Dicer Deletion in the Ear Can Cut Most Neurons and Their Innervation of Hair Cells to Project to the Ear and the Brainstem
Ebenezer N. Yamoah, Gabriela Pavlinkova, Jeong Han Lee, Jennifer Kersigo, Marsha L. Pierce, Bernd Fritzsch

TL;DR
Deleting Dicer in the ear disrupts most auditory neurons and their connections, leading to hearing loss and highlighting the role of microRNAs in ear development.
Contribution
The study reveals how Dicer deletion differentially affects auditory and vestibular neurons and their projections, emphasizing miRNA roles in ear development.
Findings
Dicer deletion eliminates spiral ganglion neurons but spares some vestibular neurons.
Hair cell development is disrupted in the cochlea and canal cristae but preserved in the utricle and saccule.
Central projections from Dicer null mice reach cochlear and vestibular nuclei but not the cerebellum.
Abstract
Dicer is crucial for the generation of microRNAs (miRNAs), which are essential for regulating gene expression and keeping neuronal health. Dicer’s conditional deletion cuts all spiral ganglion neurons but spares a small fraction of vestibular ganglion neurons, innervating the utricle and part of the saccule. Hair cells develop in the utricle, saccule, posterior crista, and the cochlea in Pax2Cre; Dicerf/f. Cochlear hair cells develop at the base and expand the OHC and IHC in the middle, or split into a base/middle and the apex. In contrast, Foxg1Cre; Dicerf/f cuts all canal cristae and cochlea hair cells, leaving a reduced utricle and an exceedingly small saccule. Likewise, Foxg1Cre; Gata3f/f shows no cochlear hair cells and is absent in the horizontal and reduced in the posterior crista. In contrast, the utricle, saccule, and anterior crista are nearly normal, underscoring the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
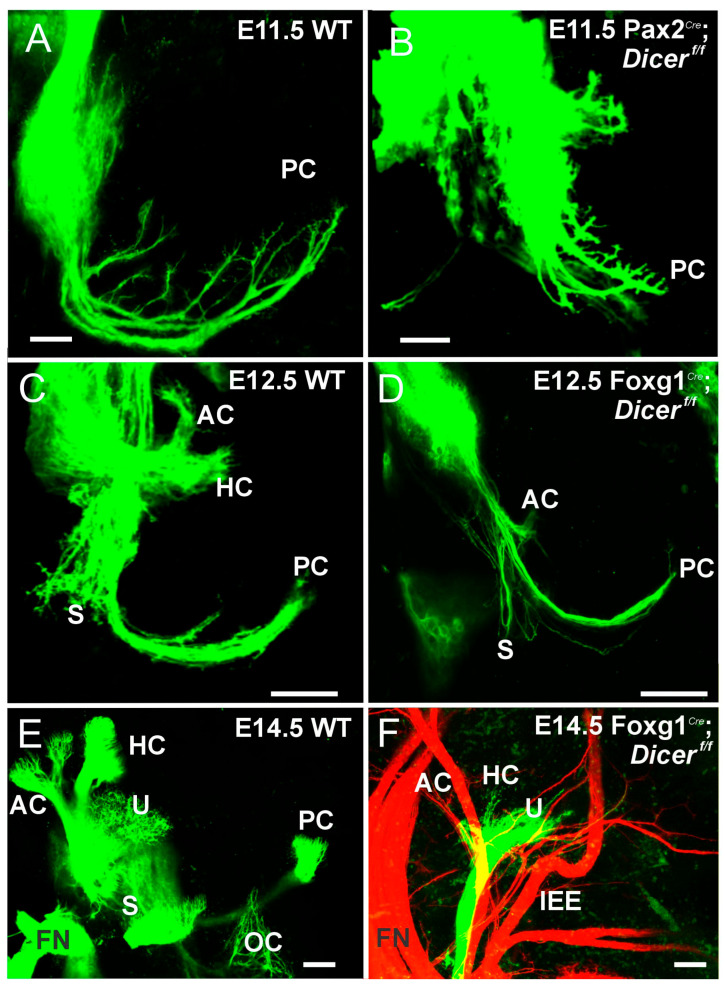 Figure 1
Figure 1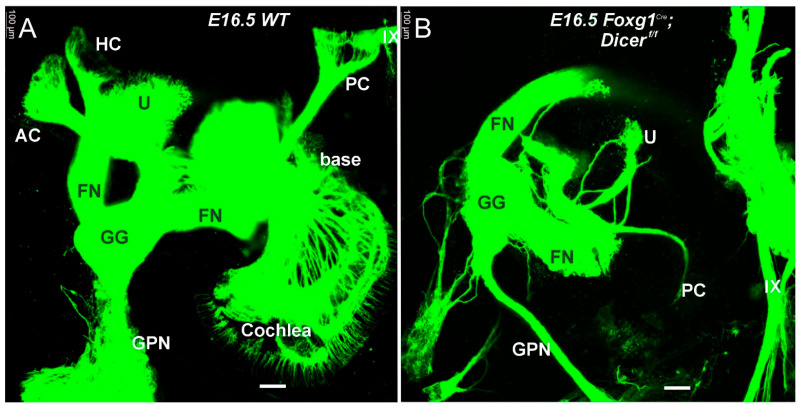 Figure 2
Figure 2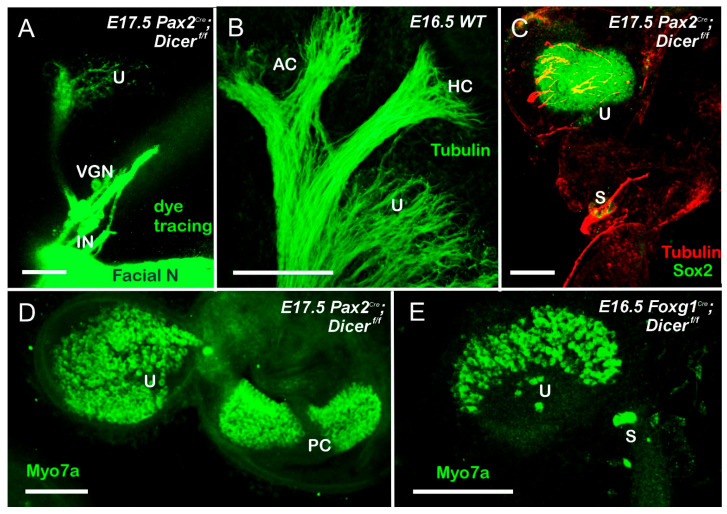 Figure 3
Figure 3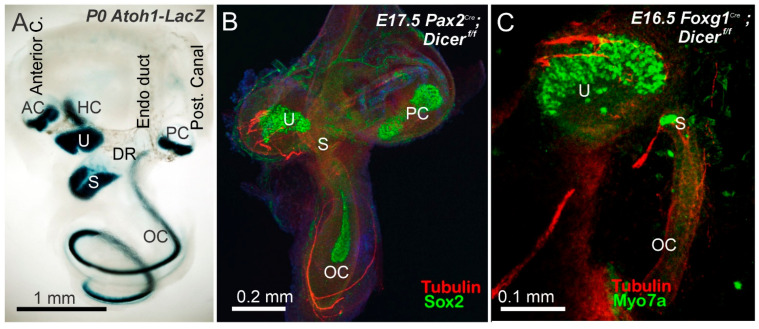 Figure 4
Figure 4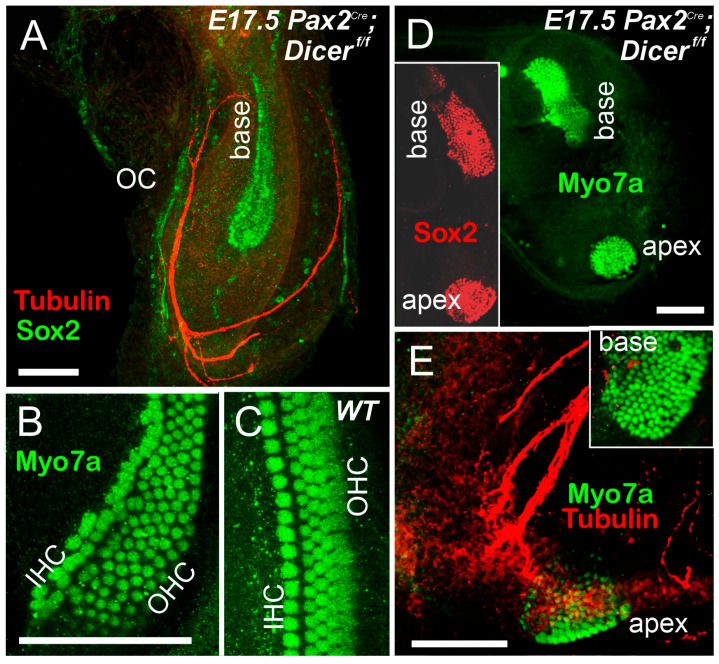 Figure 5
