Curcuminoids Activate TET Enzymes and Increase DNA Hydroxymethylation and Active Demethylation in Leukemia Cells
Sridhar A. Malkaram, Suhila Sawesi, Botao Peng, Badreldeen Rashrash, Hailey Cox, Tamer E. Fandy

TL;DR
Curcuminoids like curcumin and DMC boost TET enzyme activity, leading to increased DNA demethylation in leukemia cells.
Contribution
This study is the first to show that curcuminoids activate TET enzymes and promote active DNA demethylation in leukemia cells.
Findings
Both curcumin and DMC increased TET enzyme activity and mRNA expression in leukemia cells.
Curcumin and DMC elevated global 5hmC levels and induced active demethylation at gene promoters and other genomic regions.
Curcumin and DMC each caused promoter 5hmC increases and active demethylation in hundreds of genes.
Abstract
Curcuminoids demonstrate diverse pharmacological activity as antioxidant, neuroprotective, antitumor, and anti-inflammatory drugs. Dimethoxycurcumin (DMC) is a metabolically stable analog of curcumin, and both drugs modify the activity of several epigenetic enzymes that affect DNA methylation and histone modifications. 5-hydroxymethylcytosine (5hmC) is an epigenetic mark involved in active demethylation and in gene expression regulation. The effect of curcuminoids on the activity and expression of TET enzymes involved in 5hmC oxidation and active demethylation in leukemia cells is unclear. In this study, we investigated the impact of curcumin and DMC on the activity and expression of the three isoforms of TET enzymes. We also studied their effect on global 5hmC and performed a genome-wide analysis of 5hmC distribution at the single CpG level using oxidative bisulfite sequencing, which…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
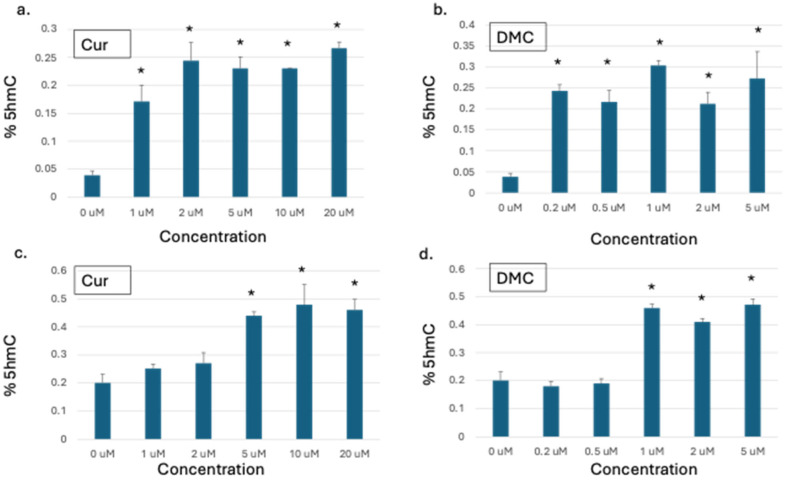 Figure 1
Figure 1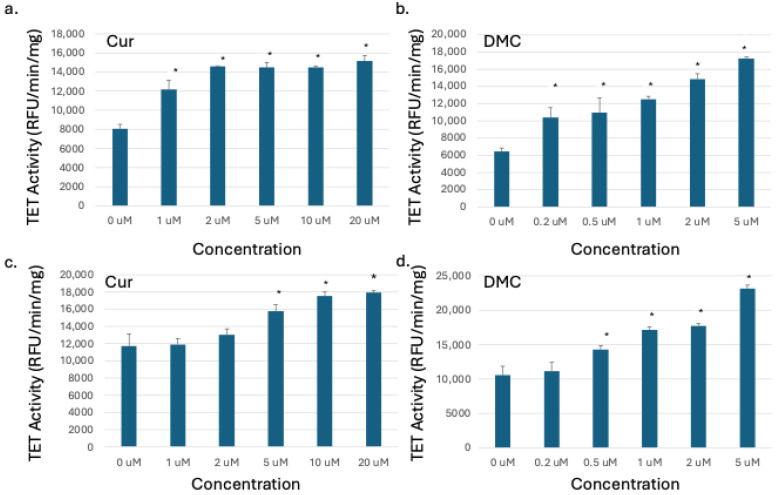 Figure 2
Figure 2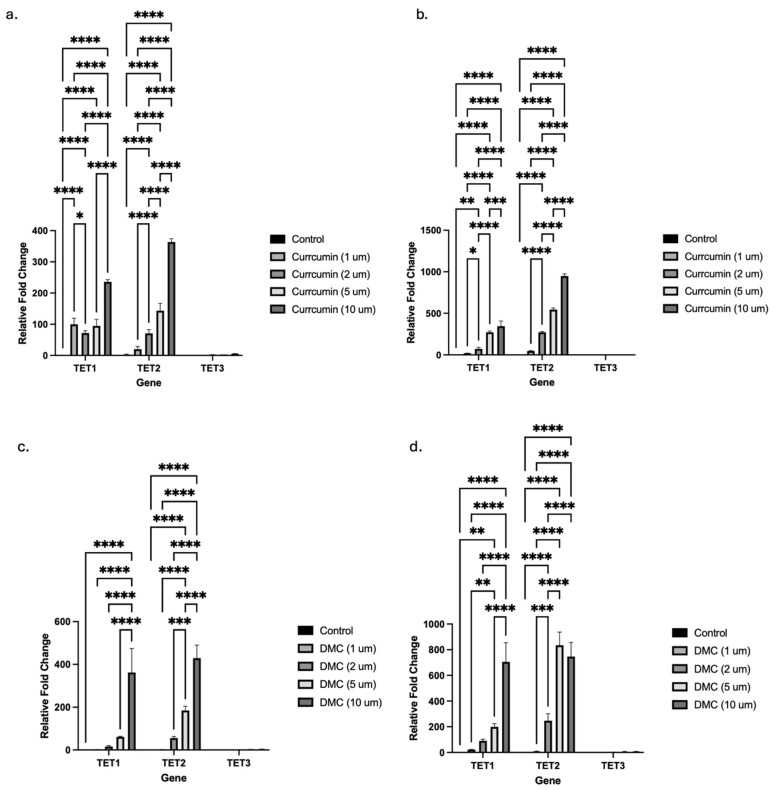 Figure 3
Figure 3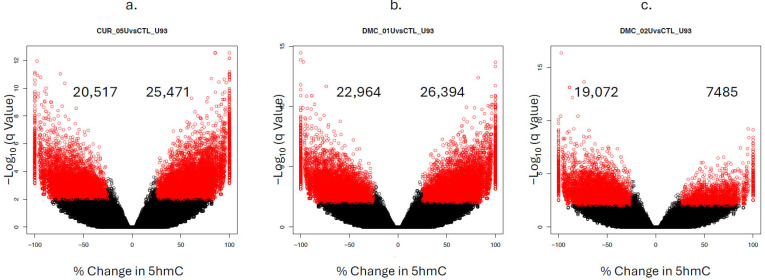 Figure 4
Figure 4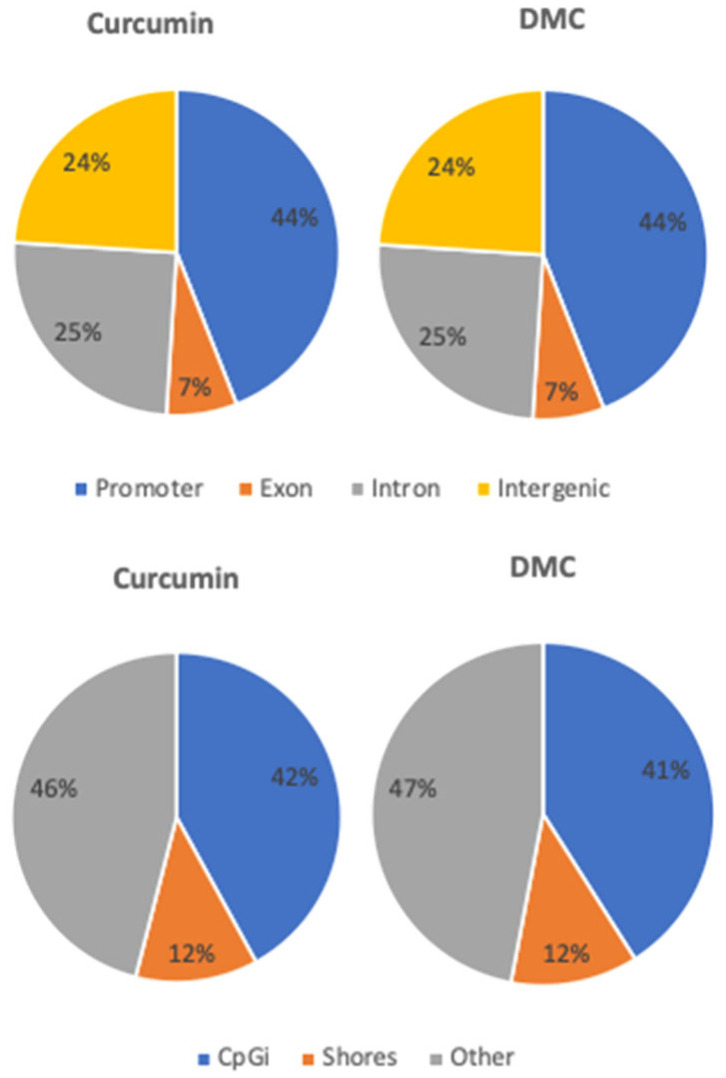 Figure 5
