A High-Multiplicity Baculovirus Method Enables Efficient Gene Delivery to Diverse Mammalian Cells In Vitro and to Multiple Organs In Vivo
Min-Hsiu Wu, Song-Tay Lee, Tsung-Hsien Chang, Wei-Sheng Chao, Nan-Kai Lin, Shoa-Lin Lin

TL;DR
A new high-multiplicity baculovirus method efficiently delivers genes to various mammalian cells in the lab and multiple organs in live animals.
Contribution
The study introduces a recombinant baculovirus vector that enables efficient and safe gene delivery in vitro and in vivo at high multiplicities.
Findings
vAcMBac-CMV-IE-EGFP achieved high transduction rates in Sf9, HepG2, and Vero E6 cells.
In vivo, EGFP expression was observed in multiple organs and neural tissues of rats.
High MOI did not significantly affect cell viability in vitro.
Abstract
Recombinant baculovirus vectors are recognized as effective gene delivery systems for mammalian cells in vitro. However, their application in vivo has been limited due to inactivation by the host’s complement system. We developed a recombinant baculoviral vector derived from Autographa californica multiple nucleopolyhedrovirus (AcMNPV), incorporating both CMV-IE and polyhedron promoter-driven green fluorescence protein (EGFP) (vAcMBac-CMV-IE-EGFP). We then evaluated the transduction efficiency and safety of vAcMBac-CMV-IE-EGFP at a high multiplicity of infection (MOI) across five distinct cell lines and in Sprague Dawley (SD) rats. In vitro, Sf9, HepG2, and Vero E6 cells showed high