Figure 5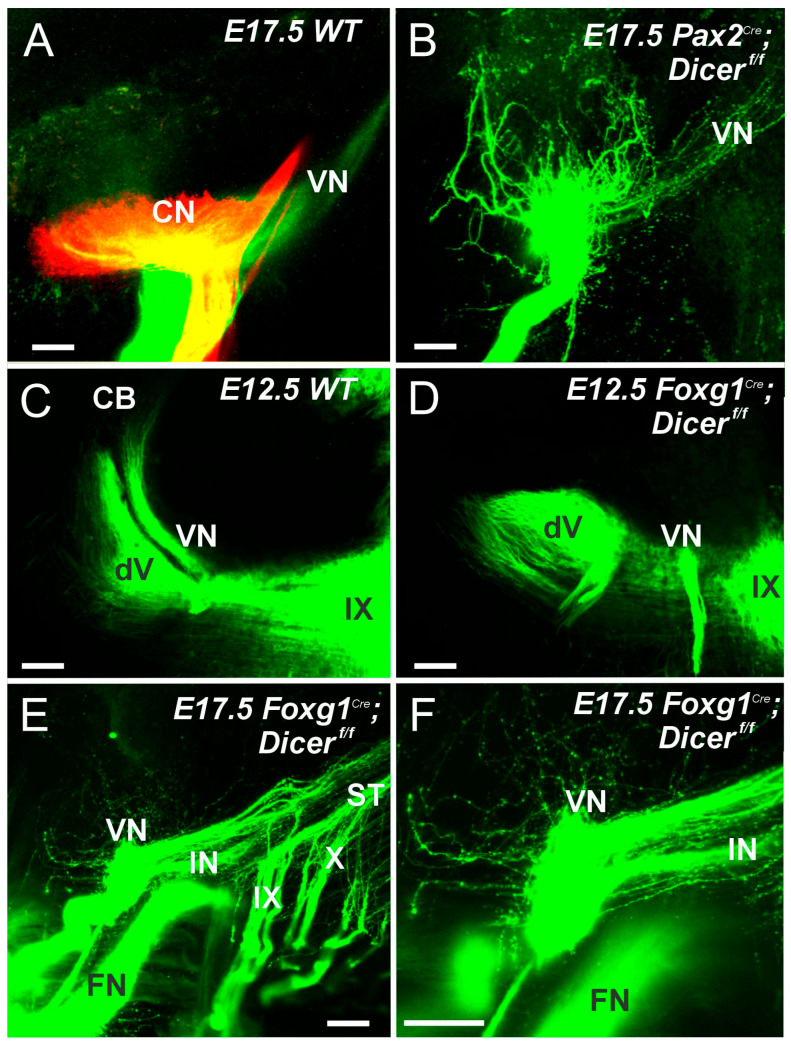 Figure 6
Figure 6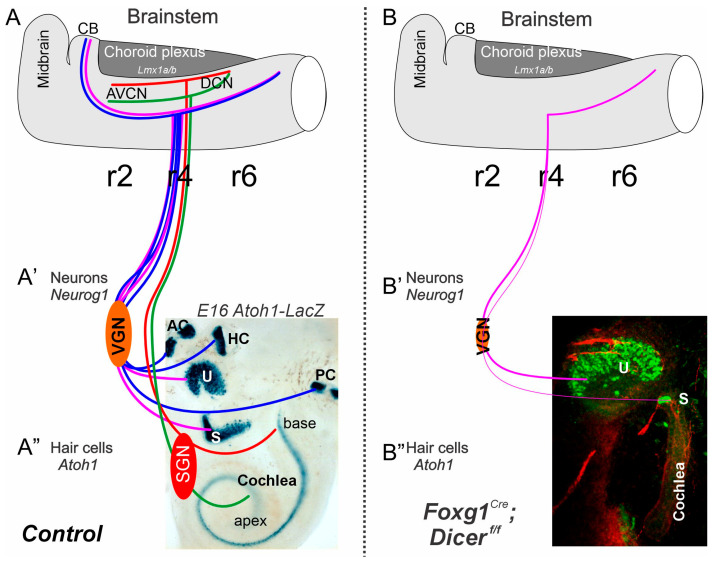 Figure 7
Figure 7- —National Institutes of Health
- —Czech Science Foundation
- —Czech Academy of Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHearing, Cochlea, Tinnitus, Genetics · Vestibular and auditory disorders · Developmental Biology and Gene Regulation
1. Introduction
Vertebrate ears extract sound, angular, and linear acceleration from mechanical stimuli and develop a 3D geometry that connects sensory epithelia (hair cells and supporting cells) via sensory neurons to the brainstem. Molecular analysis over the last 25 years has revealed aspects of the molecular basis for patterning, morphogenesis, and histogenesis of the ear and the brainstem [1,2,3]. How they are integrated into a program that ensures the right genes are regulated at the right time and in the correct place remains unclear. Essential factors that translate early patterning processes into specific areas of expression in developing prosensory epithelia, which in turn govern the morphogenesis of semicircular canals, otoconia, and the cochlea, are not fully known [4,5]. Molecular steps in histogenesis of the vestibular and cochlear begin to emerge, indicating that hair cell formation requires the coordinated transition of gene expressions such as Eya1, Sox2, Neurog1, Isl1, and Neurod1 [6,7,8] to eventually upregulate Atoh1 in postmitotic hair cells to initiate differentiation of these hair cells [2,3]. All of these processes are not only characterized by upregulation of novel gene expression but also by a coordinated downregulation of previously important gene expression, likely involving microRNA (miRNA) to eliminate already existing transcripts to hasten the transition from an expression profile characterizing epithelial cells to another characterizing sensory cells and neurons [9,10,11]. Moreover, previous work showed that the inner ear expresses about 100 miRNAs [12,13], including hair cell-specific miRNAs [14]. Overall, we underscore the complexity of genetic regulation in ear development and the critical roles of signaling pathways and miRNAs in shaping the structure and function of this sensory organ to make hair cells, its vestibular and spiral ganglion neurons, and brainstem nuclei.