Figure 5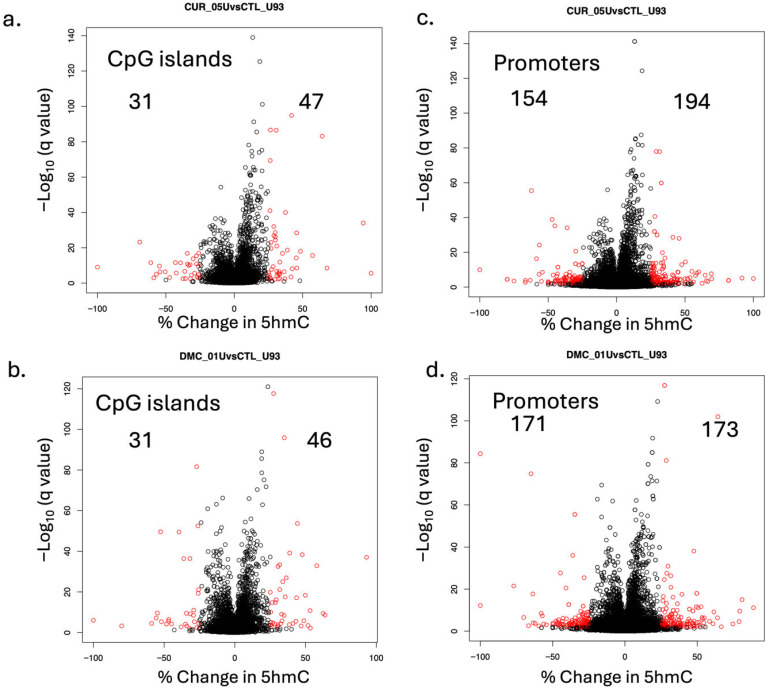 Figure 6
Figure 6- —National Institute of General Medical Sciences (NIGMS) of the National Institutes of Health (NIH)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCurcumin's Biomedical Applications · Epigenetics and DNA Methylation · Retinoids in leukemia and cellular processes
1. Introduction
Curcuminoids represent a small class of plant secondary metabolites that includes curcumin, demethoxycurcumin, and bisdemethoxycurcumin, all isolated from the turmeric rhizome [1,2]. A growing body of evidence demonstrates that curcuminoids induce a variety of epigenetic modifications [3,4,5]. Curcuminoids demonstrate extensive biological activity as antioxidant, neuroprotective, antitumor, and anti-inflammatory agents [6,7,8,9]. Curcumin is the most widely used curcuminoid, and despite its proven efficacy and safety against numerous diseases, its poor bioavailability [10], rapid metabolism [11], and rapid systemic elimination limit its therapeutic efficacy. Therefore, several curcumin analogs were synthesized to improve its bioavailability and stability.
Dimethoxycurcumin (DMC) is a more metabolically stable and potent synthetic analog of curcumin [5,12,13]. DMC showed antitumor effects and induced epigenetic alterations in tumor cells [3,5,14]. The antitumor activity of both curcumin and DMC could be attributed to their abilities to reverse the epigenetic alterations associated with tumor pathogenesis. Several studies documented the epigenetic alterations induced by these compounds in leukemia cells, such as their impact on DNA methylation [5,14,15,16], histone acetylation [17,18], and miRNA modulation [19,20]. However, their effect on DNA hydroxymethylation is largely unknown.
