study, investigated the effects of vitamin D supplementation on regulatory T lymphocytes as well as on cytokine production from Th cells and PBMCs. This study did not find any effect of vitamin D supplementation on Treg absolute count; however, they found differences in the proportion of anti-inflammatory IL-4^+^ Th cells as well as pro-inflammatory cytokines produced by PBMCs between treatment groups.
Despite the importance of vitamin D in regulating the immune system and the ongoing research regarding supplementation, no significant clinical benefits have been reported regarding MS progression. Further studies, enrolling an adequate number of participants, should be performed to evaluate the real contribution of vitamin D supplementation in MS pathogenesis.
5.2. Rheumatoid Arthritis
RA is a chronic inflammatory autoimmune disorder. RA pathogenesis is complex, but it is well known that the interplay of innate and adaptive immune cells, along with autoantibody production, such as rheumatoid factor (RF) and anti-citrullinated protein antibodies (ACPAs), drives synovial inflammation and joint destruction in RA [80].
Many immune cells contribute to RA pathogenesis, including monocytes/macrophages, which play a pivotal role by releasing inflammatory cytokines and reactive oxygen species, thus exacerbating inflammation [81,82]. Given its multiple effects on the immune system, vitamin D may influence disease course. Neve et al. reported that calcitriol-treated macrophages derived from peripheral blood of RA patients presented lower levels of both membrane and soluble TNF-α, a coordinator of the inflammatory response and a major therapeutic target for RA treatment [83]. Moreover, Zwerina et al. found increased macrophage infiltration, proinflammatory cytokine production, and osteoclastogenesis in synovia of VDR knockout human tumor necrosis factor α transgenic (VDR−/− hTNFtg) mice as compared to hTNFtg mice, suggesting a role of VDR, and therefore vitamin D, in mediating synovial inflammation and bone resorption [84].
Within the innate immune players in RA, DCs contribute to the presentation of autoantigens to B cells, promoting autoantibody production and immune complex formation. Additionally, DCs participate in the co-stimulatory activation of CD4^+^ T cells [82]. Th1 cells release TNF-α and INF-γ, contributing to initiation and sustainment of inflammation in RA, while Th17 cells, through IL-17 secretion, intensify the release of inflammatory cytokines and chemokines, thus recruiting other immune cells and contributing to cartilage degradation and bone erosion [85,86]. It has been reported that T cells in synovial fluid are relatively insensitive to calcitriol; therefore, its direct immunomodulatory effects on the tissue may be impaired. In this context, evaluation of vitamin D’s effects on peripheral cells may not provide the whole picture of the local effects of vitamin D in RA [87].
Additionally, in RA, an imbalance between Th17/Treg cells contributes to disease pathogenesis. Surely, Tregs-mediated suppressive mechanisms are essential to control autoreactivity and maintain self-tolerance. However, studies identified reduced frequency and decreased immunosuppressive activity of Tregs in RA patients’ peripheral blood [88]. It has been evidenced that vitamin D promotes IL-10^+^ Tregs differentiation and inhibits CD4^+^ T cell differentiation, which, in the context of RA, may help to restore the Th17/Treg ratio, contributing to amelioration of the disease [89]. In a separate study, T cells isolated from PBMCs of early RA patients and activated with anti-CD3/-CD28 were treated with calcitriol, with or without methotrexate (MTX). After 72 h, the proportion of Th1 and Th17 cells as well as the Th17/Treg ratio decreased in both calcitriol- and calcitriol/MTX-treated groups compared to untreated controls. However, no significant reduction was observed in the calcitriol-treated cells compared to MTX alone [90].
In a randomized, non-blinded interventional study (NCT04472481), 40 patients with active RA were randomized to receive either MTX plus hydroxychloroquine or MTX and hydroxychloroquine plus ergocalciferol (1.25 mg weekly) for 3 months. Tregs were lower in RA patients than in healthy controls at baseline. After three months of treatment, a significant elevation in Tregs in RA patients was reported, particularly in the group receiving vitamin D supplementation [91].