miRNAs have a pivotal role in a wide array of plant and animal development and organ functions. Currently, about 1900 miRNAs (see miRbase; releases 22.1) have been catalogued, of which about 500 have been described based on evidence and expression [15,16]. miRNAs require Pol II to start transcription with Drosha and DGCR8 to generate a pre-miRNA that is exported from the nucleus to the cytosol. Dicer is an enzyme that plays a crucial role in the processing of microRNAs and small interfering RNAs (siRNAs). It cleaves long double-stranded RNA (dsRNA) and precursor miRNA (pre-miRNA) molecules into short, functional RNA fragments, which are then incorporated into the RNA-induced silencing complex (RISC) to regulate gene expression [17,18]. Dicer is crucial to generate a specific miRNA population; it dictates the specificity of miRNA of approximately 22–24 bp with a terminal loop and the 3′ overhang [16]. The conserved Dicer (Dicer-1) gene encodes a 220 kDa multidomain protein that is dedicated to the miRNA pathway [19]. In the absence of Dicer, no canonical miRNA will be generated and will typically interact with other gene expression to regulate about 60% specific expression [9], except for deviating miRNAs [20]. Haploinsufficiency with only one functional allele shows specific cancer development [21,22]. Moreover, evolution is known across all archaea, bacteria and eucaryotic and has specific duplication in non-vertebrates [23,24] and is present in the common origin of life, LUCA [25,26]. Simple Dicer deletion blocks normal development beyond the earliest mice development [27]. Using conditional deletion of other genes, such as Pax2^Cre^ or Foxg1^Cre^, can eliminate the floxed Dicer and can provide the absence of specific miRNAs and has specific losses of ganglion neurons, hair cells, and brainstem nuclei [28,29,30,31].