DNA methylation was believed to be an irreversible epigenetic modification until the discovery of the ten-eleven translocation protein 1 (TET1) that catalyzes the successive oxidation of 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC), and 5-carboxylcytosine (5caC) [21]. The TET protein family has three isoforms, namely, TET1, TET2, and TET3. These 5mC oxidation products serve as an intermediate in the conversion of 5mC to unmodified cytosines, a process known as active demethylation [22,23]. TET enzyme activity has emerged as an important tumor suppressor mechanism in cancer. TET2 is a frequently mutated gene in acute myeloid leukemia (AML) [24,25,26] and glioblastoma [27] and contributes to their pathogenesis. It is now recognized that all three TET isoforms are involved in a wide range of different cancer types. Accordingly, TET enzymes are rational targets for cancer therapy.
In this manuscript, we identified curcumin and DMC as active demethylators through the activation of TET enzymes. Both drugs increased the activity and induced the expression of the three TET isoforms in leukemia cells. Additionally, we detected a global increase in the 5hmC mark by both drugs. Genome-wide analysis of 5hmC at the single CpG level in leukemia cells revealed increases in 5hmC in CpG islands and promoters of leukemia-related genes. Moreover, decreases in 5hmC, indicating active demethylation, were similarly detected in promoters and CpG islands of leukemia-related genes. In conclusion, curcumin and DMC are capable of increasing DNA hydroxymethylation with consequent active demethylation of epigenetically silenced genes, which could contribute to their antitumor effect.
2. Results
2.1. Curcumin and DMC Increased Global DNA Hydroxymethylation in Leukemia Cells
The effect of curcumin and its analogs on DNA methylation reversal is controversial. Reversal of epigenetically silenced methylated genes was reported [28,29] but was not supported by later studies [5,15]. On the other hand, the effect of curcumin on DNA 5-hydroxymethylation (5hmC), an intermediate epigenetic mark that could also influence gene expression, is unclear [30]. Accordingly, we studied the effect of curcumin and its analog DMC on global DNA 5hmC in two leukemia cell lines to monitor any changes in this epigenetic mark.