Vitamin D deficiency has been inversely correlated with RA prevalence and disease activity [92]. In particular, RA patients with high disease activity, compared to moderate and low, presented significantly lower serum levels of calcitriol, the hormonally active form of vitamin D [93]. A meta-analysis of 24 eligible studies performed between 1991 and 2015 involving a total of 3489 patients revealed lower levels of vitamin D in RA patients compared to healthy controls. Moreover, they evidenced a correlation between higher vitamin D levels and reduced symptoms, defined by lower disease activity and C-reactive protein (CRP) levels [94]. Additionally, our group reported increased PTH levels, irrespective of vitamin D levels, in patients with autoimmune rheumatic diseases, including RA, compared to controls, suggesting altered vitamin D metabolism [95]. Interestingly, normalization of vitamin D and PTH levels was achieved only after high-dose cholecalciferol supplementation (single 300,000 IU dose, followed by 800–1000 IU daily) for 6 months [96].
Many experimental data suggest a link between vitamin D levels and RA; however, studies on the effects of vitamin D supplementation in RA patients have not given clinically relevant results. Therefore, further studies, both in vitro and in vivo, should be performed in order to elucidate the possible clinical efficacy of vitamin D in the context of RA.
5.3. Systemic Lupus Erythematosus
SLE is a systemic, chronic autoimmune disease, with a prevalence ratio of 9:1 in favor of women. The etiology of SLE is not fully understood; however, it is known that genetic and environmental factors contribute to disease pathogenesis, activating the immune response and inducing B cell autoreactivity, characterized by the production of autoantibodies against nuclear and cytoplasmic antigens, such as anti-double-strand DNA antibodies (anti-dsDNA). Clinically, SLE manifests with different symptoms, which range from mild to severe. The disease commonly presents itself with cutaneous and renal manifestations (lupus nephritis, LN), but it can extend to other systems [97]. A key cutaneous manifestation of SLE is photosensitivity, which, in combination with reduced sunlight exposure and the use of sunscreen protection, may compromise vitamin D cutaneous synthesis [98]. In addition, glucocorticoid use and the proposed existence of anti-vitamin D antibodies may contribute to vitamin D deficiency [99,100,101].
The first studies about vitamin D and SLE date back to 1979, when decreased levels of the active vitamin were reported in serum of prednisone-treated juvenile SLE patients [102]. As observed in other autoimmune diseases, vitamin D deficiency has been widely described in SLE patients and has been linked to disease activity and potentially to the underlying pathogenesis [103]. Vitamin D levels inversely correlate with disease activity scores, along with anti-dsDNA and anti-complement component 1q (C1q) antibody levels [104]. However, whether vitamin D deficiency is a cause or a consequence of SLE is still not fully elucidated [105,106].
As reported in Table 2, several studies have been performed to evaluate the effects of vitamin D and other VDR agonists in SLE, initially focusing on animal models. Moreover, many observational and ex vivo studies on samples derived from SLE patients have been reported in the literature (Table 3), establishing the groundwork for subsequent interventional studies.
Back in 1992, Lemire J.M. et al. reported a decreased degree of proteinuria, autoantibody titers, and skin lesions in calcitriol-treated MRL/1 mice, an animal model of SLE, compared to untreated mice [107]. Despite these findings, no significant effects of calcitriol on proteinuria and on autoantibody production have been reported [107]. Additionally, the impact of vitamin D on renal pathology remains controversial, with conflicting results across studies. Indeed, a study on a New Zealand Black × New Zealand White F1 (NZB × W F1) murine model described aggravation of kidney histology in female mice treated with cholecalciferol, the inactive form of vitamin D [108]. More recently, Freitas et al., in pristane-induced lupus (PIL) mice, did not evidence effects of calcitriol on proteinuria, IgM and IgG deposition, and kidney histology compared to unsupplemented mice, suggesting the incapacity of vitamin D to alleviate renal injury [109]. Interestingly, beneficial effects have also been reported about the use of VDR agonists in LN. In 2022, Li X. et al. described attenuation of this condition in calcitriol-treated MRL/lpr mice. In particular, by suppressing the NF-κB and MAPK pathways, calcitriol mitigates inflammatory responses and ameliorates renal injury in lupus-prone mice [110].