There is an emphasis on the need for detailed analyses comparing the effects of Dicer deletions with other gene mutants. We will compare the effect of Dicer deletion in neurons and their peripheral and central projection with other null mutants to provide a detailed analysis of Shh [32], Neurog1 [33], Neurod1 [6,34], Pax2 [35,36], Gata3 [37], Lmx1a/b [38], and Irx3/5 DKO [39]. This will provide insights into the intricate network of genetic interactions that govern the development of sensory systems within the ear and the brainstem [40,41]. Further research in this field could lead to enhanced understanding of auditory and vestibular system disorders and potential therapeutic strategies for miRNAs [10].
2. Results
2.1. Dicer Deletion Results in the Loss of Most Vestibular Neurons
The sequence of gene expressions in neuronal generation can be tentatively categorized as follows: Eya1 > Neurog1 > Neurod1, among others [2,5,42,43]. In the absence of Eya1, there is limited expression of Neurog1 [42,44]. Specific genes, like Sox2 and Isl1, are needed for the normal development of neurons [41,42]. However, despite a later loss of neurons in Sox2 deletion [45,46], an initial neuronal formation can be demonstrated, using in situ hybridization (ISH), showing overall expression of Neurog1, Isl1, and Neurod1 [42,47]. Likewise, Neurod1 null deletion was thought to result in the loss of all neurons [48], but a more detailed analysis showed an altered neuronal connection in the ear [6,34,49]. Using Pax2^Cre^ and Foxg1^Cre^ to alter inner ear genes shows a more incomplete deletion of Pax2^Cre^; Gata3^f/f^ compared to Foxg1^Cre^; Gata3^f/f^ [37]. Moreover, using antibodies against caspase 3 shows an upregulation of vestibular neurons and eventual loss of nearly all fibers, including dye-tracing [42,47]. Other genes interfere with the normal development of vestibular fiber innervation of hair cells, such as the posterior canal [Fgf10 [50]] or the horizontal canal [Foxg1; [51]] or in the case of Sema, there is an expanded fiber projection [52] similar to the misexpression of Bdnf/Ntf-3 in a transgenic model [53,54]. Moreover, central projection showed the near-normal vestibular fibers in a Foxg1^Cre^; Sox2^f/f^ null mice that lose all central projection in about E16.5 old mice [47].
Dicer deletion takes 3–10 days to deplete specific miRNA [29] while deletion of Pax2 or Foxg1 can eliminate neurons with a shorter delay, ~12–24 h [37]. Even the earliest onset of these Dicer deletions using Foxg1^Cre^ or Pax2^Cre^ shows near-normal first neuron development and near-normal projections to the periphery, including the ear (Figure 1). However, the projection of nerve fibers is diminished. It has a limited projection to the ear, as shown by dye tracing or an antibody, innervating the utricle and saccule (Figure 1 and Figure 2). In contrast, the posterior crista forms but does not reach out to innervate the posterior hair cells (Figure 3) in E17.5 old Pax2^Cre^; Dicer^f/f^ [29]. A smaller utricle and saccule (Figure 3) ensue in Foxg1^Cre^; Dicer^f/f^, which lacks all canal crista formation [28]. Moreover, caspase 3 is upregulated in the VGNs, indicating that the neurons are dying [28,29,30]. Additionally, the smaller ear has reduced otoconia formation in the Pax2Cre; Dicer^f/f^ (Supplemental Figure S1). No otoconia can be found in Foxg1^Cre^; Dicer^f/f^, which likely requires Irx3/5 to develop otoconia, tectorial membrane, and cupulae [39]. Meanwhile, alterations in miRNA, specific for miR-124, resulted in nearly absence of ears [29]. In summary, even with a delay in Dicer loss, nearly all VGNs are lost, except for the utricle and a small saccule neuron. There is an absence of the posterior crista that reaches out toward the canal cristae in Pax2^Cre^; Dicer^f/f^, but there was no hair cell innervation, like in Fgf10 null mice [50], while all canal cristae hair cells and innervation were absent in Foxg1^Cre^; Dicer^f/f^.
2.2. Dicer Requires Spiral Ganglion Neurons
Spiral ganglion neurons (SGNs) develop between E10 and E12.5, progressing from basal neurons to the apex, driven by Neurog1 [7,55]. Specific gene deletions eliminate SGNs. For example, Pax2^Cre^; Gata3^f/f^ shows a small SGN that innervates the cochlear hair cells, while Foxg1^Cre^; Gata3^f/f^ shows the absence of all SGNs and eliminates all cochlear hair cells [37]. An absence of SGNs is shown in Shh null [32] and Lmx1a/b DKO [38], while a small population of Pax2 KO seems to form, but never reaches the usual position ventral to the brainstem [35]. A small population initially forms to develop SGNs but is lost in the cochlea before E14.5 [47]. There is increased neuronal formation, with abnormal extensions into the cochlea and brainstem [6,41].
In the absence of Dicer, no SGN develops (Figure 2, Figure 3 and Figure 4). It is dependent on Pax2^Cre^; Dicer^f/f^ [29] or Foxg1^Cre^; Dicer^f/f^ [28]. Dye tracing (Figure 1 and Figure 2) or using antibodies with tubulin shows a very few fibers that extend to the cochlear hair cells in Pax2^Cre^; Dicer^f/f^, but selective innervation is lost (Figure 4 and Figure 5). Likewise, very few fibers expand toward the cochlea [28]. In summary, without Dicer, there is nearly complete SGN loss, and the few fibers that remain are likely the inner ear efferent fibers (IEE; [56]).