U937 and HL60 human leukemia cells were treated with graded concentrations of either curcumin or DMC for 48 h, followed by DNA extraction and quantitation of global DNA 5hmC, as described in the methods. The concentrations used for both drugs induced minimal apoptosis even at the highest concentration, as described previously [3]. U937 cells showed a significant increase in global DNA 5hmC at all the concentrations used for both curcumin and DMC (Figure 1a,b). However, the 5hmC increase was comparable at the different drug concentrations used and did not show a linear dose–response relationship. On the other hand, HL60 cells showed a similar increase in global DNA 5hmC but at higher concentrations compared to U937 (Figure 1c,d). Taken together, curcumin and DMC increased global DNA 5hmC in leukemia cells.
2.2. Curcumin and DMC Increased the Activity of TET Enzymes in Leukemia Cells
The ten-eleven translocation (TET) enzyme family has three known isoforms (TET1, TET2, and TET3) involved in active DNA demethylation [31]. We detected a global 5hmC increase after curcumin or DMC treatment of leukemia cells, which is indirect evidence of TET enzyme activation. To confirm that, we measured the TET enzyme catalytic activity in the nuclear extracts of HL60 and U937 cells after 48 h treatment with graded concentrations of curcumin and DMC, as described in the methods. A significant increase in TET activity was observed by both drugs in U937 cells (Figure 2a,b). Even the lowest concentration of both drugs induced an increase in TET activity in U937. Similar to the global 5hmC results, HL60 cells were more resistant, and lower concentrations of curcumin or DMC did not induce TET activity (Figure 2c,d). Higher concentrations of both drugs induced an increase in TET activity, and the increase was concentration-dependent. In summary, an increase in TET activity was observed in the nuclear extracts of both cell lines after treatment with curcumin or DMC, which explains the previously observed increase in global DNA 5hmC.
2.3. Curcumin and DMC Induce TET Isoform Transcription in Leukemia Cells
Epigenetic modifiers, like 5-azacytidine and 5-aza-2′-deoxycytidine, induced the transcription of both the TET and DNMT enzyme isoforms in leukemia cells [5]. The observed increase in activity of TET enzymes by curcumin and DMC could be mediated through different mechanisms, like increasing the enzyme transcription, stabilization of mRNA of the enzyme, or direct binding of the drug to the enzyme. We hypothesized that curcumin and DMC induced the transcription of TET enzymes with a consequent increase in TET enzyme activity.
To test our hypothesis, we performed qRT-PCR to quantify the effect of both drugs on the transcription of the three isoforms, TET1, TET2, and TET3, in U937 leukemia cells. Curcumin treatment for 24 or 48 h significantly induced mRNA expression of both the TET1 and TET2 isoforms at all the concentrations tested (Figure 3a and Figure 3b, respectively). The highest fold increase in mRNA expression was observed at the highest curcumin concentrations (5 and 10 µM), and the 48 h treatment induced higher fold induction compared to the 24 h treatment. TET3 isoform transcription was also induced, but with much lower fold induction compared to the other isoforms (the increase is not visible on the graphs because of the scale). Similar to the other TET isoforms, TET3 mRNA induction was also concentration-dependent, with the highest fold induction (6-fold) observed at the 10 µM concentration after 48 h of treatment (Supplementary Figure S1a).
Similarly, DMC induced the transcription of both the TET1 and TET2 isoforms to a greater extent compared to TET3 after 24 and 48 h of treatment (Figure 3c and Figure 3d, respectively). TET3 mRNA was increased by different DMC concentrations up to 3-fold after 24 h and 8-fold after 48 h (the increase is not visible on the graphs in Figure 3 because of the scale, and Supplementary Figure S1b shows the effect of DMC on TET3 mRNA induction). Collectively, both drugs significantly induced the transcription of the three TET isoforms; however, TET3 isoform mRNA induction was relatively much less than the other isoforms.
2.4. Genome-Wide Mapping of 5hmC at the Single CpG Level in Leukemia Cells Treated with Curcuminoids
The above data demonstrated an increase in global 5hmC in leukemia cells after treatment with curcumin or DMC. Unfortunately, the data measured only the overall changes in 5hmC without determining the genomic locations of the changes in 5hmC. Additionally, the method of analysis was not capable of detecting active demethylation. To overcome these drawbacks, we performed single CpG resolution mapping of 5hmC in U937 cells to monitor changes after treatment with curcumin (5 µM) or DMC (1 and 2 µM) for 48 h, as described in the methods. Data analysis was performed at three levels: single CpG dinucleotide analysis, CpG island (CpGi) analysis, and gene promoter analysis.
For single CpG dinucleotide analysis, curcumin increased 5hmC at 25,471 CpG sites (positive values) and decreased 5hmC (negative values) at 20,517 CpG sites (indicating active demethylation) (Figure 4a). On the other hand, DMC (1 µM) similarly increased 5hmC at 26,394 CpG sites and decreased 5hmC at 22,964 CpG sites (Figure 4b). Increasing the concentration of DMC (2 µM) shifted the majority of 5hmC changes to negative values (19,072 vs. 7485), indicating an increase in active demethylation due to increased TET activity (Figure 4c). Furthermore, comparing the total number of CpG sites (both increases and decreases in 5hmC) between 1 µM and 2 µM DMC (Figure 4b,c) showed a decrease in the total number of CpG sites affected in the 2 µM concentration (26,557 versus 49,358 CpG sites). This difference is mainly attributed to the cytotoxicity of the higher concentration of DMC.