Paricalcitol (9-nor-1,25-diydroxyvitamin D) treatment of Murphy Roths Large/lymphoproliferation (MRL/lpr) mice improves pathological renal alterations and decreases proteinuria and anti-dsDNA antibody levels. VDR activation can reduce anti-dsDNA antibody-induced apoptosis through the inhibition of NF-κB/NLRP3 inflammasome activation, thereby ameliorating renal pathology [111].
VDR activation and effectiveness are essential for eliciting responses to VDR agonists as potential treatments for SLE and LN, highlighting the importance of assessing its expression in relevant tissues. For instance, renal tubular epithelial cells derived from patients affected by active LN display decreased VDR expression, which inversely correlates with injury severity [113]. Moreover, VDR polymorphisms, which could impair its activation, have been associated with nephritic disorders and photosensitivity [114].
Another potential mechanism by which vitamin D may protect against renal damage is through the regulation of autophagy, which contributes to SLE pathogenesis by promoting survival of self-reactive B cells [118]. Indeed, it has been reported that vitamin D can reduce autophagy of LN-patient-derived IgG in human podocytes [115]; however, this study was performed on a small cohort, and these findings are not corroborated elsewhere.
Given the contribution of activated T cells, B cells, monocytes, and NK cells to SLE pathogenesis, the immunomodulatory properties of vitamin D may ameliorate disease course. In Act1-knockout mouse, cholecalciferol restriction (0 IU/kg, low) increased memory B cells compared to mice supplemented with high-dose cholecalciferol (10 IU/kg) for 9 weeks. Moreover, the low-cholecalciferol group showed elevated IgG levels, anti-dsDNA antibodies, and IgG deposition in the glomeruli [112]. Considering the excessive autoantibody production by B cells under low-vitamin D conditions, in vitro studies on PBMCs of SLE patients have shown that calcitriol, the active form of vitamin D, and other synthetic analogues can reduce cellular proliferation, immunoglobulin production, and the frequency of both T and B cells [116]. Moreover, reduced vitamin D levels in SLE patients have been related to enhanced B cell activation, suggesting a potential role of vitamin D deficiency in B cell hyperactivation and autoantibody production [117]. Due to the potential effects of vitamin D in SLE pathogenesis, several clinical trials have also been proposed for this condition, as reported in Table 4.
A study on a cohort of 20 female patients supplemented with cholecalciferol at a dose of 100,000 IU/week for 4 weeks followed by 100,000 IU/month for 6 months (NCT01413230) showed a significant reduction in the frequency of class-switched memory B cells after 2 months and of memory B cells at 6 months. Moreover, the authors found an increased percentage of both resting and activated memory Tregs at 2 and 6 months, along with a decrease in Th1 cells, CD8^+^ T cells, and Th17 cells, compared to the placebo group [119]. Another study in a Portuguese cohort of 24 patients reported decreased levels of Th17 cells and increased levels of Tregs after 6 months of supplementation [123].
In a randomized placebo-controlled trial (NCT01892748), 40 female juvenile SLE patients were supplemented with either 50,000 IU/week of cholecalciferol, the inactive form of vitamin D, or placebo for 6 months. At the conclusion of the study, the group receiving cholecalciferol showed an improved Systemic Lupus Erythematosus Disease Activity Index (SLEDAI), better global fatigue score, and reduced positivity to anti-dsDNA antibodies [120]. Given the reported reduction in bone density and strength in juvenile SLE patients, the same cohort was also evaluated for bone microarchitecture. The results revealed a significant increase in trabecular number, while bone mineral density remained unaffected [121].
In a randomized double-blind placebo-controlled clinical trial of 48 SLE patients (NCT00710021), cholecalciferol supplementation did not significantly decrease IFN signature, defined by the expression levels of IFN-induced genes related to disease activity [122]. However, the supplemented dose, the reduced number of patients and the limited duration of the study may not have been sufficient to provide conclusive results; therefore, further studies regarding the supplementation of vitamin D in SLE should be performed. Moreover, since sex hormones influence disease pathogenesis and treatment response [124], their effects should be taken into consideration when designing interventional studies.