2.3. Absence of Specific Hair Cells After Dicer Deletion
The sequence of events leading to the development of vestibular and cochlear hair cells starts with Prox1 in the anterior and posterior canals at E11.5, which upregulates Atoh1 to initiate vestibular hair cells formation, followed by a graded progression in the cochlea [4,57,58]. Specific null mutations show the loss of hair cells in the posterior crista (PC) [Fgf10; [50]] or horizontal crista (HC; Foxg1; [51]). In contrast, an expansion can ensue in the PC in Lmx1a null mice [59], and can be nearly eliminated in the anterior (AC) and horizontal cristae, while the posterior crista can be very large in Lmx1/Lmx1b DKO [38]. A normal AC in Pax2^Cre^; Gata3^f/f^ mice show a reduced PC and HC. A typical AC forms in Foxg1^Cre^; Dicer^f/f^, but is absent in the HC with a tiny PC [37]. In contrast, only a PC develops in Pax2^Cre^; Dicer^f/f^, while all canal cristae and hair cells are absent in Foxg1^Cre^; Dicer^f/f^.
The otoconia-bearing utricle and saccule depend on genes that separate them into two populations of vestibular hair cells. Otx1 [60], Lmx1a [59] or Lmx1a/Lmo4 [61] deletion, resulting in utricular/saccular fusion. In Irx3/4 DKO mice, there is a fusion between the base of the cochlea and the saccule [39]. Most important is the absence of tectorial membrane and otoconia that are likely absent in Foxg1^Cre^; Dicer^f/f^, but a small otoconia can be seen in the Pax2^Cre^; Dicer^f/f^ mice (Supplement Figure S1; [28,29,30]). Hair cells develop in the utricle but have few vestibular hair cells (Figure 3). Moreover, a few fibers extend into the utricle, as shown with dye tracing (Figure 3A) or antibody labeling (Figure 3C and Figure 4B,C). In summary, utricular hair cells develop in either Pax2 or Foxg1^Cre^; Dicer deletion, but show a reduced presence in a small saccule that also shows a dependence on Neurog1 [7,33].
Cochlear hair cells develop into two populations: one row of inner hair cells (IHC) and three rows of outer hair cells (IHC and OHC; Figure 5C; [8,58]). Detailed analyses show a reduction in the base and loss of OHCs, while in the apex, the number of OHCs increases and has many hair cells compared to IHCs [4]. A shorter cochlea is shown in Neurog1 [33], Foxg1 [51], COUP-TF1 [62], Insm1 and Tbx2 [63] or MycN null deletions [64]. In particular, the apex expands its rows in three mutants [33,51,64], while the shorter base is near normal with one row of IHC and three rows of OHCs. Monotremes, a basic mammal, have expanded IHCs and OHCs and have a unique population in the apex, and the lagena [65,66]. Pax2^Cre^; Gata3^f/f^ shows patches of hair cells [37]. Likewise, Fgfr1 shows patches of cochlear hair cells [67], which also shows hair cell patches in Sox2^Ysb/Ysb^ mice [68,69]. An absence of cochlear hair cells is shown in Foxg1^Cre^; Gata3^f/f^, Lmx1a/b DKO [38], and an unusual Pax2 cochlear nerve underneath the brainstem that is devoid of cochlear hair cells [35].
Compared to cochlear hair cells, if present at all, they show a form of two outcomes: either a shorter, increasingly wider form or a split shorter basal turn that separates from the apex in Pax2^Cre^; Dicer^f/f^ (Figure 5A,B,D,E; [29]). Hair cells can split into the IHC and the much broader OHC, or do not show any distinction from IHC/OHC compared to control (Figure 4B,C). Fibers are expanding, likely from IEE, and receive innervation (Figure 5A,E). Foxg1^Cre^; Dicer^f/f^ lacks all cochlear hair cells [28]. In summary, one conditional deletion of Dicer results in a single or two patches of cochlear hair cells (Pax2^Cre^; Gata3^f/f^ or Pax2^Cre^; Dicer^f/f^ that resemble monotreme organization (base/middle; distinct apex), while Foxg1^Cre^; Dicer^f/f^ resembles Foxg1^Cre^; Gata3^f/f^ conditional deletions of all cochlear hair cells, resembling Shh KO, Lmx1a/b DKO and Pax2 null mice [32,35,38,70].