Both curcumin and DMC induced quite similar 5hmC increases at different genomic regions, where 44% and 7% of the increase in 5hmC happened at promoter regions and exons, respectively (Figure 5, upper panel). Similarly, 41–42% and 12% of the increase in 5hmC by both drugs occurred within CpG islands and shores, respectively (Figure 5, lower panel).
Active demethylation by both drugs, as represented by a decrease in 5hmC, was also observed in promoter regions and exons, where 41–42% of the total decrease in 5hmC occurred in promoter regions and 6–7% occurred in gene exons (Supplementary Figure S2). Moreover, 36–37% of active demethylation by both drugs occurred within CpG islands (Supplementary Figure S2).
Analysis of CpGi showed that curcumin increased 5hmC in 47 CpGis, while DMC increased 5hmC in 46 CpGis (Figure 6a and Figure 6b, respectively). Among these CpGis, four common CpGis showed an increase in 5hmC by both drugs. These four CpGis are located within the promoter region (≤1000 bp upstream TTS) of the following genes: DKKL1, SLFN12L, IKZF4, and RNA5S1. On the other hand, active demethylation was observed in 31 CpGis by both drugs (Figure 6a,b). Among these 31 CpGis, 17 common CpGis were actively demethylated by both drugs. Overall, 5 of these 17 CpGis were located in the promoter region of the following genes: LINC02610, ZNF222, WASIR1, GCFC2, and RNVU1-14.
Promoter region analysis of 5hmC in leukemia cells treated with curcumin or DMC showed significant increases and decreases in 5hmC in several gene promoters. Curcumin showed an increase in promoter 5hmC in 194 genes and a decrease (active demethylation) in 154 genes (Figure 6c). Similarly, DMC showed an increase in promoter 5hmC in 173 genes and a decrease in 171 gene promoters (Figure 6d). Among these genes, 34 and 29 leukemia-related genes showed promoter active demethylation by curcumin (Table 1) and DMC (Table 2), respectively. Supplementary Tables S1 and S2 show the list of leukemia-related genes with promoter 5hmC increase after DMC and curcumin treatment, respectively.
2.5. KEGG Pathway Enrichment Analysis
The KEGG database was used to map the set of genes that showed significant changes in 5hmC to signaling pathways. Both curcumin and DMC modulated 5hmC in a set of genes that was mapped to common signaling pathways related to AML and leukemia, like Wnt signaling, Hippo signaling, Rap1 signaling, cAMP signaling, and signaling pathways regulating pluripotency of stem cells (Supplementary Figure S3).
3. Discussion
In this manuscript, we monitored the effect of curcumin and its metabolically stable analog DMC on the enzymatic activity and transcription of different TET isoforms and how the increased TET activity impacted 5hmC changes genome-wide at the single CpG level. We used oxidative bisulfite sequencing to differentiate between 5mC and 5hmC. Both drugs increased the conversion of 5mC into 5hmC and also the active demethylation of 5hmC into cytosine. Increasing the concentration of DMC induced a significant increase in active demethylation compared to the 5hmC increase, which is consistent with the observed dose-dependent increase in TET activity by DMC. Both drugs induced active demethylation and increased 5hmC at the promoters of several leukemia-related genes. These 5hmC changes could reverse or suppress epigenetic silencing induced by DNA methylation and contribute to the antitumor effect of curcuminoids.
Initially, 5hmC was considered a passive intermediate in the process of active demethylation, but later, it was recognized as a bona fide dynamic epigenetic mark that affects gene expression, hematopoiesis, and stem cell pluripotency [32,33]. In contrast to 5mC, 5hmC is more enriched in euchromatin, with active histone marks (like H3K4me3 and H3K27ac) and actively transcribed genes. Oxidation of 5mC into 5hmC inhibits the binding of methyl-CpG-binding proteins (MBD1, MBD2, and MBD4) and the recruitment of gene repression complexes. Indeed, 5hmC is often detected in promoters and enhancers of actively transcribed genes, and its presence is associated with increased gene expression. Moreover, specific proteins like MBD3 and UHRF2 act as epigenetic readers for 5hmC and contribute to transcriptional activation [34]. Loss of UHRF2 expression has been implicated in neoplasia [35], where alterations in its ability to recognize and interpret 5hmC may contribute to promoter hypermethylation, disrupted gene expression, and tumor progression. In this study, we detected significant increases in 5hmC and active DNA demethylation after treatment with curcuminoids at promoters, CpGis, exons, introns, and intergenic regions. These 5hmC changes could be valuable in leukemia subtypes characterized by promoter hypermethylation by restoring the expression of genes silenced by DNA methylation. Nonetheless, a drawback of the observed 5hmC increase and active demethylation is the possible unintended enhanced expression of oncogenes, which is undesirable. For instance, genes like ANGPT1, PES1, and ZNF222 demonstrated promoter active demethylation by both compounds and are known for promoting cell survival and proliferation of different types of leukemia [36,37,38]. This drawback was also observed with other classes of anticancer drugs, like DNA hypomethylating agents and HDAC inhibitors, where targeted locus-specific delivery is not feasible. Predicting the net effect of the mixed active demethylation of tumor suppressor genes and oncogenes on tumor progression requires in vivo animal and clinical testing.
