Polyphenols, Polysaccharides, and Their Complexes from Aronia melanocarpa in the Chemoprevention of Colorectal Cancer
Karolina Niewinna, Katarzyna Owczarek, Zuzanna Senkowska, Urszula Lewandowska

TL;DR
This paper reviews how compounds from black chokeberry may help prevent colorectal cancer through their antioxidant and anti-inflammatory effects.
Contribution
The paper highlights the chemopreventive potential of Aronia melanocarpa compounds and their complexes in colorectal cancer.
Findings
Aronia melanocarpa contains polyphenols and polysaccharides with anticancer properties.
These compounds modulate apoptosis pathways and reduce inflammatory markers like interleukin-6 and tumor necrosis factor alpha.
Complexes of these compounds may enhance chemopreventive efficacy through synergistic mechanisms.
Abstract
Colorectal cancer (CRC) is among the three most commonly diagnosed malignancies worldwide and remains a major public health challenge, emphasizing the need for effective preventive strategies. Considering the current chemotherapy limitations of key agents, natural products widely researched as dietary supplements can complement conventional treatments. This review concentrates on Aronia melanocarpa (black chokeberry), including its fruits, leaves and pomace, as a rich source of bioactive compounds with well-documented anticancer properties. Notably, A. melanocarpa contains high levels of polyphenols such as cyanidin-3-galactoside, cyanidin-3-arabinoside, chlorogenic acid, quercetin, and epicatechin, as well as biologically active polysaccharides, including pectins and arabinogalactans. These compounds, through their antioxidant and anti-inflammatory activities, are involved in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
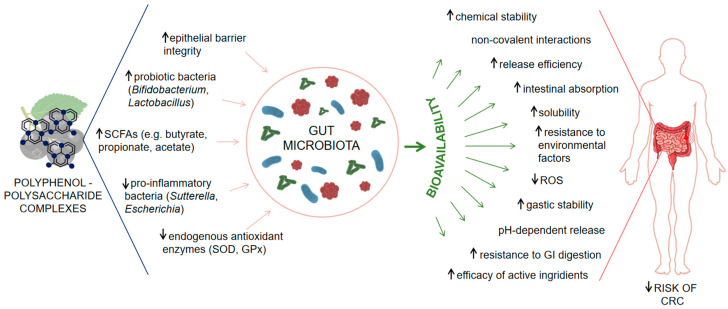 Figure 1
Figure 1- —Medical University of Lodz, Poland
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhytochemicals and Antioxidant Activities · Microbial Metabolites in Food Biotechnology · Polysaccharides and Plant Cell Walls
1. Introduction
1.1. Overview of Colorectal Cancer
Colorectal cancer (CRC) is one of the most commonly diagnosed malignancies and a leading cause of cancer-related mortality worldwide, with 1.93 million new cases and 935,000 deaths reported in 2020 [1]. While incidence remains highest in high-income countries, rising rates in low- and middle-income regions are linked to Westernized lifestyles. It should be emphasized that, based on projections related to aging, population growth, and societal development, the annual number of newly diagnosed CRC cases is expected to reach approximately 3.2 million by 2040, with annual deaths rising to about 1.6 million. The anticipated increase in CRC incidence is primarily associated with greater exposure to environmental risk factors [2].
Modifiable risk factors such as diets rich in red and processed foods, low fiber intake, obesity, physical inactivity, alcohol consumption or smoking significantly contribute to CRC development [3]. A recent study confirms that pro-inflammatory lifestyle patterns strongly correlate with increased CRC cases [4]. Chronic inflammatory bowel diseases (IBD), especially when associated with primary sclerosing cholangitis, further elevate CRC risk due to persistent mucosal inflammation [5]. Advances in molecular diagnostics and endoscopic surveillance have improved CRC risk stratification in IBD populations [6]. Factors such as age, sex, family history, and hereditary syndromes (e.g., Lynch syndrome, familial adenomatous polyposis) also play a critical role [7]. CRC is largely preventable through early detection and screening, though access imbalance carry on globally [8]. At present, considerable emphasis is placed on the prevention of CRC development, with particular focus on chemoprevention, as this approach has the potential to substantially reduce disease incidence. This strategy involves the search for agents—primarily of natural origin—to inhibit, delay, or reverse carcinogenesis, and has shown promise in high-risk populations as a complementary approach alongside screening and lifestyle interventions.
1.2. The Role of Chemoprevention in CRC
Chemoprevention is a rapidly evolving and promising strategy for the prevention of CRC, largely due to the slow progression of the disease from precancerous adenomas to invasive carcinomas. This extended latency provides a crucial opportunity to intervene using natural or synthetic agents capable of halting or delaying tumor development [9]. Among various approaches, dietary chemoprevention has garnered particular interest, supported by the well-established associations between nutrition, lifestyle and CRC risk [10].
Plant-derived bioactive compounds, particularly polyphenols, exert chemopreventive activity through multiple biological mechanisms. These include the modulation of oxidative stress, suppression of chronic inflammation and regulation of key cancer-related signaling pathways [11,12]. Notably, polyphenols such as resveratrol, quercetin, epigallocatechin gallate and curcumin have shown potent antioxidant, anti-inflammatory, pro-apoptotic, and anti-angiogenic properties in CRC studies [10,13].
In addition to direct effects on tumorigenic processes, dietary polyphenols and plant-derived polysaccharides also influence colorectal carcinogenesis via modulation of the gut microbiota. These compounds can enhance epithelial barrier integrity, regulate host immune responses and shape a microbial environment that supports anti-inflammatory and anticancer outcomes [10]. Furthermore, microbial metabolism of dietary components leads to the production of key bioactive metabolites, such as short-chain fatty acids (SCFAs)—notably butyrate, acetate and propionate acids—which have been shown to induce apoptosis, suppress inflammation and regulate gene expression in colonic epithelial cells [14].
Emerging evidence also supports the concept of synergistic interactions among multiple bioactive compounds naturally present in whole food, such as Aronia melanocarpa, enhance their chemopreventive potential. This supports the focus on this botanical source as a promising dietary strategy for CRC prevention, particularly in the context of its rising global incidence [10,11,12].
1.3. Significance of Aronia melanocarpa in Preventive Medicine
Black chokeberry (Aronia melanocarpa) is a plant resident to the Rosaceae family. This particular species is gaining increasing recognition as a functional food with exceptional potential in the prevention of chronic diseases. Its health-promoting effects stem primarily from its remarkably high content of polyphenolic compounds—particularly anthocyanins, proanthocyanidins, flavonoids and phenolic acids—which exhibit strong antioxidant, anti-inflammatory and cytoprotective properties [15]. Compared to other berries and polyphenol-rich fruits, A. melanocarpa demonstrates superior capacity to modulate pathways relevant to colorectal cancer (CRC) prevention, including oxidative stress, NF-κB-mediated inflammation, gut microbiota composition, and intestinal barrier integrity [15,16,17].