5.4. Type 1 Diabetes Mellitus
T1D is an autoimmune disorder in which immune-mediated destruction of pancreatic β cells leads to the loss of endogenous insulin production, thereby necessitating replacement therapy. In particular, macrophages, CD8^+^ T, Th1, and Th17 cell infiltration in pancreatic islets, as well as Tregs dysfunction, contribute to disease pathogenesis [125]. Many experimental studies (Table 5) and clinical interventional studies (Table 6) have been performed to investigate the effects of VDR agonists in T1D.
Most pre-clinical studies investigating vitamin D in T1D have been conducted on non-obese diabetic (NOD) mice (Table 5). In this model, early-life vitamin D deficiency has been linked to an earlier onset and increased severity of T1D [126]. Interestingly, supplementation of vitamin D to NOD mice improves glucose tolerance. It has been proposed that supplementation downregulates CatG expression, a proteolytic enzyme involved in the modulation of the inflammatory response and glucose homeostasis, thereby inhibiting T cell activation and improving β cell function, as indicated by increased serum levels of C-peptide [127]. Studies on NOD mice supplemented with 800 IU/day vitamin D show that this treatment effectively delays T1D onset and reduces disease incidence. Moreover, increased frequency of Tregs and IL-10-secreting CD4^+^ T cells has been reported [128], highlighting the anti-inflammatory properties of vitamin D and its contribution to the modulation of immune-mediated diseases such as T1D. Based on pre-clinical results in mice, studies regarding the use of vitamin D as an adjuvant immunomodulatory therapy in T1D patients have also been performed, as reported in Table 6. Overall, vitamin D supplementation appears to enhance β cell function [129,132]; however, several studies fail to show meaningful changes in C-peptide levels in treated versus untreated individuals [130]. Moreover, supplementation seems to increase Tregs frequency [132] as well as their suppressive function [133]. Finally, in juvenile patients, vitamin D supplementation seems to decrease insulin requirements [134]. Taken together, these findings suggest a potential role of vitamin D as an adjuvant treatment in T1D.
6. Translational Gap Between Pre-Clinical Models and Human Autoimmune Diseases
Even though pre-clinical models have provided promising evidence regarding the immunomodulatory role of vitamin D, these findings do not translate into consistent clinical benefits in human autoimmune diseases. Indeed, studies report modest or no effects of vitamin D supplementation in autoimmune diseases; therefore, the clinical relevance of this molecule in humans remains inconsistent. The discrepancy between human and murine models highlights the limitations of translating pre-clinical studies to humans. Among the reasons for this discrepancy, species-specific differences between human and murine immune systems as well as differences in vitamin D signaling play a major role. For instance, in human VDRE sequences are located in the promoter of the IL-1β gene, while they are absent in rodents; therefore, IL-1β is not regulated by vitamin D in mice. This highlights fundamental differences in the regulation of immune-related genes in mice and humans [135]. Moreover, the distinct composition and regulation of the VDR gene in mice and humans suggest that the same stimulus may lead to different outcomes in the two species [136]. Additionally, genetic variability, including polymorphisms affecting VDR or vitamin D-metabolizing enzymes in humans, as well as interindividual biological and environmental factors, may increase the difficulties in translating animal studies in humans. Moreover, differences in dosage and timing of supplementation may explain the different findings reported in humans and mice. For instance, to reach sufficient blood levels, mice require a daily intake of 1000 IU/kg of vitamin D [137], corresponding to unrealistically high and potentially dangerous doses in humans, for whom daily intakes of approximately 200–600 IU/day are generally recommended [138]. Furthermore, it is essential to carefully control vitamin D dosage in humans to avoid the hypercalcemic side effects, which are less pronounced or more easily tolerated in murine models [139].
Taken together, these considerations highlight the importance of cautiously interpreting pre-clinical results, acknowledging species-specific mechanisms, and designing clinical studies accounting for the human immune system’s complexity and biological heterogeneity.