TET enzymes are essential for the oxidation of 5mC into 5hmC and the sequential oxidative steps of active demethylation [22]. Loss-of-function mutations in TET2 were frequently observed in myeloid malignancies and resulted in decreased 5hmC levels, increased 5mC levels, and impaired hematopoietic differentiation [39]. Our study suggests that the use of curcuminoids in leukemia cells with wild-type or partially functioning TET2 could increase 5hmC levels and induce active demethylation of silenced promoter hypermethylated genes. Similarly, IDH1 and IDH2 gain-of-function mutations in leukemia lead to the accumulation of the competitive inhibitor of TET enzymes, 2-hydroxyglutarate (2-HG), with consequent DNA hypermethylation. The observed increase in the transcription and the activity of TET enzymes by curcuminoids could overcome the competitive inhibition by the oncometabolite 2-HG. There are several mechanisms that could contribute to the observed enhanced TET transcription. Increased transcription factor binding, histone modifications, chromatin remodeling, enhancer activation, and promoter–enhancer looping are possible mechanisms that could contribute to enhanced TET transcription. We previously reported that both curcumin and DMC inhibited the methylation of the transcriptional repressor marks H3K9 and H3K27 [3]. Direct promoter/transcription factor binding is also a possible mechanism, but it would be problematic to investigate due to the promiscuous non-specific binding of curcumin [40].
Previous studies demonstrated a relationship between chemotherapy resistance and 5hmC levels. Chemoresistance in hepatocellular carcinoma was associated with reduced 5hmC levels [41]. Moreover, AML patients treated with 5-azacytidine in combination with standard chemotherapy showed improved outcomes in patients with favorable 5hmC patterns [42]. Accordingly, the combination of curcuminoids with chemotherapy could overcome chemoresistance associated with reduced 5hmC levels through activation of TET enzymes and increasing 5hmC levels.
Our results indicate that the effect of curcumin and DMC on 5hmC distribution was not identical but shared several similarities. These differences were expected due to differences in dosing, metabolic stability, and TET activation. In support of that, increasing the concentration of DMC was also associated with changes in 5hmC distribution compared to the lower concentration. Nevertheless, comparing the overall changes in 5hmC (active demethylation and increase in 5hmC) at promoters, CpGis, shores, introns, and exons showed quite similar changes by both drugs. To the best of our knowledge, there are no previous reports on the effect of DMC on 5hmC to compare with our findings. However, a few studies reported the effect of curcumin on 5hmC in other tissues. A previous study demonstrated a similar effect of curcumin on 5hmC in cultured vascular smooth muscle cells (VSMCs), where curcumin treatment blocked the effect of the CCDC80-induced reduction in TET2 and 5hmC by restoring the expression of TET2 in VSMCs [43]. Moreover, curcumin was shown to upregulate the expression of both the TET2 and TET3 enzymes in gastric cancer cell lines, with consequent active demethylation and re-expression of the tumor suppressor RB1 [44].
DMC differs from curcumin in chemical structure by replacing the two phenolic hydroxyl groups in curcumin with two methoxy groups. Accordingly, DMC is less vulnerable to phase-II conjugation and oxidative degradation [5]. The higher metabolic stability of DMC is concordant with the comparable 5hmC changes induced by both curcumin and DMC, using a DMC concentration that is five times less than that of curcumin. Moreover, DMC induced significantly higher apoptosis induction in leukemia cells compared to curcumin at equimolar concentrations [5]. Across other types of cancer, DMC showed higher potency compared to curcumin at an equimolar dose. For instance, in breast cancer cells, DMC induced paraptosis-like cell death and induced more potent inhibition of the proteasome compared to curcumin [45]. The epigenetic changes induced by both compounds shared some similarities but were not identical. Although both compounds increased the activity of TET enzymes, global 5hmC, and active demethylation, the affected genes were different, with minor overlapping. Similarly, mass spectrometry analysis of a variety of histone post-translational modifications induced by both drugs showed major differences despite similar inhibition of histone lysine methyltransferase (HKMT) and activation of different histone lysine demethylases, like JARID and LSD1 [3].
In this study, we used two different approaches—the global and locus-specific approaches—to study the impact of curcumin and DMC on 5hmC. The global approach was used to compare the global 5hmC changes across different concentrations of both compounds and leukemia cell lines. Despite its affordability and ability to detect 5hmC changes across different concentrations, it did not capture locus-specific 5hmC changes and just provided the net change in 5hmC without discriminating between 5hmC-decreased and 5hmC-increased loci. Accordingly, 5hmC mapping at the single CpG level using oxidative bisulfite sequencing was used to overcome this drawback.
