Is the Reindeer Lichen Cladonia arbuscula Really Producing Isousnic Acid? A Chemotaxonomy Query
Dagmar Ísleifsdóttir, Maonian Xu, Maia Biwersi, Marie-Jeanne Leblanc, Starri Heiðmarsson, Snæbjörn Pálsson, John L. Sorensen, Elvar Örn Viktorsson, Elín Soffía Ólafsdóttir

TL;DR
This study investigates whether the lichen Cladonia arbuscula produces isousnic acid, using DNA barcoding and advanced chromatography to confirm its presence in trace amounts.
Contribution
The study confirms the presence of isousnic acid in Cladonia arbuscula using rigorous chromatographic methods and DNA barcoding.
Findings
Isousnic acid was detected in trace quantities in Cladonia arbuscula using advanced chromatographic purification.
Routine metabolite profiling failed to detect isousnic acid due to its low concentration.
A new UHPLC method was developed to distinguish isousnic acid from usnic acid.
Abstract
Isousnic acid (isoUA) has been detected in a few usnic acid (UA)-producing lichens with chemotaxonomic values. IsoUA was first isolated from a specimen belonging to Cladonia arbuscula s.l. (referred to as C. mitis in the publication). However, the isolation and detection of isoUA in this Cladonia species have not been reproduced and confirmed with clear evidence. This study focused on C. arbuscula s.l. collected in Iceland and aimed to (1) identify the lichen specimen using DNA barcoding and (2) investigate whether isoUA is produced using a series of chromatographic methods. The fungal nuclear ribosomal internal transcribed spacer (nrITS) barcode was sequenced, and the specimen was identified as C. arbuscula, following recent circumscription recommendations. Routine metabolite profiling did not detect isoUA, and it could only be identified after vigorous chromatographic purification and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