In vitro studies have demonstrated that extracts from chokeberry leaves and berries can induce apoptosis, inhibit proliferation, and suppress migration and invasiveness of cancer cells by modulating pathways such as NF-κB, epithelial–mesenchymal transition (EMT), and angiogenesis [18,19]. Different studies also highlight chokeberry polyphenols’ ability to inhibit LPS-induced inflammation in colon epithelial cells and macrophages, further reinforcing their relevance in CRC-related inflammatory pathways [20]. These anticancer effects align with broader polyphenol mechanisms, such as cell cycle arrest, induction of apoptosis, inhibition of angiogenesis, and suppression of tumor-promoting signaling pathways—previously described in comprehensive reviews [21].
In vivo studies indicate that chokeberry supplementation can improve oxidative stress markers, lipid profiles, glucose metabolism, and gut microbiota composition, supporting gastrointestinal health and intestinal barrier integrity [16,17].
Clinical studies in humans, more limited, have reported beneficial effects on metabolic and inflammatory markers, including reductions in C-reactive protein (CRP), improvements in lipid and glucose profiles, and enhanced antioxidant enzyme activity (SOD, GPx, CAT) following juice or extract consumption [22].
Extracts from various aronia species show cytotoxic and antiproliferative activity against several cancer cell types—including colorectal, breast and prostate cancer—highlighting their broad chemopreventive potential [20]. These effects are linked to the high content of polyphenols and their ability to modulate oxidative stress and inflammation. Systematic reviews further confirm that chokeberry supplementation reduces markers of inflammation and oxidative stress while enhancing endogenous antioxidant enzyme activity [23,24].
2. Chemical Composition of Aronia melanocarpa
Aronia melanocarpa possesses a complex and well-defined chemical composition that includes a wide spectrum of bioactive ingredients. Its fruits and pomace are particularly rich in polyphenolic compounds, such as anthocyanins (mainly cyanidin derivatives), proanthocyanidins, flavonols (including quercetin and kaempferol glycosides) and phenolic acids (notably chlorogenic, neochlorogenic and caffeic acids) [15,25,26]. These compounds are found in both free and bound forms and vary in concentration depending on the plant part (leaves, fruit, pomace) and processing method (Table 1).
Besides polyphenols, aronia contains significant levels of polysaccharides, pectins and dietary fiber, which are mostly retained in pomace after juice extraction [40,41]. The fruit matrix also includes organic acids (e.g., malic and citric acids), natural sugars (mainly glucose and fructose) and trace amounts of unsaturated fatty acids. Furthermore, aronia provides essential minerals, such as potassium, magnesium, calcium, phosphorus and iron, as well as small quantities of vitamins, particularly vitamin C and some B-complex vitamins [16].
2.1. Polyphenolic Compounds in Aronia melanocarpa: Types and Biological Activity
Polyphenols are a structurally diverse group of secondary plant metabolites known for their antioxidant, anti-inflammatory and antiproliferative activities, particularly relevant in the context of CRC chemoprevention [42]. This category includes subclasses such as flavonoids, phenolic acids, stilbenes, lignans and less commonly in food matrices, curcuminoids [43]. These compounds vary not only in their chemical structures but also in their concentration. Aronia melanocarpa is among the richest natural sources of polyphenols, whose presence and biological effects have been extensively documented.
Flavonoids, characterized by a C6-C3-C6 carbon skeleton comprising two phenyl rings connected via a heterocyclic pyran or pyrone ring. They are subclassified based on ring modifications such as hydroxylation, glycosylation, or methylation into flavonols, flavones, flavanols, isoflavones, anthocyanins and flavanones [43].
Flavonols, e.g., quercetin and kaempferol derivatives, occur in A. melanocarpa mainly as quercetin-3-O-glucoside, quercetin-3-galactoside, quercetin-3-rutinoside, quercetin-3-robinobioside, quercetin-3-vicianoside, isorhamnetin-3-O-glucoside and kaempferol-3-O-glucoside [23,44]. Concentrations in fruits range from 0.12 to 1.4 mg/g dry weight (DW), while in leaves they may reach 1.8–7.86 mg/g DW [16,27,36]. In addition, comparative studies of leaf extracts from aronia and other polyphenol-rich species have confirmed the presence of high levels of flavonoids, phenolic acids and ellagitannins, correlating with notable antibacterial activity against Gram-positive pathogens [45]. These flavonols have been implicated in modulation of signaling cascades such as Phospho Inositide 3-Kinase/Protein kinase B (PI3K/Akt) and p53, contributing to cell cycle arrest and apoptosis in CRC models. Additionally, quercetin has been shown to enhance the effectiveness of chemotherapeutic agents by inhibiting multidrug resistance transporters, including P-glycoprotein, through both decreased transporter function and reduced expression in various cancer cell models [46].
Flavanols (flavan-3-ols), including monomers such as (+)-catechin and (−)-epicatechin and their polymers (proanthocyanidins), are abundant in A. melanocarpa. The dominant polymeric procyanidins account for a substantial portion of the antioxidant capacity (e.g., ~66% of total polyphenols in fruits). Total flavanol content in chokeberry fruits can vary broadly; for example, polymeric procyanidins (mainly epicatechin units) dominate, with free epicatechin in lower amounts [16]. They exert strong radical-scavenging and metal-chelating effects, inhibit NF-κB signaling and downregulate pro-inflammatory cytokines (e.g., IL-6, TNF-α) [20]. Flavanols may also beneficially modulate gut microbiota composition and enhance SCFAs production [14].
Anthocyanins are water-soluble pigments responsible for the deep color of chokeberries. In Aronia melanocarpa, anthocyanins are the predominant polyphenolic group, accounting for up to 60% of the total phenolic content [17,44]. The main anthocyanins are cyanidin derivatives, particularly cyanidin-3-galactoside, also cyanidin-3-arabinose, cyanidin-3-glucoside and cyanidin-3-xyloside. In some studies, cyanidin-3-O-galactoside extract of aronia berries reach 917.31 mg/g DW total anthocyanins, after purification [47]. These compounds exhibit potent antioxidant activity, regulate inflammatory cytokines such as TNF-α, IL-6 and have been shown to induce apoptosis in CRC cell lines [48].