CpGi analysis of 5hmC revealed both increases in 5hmC and active demethylation in the promoter region of several leukemia-related genes after curcuminoid treatment. For instance, the BEX1 promoter showed an increase in 5hmC and was reported to act as a novel suppressor of oncogenic FLT3-ITD-driven AML [46]. Similarly, the Wnt signaling pathway inhibitor sFRP4 showed an increase in 5hmC and was reported to be promoter-methylated in chronic myeloid leukemia. PRKX also showed an increase in 5hmC and was shown to play an essential role in myeloid cell differentiation, which is usually blocked or impaired in AML [47]. Other genes like ZNF292, PRDM1, and CDH11 demonstrated promoter active demethylation and were previously linked to different leukemia types, where PRDM1 and CDH11 were reported to be frequently epigenetically silenced in these tumors [48,49].
In conclusion, curcuminoids activated and increased the expression of all the TET isoforms in leukemia cells, with a consequent increase in 5hmC and active demethylation of several leukemia-related genes and genes mapped to common signaling pathways related to leukemia. Our findings provide a rationale for future preclinical testing of the combination of curcuminoids with DNA hypomethylating agents to harness their combined effect on active demethylation and methylation reversal of hypermethylated genes.
4. Materials and Methods
4.1. Chemicals
Curcumin analytical standard grade (Sigma-Aldrich, Milwaukee, WI, USA) and DMC (Cayman, Ann Arbor, MI, USA) were both dissolved in DMSO as a 10 mM stock solution and then aliquoted and stored in the dark at −80 °C.
4.2. Cell Culture and Treatments
Human promyeloid leukemia (HL60) and human monocytic leukemia (U937) were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA) and cultured in RPMI-1640 medium (Sigma-Aldrich, Milwaukee, WI, USA) containing 10% fetal bovine serum (FBS) and 2.5 mM L-glutamine. The cells were maintained in a humidified incubator with 5% CO_2_ at 37 °C. The cells were treated with different micromolar concentrations of DMC, curcumin, or DMSO (control), followed by nuclear protein extractions to quantify global 5hmC or measure TET enzymatic activity, as described below.
4.3. Nuclear Protein Extraction
Nuclear extracts from the treated and untreated HL60 and U937 leukemia cells were prepared using an EpiQuikTM nuclear extraction kit (Epigentek, Farmingdale, NY, USA) as per the manufacturer’s protocol. Approximately 2 × 10^6^ cells were collected and centrifuged at 1000 rpm for 5 min. The nuclear extract was prepared and stored at −80 °C until use. The protein concentration of the nuclear extract was quantitated using a Quick Start Bradford Protein Assay (BioRad, Hercules, CA, USA).
4.4. Global DNA Hydroxymethylation Quantification
Fluorometric detection of global DNA 5-hydroxymethylation of cytosine (5hmC) was performed using a commercially available kit (Epigentek, NY, USA). Briefly, genomic DNA was extracted from HL60 and U937 cells treated with curcumin, DMC, or DMSO (control) and incubated in microplate wells treated to have high affinity for hydroxymethylated DNA but not methylated or unmethylated DNA. The bound 5hmC was quantified using a capture antibody followed by a detection antibody that was fluorescence-labeled.
4.5. TET Activity Quantification Assay
A fluorometric assay (Abcam, Waltham, MA, USA) was used to detect the activity of the TET isoforms using nuclear extracts from HL60 and U937 leukemia cells. Briefly, microplate wells coated with a methylated substrate were incubated with nuclear extracts from the leukemia cells treated with graded concentrations of curcumin, DMC, or DMSO (control) for 48 h. The methylated substrate was converted into a hydroxymethylated substrate, which is proportional to the enzymatic activity and can be quantified by incubation with a specific capture antibody followed by a fluorescent-labeled detection antibody. The activity for the TET enzymes was determined using the following formula:
where * Incubation time = 120 min and RFU = Relative Fluorescence Unit.
4.6. Real-Time Quantitative RT-PCR
Total RNA was extracted from the treated and control cells using RNeasy columns (Qiagen, Chatsworth, CA, USA). One-step qRT-PCR was performed as recommended by the manufacturer of a commercially available kit (ThermoFisher, Waltham, MA, USA) using a QuantStudio 5 Real-Time thermal cycler. The following primers were used for TET1: forward 5′cacaccagctccactgaaga3′ and reverse 5′ctccatcacaggagcagaca3′; for TET2: forward 5′ttggacttctgtgctcatgc3′ and reverse 5′ctcctgagcttccacactcc3′; and for TET3: forward 5′tgtgacgttgtcgagagagg3′ and reverse 5′attcccctctgtgtgtcctg3′. GAPDH was used as a loading control.