transduction rates (95.52 ± 4.86%, 80.53 ± 3.31%, and 80.87 ± 2.50%, respectively), significantly outperforming the other cell types tested, and cell viability remained largely unaffected even at an MOI of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
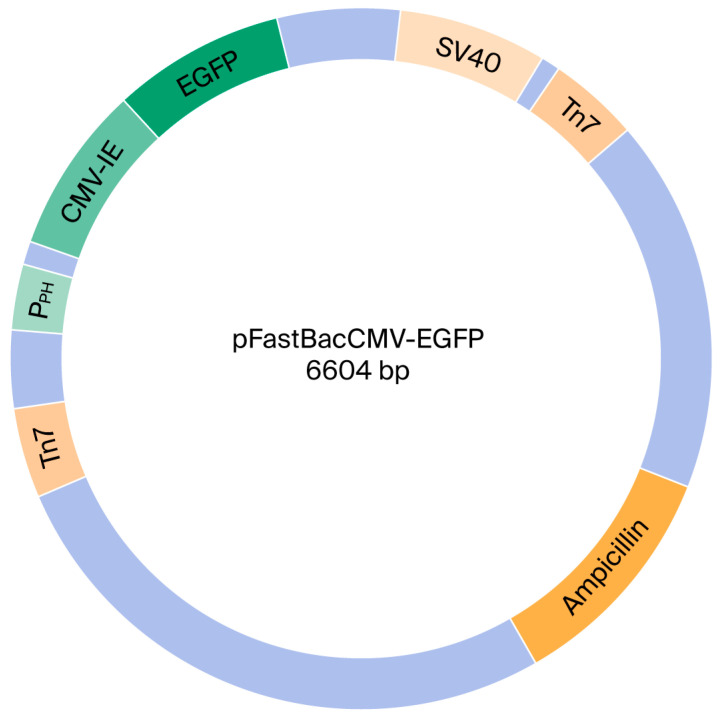 Figure 1
Figure 1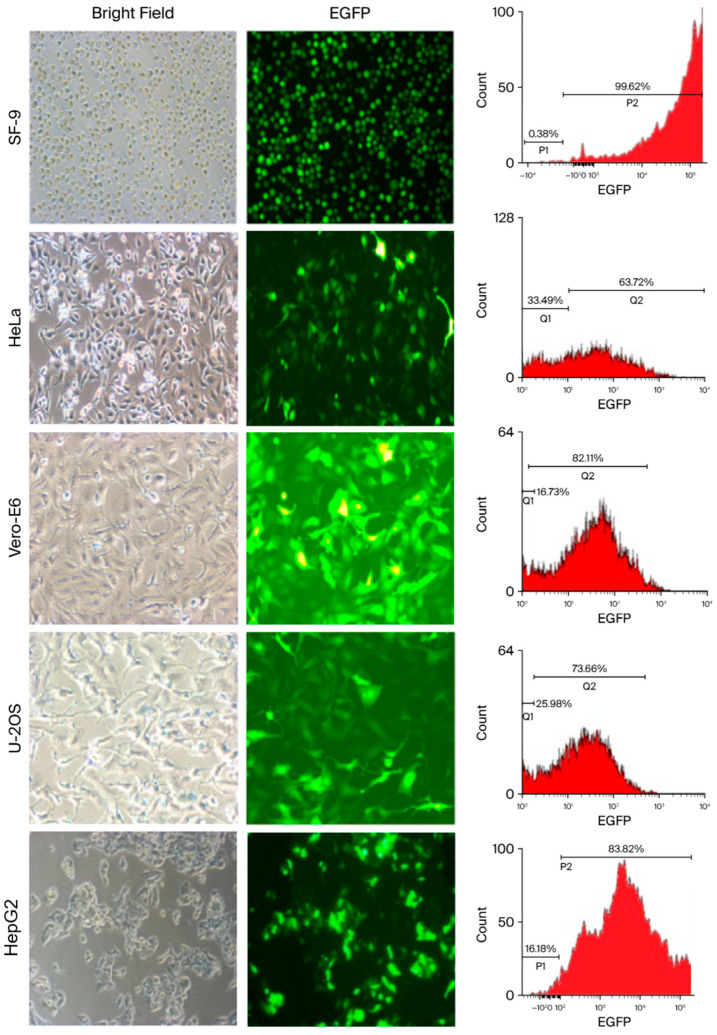 Figure 2
Figure 2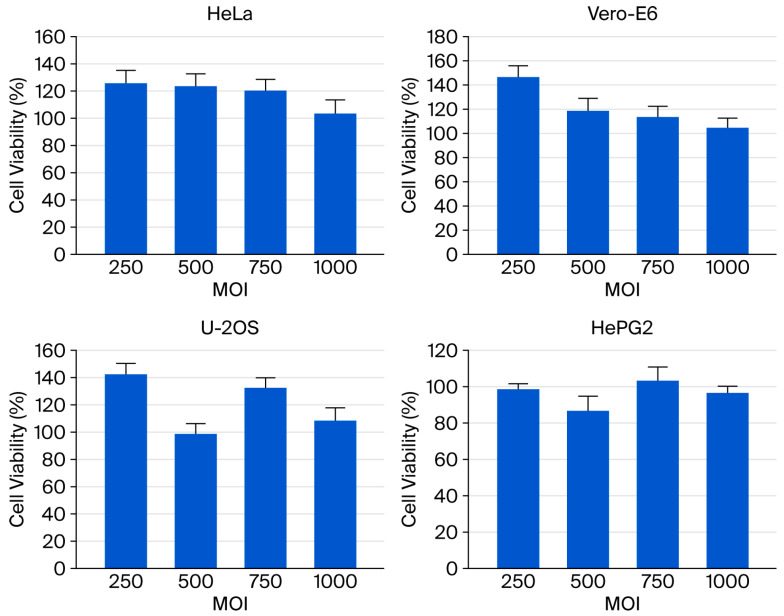 Figure 3
Figure 3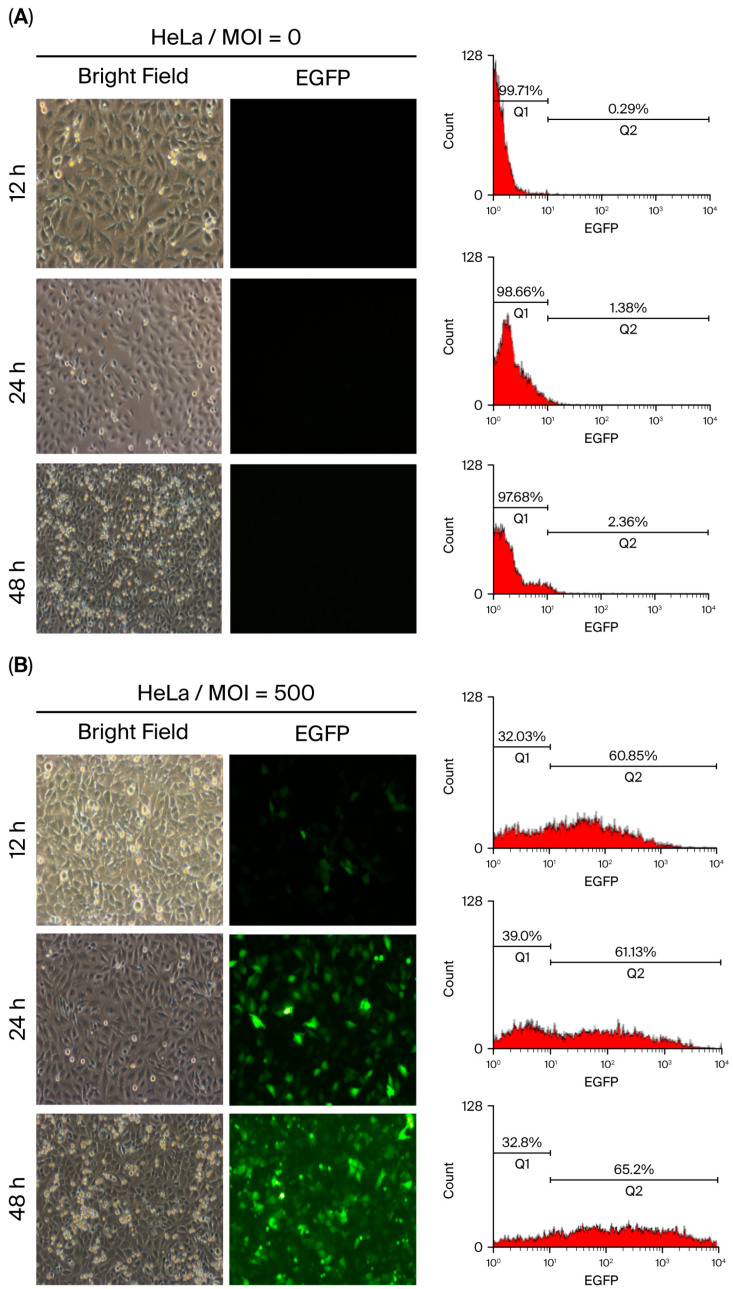 Figure 4
Figure 4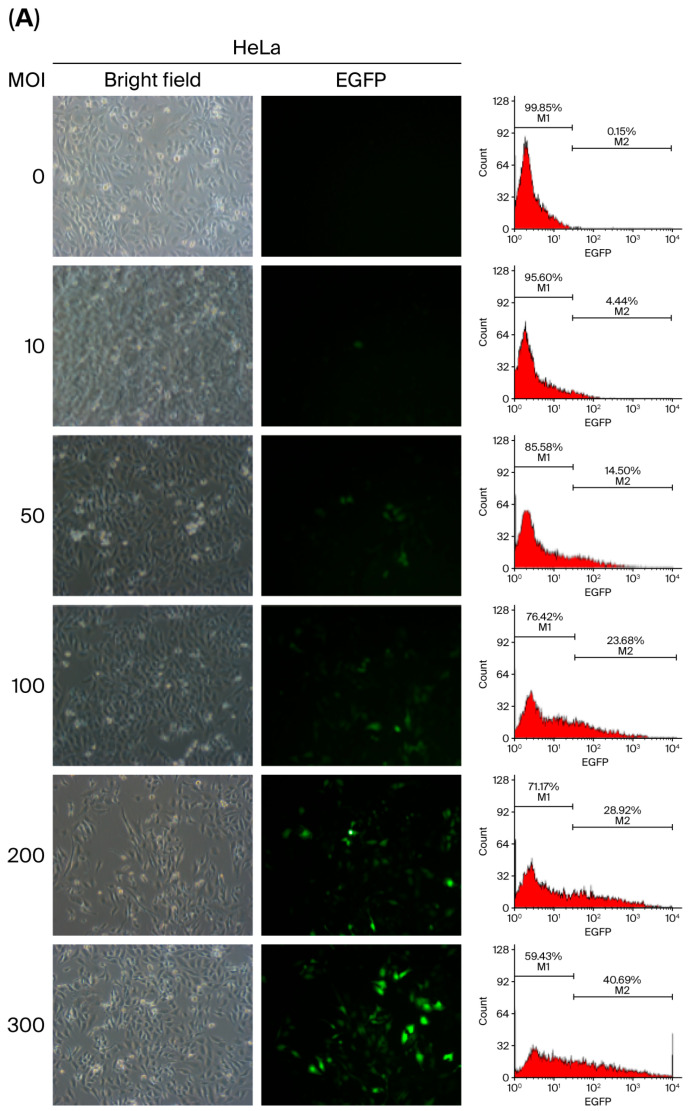 Figure 5
Figure 5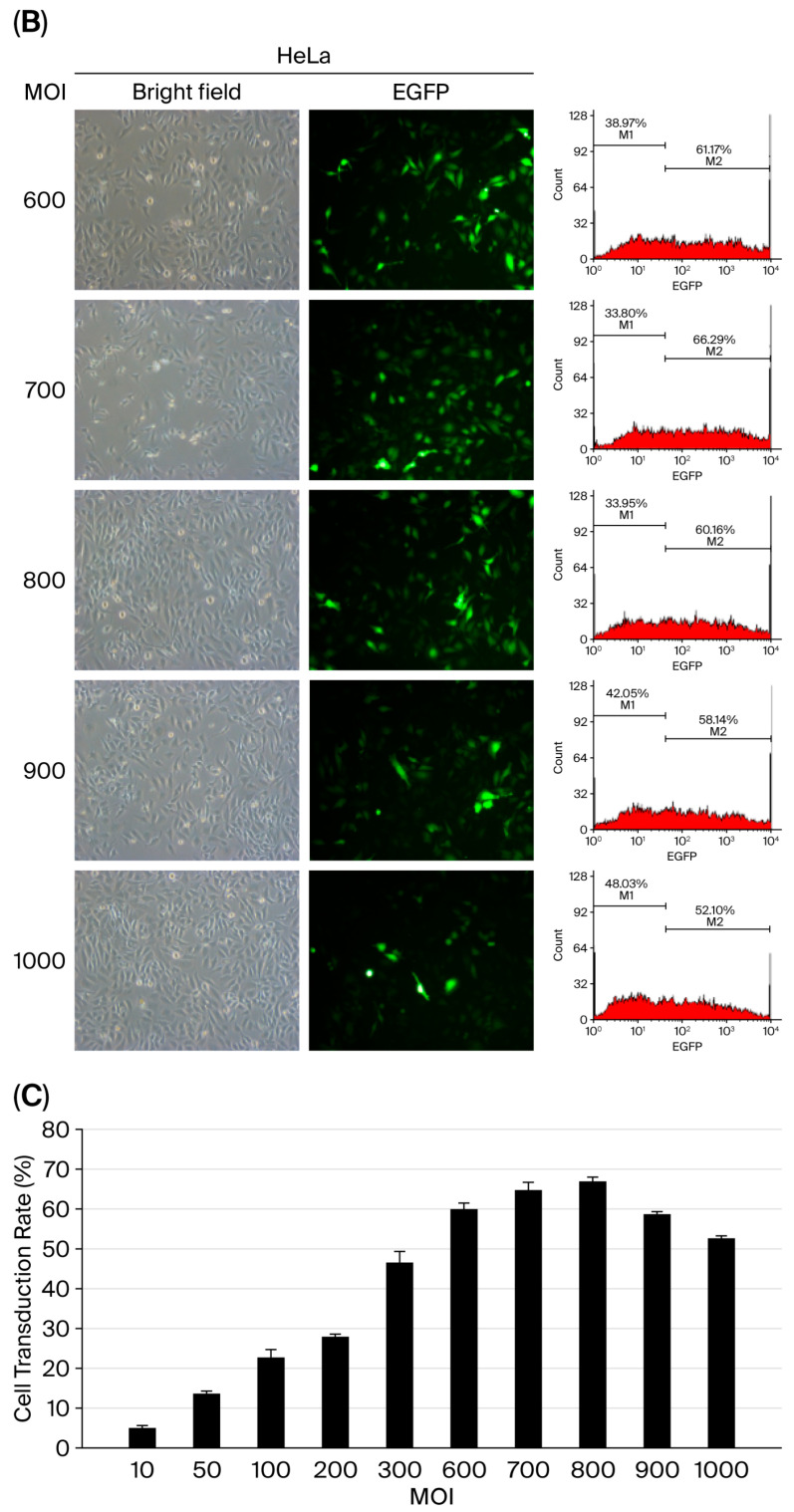 Figure 6
Figure 6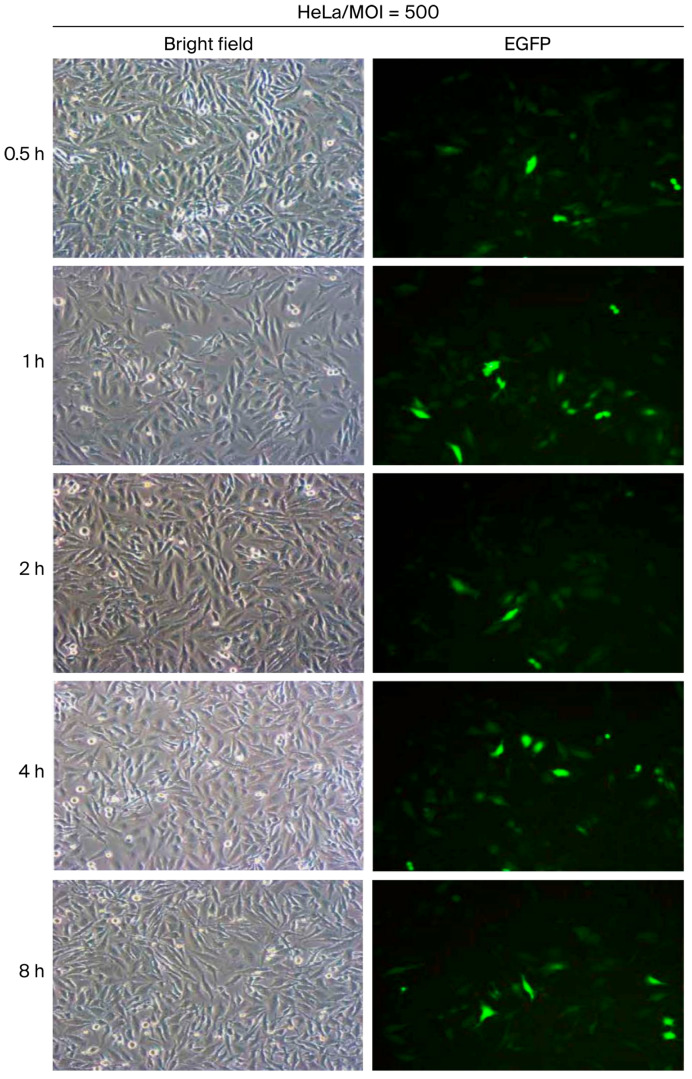 Figure 7
Figure 7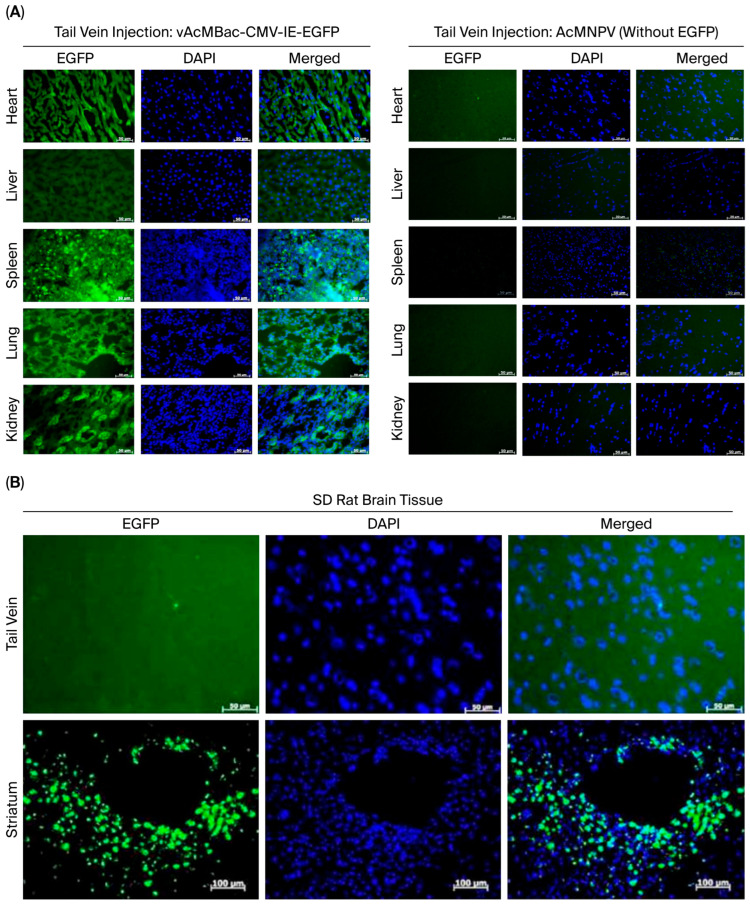 Figure 8
Figure 8- —Yuan’s General Hospital
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsViral Infectious Diseases and Gene Expression in Insects · Virus-based gene therapy research · Cancer Research and Treatments
1. Introduction
Baculoviruses, members of the Baculoviridae family, are best represented by Autographa californica multiple nuclear polyhedrosis virus (AcMNPV), an enveloped virus with a circular double-stranded DNA genome of 80–180 kb. The insect specificity of baculoviruses has led to their application as safe potential insecticides for use against forestry and agricultural pests [1]. This use has prompted an investigation into the possibility of baculovirus replication in vertebrate systems. Low levels of AcMNPV replication have been reported in Chinese