2.4. Vestibular Fibers Reach out to the Caudal Brainstem but Never Innervate the Cerebellum
A distinct cell population generates Atoh1 to produce cochlear nuclei (CN), which also gives rise to a more ventral population of neurons. In Ptf1a null mice, neurons migrate to the CN [71,72]. Interestingly, even if Atoh1 is deleted in all CN and all HC, the central projection shows the same topology as usual, which likely receives the Ptf1a nuclei [73,74]. In contrast to the elementary organization of Atoh1/Ptf1a, the vestibular nuclei are more complex and spread much more widely [8,75,76]. While the CN consists of only four nuclei (rhombomeres (r) 2–5), the VN extends from the cerebellum to r9. Selective deletion using specific deletion of Atoh1 can show the unique absence of specific rhombomeres that alter the central projection [40]. Central projection can be a misrouted projection in Gata3, Neurod1 or Isl1 conditional deletions [6,37,41]. In addition, using the conditional deletion of Dicer showed a reduction in neurons in the brainstem [31]. Pax2^Cre^; Dicer^f/f^ have a broad expression in the midbrain, the brainstem, and the ear, and eliminate the cerebellum and much of the brainstem with the Dicer deletion [29]. In contrast, Foxg1^Cre^; Dicer^f/f^ is highly expressed in the forebrain, the anterior part of the eye, the cerebellum, and the ear [28]. A significant difference in Dicer expression should be seen in the brainstem between Pax2 and Foxg1. Results show a central projection of control mice labeled from dye tracing of cochlear and vestibular fibers (Figure 6A). In contrast, central projections of Pax2^Cre^; Dicer^f/f^ are ramified across CN fibers that do not show a tonotopic organization, do not reach the cerebellum, and show a caudal projection of VN (Figure 6B). The ramifications of fibers in the absence of SGN are still unclear. Central projection of Foxg1^Cre^; Dicer^f/f^ shows only a minimal projection that exclusively innervates caudal vestibular fibers. Compared to the central projection in E12.5, old mice show projections in parallel to the trigeminal fibers (Figure 6C), while no segregation of trigeminal and vestibular projections is shown in Foxg1^Cre^; Dicer^f/f^ (Figure 6D). A closer examination of the inner ear fibers with the intermediate nerve and facial nerve shows that all remaining central projections expand exclusively to the caudal rhombomere 4–9 (Figure 6E,F), which shows a tiny projection around the CN. In summary, central projection of cochlear and vestibular fibers can show segregation, while in Pax2^Cre^; Dicer^f/f^, there is no sign of a topology and ramifying fibers around the CN and follow caudal VN. In contrast, in Foxg1^Cre^; Dicer^f/f^, they have only very few fibers ramifying across the CN and have a caudal projection next to intermediate nerves to innervate the taste buds.
3. Discussion
Dicer is a central protein that allows the conversion of pre-miRNAs into miRNAs [77,78,79,80]. Despite the central function of Dicer [16], other miRNAs can further upregulate after Dicer deletion [20,81]. In addition, after conditional deletion of Dicer, miRNA can be expressed for hours and days, which will diminish with the presence of miRNA [29]. Given that over 100 miRNAs have been partially identified for the ear [9]. In contrast, the total miRNAs can vastly increase beyond the current identified ~500 miRNAs [16], which leaves open how many miRNAs survive longer or shorter after Dicer deletion. With this warning in mind, we aim to provide an overview of two conditional deletions of Dicer using Pax2^Cre^ and Foxg1^Cre^ for conditional deletion of Dicer.
3.1. The Absence of Dicer Results in the Loss of Nearly All Ear Neurons
Nearly all vestibular neurons show a rapid loss of early expression (Figure 1 and Figure 2) that disappears in miR-124 [29]. Spiral ganglion neurons develop in either Pax2^Cre^ or Foxg1^Cre^ with Dicer deletion. We know that the earliest vestibular neurons develop at about E9 [33,42], while SGNs develop later at approximately E10 [42,55]. This progression of Dicer deletion fits with Foxg1^Cre^; Sox2^f/f^, which initially develop near-normal, but do not expand to innervate the cochlear hair cells [47], or Foxg1^Cre^; Gata3^f/f^ eliminates all SGNs [37]. Deletion is more effective in eliminating all SGNs in Lmx1a/b DKO mice [38] or Pax2 KO [35]. The remaining vestibular neurons in older mice show the innervation of the utricle (Figure 3 and Figure 4) consistent with other vestibular deletions. Dye insertion and tubulin antibody labeling, combined with caspase 3 expression, document the loss of nearly all peripheral inner ear neurons [28,29,30]. The survival of specific small vestibular neurons after Dicer deletion can be attributed to several potential factors:
- Cell-Specific Functions of Dicer: Certain vestibular neurons may have a distinct set of survival pathways or compensatory mechanisms that allow them to thrive despite the loss of Dicer.
- Differential Expression of miRNAs: The small vestibular neurons innervating the utricle may be less dependent on specific microRNAs for their survival compared to other neurons. They may express distinct sets of miRNAs that confer resilience to the effects of Dicer deletion.
- Regional Differences: The utricle may provide unique environmental factors or signaling cues that promote the survival of these specific neurons, such as growth factors or extracellular matrix components that are not present in other areas of the vestibular system, such as Bdnf and Ntf3 [82].
- Developmental Timing: The timing of Dicer depletion may influence neuron survival. If Dicer is removed at a stage where specific small vestibular neurons have already formed critical connections or survival mechanisms, they might tolerate the loss better than others.
- Intrinsic Neuronal Properties: The intrinsic properties of these neurons, such as their metabolic demands or signaling pathways, could make them more resilient to stress induced by the lack of miRNA regulation.
Further work is needed to clarify the survival of a very few vestibular neurons that innervate the utricle nearly exclusively.
3.2. Absence of Dicer Results in Differential Loss of Vestibular and Cochlear Hair Cells
Hair cell development starts in the anterior and posterior crista, where Prox1 is expressed, followed by Sox2 and Atoh1 for vestibular and cochlear hair cells [57,83,84,85]. Three distinct hair cells form: the canal cristae respond to angular acceleration, the utricle and saccule respond to linear acceleration and gravity, and the cochlea responds to different frequencies for hearing. Certain deletions are known, such as specific loss of posterior crista in Fgf10 or horizontal crista in Foxg1 null mice [50,51], while other deletion shows the absence of cochlear hair cells in Lmx1a/b DKO, Pax2 KO or Foxg1^Cre^; Gata3 ^f/f^ mice [35,37,38,86].