Phenolic acids, a major class of non-flavonoid polyphenols are structurally characterized by hydroxyl and carboxyl functional groups and are subdivided into two main categories: hydroxybenzoic acids and hydroxycinnamic acids [43]. Black chokeberry is particularly notable for its high phenolic acid content, with chlorogenic acid and neochlorogenic acid being the dominant representatives. Chlorogenic acid concentrations range from 0.8 to 1.5 mg per g fresh weight (FW), while neochlorogenic acid is typically present at levels of 0.4 to 0.7 mg/g FW [16]. These hydroxycinnamic acids are recognized for their strong antioxidant capacity, primarily through free radical scavenging and metal ion chelation. Furthermore, they exert chemopreventive effects by inducing phase II detoxification enzymes, such as glutathione S-transferase, which contributes to the protection of colonic epithelial cells against oxidative DNA damage and carcinogenesis [49].
Lignans are diphenolic compounds biosynthesized via the shikimate pathway. Upon ingestion, they are metabolized by gut microbiota into enterolignans, such as enterodiol and enterolactone, which exhibit phytoestrogenic, antioxidant and anticancer activities [50]. While Aronia melanocarpa is not generally considered a major source of lignans and most compositional studies do not report their presence in significant concentrations in fruit tissues [27], some research suggests that minor lignan components (such as secoisolariciresinol) may occur in small quantities and contribute to the overall antioxidant and anticancer potential of chokeberry extracts [25], but evidence remains limited.
There are also studies comparing the biological activities and synergistic effects of aronia extracts with other polyphenols, such as stilbenes and curcuminoids. Stilbenes are structurally defined by two aromatic rings connected via a two-carbon ethylene bridge, with resveratrol being the most prominent compound, widely recognized for its antioxidant, anti-inflammatory, and cardioprotective properties. Dietary sources of stilbenes include berries [51]. Although Aronia melanocarpa is frequently compared to resveratrol in antioxidant and antiplatelet activity studies, current phytochemical analyses do not confirm the presence of resveratrol or other stilbenes in chokeberry fruits in quantifiable amounts [25,52]. In functional assays, chokeberry extracts have shown greater inhibitory effects on platelet aggregation than resveratrol used as a positive control, but this refers to comparative bioactivity rather than endogenous stilbene content [53].
Curcuminoids (e.g., curcumin, demethoxycurcumin, bisdemethoxycurcumin) are a small class of polyphenolic compounds extracted primarily from Curcuma longa (turmeric). Curcumin recognized for its potent anti-inflammatory, antioxidant and chemopreventive properties, acting through mechanisms such as NF-κB inhibition and modulation of oxidative stress pathways [54]. These compounds are not naturally present in aronia; however, in nutraceutical formulations or synergy-based studies, chokeberry extracts are often combined with curcuminoids to target overlapping molecular pathways—particularly those involved in inflammation, oxidative stress regulation and tumor suppression—thus enhancing the overall protective effects [25].
Recent clinical evidence also supports the role of polyphenol-based interventions, including curcuminoids, in the prevention and adjunct treatment of CRC. According to López-Gómez and Uranga (2024), combinations of dietary polyphenols such as curcumin with other bioactive compounds can improve treatment outcomes by modulating gut microbiota, reducing pro-inflammatory cytokines and enhancing epithelial barrier integrity—mechanisms that align with those attributed to Black chokeberry [42].
2.2. Polysaccharides in Aronia melanocarpa
Aronia melanocarpa contains a complex and diverse polysaccharide profile that plays an important role in the nutritional and health-promoting properties of the fruit. The dietary fiber in aronia pomace (the residue after juice extraction) is dominated by cellulose, accounting for approximately 300–350 mg/g DM and hemicelluloses (about 150–200 mg/g DM), including polysaccharides such as arabinogalactans, galactomannans and xyloglucans [34,55]. Pomace also contains significant amounts of pectins, averaging 70–100 mg/g DM, which represent an important soluble fiber fraction [34].
Monosaccharide analysis revealed that the main monosaccharide components of aronia polysaccharides are: arabinose (20–28%), galactose (25–30%), glucose (10–15%), xylose (5–8%), along with smaller amounts of fucose, rhamnose, mannose and galactosamine [34]. The two main polysaccharide fractions obtained from aronia showed average molecular weights of approximately 51,089 Da and 21,214 Da, respectively. Both fractions exhibited low polydispersity index values, indicating that they were molecularly uniform and composed of relatively homogeneous polysaccharide molecules [56].
2.3. Synergy Between Polyphenols and Polysaccharides in Natural Products
Recent research has highlighted the importance of synergistic interactions between polyphenols and polysaccharides in natural products, where interactions enhance both the bioavailability and biological efficacy of bioactive compounds. Polyphenols such as anthocyanins, when complexed with polysaccharides like arabinogalactans or xyloglucans, exhibit increased chemical stability and solubility, improving their resistance to gastrointestinal degradation and facilitating intestinal absorption. Wang et al. (2024) demonstrated that anthocyanin-polysaccharide complexes prepared from chokeberry showed significantly enhanced antioxidant activity compared to free anthocyanins, with a 1:1 ratio producing the most favorable structural and functional properties. These complexes formed amorphous, smooth particles, suggesting non-covalent interactions—primarily hydrogen bonding and hydrophobic forces—underlying their stability [55]. In addition to improving antioxidant potential, these complexes influence host health through gut microbiota modulation [57].
Several studies have shown that polyphenol-polysaccharide rich chokeberry extracts promote beneficial microbial shifts, including increased abundance of Lactobacillus and Bifidobacterium and decreased levels of gut inflammation such as Sutterella and Escherichia [58]. These microbial changes are accompanied by increased production of SCFAs, such as butyrate, which play key roles in maintaining intestinal barrier function and regulating immune responses. Zhu Y et al. (2022) further demonstrated that chokeberry polyphenols can regulate lipid metabolism by modulating the glycerophospholipid metabolic pathway, offering potential benefits for cardiometabolic health [59]. According to Shahidi and Athiyappan (2025), the functional interplay between polyphenols and polysaccharides may be leveraged in the development of novel food systems and delivery platforms that enhance the therapeutic potential of natural bioactives [60]. Collectively, these findings underscore that the health benefits of natural products should be understood in the context of their molecular synergy, rather than as isolated constituents.
3. Action of Polyphenols in Chemoprevention of CRC
3.1. Antioxidant and Anti-Inflammatory Properties
Polyphenols extracted from black chokeberry exhibit strong biological properties in the chemoprevention of CRC. One of the key mechanisms is their ability to neutralize reactive oxygen species (ROS), which are known to cause oxidative DNA damage and initiate carcinogenesis. Aronia polyphenols—particularly anthocyanins and proanthocyanidins—act as radical scavengers, chelate pro-oxidant metals and inhibit ROS-producing enzymes such as NADPH oxidase and cyclooxygenase [55,61]. Additionally, they upregulate the Nrf2/ARE pathway, leading to increased expression of endogenous antioxidant enzymes like superoxide dismutase (SOD) and glutathione peroxidase (GPx), thereby reinforcing cellular antioxidant defenses [37,40,55]. Owczarek et al. (2022) studies evaluated the phenolic composition, antioxidant and cytotoxic activity of A. melanocarpa leaf extracts, showing selective cytotoxicity toward cancer cell lines associated with significant ROS modulation [62]. This provides evidence that not only fruit extracts but also leaf-derived polyphenols can contribute to oxidative stress regulation in pathological contexts [62].