hamster cell lines [2]. Although numerous studies have identified various mammalian cell types as being susceptible to baculovirus transduction, there is no evidence of viral replication, persistence, or cytopathic effects in these cells [3,4,5,6]. In 1995, Hofmann et al. created recombinant AcMNPV equipped with a cytomegalovirus (CMV) promoter to activate the truncated large T antigen of SV40 in human hepatocytes, thereby demonstrating baculovirus-mediated gene transfer into mammalian cells in vitro [7]. Since then, an increasing number of mammalian cell lines have been shown to support AcMNPV transduction [8,9,10,11,12]. These studies have encouraged the development of baculovirus as a potential gene delivery vector for both in vitro and in vivo gene therapy applications.
The restriction of in vivo delivery of genes by baculovirus vectors into mammalian cells via systemic administration has been attributed to two major factors: inactivation of the virus by the mammalian complement system [13,14,15,16,17] and reduced transgene expression efficiency due to induction of innate antiviral immune responses, such as interferons (IFNs) [18,19,20,21,22,23,24]. However, the use of baculovirus vectors in ex vivo applications and in immune-privileged sites—such as tissue-engineered cartilage and bone—has met with promising success [25].
The baculovirus-based gene expression method has been shown to ameliorate ischemia-reperfusion injury (IRI) in organs during transplantation procedures [26]. Moreover, recombinant baculoviruses have demonstrated the ability to penetrate mammalian kidneys and livers during cold preservation [27]. Baculoviruses’ potential as gene therapy vectors for brain cancer has also been evaluated [28]. In addition, the baculovirus expression vector has emerged as a promising platform for vaccine production, offering high safety, rapid scalability, and flexible product design [29]. Collectively, these studies highlight the broad applicability of baculovirus-based vectors.
Recently, several studies have shown that recombinant baculoviruses can mediate in vivo gene transfection. One such study demonstrated that the delivery of baculovirus to adventitial cells of rabbit carotid arteries via a collar-mediated approach was efficient, although transgene expression was transient. In this study, LacZ-baculoviruses were delivered using a silastic collar placed around the rabbit carotid arteries—a design that effectively avoids contact with blood components [30]. In subsequent studies, researchers used recombinant AcMNPV as a transfer vector for in vivo targeting of different organs, including the brain, eyes, skeletal muscles, cerebral cortex, and testes [31,32,33,34,35,36]. However, these studies relied on direct injection of recombinant baculoviruses into target organs to transfer genes. Whether baculovirus-based vectors can mediate gene expression in animal organs following intravenous administration remains unknown.