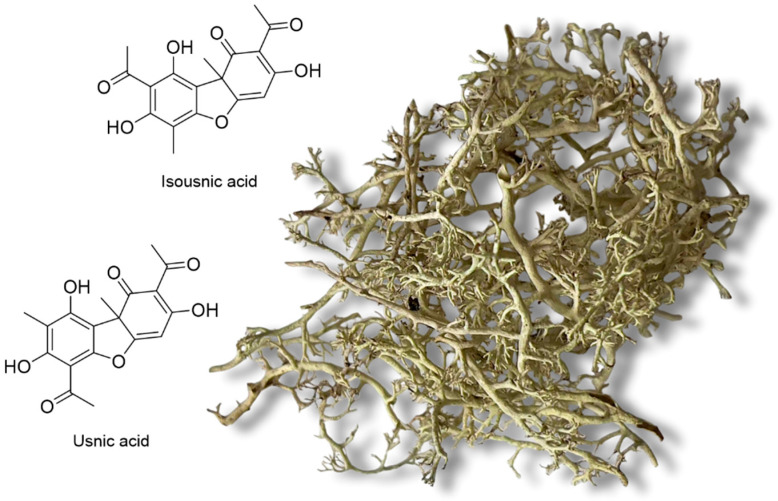 Figure 1
Figure 1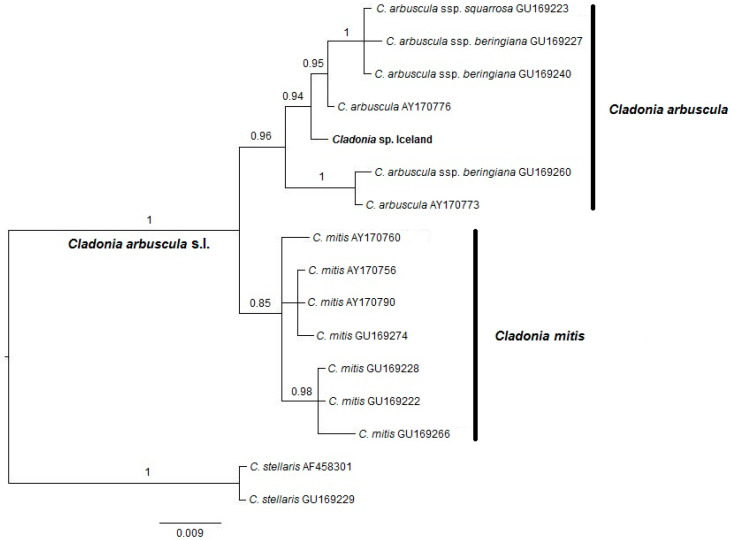 Figure 2
Figure 2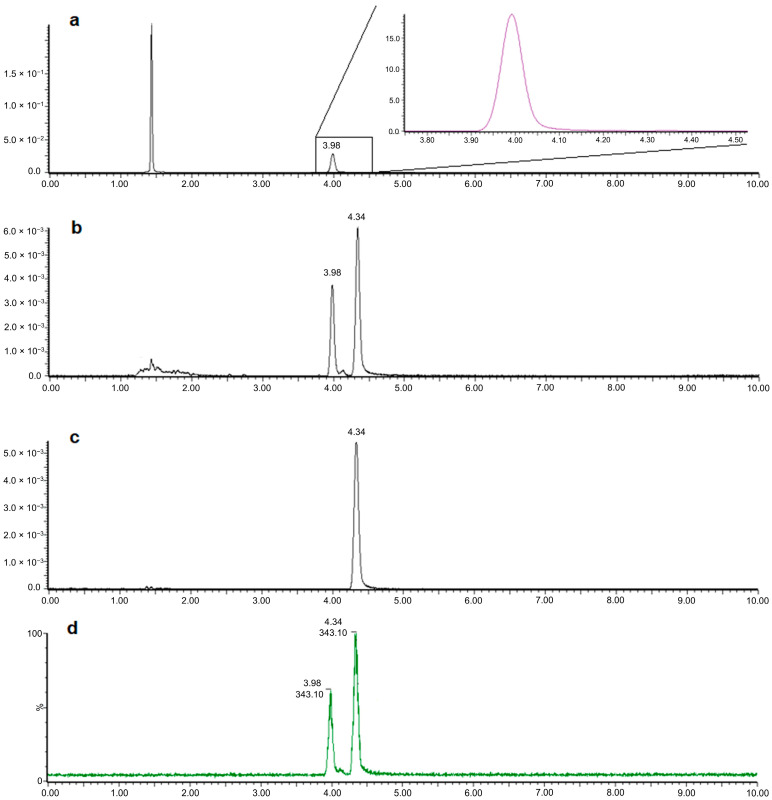 Figure 3
Figure 3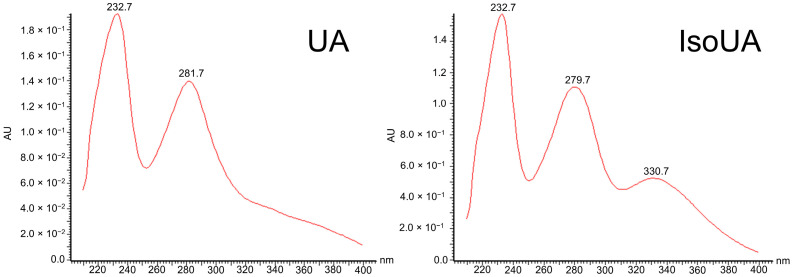 Figure 4
Figure 4- —Icelandic Research Fund
- —University of Iceland Science Park Fund
- —University of Iceland Research Fund
- —Landsvirkjun’s Energy Research Fund
- —Natural Sciences and Engineering Research Council of Canada Alliance International Collaboration
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLichen and fungal ecology · Plant Ecology and Soil Science · Diatoms and Algae Research
1. Introduction
Lichens are classic examples of successful microbial symbiosis, adapting to extreme environments and producing a wide range of unique, specialized metabolites [1,2,3]. These lichen-specific compounds, including depsides, depsidones, and dibenzofurans, serve diverse ecological functions, such as protection against UV radiation, herbivory, and allelopathy [4,5,6,7]. They also display significant biological potential, with reported antimicrobial, anti-inflammatory, cytotoxic, and antioxidant activities [3,8,9]. Lichen chemistry played an indispensable role in the early lichen taxonomy before the advent of DNA sequencing, with most morphological species found to be chemically uniform [10]. However, chemical variations can be found within a single lichen taxon, leading to controversy over whether chemical varieties should be treated as separate species or subspecies [11]. Therefore, it is suggested that chemical characters should be combined and correlated with other independent characters for taxonomic investigations [12].