In addition to their antioxidant role, Aronia melanocarpa polyphenols exert potent anti-inflammatory effects [63]. They suppress the expression of key pro-inflammatory mediators such as TNF-α, IL-6 and inducible nitric oxide synthase (iNOS) by downregulating signaling pathways like NF-κB and MAPK [64,65]. Ultrasound-assisted extracts from aronia pomace have demonstrated efficacy in reducing inflammation in vitro, supporting their therapeutic relevance in CRC [64]. Also, valorization of aronia pomace as a by-product has been explored. Lazăr et al. (2020) characterized pomace fractions for functional ingredient development, confirming retained antioxidant capacity and polyphenol potential [66]. Furthermore, encapsulation strategies using hemicellulose matrices have shown promise in enhancing the bioactivity and targeted delivery of polyphenols to inflamed intestinal tissues, thereby improving their potential for managing inflammatory bowel disease and CRC [67].
Interestingly, recent studies suggest that the biological activity of aronia polyphenols may be dose-dependent. While moderate concentrations boost cellular antioxidant capacity, higher doses in cancer cells can paradoxically induce ROS-mediated apoptosis, selectively eliminating malignant cells [61]. Notably, comparative phytochemical profiling reveals that aronia fruits contain significantly higher levels of polyphenols and antioxidant potential than many commonly consumed berries and fruits, highlighting their unique value in functional food and nutraceutical approaches to CRC prevention [37,68].
3.2. Modulation of Apoptotic Pathways in CRC Cells
Polyphenols have significant anticancer effects in CRC cells by modulating key apoptotic pathways. One primary mechanism involves the activation of caspase-dependent apoptosis, particularly caspase-3, -8 and -9, which leads to programmed cell death in tumor cells [61,69]. Coinciding, polyphenols influence the balance between pro-apoptotic (e.g., Bax, Bad) and anti-apoptotic proteins (e.g., Bcl-2, Bcl-xL), often tipping the scale toward cell death by promoting mitochondrial membrane permeabilization and cytochrome c release [55].
Specifically, aronia polyphenols—especially anthocyanins and proanthocyanidins—have been shown to downregulate Bcl-2 and upregulate Bax in CRC cells, thereby sensitizing them to apoptosis [40]. This modulation is often mediated via upstream signaling pathways, including the inhibition of PI3K/Akt and NF-κB, which are commonly upregulated in colorectal tumors and linked to cell survival and chemoresistance [61,69,70]. Moreover, some studies report that high concentrations of polyphenols can induce ROS-mediated apoptosis, selectively damaging cancer cells without affecting normal colonocytes, suggesting dual antioxidant/pro-oxidant behavior depending on context and dosage [55,61].
3.3. Inhibition of Cancer Cell Proliferation and Metastasis
Polyphenols from natural dietary sources demonstrate strong antiproliferative and antimetastatic effects against CRC cells by modulating key molecular signaling pathways involved in tumor progression (Table 2).
Studies on anthocyanin-rich extracts from Aronia melanocarpa fruit have shown suppression of CRC cell growth via downregulation of Cyclin D1, activation of cell cycle inhibitors p21 and p27 and inhibition of PI3K/Akt and MAPK/ERK signaling cascades, which are critical for tumor cell survival and proliferation [55,61]. Additionally, their fruit polyphenols downregulate NF-κB and STAT3, two key regulators of inflammatory and proliferative responses in colorectal carcinogenesis [61].
Crucially, aronia extracts also target mechanisms involved in tumor metastasis. The suppression of matrix metalloproteinases MMP-2 and MMP-9, enzymes essential for extracellular matrix degradation and cancer cell invasion, has been observed following treatment with aronia polyphenols [40]. These compounds also interfere with epithelial–mesenchymal transition (EMT) by increasing the expression of E-cadherin and suppressing mesenchymal markers such as Snail and ZEB1, thereby limiting cancer cell motility and invasiveness [72].
Beyond fruit-derived compounds, recent research has highlighted the anti-metastatic activity of aronia leaf extracts, which similarly reduce CRC cell migration and invasion. This activity has been linked to reduced MMP activity and modulation of EMT-related genes, providing additional evidence for the therapeutic potential of different aronia plant parts [24].
4. Role of Polysaccharides in CRC Prevention
4.1. Influence on Gut Microbiota and Its Role in CRC Prevention
Recent advances have reinforced the central role of dietary polysaccharides in preventing CRC through modulation of the gut microbiota. Carbohydrates complex serve fermentable substrates for beneficial bacteria such as Bifidobacterium, Lactobacillus and Roseburia, promoting the production of SCFAs, especially butyrate [73,74]. Butyrate, in turn, exerts anti-inflammatory and anti-proliferative effects in colonic epithelial cells, enhancing epithelial barrier integrity, modulating immune responses and inducing apoptosis in dysplastic or cancerous cells [14,73]. A review in the British Journal of Nutrition further emphasizes butyrate’s role in CRC prevention by inhibiting histone deacetylases, reducing pro-inflammatory signaling and downregulating genes involved in tumorigenesis [75]. Moreover, recent findings by Liu et al. (2024) suggest that natural polysaccharides can reshape the gut microbial landscape in favor of anti-tumor immunity, reduce the abundance of pathogenic species and support the efficacy of immunotherapy. These multifaceted microbial and immunomodulatory effects underscore the therapeutic relevance of polysaccharides as functional dietary components in CRC prevention strategies [76].
4.2. Anti-Inflammatory and Antioxidant Properties of Polysaccharides
Dietary polysaccharides derived from natural plant sources, often coexisting with polyphenolic compounds such as kaempferol, epicatechin, resveratrol and curcumin, exhibit potent anti-inflammatory and antioxidant properties, making them promising candidates for the prevention and management of chronic intestinal inflammation—a recognized risk factor in the pathogenesis of CRC. These polysaccharide-polyphenol complexes synergistically modulate inflammatory signaling by inhibiting the activation of NF-κB, a key transcription factor involved in the expression of pro-inflammatory cytokines including IL-6 and TNF-α [77,78].
Simultaneously, polysaccharides contribute to oxidative stress reduction by scavenging ROS, stabilizing redox homeostasis and enhancing the activity of endogenous antioxidant enzymes such as SOD and GPx. These mechanisms not only prevent oxidative DNA damage but also support the maintenance of mucosal integrity and attenuate inflammation-induced cytotoxicity in colonic epithelial cells [79]. Findings from in vivo studies corroborate these effects, where supplementation with plant-derived polysaccharides has been shown to reduce inflammatory cell infiltration, preserve epithelial barrier function, and restore intestinal homeostasis [80]. These data underscore the chemopreventive potential of polysaccharide-rich plant extracts, particularly those enriched with complementary phytochemicals, in reducing the burden of CRC through dual modulation of oxidative and inflammatory pathways.