In this study, we first constructed a recombinant AcMNPV, designed as a dual-expression vector containing both the CMV immediate-early promoter (CMV-IE, Pcmv) and the polyhedron promoter (Pph). We then optimized key parameters to enhance baculovirus transduction efficiency across various mammalian cell types. Finally, we evaluated in vivo baculovirus-mediated gene delivery by administering the vector via tail vein injection into Sprague Dawley (SD) rats, assessing enhanced green fluorescent protein (EGFP) expression in the heart, liver, spleen, lungs, and kidneys. In parallel, we performed stereotaxic injections into the striata of SD rats to monitor EGFP expression in neural tissues.
2. Results
2.1. Construction of a Dual-Promoter Recombinant AcMNPV Expression Vector
A recombinant AcMNPV containing dual promoters was created and designated vAcMBac-CMV-IE-EGFP. This construct harbors an EGFP expression cassette whose activity is driven by both the CMV-IE and Pph promoters (Figure 1), as detailed in the Materials and Methods Section. The ability of vAcMBac-CMV-IE-EGFP to infect both insect cells and mammalian cells was evaluated by assessing EGFP expression using fluorescence microscopy and flow cytometry after 48 h of viral infection at a multiplicity of infection (MOI) of 500. The transduction efficiency in the insect cell line Sf-9 reached 95.52 ± 4.86%, while the efficiencies achieved in the mammalian cell lines HeLa, Vero E6, U-2 OS, and HepG2 were 60.87 ± 2.59%, 80.87 ± 2.50%, 70.14 ± 1.70%, and 80.53 ± 3.31%, respectively (Figure 2 and Table 1). The observation of over 50% transduction efficiency in Sf-9 cells and in most tested mammalian cell lines indicates that both the polyhedron (Pph) and CMV-IE promoters in the viral construct were functionally active.
2.2. High-Dose Baculovirus Transduction Exhibits Minimal Cytotoxicity in Mammalian Cells
Although baculovirus’s inherent inability to replicate and low cytotoxicity in mammalian cells have been previously reported, the effects of extremely high viral dosages on transduction have not been thoroughly examined. In this study, the cytotoxicity of the baculoviral vector was assessed in four mammalian cell lines at MOI values ranging from 250 to 1000 using the MTT assay. After 48 h of viral infection, no obvious cytopathic effects were observed in any of the tested cells, which maintained viability above 95% even at the highest MOI tested (Figure 3). These findings indicate that recombinant baculovirus infection in mammalian cells exhibits minimal cytotoxicity, supporting its potential as a safe gene delivery method.
2.3. Temporal Dynamics of Baculovirus-Mediated Transduction Efficiency
The transduction efficiency of baculovirus is typically assessed by quantifying EGFP-expressing cells using fluorescence microscopy at 48 h post-infection (p.i.), when infected cells generate sufficient fluorescence intensity for detection. However, vAcMBac-CMV-IE-EGFP is expected to be internalized by cells at earlier time points. To investigate this, HeLa cells were inoculated with or without vAcMBac-CMV-IE-EGFP (MOI = 500), and the EGFP-expressing cells were analyzed at 12, 24, and 48 h p.i. using both flow cytometry and fluorescence microscopy.
Uninfected HeLa cells served as negative controls and showed no EGFP signal (Figure 4A). Flow cytometry analysis revealed that approximately 58.87 ± 1.83% of inoculated cells expressed EGFP at 12 h p.i., with fluorescence intensities ranging from 10^1^ to 10^4^. However, only a few EGFP-expressing cells were visible via fluorescence microscopy at this time point. At 24 h p.i., 59.40 ± 1.79% of cells expressed EGFP, and at 48 h p.i., the proportion increased to 62.86 ± 1.43%, with both having similar fluorescence intensity ranges (Figure 4B and Table 2). These results indicate a progressive increase in EGFP-expressing cells over time. While fluorescence microscopy observation at 48 h p.i. is sufficient to evaluate baculovirus transduction efficiency, flow cytometry enables earlier detection at 12 h p.i. with greater sensitivity, suggesting that flow cytometry may be more effective than fluorescence microscopy for early assessment of transduction efficiency.
2.4. Optimal Parameter for Assessing the Transduction Efficiency of Baculovirus
The transduction efficiency of baculoviral vectors is influenced by inoculation parameters such as viral load and exposure duration. To determine the optimal conditions for baculovirus-mediated gene delivery in mammalian cells, we used HeLa cells due to their intermediate transduction efficiency (60.87 ± 2.59%) at an MOI of 500, as previously described in Figure 2. A range of viral doses (MOI 10 to 1000) was tested, and transduction efficiency was assessed by quantifying EGFP-positive cells using flow cytometry and fluorescence microscopy at 48 h p.i. The results showed a progressive increase in transduction efficiency with an increasing MOI (Figure 5A,B), reaching a peak of 64.41% at an MOI of 700. However, efficiency declined to 51.35% at MOI 1000 (Figure 5C), suggesting there is a threshold beyond which higher viral loads may reduce transduction performance.
Prolonging the virus incubation period is generally expected to enhance transduction efficiency. However, the optimal incubation duration at a given viral dosage remains to be defined. In this study, HeLa cells were inoculated with vAcMBac-CMV-IE-EGFP at a multiplicity of infection (MOI) of 500 and incubated with gentle rocking for 0.5, 1, 2, 4, or 8 h to determine the optimal exposure time. The transduction efficiency was 44.8% after 0.5 h of incubation and increased to 50.4%, 60.7%, and 60.3% following 1, 2, and 4 h of incubation, respectively. However, extending the incubation period to 8 h resulted in a slight decline in efficiency to 56.2% (Figure 6 and Table 3). According to these results, transduction efficiency in mammalian cells, as evaluated in HeLa cells, was optimal at an MOI of 600–800 with a 2–4 h incubation period in which gentle rocking was employed.
2.5. Recombinant AcMNPV as an In Vivo Gene Delivery Vector Targeting Mammalian Organs
We demonstrated that several mammalian cell lines inoculated with a high viral dose of up to MOI 1000 exhibited no evidence of cytopathic effects (Figure 3). To assess whether a high-MOI infection could mediate transgene expression in vivo, Sprague Dawley (SD) rats were intravenously injected via the tail vein with 3.5 mL of vAcMBac-CMV-IE-EGFP at a concentration of 5 × 10^7^ PFU/mL. At 72 h p.i., the animals were sacrificed, and EGFP expression was evaluated in various organs. The control Sprague Dawley (SD) rats underwent the same procedures as the experimental group but were inoculated with AcMNPV lacking the EGFP gene. EGFP expression was clearly detected in the heart, liver, spleen, lungs, and kidneys of SD rats inoculated with vAcMBac-CMV-IE-EGFP, whereas no EGFP signal was observed in rats that received AcMNPV without the EGFP gene (Figure 7A). We did not observe an EGFP signal in the brains of SD rats after tail vein inoculation of vAcMBac-CMV-IE-EGFP. However, direct injection of 20 μL of vAcMBac-CMV-IE-EGFP into the striatum (a part of the basal ganglia) successfully transduced neural cells in vivo (Figure 7B). These data suggest that recombinant baculovirus can be used for transgene delivery in vivo.