Among the lichen metabolites, the dibenzofuran usnic acid (UA) (Figure 1) is well known for its antibacterial, phytotoxic and insecticidal activity with important ecological roles [13,14,15]. Although UA has occasionally been shown to be accompanied by its structural isomer, isousnic acid (isoUA) (Figure 1), this isomer has not been widely studied for its bioactivity or ecological functions, probably due to its natural paucity. IsoUA has an important chemotaxonomic value in different lichenized fungal lineages, such as the genus Sphaerophorus (Sphaerophoraceae) [16] and Lecanora (Lecanoraceae) [17].
IsoUA was first isolated from a lichen reported as Cladonia mitis by Japanese phytochemists over half a century ago [18]. However, no evidence (e.g., morphological characters according to monographs) was provided for the identification of the sourced lichen materials in that study, and extensive research efforts have been made in relation to the re-circumscription of C. mitis [19], which is suggested to be treated as a subspecies of Cladonia arbuscula s.l. by Piercey-Normore et al. [20]. In addition, the yield and purification process of isoUA were not described in sufficient detail in the literature to enable reproduction of the results. Since then, the detection of isoUA from this lichen taxon has not been reproduced using chromatographic methods with various sensitivities for purification and detection [18,21,22], except for one putative identification using normal-phase chiral high-performance liquid chromatography (HPLC), which did not include an isolated analytical standard for comparison [23]. Thus, it remains ambiguous whether this lichen truly produces isoUA or not.
The current study focused on the lichen C. arbuscula s.l. collected in Iceland and aimed to (1) identify the specimen using DNA barcoding and (2) investigate whether isoUA is present in this lichen using a series of chromatographic and analytical tools. We intended to explore the effectiveness of integrating DNA-based specimen identification with metabolite profiling to unambiguously link the identified metabolites to the phylogenetically identified lichen species.
2. Results
2.1. Identification of the Lichen Using DNA Barcoding
To verify the species identity of our Icelandic specimen, we performed DNA-based identification. This is particularly important within the Cladonia arbuscula complex, where C. mitis and C. arbuscula are morphologically very similar and can produce overlapping chemotypes [19,20]. The nrITS sequence obtained from the Icelandic specimen has been deposited in GenBank under accession number PX470057. BLAST search (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 26 December 2025) revealed two 100% matches to Cladonia arbuscula voucher sequences (KY266933 and AY17775), and 99.8% identity with C. arbuscula ssp. beringiana (GU169252). The sequence also showed 99.1% identity with a Cladonia submitis voucher (OL694697) and 98.3% identity with a Cladonia mitis strain (OL605254). The BLAST results for the RNA polymerase II second largest subunit (RPB2) sequence (GenBank accession number PX549135) showed the highest identity with C. submitis (99.6%; OL963671 and OL963660), followed closely by C. arbuscula isolates (99.5%). No C. mitis sequences were recovered among the top 50 BLAST hits for RPB2. The Bayesian tree constructed from the fungal nrITS alignment shows two monophyletic clades, consisting of the C. arbuscula clade and the C. mitis clade. The former shows much higher posterior probability than the latter. It confirms that the Icelandic specimen belongs to the C. arbuscula clade (Figure 2). A neighbor-joining tree was also constructed using the nrITS data matrix, which shows the same topology, with our specimen placed in the C. arbuscula clade (Figure S1).
2.2. Purification and Identification of isoUA from Cladonia arbuscula
For the purification of isoUA, we started with automated flash chromatography, which showed a major peak of UA with some tailing. Fractions that potentially contained isoUA were collected after the UA peak (Supplementary File, Figure S2). To remove the contaminating UA, we continued the purification with semi-preparative HPLC, which provided the baseline separation of UA (19.17 min) and the putative isoUA peak (21.41 min) (Supplementary File, Figure S3). After evaporation, 0.4 mg of isoUA was obtained.
Routine ultra-high-performance liquid chromatography–photodiode array detection–mass spectrometry (UHPLC-PDA-MS) profiling of C. arbuscula extract showed the presence of UA (t_R_ = 3.98 min), with no sign of isoUA (Figure 3a). However, after combining 34-times collection of effluents (following the UA peak) from the flash chromatography, the isoUA peak became visible at 4.34 min (Figure 3b). With further semi-preparative HPLC purification, isoUA was obtained (Figure 3c) with over 99% purity at 280 nm. Its identity was confirmed by comparing the retention time with a reference lichen (B. ramuliferum) that produces isoUA (Supplementary Files, Figure S4), and by MS data (Figure 3d). The high-resolution MS spectrum of the purified isoUA is shown in the Supplementary File, Figure S5. In addition, we importantly observed a third UV absorbance maximum for isoUA at 330 nm, which is not observed for UA (Figure 4).