4.3. Potential to Inhibit CRC-Related Pathways
Beyond their well-documented anti-inflammatory and antioxidant actions, dietary polysaccharides derived from plants exhibit a multifaceted potential to inhibit molecular pathways directly involved in CRC development and progression. Emerging evidence from recent reviews and experimental studies demonstrates that these polysaccharides can suppress CRC cell proliferation by modulating several oncogenic signaling cascades critical for tumor growth and survival. Notably, polysaccharides have been shown to downregulate the PI3K/Akt pathway, which is often hyperactivated in CRC and contributes to enhanced cell proliferation, survival and chemoresistance [81]. Additionally, they interfere with the IL-6/STAT3 axis, a key driver of chronic inflammation-induced tumorigenesis, thereby reducing pro-survival and pro-inflammatory gene expression that promotes tumor progression.
Polysaccharides also modulate mTOR signaling, a central regulator of cellular metabolism and growth, thus impairing cancer cell anabolism and proliferation. The inhibition of TLR4/JNK pathways further contributes to their anti-tumor effects by suppressing inflammatory responses and inducing apoptosis in cancer cells. These compounds can promote programmed cell death through both apoptosis and autophagy, enhancing the elimination of malignant cells [81]. Importantly, polysaccharides influence non-coding RNAs, including microRNAs and long non-coding RNAs, which regulate intrinsic cell death mechanisms and tumor suppressor gene expression, highlighting a complex layer of epigenetic regulation in CRC chemoprevention.
Moreover, the ability of polysaccharides to inhibit metastatic processes has been linked to the disruption of Smad2/3 and IRS1 signaling, pathways implicated in EMT and cellular migration. By impeding these pathways, polysaccharides reduce tumor invasiveness and dissemination, which are critical steps in CRC progression and patient prognosis [81].
5. Polyphenol–Polysaccharide Complexes from Aronia melanocarpa: Synergistic Effects
5.1. Enhancement of Bioavailability and Stability of Active Compounds
Anthocyanins derived from Aronia melanocarpa exhibit significant therapeutic potential; however, their poor stability and low bioavailability under physiological and environmental conditions limit clinical application. Recent strategies based on polysaccharide complexation have demonstrated promising results in enhancing anthocyanin stability. For instance, the use of carboxymethyl cellulose (CMC), pectin and xanthan gum (XG) enabled the formation of stable anthocyanin-polysaccharide complexes through hydrogen bonding, cation-π interactions and electrostatic forces. These complexes markedly improved resistance to heat, light, oxidation, and metal ions, while significantly enhancing antioxidant activity-especially with XG, which also improved bile salt-binding and tyrosinase inhibition [82].
Further advancements include encapsulation techniques using maltodextrin combined with polysaccharides such as CMC, XG, or gum arabic. These formulations achieved encapsulation efficiencies of 94–98%, with anthocyanin retention reaching 88–91% after 100 days of storage at 25 °C, compared to only 47% in non-encapsulated samples. Notably, cyanidin-3-galactoside and cyanidin-arabinosides demonstrated higher stability than their glucoside and xyloside counterparts [83].
A particularly promising approach involves amylopectin nanoparticle delivery systems, which bind anthocyanins more than 80% efficiency. This delivery system increased antioxidant retention, protected anthocyanins from digestive degradation and enhanced intestinal absorption [84]. A subsequent pharmacokinetic study confirmed that amylopectin encapsulation improved oral bioavailability by 440–593%, prolonged plasma retention time and reduced excretion of unmetabolized anthocyanins, demonstrating effective sustained release [85]. Collectively, these strategies represent advanced delivery systems that significantly improve the functional performance of Aronia melanocarpa anthocyanins in food, pharmaceutical and nutraceutical formulations.
5.2. Combined Action on CRC Prevention and Synergistic Effects
The integration of Aronia melanocarpa-derived polyphenols and polysaccharides reveals compelling synergistic mechanisms in CRC prevention. Specifically, Li et al. (2024) demonstrated that the combination of Aronia Berry Extract (ABE) with oligomeric proanthocyanidins (OPCs) significantly reduced CRC cell viability and colony formation, while amplifying apoptosis in vitro and in 3D organoid models. This effect was mechanistically traced to downregulation of lamin B1 (LMNB1) and subsequent suppression of AKT phosphorylation, emphasizing multi-targeted pathway modulation more effective than with either agent alone [86].
Complementing this, Wei et al. (2020) showed that anthocyanins from aronia induce apoptosis in Caco-2 cells by inhibiting the Wnt/β-catenin pathway—specifically, reducing cytoplasmic β-catenin levels, arresting the cell cycle and triggering apoptotic processes [87].
Although anthocyanin–polysaccharide complexes from aronia have not yet been directly tested in CRC models, Wang et al. (2024) provided strong evidence that such complexes substantially enhance anthocyanin stability, antioxidant activity and resistance to degradation—all essential precondition for effective in vivo action. This improved stability likely prolongs anthocyanin exposure to colon cells, thereby strengthening inhibition of oncogenic pathways like Wnt/β-catenin [55].
Altogether, these findings indicate a synergistic interplay: polyphenol-polyphenol combinations (e.g., ABE + OPCs) efficiently suppress oncogenic signaling (LMNB1-AKT), while polysaccharide stabilization enhances persistence and bioactivity of anthocyanins in the colon, permitting more effective engagement of apoptotic and anti-proliferative mechanisms [86,88].
Figure 1 summarizes supporting evidence explaining how polyphenol–polysaccharide complexes from Aronia melanocarpa enhance polyphenol bioavailability and modulate the gut microbiota in the prevention of CRC.
6. In Vitro and In Vivo Studies on Aronia melanocarpa in CRC Chemoprevention
6.1. In Vitro Models: Impact on CRC Cell Lines and Tumor Markers
Cell culture studies have demonstrated that extracts from black chokeberry, rich in polyphenols such as anthocyanins, flavanols and phenolic acids, significantly suppress the proliferation of colorectal cancer cell lines such as Caco-2, HT-29 and SW-480 by inducing apoptosis and causing cell cycle arrest. Bermúdez-Soto et al. (2007) reported that prolonged exposure of Caco-2 cells to chokeberry juice led to the upregulation of the tumor suppressor protein, contributing to reduced cancer cell growth [89]. Likewise, Wei et al. (2020) demonstrated that anthocyanins from Aronia melanocarpa suppress the Wnt/β-catenin signaling pathway, resulting in decreased cytoplasmic β-catenin levels and downregulation of pro-proliferative target genes, thereby enhancing cancer cell apoptosis [87].