3. Discussion
Gene therapy has advanced to the point where it can be used to treat a variety of illnesses, including genetic deficiencies, cancer, and rare diseases, and it can even be used in wound repair. Its success largely depends on the development of suitable delivery vehicles that can efficiently and safely introduce foreign genes into mammalian cells. Both viral and nonviral vectors have been employed in this field [37], with viral vectors generally considered the more effective platforms for gene delivery [38,39]. Several viral vectors, including adeno-associated vectors (AAVs), lentivirus, and retrovirus, have been approved by the US Food and Drug Administration (FDA) for gene therapy implementation [40]. However, AAV vectors face challenges in large-scale production and carry risks of toxicity and inflammation. Lentiviral and retroviral vectors also suffer from limited cloning capacity, and their genomes may integrate into host DNA, raising the possibility of insertional mutagenesis. In addition, the replication properties of retroviruses have raised safety concerns regarding oncogenesis, including virus-induced T-cell lymphomas. These disadvantages restrict their broader application [41].
Engineered recombinant baculoviruses equipped with mammalian cell-active promoters can enter a broad range of mammalian cells and transiently express foreign genes without undergoing viral replication. Their inability to replicate in mammalian cells, combined with their low cytotoxicity, makes them relatively safe, scalable, and cost-effective, thereby establishing baculoviruses as valuable vectors for gene therapy [7,8,10,11].
In this study, we generated a recombinant AcMNPV carrying the shuttle promoters CMV-IE (Pcmv) and polyhedron (Pph) to potentiate EGFP expression. We hypothesized that vAcMBac-CMV-IE-EGFP expressed EGFP in mammalian cells under the control of the CMV-IE promoter/enhancer, while expression in insect cells was driven by the polyhedron promoter (Pph). We demonstrated that this recombinant baculovirus, vAcMBac-CMV-IE-EGFP, mediated EGFP expression in both insect and mammalian cells. In the permissive insect cell line Sf-9, transduction efficiency approached 96%, whereas in four non-permissive mammalian cell lines (Vero E6, HeLa, U-2 OS, and HepG2), effective transduction rates ranged from 61% to 81%. These results confirmed robust EGFP expression in both permissive and non-permissive mammalian cells, highlighting the therapeutic potential of baculovirus, although efficiency varied by cell type (Figure 2).
Regarding the therapeutic potential of baculoviral vectors, safety is an important concern. Many researchers have reported that baculoviruses replicate only in insects and not in mammalian cells [42,43]. In this study, all tested mammalian cells infected with an extremely high dosage of vAcMBac-CMV-IE-EGFP at an MOI of up to 1000 showed no evidence of cytopathic effects (Figure 3). Our findings indicate that vAcMBac-CMV-IE-EGFP transduction in mammalian cells is safe even at extremely high concentrations, a finding consistent with previous reports [44].
The mechanisms underlying baculoviruses’ binding with and entry into mammalian cells remain incompletely understood. Several studies have investigated potential receptors on mammalian cells and viral molecules that facilitate baculoviruses’ attachment to the cell membrane. It has been reported that baculovirus gp64 and cell-surface heparan sulfate are required for viral entry [45,46,47], with GP64 binding to heparan sulfate proteoglycans (HSPGs) [48]. These findings have prompted efforts to modify baculoviral surface molecules, such as incorporating vesicular stomatitis virus envelope G protein (VSVG) or increasing gp64 expression, to enhance gene transfer efficiency [47,49]. Following attachment to cell-surface molecules, baculoviruses are internalized via lipid rafts [45]. Infection of insect cells and transduction of mammalian cells by baculoviruses have been extensively shown to occur primarily through endocytosis [49,50,51]. However, Dong et al. demonstrated that AcMNPV enters Sf9 cells through direct fusion at low pH, and that acidic conditions can significantly enhance baculovirus entry into mammalian cells, thereby increasing transduction efficiency [52,53,54].
The transduction efficiency of baculoviruses can be improved by optimizing transduction conditions. A previous study reported that when HeLa cells were transduced with baculovirus, incubation for 4 h at 25 °C in Dulbecco’s phosphate-buffered saline (D-PBS) resulted in higher expression of the reporter protein GFP [53].
Although baculovirus infection has been shown to induce antiviral responses in mammalian cells, we hypothesized that administering a high dosage of recombinant AcMNPV could overcome inactivation mediated by innate immune mechanisms, such as the complement system and interferons (IFNs). Our in vivo data demonstrated that Sprague Dawley (SD) rats inoculated via the tail vein with 3.5 mL of vAcMBac-CMV-IE-EGFP at a concentration of 5 × 10^7^ PFU/mL exhibited clear EGFP expression in the heart, liver, spleen, lungs, and kidneys but not in the brain. These findings suggest that systemic delivery of recombinant baculovirus vectors at high, non-lethal doses can allow effective transgene expression in vivo, providing evidence of their potential utility in gene therapy applications.
There are two limitations that should be mentioned. Firstly, although this study demonstrated that EGFP protein expression was detected in the heart, liver, spleen, lungs, and kidneys tissues from SD rats inoculated with vAcMBac-CMV-IE-EGFP (Figure 7A, left panels), it did not demonstrate the EGFP protein expression in those without harboring the EGFP gene (Figure 7A, right panels). However, we have not assessed the RT-PCR, Western blot, ELISA, or fluorescence assay in a plate reader of the rat tissues. This study has not provided more supportive data on EGFP gene expression analysis, which is a limitation of this study. Secondly, we carried out a comparison of EGFP protein expression in the striatum by direct injection versus tail vein injection of vAcMBac-CMV-IE-EGFP. This study only performed a qualitative comparison of the fluorescence imaging without any quantitative comparison between these two experiments. This was another limitation.