Compared to the two Dicer conditional deletions, a difference is noticeable: only the posterior crista forms in Pax2^Cre^:Dicer^f/f^, while all canal cristae are absent in Foxg1^Cre^; Dicer^f/f^. The presence of the posterior crista can likely depend on differential expression of Pax2, which has the same effect as Pax2^Cre^; Gata3^f/f^ [37,87] compared to Foxg1^Cre^; Gata3^f/f^ [37]. It shows nearly complete loss of the posterior crista. It is unclear why the utricle stays and has vestibular hair cells while the nearby saccule is reduced to a few hair cells (Figure 3, Figure 4 and Figure 7). As pointed out on the survival of a very few vestibular neurons, the same arguments can be raised for the few surviving vestibular hair cells in the utricle (and posterior crista).
A distinction of cochlear hair cells shows nicely in the comparison (Figure 4): Pax2^Cre^; Dicer^f/f^ has formed cochlear hair cells, while Foxg1^Cre^; Dicer^f/f^ has no cochlear hair cells (Figure 4, Figure 5 and Figure 7). A similar effect is in Pax2^Cre^; Gata3^f/f^ shows a patch of neurons, while Foxg1^Cre^ eliminates all cochlear hair cells, which is likely related to the slight progression of conditional deletions [37]. Additional information that regulates Neurod1, Notch, Jag1, Hes, and others play a role in abnormal cochlear patterning, increased sensory proliferation, and extra inner and outer hair cells [42,51,62,63,88,89,90] that requires a deeper understanding of the results of Dicer deletion. What is most interesting is the split into two populations in Gata3 or Dicer conditional deletion using Pax2^Cre^: a split into base/middle and an apex (Figure 5 and Figure 7). In addition, a base/middle and apex are shown in MycN null mice [64], while patches of hair cells are documented in Fgfr1 deletion [67] or Sox2^Ysb/Ysb^ deletions [68]. Whether or not this apex patch is compared to the lagena in monotremes remains to be seen [4,65].
3.3. The Absence of Rostral Projection Shows Fibers Toward the Caudal Brainstem
Central projection receives a distinct tonotopic organization from the SGN to innervate the cochlear nuclei (CN) of rhombomere 2–5 (Figure 7; [1]), while vestibular projection, without a topological organization, spreads from the cerebellum to the caudal tip of rhombomere 9 to innervate vestibular nuclei [4,42,76]. Specific deletions of Neurod1, Isl1, and Gata3 show a ramification to innervate the CN, [6,37,41,91], even if certain nuclei are absent in the conditional deletion of specific rhombomeres of Atoh1 [40]. Even a near-normal topology can happen without the CN and hair cell development [73,74]. Moreover, in the absence of Lmx1a/b, DKO mice show no SGNs, no cochlear hair cells, and no cochlear nuclei showing branches of vestibular fibers to ramify across the cerebellum and cross to the contralateral side [38].
Pax2^Cre^ or Foxg1^Cre^, after Dicer deletion, should be differentially affected in the brainstem (Figure 6). Pax2 is broadly expressed in the cerebellum, the ear, and the brainstem, while Foxg1 is expressed in the forebrain, part of the eyes, the ears, and the cerebellum [28,29]. Another conditional deletion of Dicer using Egr2^Cre^ shows a reduced volume formation of the cochlear nuclei. It reduces specific SOCs, whereas delayed upregulation of Atoh1^Cre^ via Dicer deletion results in near-normal cochlear nuclei [31]. In the absence of SGNs, we show a different central projection that ramifies across the CN in Pax2^Cre^; Dicer^f/f^ (Figure 6; [29]) and shows a comparable ramification that has enough SGNs in Pax2^Cre^; Gata3^f/f^ to innervate the CN [37]. Most interesting is the central projection in Foxg1^Cre^; Dicer^f/f^ that lacks all cochlear nuclei and cochlear hair cells and has a small population of VGN to reach out to the VN (Figure 6E,F and Figure 7). Nearly all fibers reach out toward rhombomere 4–9 and show a very few branches that ramify around the CN. It is possible that, in VGN and the small population of the utricle, projection is only caudal, while in normal central projection, it splits into rostral and caudal neurons that show a unique pattern of the utricle [92,93,94]. Moreover, the polarity is different in the utricle between the fibers innervated from the cerebellum versus the caudal brainstem. Unfortunately, conditional deletion does not survive beyond birth in postnatal mice, preventing further analysis of the remaining fibers and their peripheral and central projections and functions [93].
4. Materials and Methods
The absence of Dicer results in early embryonic lethality, but the use of Dicer^f/f^ allows examination of later deletions [27]. We employed conditional deletion of Dicer to show the loss of later expression of two genes, Pax2^Cre^ and Foxg1^Cre^ [95,96], to eliminate Dicer expression (Pax2^Cre^; Dicer^f^^/f^ and Foxg1^Cre^; Dicer^f^^/f^). Embryos were collected from pregnant dams on embryonic days (E) 11.5, 12.5, 14.5, 16.5, and 18.5 that would be dead before birth. There is no replacement for animals, and the minimal number of mice was used. Moreover, mice were housed in a 12/12 cycle provided with food and water in an enriched environment. All animal care and procedures were reviewed by an ethics committee and approved by the University of Iowa Institutional Animal Care and Use Committee (IACUC; #0021971; 19 February 2020).
Pregnant dams were anesthetized with 1.25% Avertin (tribromoethanol) solution at a dose of 0.025 mL/g by IP injection. Deeply anesthetized mice were not responding to pinches. Mice were euthanized after being deeply anesthetized, free of pain, suffering, and distress. Both male and female embryos were used. Embryos were dissected from the uterus and perfusion fixed with 4% PFA in 0.1M phosphate buffer (pH 7.4) with 300 mM sucrose [97]. We used about three Dicer deletion mice of each Pax2 and Foxg1 null and control mice spanning the range of E11.5–E18.5. The smaller size of the ears makes it difficult to dissect the ears, especially in the smallest Foxg1^Cre^; Dicer^f^^/f^. Given the small ears, we used them as whole mounts to see the 3D organization and the remaining innervation.