Further research by Cvetanović et al. (2018) revealed that among different plant parts, leaf extracts showed the strongest anti-metastatic properties by inhibiting cancer cell migration and invasion through downregulation of matrix metalloproteinases MMP-2 and MMP-9, which are essential for extracellular matrix degradation [90]. Similarly, Gill et al. (2021) compared various chokeberry species and confirmed that black chokeberry extract had the highest antiproliferative activity against HT-29 cells, correlating with its high total phenolic content and antioxidant potential [18]. These findings underscore the importance of specific polyphenolic constituents (especially chlorogenic and caffeic acids) in mediating aronia’s anticancer properties [20].
Complementing these earlier studies, Kulling and Rawel (2008) showed that flavanol-rich chokeberry extracts induce cell cycle arrest and apoptosis in CRC cells, while also modulating the expression of genes associated with cellular survival and oxidative stress response [28].
Recent findings by Owczarek et al. (2022) further support these conclusions. In their study, two types of aronia leaf extracts—crude (ACE) and purified (APE)—were tested on CRC cell lines SW-480 and HT-29. The APE extract demonstrated significantly stronger cytotoxic effects in a dose and time-dependent manner, with an IC_50_ of approximately 194 µg/mL for SW-480 cells after 48 h of exposure. Notably, these effects were selective, showing minimal toxicity toward normal colon epithelial cells (CCD 841 CoN), suggesting potential therapeutic relevance. The enhanced activity of APE was attributed to its enriched content of phenolic acids and antioxidant compounds, which likely play a central role in modulating tumor cell viability, inflammation and oxidative stress [62].
In 2024, Stanca and his colleagues provided additional evidence for the anticancer potential of chokeberry by demonstrating that polyphenol and anthocyanin-rich pomace extracts from Aronia melanocarpa significantly reduced the viability of CRC cells (C2BBe1) in vitro. Their study revealed that chokeberry extracts exert cytotoxic effects in a dose-dependent manner, modulate key signaling pathways such as Akt and Erk1/2, and downregulate the expression of pro-inflammatory cytokines and matrix metalloproteinases (MMP-1, MMP-2, MMP-3), indicating that their action involves both antiproliferative and anti-inflammatory mechanisms [91].
6.2. Animal Model Studies
In vivo studies provide additional support for earlier in vitro findings. Valcheva-Kuzmanova and colleagues provide compelling evidence of the broad-spectrum health-promoting properties of Black chokeberry, particularly in the context of gastrointestinal disorders and metabolic dysfunctions. Investigations demonstrated potential of aronia juice against indomethacin-induced gastric mucosal damage, primarily through suppression of oxidative stress and upregulation of endogenous antioxidant enzymes such as SOD and catalase [92,93,94]. More recent studies using a 2,4,6-Trinitrobenzenesulfonic acid (TNBS)-induced colitis model in rats revealed that aronia juice ameliorates the symptoms of IBD by reducing inflammatory cytokine expression, improving histopathological features of the colon and restoring redox homeostasis [94].
More recent research by Zhao et al. (2021) and Zhu et al. (2022) revealed that aronia-derived polysaccharides positively modulate the gut microbiota by promoting the growth of Bacteroides species and increasing SCFAs production. These SCFAs enhance intestinal barrier integrity and regulate inflammatory pathways, notably through AMPK/SIRT1/NF-κB signaling, which are crucial in CRC pathogenesis [41,59]. Similarly, Kaczmarczyk et al. (2025) reviewed animal and human studies confirming that A. melanocarpa supplementation reduces oxidative stress and inflammation while improving gut microbiota composition [24].
Additional in vivo models of colitis induced by dextran sodium sulfate or TNBS have demonstrated that aronia extracts reduce pro-inflammatory cytokines (e.g., TNF-α, IL-1β), oxidative stress markers (MDA) and restore endogenous antioxidant enzymes such as catalase and GPx [95,96]. Moreover, these treatments enhanced the expression of tight junction proteins (ZO-1, occludin, claudin-1), thereby improving intestinal barrier function and elevating SCFAs levels, further supporting gut homeostasis [97].
T-cell transfer-induced colitis models provided additional mechanistic insights, showing that dietary supplementation with 4.5% aronia powder attenuated colonic inflammation by reducing TNF-α and IFN-γ levels, lowering MDA and preserving antioxidant defenses (GPx, reduced glutathione, Nfe2l2) in intestinal tissues and associated lymph nodes [98]. Notably, aronia fruit juice demonstrated superior efficacy to sulfasalazine in TNBS-induced colitis rats by significantly ameliorating clinical symptoms and decreasing thiobarbituric acid reactive substances, confirming its potent antioxidant and anti-inflammatory properties [94].
Complementary in vitro experiments with anthocyanin-rich aronia extracts further support these findings by demonstrating suppression of inflammatory mediators IL-1β, TNF-α and MDA in Lipopolysaccharide-stimulated macrophages without cytotoxicity [40]. Collectively, these animal model studies elucidate multiple molecular mechanisms through which aronia exerts therapeutic effects, including modulation of inflammatory signaling (e.g., NF-κB pathway), reduction in oxidative damage, enhancement of gut barrier integrity and beneficial alteration of microbiota composition.
6.3. Clinical Trials on Aronia melanocarpa in Cancer Prevention
Clinical and preclinical studies based on Aronia melanocarpa extracts have explored its health benefits across various populations, including healthy individuals, athletes, and those with metabolic disorders or cardiovascular conditions. Early clinical evidence demonstrated significant anti-inflammatory effects of aronia supplementation in high-risk populations. In a 6-week placebo-controlled trial, Naruszewicz et al. (2007) found that daily intake of 255 mg of aronia extract significantly reduced high-sensitivity C-reactive protein (hs-CRP) and IL-6 in post-myocardial infarction patients [99]. Similarly, Broncel et al. (2010) reported that 8 weeks of supplementation with 300 mg/day of extract in 25 patients with metabolic syndrome did not significantly alter CRP levels but improved the activity of endogenous antioxidant enzymes such as SOD and GPx, while CAT activity decreased, highlighting enzyme-specific responses [100].
In healthy populations, the effects of aronia are more modest but still evident. A 12-week single-arm study by Kardum et al. (2014) showed increased SOD and GPx activity in 29 healthy women consuming 100 mL of chokeberry juice daily [101]. In hypertensive individuals, a 4-week intervention with 200 mL/day of aronia juice led to a significant reduction in blood pressure and improvements in lipid parameters [102]. Findings in diabetic populations have been more variable. Milutinović et al. (2019) reported that a 12-week intervention with 150 mL/day of juice in type 2 diabetes patients led to improvements in LDL cholesterol, HbA1c, and select hematological parameters, suggesting potential benefits for metabolic and cardiovascular health [103].