4. Materials and Methods
4.1. Cells
The insect cell line Sf-9, isolated from ovarian tissue of Spodoptera frugiperda, was purchased from Bioresource Collection and Research Center, Hsinchu, Taiwan (catalog number CCRC60011), and cultured in TC-100 insect cell culture medium at 26 °C with 10% heat-inactivated fetal bovine serum (FBS) (Gibco). The mammalian cell lines Vero-E6 and U-2 OS were provided by Professor Yu-Chan Chao from the Institute of Molecular Biology, Academia Sinica, Nankang, Taipei, Taiwan. The HeLa and HepG2 cell lines were provided by Chi-Mei Medical Center, Tainan, Taiwan. Vero-E6 cells isolated from African green monkey kidneys were incubated in Minimum Essential Medium (MEM) with 2 mM L-glutamine, 1.5 g/L sodium bicarbonate, 1% sodium bicarbonate, and 0.1 mM nonessential amino acids, supplemented with 10% (v/v) FBS. U-2 OS, a human osteogenic sarcoma cell line, was incubated in McCoy 5A medium with 1.5 mM L-glutamine and 10% (v/v) FBS. HeLa cells from a human cervical epithelioid carcinoma were maintained at 37 °C in MEM with 2 mM L-glutamine, 1.5 g/L of sodium bicarbonate, 0.1 mM non-essential amino acids, and 1.0 mM sodium pyruvate, supplemented with 10% (v/v) FBS. HepG2, a human hepatocellular carcinoma cell line, was maintained in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 2 mM L-glutamine, 10 mM HEPES, and 10% (v/v) FBS. All mammalian cell lines were maintained at 37 °C in a 5% C02-95% air environment.
4.2. Plasmids
pAtEG containing an insert of the EGFP gene was kindly provided by Professor Yu-Chan Chao from the Institute of Molecular Biology, Academia Sinica, Nankang, Taipei, Taiwan. pFastBac was purchased from Life Technologies, Grand Island, NY, USA.
4.3. Construction of Recombinant AcMNPV with Dual Promoters Harboring EGFP Expression Cassette
We modified the transvector pFastBac1, one component of Bac-to-Bac^®^ Baculovirus Expression System (Life Technologies Corporation, Grand Island, NY, USA), for the construction of recombinant AcMNPV via insertion of the CMV-IE (Pcmv) promoter in the anterior part of the multiple cloning sites through BamHI and EcoRI restriction endonuclease enzyme sites. The CMV-IE fragment was amplified by PCR from plasmid pAtEG. The primer sequences subjected to amplification of CMV-IE fragments were as follows: forward primer, 5′CGAGGATCCACAAACTGGAAATGTCTATC3′, and reverse primer, 5′GCTGAATTCGGTATATCTCCTTTGATTGT3′. The generated transvector pFastBac1-CMV-IE was equipped with dual promoters, Pph and Pcmv, to express foreign genes in insect Sf-9 cells and mammalian cells, respectively. The posterior parts of the multiple cloning sites of pFastBac1-CMV-IE were kept for cloning foreign genes. For the construction of recombinant AcMNPVs harboring the enhanced green fluorescence protein (EGFP) expression cassette, the EGFP coding region was obtained via PCR from pAtEG and then cloned into the pFastBac1-CMV-IE between Spe I and Xho I restriction endonuclease enzyme sites to generate pFastBac1-CMV-IE-EGFP. The primer sequences for amplification of the EGFP gene were as follows: forward primer, 5′TATACTAGTCATGGTGAGCAAGGGCGAGG3′, and reverse primer, 5′CGTCTCGAGTTACTTGTACAGCTCGTCCA’.
Then, pFastBac1-CMV-IE-EGFP was transformed into E. coli component, the MAX Efficiency^®^ DH10Bac™, of Bac-to-Bac^®^ Baculovirus Expression System. DH10Bac™ was internalized as a modified AcMNPV (bacmid), a large extra genome of E. coli cells, and a helper plasmid that expresses transposase. The sequences of Tn7 L and Tn7 R transposition sites were cloned into both pFastBac1 and bacmid. Furthermore, a lacZ (b-galactosidase gene) was designed to be located between Tn7 L and Tn7 R in the bacmid. When the pFastBac1-CMV-IE-EGFP was transformed into the DH10Bac, the transposase served to replace the EGFP gene expression cassette from pFastBac1-CMV-IE-EGFP with the lacZ gene located in the bacmid. Ideally, the successful recombinant bacmid should lose lacZ gene function and be identified by blue and white colony screening. The white colony, which harbors the recombinant bacmid, was selected and cultured for further purification. The purified bacmid DNA, i.e., bFastBac-CMV-IE-EGFP harboring the EGFP gene expression cassette, was further transfected into insect cells, Sf-9, to replicate recombinant AcMNPV, designated as vAcMBac-CMV-IE-EGFP.
4.4. Baculovirus Production in Insect Cells and Titration
Wild-type AcMNPV and its derived recombinant viruses were generated and propagated in Sf-9 cells according to the standard protocols described by O’Reilly et al. [55]. The titers of the virus clones were determined via both quantitative PCR (qPCR) and the 50% tissue culture infective dose (TCID50) [56].
4.5. In Vitro Transduction of Insect and Mammalian Cells with Recombinant AcMNPV
A monolayer of cells was cultured in a 6-well plate in a matched medium of each cell line for 12 h before viral inoculation. Then, the culture medium was removed and replaced with 1000 μL of fresh FBS-free medium containing recombinant AcMNPV, vAcMBac-CMV-IE-EGFP, at a multiplicity of infection (MOI) ranging from 250 to 1000. After an adsorption period with gentle rocking, ranging from 0.5 h to 8 h, the inoculum was removed, and cells were washed with phosphate-buffered saline (PBS) with a pH of 7.4. Then, a fresh medium with FBS was added. The insect cells were incubated at 26 °C, and mammalian cells were cultured at 37 °C, with 5% CO_2_.