4.7. Single CpG Resolution Analysis of 5hmC Using Oxidative Bisulfite Sequencing
The ovation reduced representation bisulfite sequencing (RRBS) Methyl-Seq System (Nugen, Tecan Genomics, Redwood City, CA, USA) was used. RRBS is a technique that reduces the amount of sequencing compared to whole-genome sequencing and utilizes the methylation-insensitive restriction enzyme MspI, which recognizes CCGG [50]. Regular bisulfite sequencing is not capable of differentiating 5-methylcytosine (5mC) from 5hmC because cytosine will not be converted into thymine in both cases. In order to differentiate between 5mC and 5hmC, each sample was divided into two equal portions. The first portion (mock sample) underwent regular bisulfite treatment, followed by next-generation sequencing (provides a total of 5mC and 5hmC). The second portion (oxidative sample) underwent an oxidation step according to the manufacturer’s instructions to convert 5hmC into 5fC, followed by bisulfite treatment and next-generation sequencing. The conversion of 5hmC into 5fC allows for the conversion of cytosine into thymine after bisulfite treatment, and, consequently, comparing the cytosines in the first and second portions differentiates the 5hmC mark from the 5mC.
Paired-end bisulfite sequencing reads were pre-processed with FastQC v0.11.9, cutadapt v2.10, and Trim Galore v0.6.10 python functions to remove adapter content and simultaneously trim low-quality ends and filter for reads below Q20. The additional diversity adaptors were trimmed using a NuMetRRBS python script using recommended settings for paired-end reads [51]. The hg38 version of the human genome was used to align the reads. Before the alignment, the genome was converted to bisulfite-compatible versions using Bismark v0.22.3. The reads were aligned to the genome sequences using bowtie2 v2.4.4 via the Bismark scripts using default parameters. Methylation calling and extraction of methylation levels were performed using a Bismark methylation extractor. The methylation data (CpG sites) from all samples were merged to find the common sites present in all samples. After filtering for sites with <2 standard deviations and known C2T mutation sites, there were about 288,097 CpG sites left for further analysis. Analysis of CpG methylation with respect to genomic features was performed using the methylKit v1.34.0 and methylGSA v1.20.0 R packages.
Initially, the mock and oxidative samples were merged to obtain the common CpG sites. The difference in the methylation level at CpG sites between the mock and oxidative samples was taken as the hydroxymethylation level. The differential hydroxymethylation levels between the control and treatments were computed using the MethylKit v1.34.0 R package. Differential analysis was performed at 4 levels: base pairs, CpG islands, promoters, and 1000 bp tiles. The R package methylGSA v1.20.0, was used to compute Gene Set Enrichment (GSA) and pathway enrichment for the significantly different sites.
4.8. Statistical Analyses
All cell culture experiments were performed in triplicate. Data represent the mean ± standard deviation (S.D.). Multiple comparisons between the effects of different concentrations of curcumin or DMC on the mRNA expression of TET proteins were performed using one-way ANOVA followed by post hoc analysis using the Bonferroni test.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Rao C.V. Rivenson A. Simi B. Reddy B.S. Chemoprevention of colon carcinogenesis by dietary curcumin, a naturally occurring plant phenolic compound Cancer Res.1995552592667812955 · pubmed ↗
- 2Sudarshan K. Yarlagadda S. Sengupta S. Recent Advances in the Synthesis of Diarylheptanoids Chem. Asian J.202419 e 20240038010.1002/asia.20240038038744677 · doi ↗ · pubmed ↗
- 3Sawesi S. Malkaram S.A. Abd Elmageed Z.Y. Fandy T.E. Modulation of the activity of histone lysine methyltransferases and demethylases by curcumin analog in leukaemia cells J. Cell Mol. Med.2022265624563310.1111/jcmm.1758936300880 PMC 9667515 · doi ↗ · pubmed ↗
- 4Uddin M.G. Fandy T.E. DNA methylation inhibitors: Retrospective and perspective view Adv. Cancer Res.202115220522310.1016/bs.acr.2021.03.00734353438 PMC 10275377 · doi ↗ · pubmed ↗
- 5Hassan H.E. Carlson S. Abdallah I. Buttolph T. Glass K.C. Fandy T.E. Curcumin and dimethoxycurcumin induced epigenetic changes in leukemia cells Pharm. Res.20153286387510.1007/s 11095-014-1502-425186441 PMC 11173366 · doi ↗ · pubmed ↗
- 6Amalraj A. Pius A. Gopi S. Gopi S. Biological activities of curcuminoids, other biomolecules from turmeric and their derivatives—A review J. Tradit. Complement. Med.2017720523310.1016/j.jtcme.2016.05.00528417091 PMC 5388087 · doi ↗ · pubmed ↗
- 7Jakubczyk K. Druzga A. Katarzyna J. Skonieczna-Zydecka K. Antioxidant Potential of Curcumin-A Meta-Analysis of Randomized Clinical Trials Antioxidants 20209109210.3390/antiox 911109233172016 PMC 7694612 · doi ↗ · pubmed ↗
- 8Peng Y. Ao M. Dong B. Jiang Y. Yu L. Chen Z. Hu C. Xu R. Anti-Inflammatory Effects of Curcumin in the Inflammatory Diseases: Status, Limitations and Countermeasures Drug Des. Dev. Ther.2021154503452510.2147/DDDT.S 327378 PMC 857202734754179 · doi ↗ · pubmed ↗