3. Discussion
Accurate identification of lichen or plant material is a prerequisite for the reproducibility of scientific results. It is especially important when it comes to taxa with subtle morphological differences among closely related taxa. The reindeer lichen C. arbuscula is taxonomically challenging, as the nucleotide variation in the fungal nrITS locus does not always correlate with morphological variation [19]. Some phylogenetic analyses have indicated a distinct C. mitis clade with clear splitting from a more diffuse C. arbuscula s.l. clade [20]. We used the same sequence matrix as used in that study to identify our Icelandic Cladonia specimen. It was shown to belong to C. arbuscula s.l., clearly separated from the circumscribed C. arbuscula ssp. mitis clade (Figure 2) [20]. We also produced the RPB2 sequence from the specimen with high similarity (99.6% match) to one reference sequence of C. submitis. However, the limited availability of C. arbuscula RPB2 sequences currently prevents a more comprehensive phylogenetic analysis using this locus. In two phylogenetic studies, C. submitis seemed to be more closely related to the C. arbuscula s.l. lineage than the C. mitis clade [24,25]. DNA barcoding and phylogenetic analysis are feasible and rapid methods for specimen identification, which allows comparison with other reference sequences deposited online. It is especially useful when distinguishing specimens with subtle morphological differences.
Although effective extraction of plant polyphenolics commonly uses aqueous acetone [26], the extraction of UA (and isoUA) requires more lipophilic solvents due to the intramolecular hydrogen bonding that decreases the polarity of the molecule. In this study, ethyl acetate was selected over acetone, as UA exhibits higher solubility in this solvent compared with others [27]. In addition, the use of fully homogenized powdered lichen materials is essential, as it maximizes the extraction yield of UA-like cortical compounds [28].
IsoUA is more lipophilic than UA, as indicated by its faster elution in both silica-based thin-layer chromatography [18] and normal-phase HPLC [14]. Similarly, in our analyses using reversed-phase methods, IsoUA consistently eluted later than UA. Therefore, the identification of isoUA is primarily based on the chromatographic behavior and retention time. In our study, however, we also observed distinct differences in the UV spectra of the two isomers. Notably, isoUA exhibits a unique third UV absorption maximum at 330 nm, which is shown here more clearly than in the UV spectra previously reported by Kinoshita et al. [23]. This characteristic UV signal could serve as a key spectroscopic marker facilitating the rapid chromatographic identification of isoUA in future studies, particularly since UV detectors are standard equipment in most analytical laboratories.
The isoUA in our Icelandic Cladonia specimen is produced in a very low quantity of 24 µg/g or 24 ppm, which likely explains the absence of the compound in the recent literature [18,21,22] working on the same or similar lichens. The reference study by Kinoshita et al. [23], which reported the presence of isoUA in C. mitis, did not show the chromatogram or specify the amount of isoUA obtained, and therefore, it is hard to compare our results with this reference study. Chromatographic separation with reference to the lichen B. ramuliferum and high-resolution mass spectrometry both confirmed the identity of isoUA in our study. However, the low yield and time-consuming repetitive purification process did not allow for alternative structural elucidation using multiple other spectroscopic methods, including but not limited to nuclear magnetic resonance and electronic circular dichroism. The compound from the Icelandic specimen is probably (+)-isoUA, as indicated by the retention time in the chiral HPLC study [23]. From the same study, it appears that the enantiomeric form of isoUA corresponds to the enantiomer of UA produced by the lichen.
IsoUA is known to have important chemotaxonomic values in the lichen genera Bunodophoron (Sphaerophoraceae) [16] and Lecanora (Lecanoraceae) [17]. Within Lecanora, isoUA production is confined to one of the three recognized Lecanora species groups, which include L. albellula, L. coniferarum, L. laxa, and several other Lecanora species [17]. Its presence can be correlated with the shape of the conidia for species group distinction [17]. In an earlier study, at least three Bunodophoron species were found to produce isoUA as a major metabolite, whereas other species lacked it [16]. Although isoUA is also produced in several Cladonia taxa [29], its presence and enantiomeric form in chemotaxonomy have not been explored. This calls for a highly efficient and sensitive chiral HPLC method to separate all four enantiomers. Our previously published chiral reversed-phase HPLC method [30] does not allow simultaneous separation of UAs and isoUAs. Therefore, an advanced chiral HPLC method that separates all four isomers (i.e., both enantiomers of UA and isoUA) in a single run is under development in our laboratory.