More recent trials have strengthened the evidence for aronia’s bioactivity, particularly in metabolically compromised individuals. Tasić et al. (2021) demonstrated that standardized extract for four weeks in 143 participants with metabolic syndrome significantly improved blood pressure, glucose regulation, and lipid profile key metabolic disturbances associated with increased CRC risk [104].
Gancheva et al. (2021), in a 12-week parallel trial in overweight adults, found reduced CRP and increased SOD activity, indicating a decrease in systemic inflammation and oxidative stress. Catalase activity remained largely unchanged, suggesting that aronia’s antioxidant effects may be selective for specific enzymatic pathways [105]. In contrast, Milosavljević et al. (2021) investigated the effects of one-month supplementation with 30 mL/day of a standardized Aronia melanocarpa extract in 30 anemic hemodialysis patients. They observed a significant increase in CAT and reduced glutathione levels, along with a decrease in SOD anion, but systemic inflammation was not significantly improved [106].
In physically active individuals, who often experience exercise-induced oxidative and inflammatory stress, aronia also shows potential. Two studies by Stankiewicz et al. (2021, 2023) in male athletes investigated the effects of 7- and 12-week supplementation with either chokeberry juice (200 mL/day) or freeze-dried extract (6 g/day). In the 2023 study, supplementation led to reductions in the pro-inflammatory cytokine IL-6 and increases in the anti-inflammatory cytokine IL-10. However, this effect was not observed in the earlier 2021 trial [107,108].
Chung et al. (2023), in an 8-week trial in healthy individuals following exercise, found increased GPx activity without significant changes in IL-6 or catalase, suggesting antioxidant benefits even without overt inflammation reduction [109]. In populations with low baseline inflammation, results have been mixed. A 90-day randomized crossover trial by Sangild et al. (2023) in hypercholesterolemic adults receiving 150 mg/day of an anthocyanins standardized Aronia spp. extract reported a significant increase in glutathione levels and improvement in cytoprotective targets in specific subgroups of men over 40 years of age [110].
Among the most recent findings, Chamberlin et al. (2024) conducted a 30-day randomized, placebo-controlled trial in 14 healthy adults, showing that daily intake of 100 mL of polyphenol-rich aronia juice significantly reduced postprandial glucose (β = −3.03, p < 0.01), prevented increases in fasting total cholesterol (β = −0.50, p = 0.03), and modulated both serum and gut metabolites involved in central carbon and lipid metabolism. Importantly, aronia consumption was associated with decreased levels of pro-inflammatory metabolites, underscoring its systemic anti-inflammatory effects [22].
To consolidate the growing body of evidence, Sarıkaya et al. (2025) conducted a systematic review of 18 randomized controlled trials published between 2022 and 2023. The review evaluated interventions using aronia juice, extract and also over-dried powder. The results confirm consistent reductions in inflammatory markers such as TNF-α, IL-6, and CRP, as well as significant enhancements in antioxidant enzyme activity (SOD, CAT, GPx) across diverse populations [23]. These findings are consistent with broader clinical evidence on the role of polyphenol-rich foods as summarized in a recent systematic review by López-Gómez & Uranga (2024). Dietary polyphenols, including those found in Aronia melanocarpa, have shown promising results in the prevention and adjunctive treatment of CRC, primarily through anti-inflammatory pathways, modulation of gut microbiota, and maintenance of epithelial barrier integrity [42]. Furthermore, a meta-analysis by Xu et al. (2021) demonstrates that dietary polyphenol supplementation significantly affects iron metabolism and erythropoiesis across diverse populations, suggesting that polyphenols can modulate physiological parameters relevant to cancer risk in a context-dependent manner [111].
7. Study Limitations and Future Perspectives
Aronia melanocarpa is a promising functional food ingredient due to its exceptionally high content of polyphenols, including anthocyanins, proanthocyanidins, and phenolic acids. However, its practical application faces some challenges. One of the main considerations is the relatively low bioavailability of these compounds, which may limit their biological efficacy in humans. Many polyphenols are only partially absorbed in the gastrointestinal tract and subsequently metabolized or degraded during processing and storage. Encapsulation strategies, such as binding anthocyanins to amylopectin nanoparticles, enhance systemic bioavailability in vivo [82,84,85].
Another important factor is the variability of bioactive composition in the raw material—the polyphenol profile can differ depending on the aronia cultivar, cultivation conditions, fruit maturity, and extraction methods, making it difficult to standardize preparations and compare clinical study results [30,37].
Technological and nutritional aspects can also affect their practical use: Aronia extracts are naturally astringent and bitter, which may influence consumer acceptance in food products. Strategies such as microencapsulation or freeze-drying can improve the stability of phenolic compounds and their bioavailability, though they may add complexity to production and formulation [67,112].
Despite strong mechanistic support, meta-analyses of randomized human trials suggest that clinical evidence for some of aronia’s health benefits remains limited or variable. For example, effects on cardiometabolic markers are promising but not always consistent, with many studies featuring small sample sizes, short intervention periods, and differences in dosage. Regulatory aspects also need consideration, as the classification of Aronia extracts—whether as a functional food, dietary supplement, or medicinal ingredient—can influence requirements for clinical testing, labeling, and health claims [23,42].
In terms of safety, Aronia melanocarpa is generally well tolerated. Nevertheless, some aspects deserve attention, particularly regarding long-term or high-dose intake. The polyphenols and tannins naturally present in chokeberry may modestly reduce non-heme iron absorption, an effect supported by studies and meta-analyses of polyphenol supplementation [113,114]. While this is unlikely to be clinically significant for most healthy adults, it may be relevant for populations at risk of iron deficiency.
Additionally, the literature indicates that some polyphenols exhibit prooxidant activity at high concentrations, resulting in increased production of reactive oxygen species and induction of apoptosis in cancer cells. Mechanistically, this effect may result from autoxidation of phenolic compounds, redox cycling involving transition metals, and inhibition of cellular antioxidant systems [115]. It is essential to note that most reports of prooxidant activity originate from in vitro studies conducted at concentrations significantly higher than those typically achievable after oral administration (i.e., studies ranging from tens to hundreds of µM or even mM), whereas plasma concentrations following dietary polyphenol consumption are generally in the µM range or lower [116,117]. Furthermore, in vivo factors such as hepatic metabolism, plasma protein binding, and transformation by gut microbiota significantly influence the biological activity and availability of free polyphenols. This phenomenon can be considered in the context of hormesis: low doses of polyphenols often exhibit cytoprotective and antioxidant effects, whereas higher doses can induce oxidative stress and apoptosis [62]. This has two practical implications. First, translational research should carefully assess the transfer of results from in vitro models to in vivo and then to humans; second, prooxidant effects at high doses may have therapeutic applications (e.g., targeted destruction of cancer cells) but also carry the risk of toxicity to normal cells and potential drug interactions (e.g., during chemotherapy). Therefore, detailed pharmacokinetic and toxicological studies are necessary to establish safe exposure ranges and potential dose or concentration thresholds for different preparations and clinical settings [113,118].