4.6. Flow Cytometry
The cells were examined for enhanced green fluorescent protein (EGFP) expression at 48 h post-infection (p.i.) via fluorescence microscopy and flow cytometry. The cells were then harvested with trypsin, washed with PBS twice, and resuspended in 1 mL of DPBS. The cells were collected and analyzed using an LSR II flow cytometer (Becton-Dickinson, San Jose, CA, USA). The EGFP proteins within the cells were excited at 488 nm and detected at 525 nm. About 10^4^ cells with three repeats were collected per specimen. Cells without virus transduction were used as the negative control. Transduction efficiency was obtained as a percentage: the number of EGFP-positive cells over the number of total cells collected. The level of EGFP expression was measured as the mean fluorescence intensity multiplied by the total cell number for each sample. Data are expressed as means ± standard deviation (SD).
4.7. 3-(4,5-Dimethylthiazol-2-Yl)-2,5-Diphenyltetrazolium Bromide (MTT) Assay
Cell viability subject to AcMNPV transduction was examined via MTT assay. An appropriate number of cells at about 50% confluence were cultured in a 96-well plate in a suitable medium for 12 h. Then, viral transduction, as described above, was performed. At 72 h after inoculation, 20 μL of MTT solution (Merck, Darmstadt, Germany) (5 mg/mL PBS) was added to each well, and the plate was incubated at 37 °C for 4 h. After medium removal, 200 μL of DMSO was added to each well, and the plate was gently shaken for 5 min. Absorbance was determined at 540 nm. Quadruplicate wells were applied to each MOI for a specific transduction. The 0.002% (v/v) DMSO (vehicle)-treated non-transfected cells were employed as the control.
4.8. In Vivo Transduction of SD Rats with Recombinant AcMNPV
The in vivo transduction of vAcMBac-CMV-IE-EGFP was conducted with male Sprague Dawley (SD) rats weighing 350–400 g via either tail vein or striatum injection of the virus with an infectious titer of 5 × 10^7^ PFU/mL. For tail vein injection, SD rats in the study group (n = 6) were injected with vAcMBac-CMV-IE-EGFP, which harbors an EGFP expression cassette. The rats in the control group (n = 6) were injected with AcMNPV, which does not harbor the EGFP gene. The SD rats without anesthesia were injected with 3.5 mL of AcMNPV via tail vein inoculation. For striatum infection (n = 6), 20 μL vAcMBac-CMV-IE-EGFP was injected into the striatum of SD rats, which were anesthetized with Urethane (100 mg/kg). After 72 h p.i., the infected rats were anesthetized with urethane (100 mg/kg) and sacrificed. The rats were immediately transferred to a sterile bench for cardiac perfusion with physiological saline to remove blood, followed by fixation with 4% paraformaldehyde. Brain, heart, lung, liver, spleen, and kidney tissues were carefully dissected under sterile conditions and placed in pre-cooled PBS. Fragile organs such as the brain and lungs were handled gently to avoid structural damage, while the liver and kidney samples were trimmed into standardized blocks (approximately 1 cm × 1 cm × 0.3 cm). All tissues were rinsed in PBS for 5 min to remove residual blood and fluids and then blotted dry with sterile filter paper. Processed samples were embedded in OCT compound and frozen for 15–20 min until solidified, sectioned at 5–10 µm thickness (5 µm for brain and 8 µm for liver and kidney), and subjected to EGFP immunofluorescence and nuclear DAPI staining. Fluorescence signals were observed directly under a fluorescence microscope using excitation at ~488 nm to detect EGFP expression. All animal experiments were approved by the Institutional Animal Care and Use Committee (IACUC) of Southern Taiwan University.
4.9. Statistical Analysis
Statistical analysis of flow cytometry data between the control group and the vAc-MBac-CMV-IE-EGFP transduction group was performed using Student’s t-test, with differences considered significant at p < 0.05.
5. Conclusions
We developed a high-titer, dual-promoter recombinant baculovirus (vAcMBac-CMV-IE-EGFP) capable of efficient and safe gene delivery. The use of high MOI baculovirus resulted in high transduction efficiencies in vitro, with over 80% in HepG2 and Vero E6 cells, and low cytotoxicity even at an MOI of 1000. In vivo, tail vein injection in rats resulted in robust EGFP expression across multiple organs, demonstrating that systemic delivery at high, non-lethal doses can overcome immune inactivation. Moreover, successful localized delivery to the rat striatum highlights this method’s potential for targeted applications. These findings establish this high MOI baculovirus infection as a versatile and promising platform for both systemic and tissue-specific gene therapy.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Moscardi F. Assessment of the application of baculoviruses for control of Lepidoptera Annu. Rev. Entomol.19994425728910.1146/annurev.ento.44.1.25715012374 · doi ↗ · pubmed ↗
- 2Mc Intosh A.H. Shamy R. Biological studies of a baculovirus in a mammalian cell line Intervirology 19801333134110.1159/0001491436998919 · doi ↗ · pubmed ↗
- 3Groner A. Granados R.R. Burand J.P. Interaction of Autographa californica nuclear polyhedrosis virus with two nonpermissive cell lines Intervirology 19842120320910.1159/0001495226376420 · doi ↗ · pubmed ↗
- 4Ignoffo C.M. Rafajko R.R. In vitro attempts to infect primate cells with the nucleopolyhedrosis virus of Heliothis J. Invertebr. Pathol.19722032132510.1016/0022-2011(72)90163-24632202 · doi ↗ · pubmed ↗
- 5Tjia S.T. zu Altenschildesche G.M. Doerfler W. Autographa californica nuclear polyhedrosis virus (Ac NPV) DNA does not persist in mass cultures of mammalian cells Virology 198312510711710.1016/0042-6822(83)90067-36402854 · doi ↗ · pubmed ↗
- 6Volkman L.E. Goldsmith P.A. In Vitro Survey of Autographa californica Nuclear Polyhedrosis Virus Interaction with Nontarget Vertebrate Host Cells Appl. Environ. Microbiol.1983451085109310.1128/aem.45.3.1085-1093.198316346229 PMC 242411 · doi ↗ · pubmed ↗
- 7Hofmann C. Sandig V. Jennings G. Rudolph M. Schlag P. Strauss M. Efficient gene transfer into human hepatocytes by baculovirus vectors Proc. Natl. Acad. Sci. USA 199592100991010310.1073/pnas.92.22.100997479733 PMC 40743 · doi ↗ · pubmed ↗
- 8Boyce F.M. Bucher N.L. Baculovirus-mediated gene transfer into mammalian cells Proc. Natl. Acad. Sci. USA 1996932348235210.1073/pnas.93.6.23488637876 PMC 39799 · doi ↗ · pubmed ↗