4. Materials and Methods
4.1. Lichen Sampling
The lichen (morphologically identified as Cladonia arbuscula s.l.) was collected in the northern part of Iceland in Skagafjörður (Arnarstapi, N: 65.5301, W: −19.5154) on 10 October 2024 (Figure 1). It grows on old glacier moraine with little vascular plant cover, along with Alectoria sarmentosa ssp. vexillifera and Cetraria lichens (e.g., C. islandica). The voucher specimen is deposited in the herbarium of the Icelandic Institute of Natural History, Akureyri division, with the voucher number LA32065. The lichen specimen, Bunodophoron ramuliferum (L45297 TROM), was borrowed from the Tromsø herbarium as a reference lichen, as it produces isoUA as a major metabolite [31].
4.2. Identification of the Lichen by DNA Barcoding
Genomic DNA was extracted from 15 mg of thallus tissue following the CTAB protocol [32]. Two barcoding loci, nrITS and RPB2, were amplified using mycobiont-specific primers developed for Cladonia lichens [33]. Touchdown PCR amplifications were carried out [34] and sequenced via Sanger sequencing (Macrogen Europe BV, Amsterdam, the Netherlands). The raw reads were trimmed, aligned, and curated in Geneious Prime^®^ 2024.0.5, and the resulting sequences were compared against GenBank references using BLAST. We used the nrITS alignment data matrix from Piercey-Normore et al. [20], which includes worldwide Cladonia voucher specimens and separates C. arbuscula s.l. and C. mitis into monophyletic groups. We then manually aligned our Icelandic sequence to this matrix to determine whether the specimen belongs to C. arbuscula s.str. or C. mitis. A Bayesian tree was constructed following the procedure we have previously described [34] using the software MrBayes 3.2 [35]. The software MEGA-X version 10.2.2 [36] was used to construct the neighbor-joining tree using maximum composite likelihood method with 500 bootstrap replicates. Following a guideline [37], we used the uncorrected p-distance for distance matrix calculations, without any pre-assumed evolutionary model. The neighbor-joining tree was constructed to graphically summarize the genetic distance data. To reduce the risk of specimen misidentification, a Bayesian tree was also constructed following the procedure we have previously described [34]. The evolutionary model was estimated using PartitionFinder2 [38].
4.3. Sample Preparation and Purification
A total of 16.5 g of dried lichen material was ground into a fine powder and macerated overnight in ethyl acetate (600 mL). The resulting extract was filtered, evaporated to dryness, and redissolved in a minimal volume of acetonitrile (ACN). To concentrate any isoUA in the extract, we first used automated flash chromatography (puriFlash^®^ 5.250) via with a C18-XS column (15 µm, 250 × 21.2 mm; Interchim, Montluçon, France) to collect any metabolites eluting after the major component UA. The mobile phase consisted of solvent A (ACN + 0.1% formic acid (FA)) and solvent B (H_2_O + 0.1% FA), used in an 80:20 (A:B, v/v) ratio. The flow rate was set to 16 mL/min, with UV detection at 280 nm. The injection volume was 2 mL. Since isoUA is more lipophilic than UA, all the fractions eluting after the predominant UA peak were collected and pooled for each run. A total of 34 injections were made to concentrate the isoUA using the flash chromatography system (Supplementary File, Figure S2). The solvent was then evaporated, and the residue was redissolved in ACN. As the collected isoUA was contaminated with UA after flash chromatography, we continued with semi-preparative HPLC (Dionex Ultimate 3000; Thermo Scientific, Watham, MA, USA) using a Luna Phenyl-Hexyl column (250 × 10 mm, 5 µm; Phenomenex, Torrance, CA, USA) for further purification of the isoUA. The mobile phase consisted of solvent A (10% ACN/H_2_O + 0.1% FA) and solvent B (ACN + 0.1% FA) in a 30:70 (A:B, v/v) ratio. The flow rate was 2.5 mL/min, and the UV detection wavelength was set to 280 nm. Samples were dissolved in ACN/H_2_O (70:30, v/v) with 0.1% FA, and 1 mL was injected per run. The injection process was repeated eight times to obtain purified isoUA (Supplementary File, Figure S3). The purified isoUA was identified by comparing its retention time, UV spectrum and molecular mass with those obtained from the isoUA-producing reference lichen (i.e., B. ramuliferum) by metabolite profiling, as shown in Figure 3.