7. Conclusions
Evidence supports the role for vitamin D as a modulator of immune responses, with known effects on T cell polarization and inflammatory signaling pathways. However, the translation of these mechanistic insights into clinically meaningful benefits in autoimmune diseases remains limited. At present, results are controversial, depicting the potential immunomodulatory role of vitamin D in in vitro and in murine studies, without reaching consistent results in most clinical trials. Differences between animal models and humans should be taken into consideration while analyzing the effects of a particular compound. In humans, low vitamin D levels have been widely associated with disease activity and severity across multiple autoimmune diseases. However, interventional trials on vitamin D supplementation have given inconsistent results, considering clinical endpoints. Importantly, mechanistic evidence in pre-clinical models relies on the use of different forms of vitamin D or other VDR agonists, while clinical studies frequently rely on cholecalciferol supplementation at different dosages and schedules.
These considerations highlight the limitations of the current literature. First, as previously stated, the term “vitamin D” is frequently used imprecisely, encompassing biologically distinct compounds, which are not always specified and may have a different mechanism of action or bioavailability. Second, observational studies on vitamin D deficiency may not account for the potential confounding induced by reduced sunlight exposure, latitude, chronic inflammation, or medications taken by study participants. Moreover, clinical trials are highly heterogeneous regarding supplementation dose, duration, baseline vitamin D status, outcome measures, and statistical power, thus limiting comparability across studies and reducing the ability to detect clinically meaningful effects. Finally, improvement in immunological or molecular markers in pre-clinical models does not reliably predict the long-term clinical outcomes in the context of complex diseases, underscoring the gap between experimental outcomes and disease modifications. Taken together, current evidence does not support the benefits of vitamin D supplementation as a disease-modifying agent. Rather, vitamin D may be considered to correct deficiency and potentially modulate disease susceptibility or progression, particularly in early or pre-clinical stages.
Based on all the abovementioned considerations, future studies should aim to standardize supplementation strategies and trial designs accounting for disease heterogeneity, vitamin D metabolism, genetic background, and immune phenotype.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Papagni R. Pellegrino C. Di Gennaro F. Patti G. Ricciardi A. Novara R. Cotugno S. Musso M. Guido G. Ronga L. Impact of Vitamin D in Prophylaxis and Treatment in Tuberculosis Patients Int. J. Mol. Sci.202223386010.3390/ijms 2307386035409219 PMC 8999210 · doi ↗ · pubmed ↗
- 2Chun R.F. Liu P.T. Modlin R.L. Adams J.S. Hewison M. Impact of Vitamin D on Immune Function: Lessons Learned from Genome-Wide Analysis Front. Physiol.2014515110.3389/fphys.2014.0015124795646 PMC 4000998 · doi ↗ · pubmed ↗
- 3Sun L. Arbesman J. Piliang M. Vitamin D. Autoimmunity and Immune-Related Adverse Events of Immune Checkpoint Inhibitors Arch. Dermatol. Res.202131311010.1007/s 00403-020-02094-x 32519001 · doi ↗ · pubmed ↗
- 4Formisano E. Proietti E. Borgarelli C. Pisciotta L. Psoriasis and Vitamin D: A Systematic Review and Meta-Analysis Nutrients 202315338710.3390/nu 1515338737571324 PMC 10421389 · doi ↗ · pubmed ↗
- 5Brożyna A.A. Slominski R.M. Nedoszytko B. Zmijewski M.A. Slominski A.T. Vitamin D Signaling in Psoriasis: Pathogenesis and Therapy Int. J. Mol. Sci.202223857510.3390/ijms 2315857535955731 PMC 9369120 · doi ↗ · pubmed ↗
- 6Vernia F. Valvano M. Longo S. Cesaro N. Viscido A. Latella G. Vitamin D in Inflammatory Bowel Diseases. Mechanisms of Action and Therapeutic Implications Nutrients 20221426910.3390/nu 1402026935057450 PMC 8779654 · doi ↗ · pubmed ↗
- 7Gorini F. Tonacci A. Vitamin D: An Essential Nutrient in the Dual Relationship between Autoimmune Thyroid Diseases and Celiac Disease-A Comprehensive Review Nutrients 202416176210.3390/nu 1611176238892695 PMC 11174782 · doi ↗ · pubmed ↗
- 8Infantino C. Francavilla R. Vella A. Cenni S. Principi N. Strisciuglio C. Esposito S. Role of Vitamin D in Celiac Disease and Inflammatory Bowel Diseases Nutrients 202214515410.3390/nu 1423515436501183 PMC 9735899 · doi ↗ · pubmed ↗