Finally, mild gastrointestinal effects, such as temporary astringency or discomfort, have occasionally been reported in studies using higher doses of chokeberry products [119]. Since most clinical trials have been relatively short (<12 weeks), future studies would benefit from monitoring long-term safety and potential impacts on micronutrient status.
8. Conclusions
Aronia melanocarpa (Michx.) Elliot exhibits multifaceted health-promoting properties, particularly in the context of inflammation-associated disorders such as CRC. Its berries contain high levels of anthocyanins (e.g., cyanidin-3-galactoside, cyanidin-3-arabinoside), phenolic acids (notably chlorogenic acid) and flavonoids such as quercetin and epicatechin, all of which exhibit strong antioxidant, anti-inflammatory and antiproliferative effects through mechanisms such as NF-κB inhibition, induction of apoptosis and modulation of gene expression. In addition, aronia polysaccharides (e.g., pectins, arabinogalactans) support gut microbiota by promoting beneficial taxa such as Bacteroides, increasing SCFAs production and strengthening intestinal barrier integrity. Together, these synergistic actions help reduce oxidative stress, inflammation and epithelial permeability—key features of CRC pathogenesis.
Polyphenols and polysaccharides thus complement one another: polyphenols reduce oxidative and inflammatory processes, while polysaccharides improve microbiota composition, thereby enhancing polyphenol metabolism and bioavailability, and resulting in a mutually reinforcing anti-inflammatory and anti-tumorigenic effect relevant to CRC prevention.
Beyond fresh fruits, Aronia by-products such as pomace (pressing residue) offer an underutilized but potent source of polyphenols and fibers. Pomace retains a significant fraction of anthocyanins and proanthocyanidins (up to 40% of total antioxidant activity) and exhibits superior radical scavenging ability compared to many other fruit-based matrices. Seasonal harvesting and juice processing generate large amounts of pomace, making it an economically attractive raw material for functional foods, nutraceuticals and pharmaceutical formulations. Leaf extracts, although less studied, have also shown antioxidant and antiproliferative activity, offering an additional seasonal fraction with potential biomedical value. These characteristics suggest clear practical applications: pomace and leaves can be incorporated into fiber-enriched formulations, polyphenol-rich dietary supplements, or functional food matrices designed to modulate inflammation and support gut health, thereby offering translational potential for CRC chemoprevention.
However, data specifically evaluating aronia pomace in CRC prevention are still lacking, highlighting a critical research gap. Bioavailability also presents a challenge, as polyphenols are susceptible to degradation during processing and digestion. Recent studies suggest that encapsulation technologies (e.g., zein nanoparticles, spray-drying) may help preserve their stability and activity throughout the gastrointestinal tract.
Aronia melanocarpa is generally recognized as safe and well tolerated, although some considerations regarding long-term or high-dose intake should be noted. Polyphenols and tannins naturally occurring in chokeberry may modestly reduce non-heme iron absorption, which could be relevant for populations at risk of iron deficiency, though clinically significant effects in healthy individuals are unlikely. Occasional mild gastrointestinal discomfort has been reported in studies using higher doses of chokeberry products, and potential interactions with drug-metabolizing enzymes remain theoretical, as no serious adverse events have been documented. Including routine safety monitoring in future long-term intervention trials would strengthen the evidence base for aronia supplementation.
In summary, the combination of potent phytochemicals, microbiota-modulating polysaccharides, and a growing body of scientific evidence indicates that Aronia melanocarpa as a promising dietary component for the prevention and potentially as an adjunctive treatment of CRC and other inflammation-related malignancies. The valorization of pomace and leaves enhances year-round applicability and supports sustainable production of bioactive-rich materials for health-oriented purpose applications. While its safety profile appears favorable, further research is warranted to confirm long-term tolerability, optimize polyphenol bioavailability, and conduct more rigorous and extended human trials to translate preclinical results into measurable health benefits.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sung H. Ferlay J. Siegel R.L. Laversanne M. Soerjomataram I. Jemal A. Bray F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries CA Cancer J. Clin.20217120924910.3322/caac.2166033538338 · doi ↗ · pubmed ↗
- 2Morgan E. Arnold M. Gini A. Lorenzoni V. Cabasag C.J. Laversanne M. Vignat J. Ferlay J. Murphy N. Bray F. Global burden of colorectal cancer in 2020 and 2040: Incidence and mortality estimates from GLOBOCAN Gut 20237233834410.1136/gutjnl-2022-32773636604116 · doi ↗ · pubmed ↗
- 3World Cancer Research Fund American Institute for Cancer Research Diet, Nutrition, Physical Activity and Colorectal Cancer World Cancer Research Fund International London, UK 2018 Available online: https://www.wcrf.org/dietandcancer/colorectal-cancer/(accessed on 2 October 2025)
- 4Zheng J. Zhao L. Dong J. Giovannucci E. Development and validation of Comprehensive Inflammatory Lifestyle Score and association with colorectal cancer risk Cancer Epidemiol. Biomark. Prev.2025341810181910.1158/1055-9965.EPI-25-038640704925 · doi ↗ · pubmed ↗
- 5Abdalla M. Eberhardson M. Landerholm K. Andersson R.E. Myrelid P. Impact of inflammatory bowel disease and primary sclerosing cholangitis on colorectal cancer risk: National cohort study Clin. Gastroenterol. Hepatol.2025 in press 10.1016/j.cgh.2025.06.03740706734 · doi ↗ · pubmed ↗
- 6Iacucci M. Nardone O.M. Ditonno I. Capobianco I. Pugliano C.L. Maeda Y. Majumder S. Zammarchi I. Santacroce G. Ghosh S. Advancing inflammatory bowel disease-driven colorectal cancer management: Molecular insights and endoscopic breakthroughs towards precision medicine Clin. Gastroenterol. Hepatol.2025 in press 10.1016/j.cgh.2025.06.03540706730 · doi ↗ · pubmed ↗
- 7Dekker E. Tanis P.J. Vleugels J.L.A. Kasi P.M. Wallace M.B. Colorectal cancer Lancet 20193941467148010.1016/S 0140-6736(19)32319-031631858 · doi ↗ · pubmed ↗
- 8Arnold M. Sierra M.S. Laversanne M. Soerjomataram I. Jemal A. Bray F. Global patterns and trends in colorectal cancer incidence and mortality Gut 20176668369110.1136/gutjnl-2015-31091226818619 · doi ↗ · pubmed ↗