4.4. Metabolite Profiling
Our routine UHPLC-PDA-MS profiling started with lichen extracts, which were re-constituted from the acetone extract of ca. 15 mg lichen thallus materials [39,40]. Briefly, the intact lichen thalli were ground under liquid nitrogen and extracted twice with acetone. Then, the acetone was evaporated, and the dried residues were dissolved in methanol/ACN (50:50, v/v). An aliquot of the reconstituted solution was diluted 20 times with 50% ACN in H_2_O and filtered before HPLC-PDA-MS profiling. One Kinetex F5 column (3 × 150 mm, 2.6 µm; Phenomenex, Macclesfield, UK) was used for the separation of the UA and isoUA isomers. An isocratic elution was used under ambient temperature (22 ± 1 °C) at 0.45 mL/min flow rate, and the mobile phase consisted of H_2_O:ACN (25:75, v/v) with 0.1% FA. The UV spectrum was recorded from 200 to 500 nm, and the MS full scan was monitored from a 200 to 800 mass to charge ratio (m/z) in the negative ion mode. The isomeric UA and isoUA were monitored at the same molecular ion at 343 m/z, and their resolution primarily depended on chromatographic separation.
5. Conclusions
Overall, our study found that isoUA is produced in a very low quantity (24 ppm) in the well-characterized, DNA-barcoded Cladonia arbuscula from Iceland. This low quantity may therefore easily be overlooked during routine metabolite profiling. Its presence and identity were confirmed by UPLC-PDA-MS analysis after vigorous chromatographic purification steps. The characteristic UV spectrum obtained for isoUA could serve as a key spectroscopic identification marker in future studies. Our study emphasizes the need for more sensitive methods in lichen chemotaxonomy when chemical markers are present in extremely low quantities and highlights the value of integrating DNA barcoding with metabolite profiling in lichen chemistry research.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hawksworth D.L. Grube M. Lichens redefined as complex ecosystems New Phytol.20202271281128310.1111/nph.1663032484275 PMC 7497170 · doi ↗ · pubmed ↗
- 2Singh G. Dal Grande F. Martin F.M. Medema M.H. Breaking into nature’s secret medicine cabinet: Lichens—A biochemical goldmine ready for discovery New Phytol.202524643744910.1111/nph.7000340007421 PMC 11923402 · doi ↗ · pubmed ↗
- 3Xu M. Heidmarsson S. Olafsdottir E.S. Buonfiglio R. Kogej T. Omarsdottir S. Secondary metabolites from cetrarioid lichens: Chemotaxonomy, biological activities and pharmaceutical potential Phytomedicine 20162344145910.1016/j.phymed.2016.02.01227064003 · doi ↗ · pubmed ↗
- 4Nguyen K.-H. Chollet-Krugler M. Gouault N. Tomasi S. UV-protectant metabolites from lichens and their symbiotic partners Nat. Prod. Rep.2013301490150810.1039/c 3np 70064 j 24170172 · doi ↗ · pubmed ↗
- 5Solhaug K.A. Gauslaa Y. Secondary Lichen Compounds as Protection Against Excess Solar Radiation and Herbivores Progress in Botany 73 Lüttge U. Beyschlag W. Büdel B. Francis D. Springer Berlin/Heidelberg, Germany 2012283304
- 6Rundel P.W. The ecological role of secondary lichen substances Biochem. Syst. Ecol.1978615717010.1016/0305-1978(78)90002-9 · doi ↗
- 7Bačkor M. Goga M. RučováD. UrminskáD. BačkorováM. Klejdus B. Allelopathic effects of three lichen secondary metabolites on cultures of aposymbiotically grown lichen photobionts and free-living alga Scenedesmus quadricauda S. Afr. J. Bot.202316268869310.1016/j.sajb.2023.10.005 · doi ↗
- 8Molnár K. Farkas E. Current Results on Biological Activities of Lichen Secondary Metabolites: A Review Z. Naturforsch. C J. Biosci.20106515717310.1515/znc-2010-3-40120469633 · doi ↗ · pubmed ↗