Immunostaining: We used immunostaining to show specific information. Briefly, antibodies were used to show the nerve fibers with tubulin (SigmaAldrich, St. Louis, MO, USA; anti-α-tubulin #T5168; the RRID–AB_477579 ratio; dilution: 1:500) and show the organization of hair cells using Myo7a (Proteintech, Rosemont, IL, USA; anti-Myo7a #83807; the RRID–AB_3671394 ratio; dilution: 1:700) and Sox2 (Biotechn, Mumbai, India; anti-Sox2 #AF2018; the RRID–AB_355110 ratio; dilution: 1:1000). Tissues were fixed with 4% PFA in 0.1 M phosphate buffer (pH 7.4) and defatted with ethanol, blocked with goat serum with 0.5% Triton X-100, and incubated for one day with primary antibody. Secondary antibody (Thermo Fischer Scientific, Waltham, MA, USA) was conjugated to either Alexa 488, Alexa 543 or Alexa 648 using a dilution of 1:500. Tissues were mounted in glycerol and viewed using a Zeiss confocal microscope (Oberkochen, Germany). In addition, we used Atoh1^LacZ^ to label hair cells [7].
Dye tracing: We used lipophilic dye tracing to label fibers and neurons from brainstem dye insertion to the ear and vice versa [39,97]. The ears were used to enable whole-mount imaging in younger mice, while tissue from older mice was prepared to visualize the ganglia and their innervation to the hair cells. Central projections were split at the midline and flattened to display fiber input. In the oldest mice of Foxg1^Cre^; Dicer^f/f^, which were too small, we inserted dyes to label the VII, VIII, and IX nerves. Images were captured with a confocal system, either (Jena, Germany), using 10× and 20× objectives (NA 0.4 or 0.8). The images were compiled into plates using CorelDraw, 2025.
5. Conclusions
Dicer is essential for the development of miRNA in the brain, eyes, ears, and other body parts. Using conditional deletion of Dicer in specific tissues allows at least the earliest stages of ear development and the formation of its central projections. Despite Dicer deletion, some vestibular ganglion neurons stay, and there is still innervation of vestibular hair cells within the utricle. Although cochlear hair cells do not show innervation, their development follows a pattern of segregation into basal/middle and apex regions. This suggests that the level of developmental organization persists despite the absence of Dicer. Projections from the utricle mainly target the caudal brainstem. These fibers may innervate vestibular hair cells exclusively at specific polarities, showing specialization in neural connections during development.
The findings highlight the complexity of ear and neural development, showing that specific structures can still form and function even with significant genetic changes. Currently, we understand the role of Dicer in LUCA [25,26] and the genetics of vertebrate ears [1,2,3,42] that show the role of miRNA [9,12,14,28,29]. Further research could reveal new pathways and therapeutic targets for treating hearing and balance disorders related to miRNA regulation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Pyott S.J. Pavlinkova G. Yamoah E.N. Fritzsch B. Harmony in the molecular orchestra of hearing: Developmental mechanisms from the ear to the brain Annu. Rev. Neurosci.20244712010.1146/annurev-neuro-081423-09394238360566 PMC 11787624 · doi ↗ · pubmed ↗
- 2Yamoah E.N. Pavlinkova G. Fritzsch B. Molecular Cascades That Build and Connect Auditory Neurons from Hair Cells to the Auditory Cortex J. Exp. Neurol.2025611112010.33696/Neurol.6.11440823538 PMC 12356050 · doi ↗ · pubmed ↗
- 3Petit C. Bonnet C. Safieddine S. Deafness: From genetic architecture to gene therapy Nat. Rev. Genet.20232466568610.1038/s 41576-023-00597-737173518 · doi ↗ · pubmed ↗
- 4Lee J.H. Yamoah E.N. Kersigo J. Elliott K. La Roda N. Pavlinkova G. Fritzsch B. The segregation of Calb 1, Calb 2, and Prph neurons reveals distinct and mixed neuronal populations and projections to hair cells in the inner ear and central nuclei Dev. Dyn.2025 Epub ahead of printing 10.1002/dvdy.70093 PMC 1290776341165193 · doi ↗ · pubmed ↗
- 5Zine A. Fritzsch B. Early steps towards hearing: Placodes and sensory development Int. J. Mol. Sci.202324699410.3390/ijms 2408699437108158 PMC 10139157 · doi ↗ · pubmed ↗
- 6Macova I. Pysanenko K. Chumak T. Dvorakova M. Bohuslavova R. Syka J. Fritzsch B. Pavlinkova G. Neurod 1 is essential for the primary tonotopic organization and related auditory information processing in the midbrain J. Neurosci.201939984100410.1523/JNEUROSCI.2557-18.201830541910 PMC 6363931 · doi ↗ · pubmed ↗
- 7Matei V. Pauley S. Kaing S. Rowitch D. Beisel K.W. Morris K. Feng F. Jones K. Lee J. Fritzsch B. Smaller inner ear sensory epithelia in Neurog 1 null mice are related to earlier hair cell cycle exit Dev. Dyn.200523463365010.1002/dvdy.2055116145671 PMC 1343505 · doi ↗ · pubmed ↗
- 8Elliott K.L. PavlínkováG. Chizhikov V.V. Yamoah E.N. Fritzsch B. Development in the mammalian auditory system depends on transcription factors Int. J. Mol. Sci.202122418910.3390/ijms 2208418933919542 PMC 8074135 · doi ↗ · pubmed ↗