Genetic Characterisation of Closely Related Lactococcus lactis Strains Used in Dairy Starter Cultures
Yuliya E. Uvarova, Tamara M. Khlebodarova, Asya R. Vasilieva, Aleksandra A. Shipova, Vladimir N. Babenko, Andrey V. Zadorozhny, Nikolay M. Slynko, Natalia V. Bogacheva, Ekaterina Y. Bukatich, Valeriya N. Shlyakhtun, Anton V. Korzhuk, Elena Y. Pavlova, Danil O. Chesnokov

TL;DR
This study explores the genetic makeup of Lactococcus lactis strains in a Russian artisanal cheese starter, revealing three distinct strains and their lack of typical aroma compound production.
Contribution
The study provides a detailed genetic and functional characterization of closely related Lactococcus lactis strains in a specific cheese starter.
Findings
Metagenomic analysis identified three Lactococcus lactis genomes in the cheese starter.
Isolates were grouped into three distinct strains based on rpoB gene analysis.
None of the strains produced diacetyl or CO2, characteristic of L. l. subsp. diacetilactis.
Abstract
The complex microbiota of cheese starters plays a key role in determining the structure and flavour of the final product, primarily through their acid-forming capacity, protease activity, and exopolysaccharide synthesis. However, the specific microbial communities underlying the unique qualities of artisanal cheeses remain poorly understood. This study presents the microbiological and molecular genetic characterisation of the microbiome isolated from an artisanal cheese starter in Kosh-Agach, Altai, Russia. Metagenomic analysis of this starter revealed the presence of three bacterial genomes corresponding to those of Lactococcus lactis. Pure cultures from this starter were obtained by sequential subculture, and seventeen colonies displaying distinct characteristics on differential media were selected. Genome sequencing was performed for each colony. Bioinformatic analysis based on the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
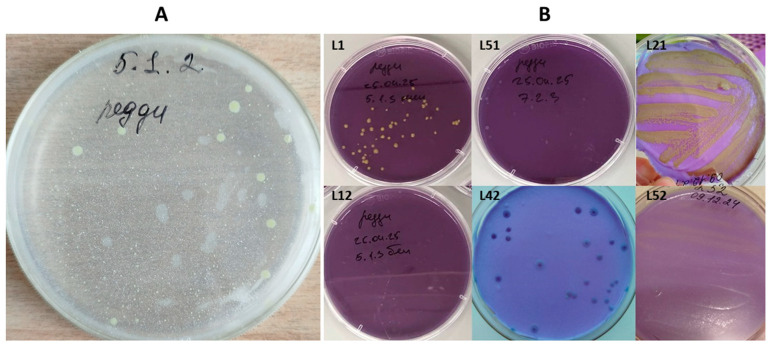 Figure 1
Figure 1 Figure 2
Figure 2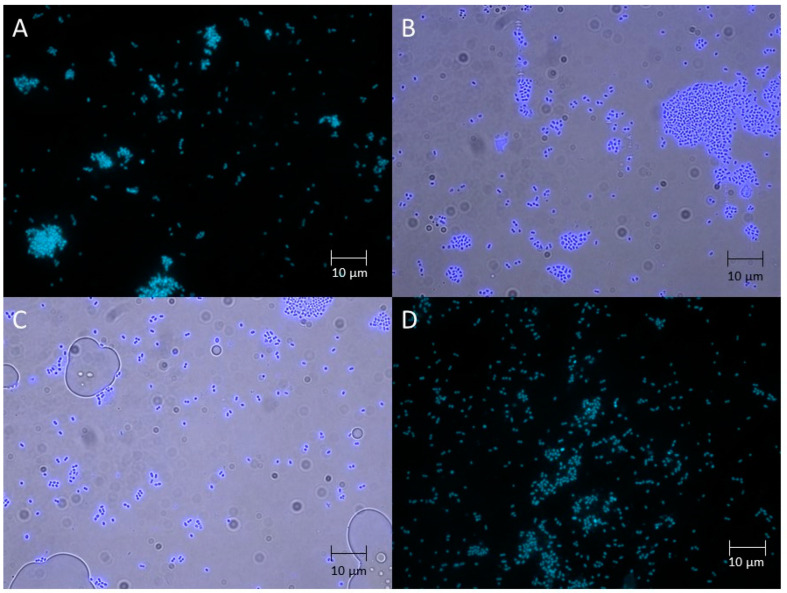 Figure 3
Figure 3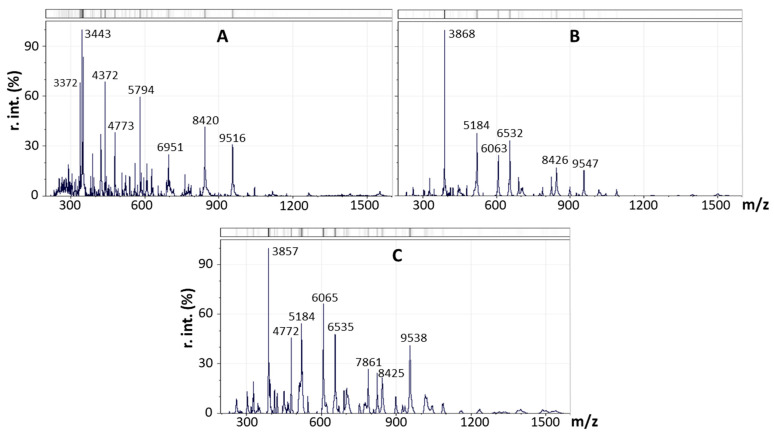 Figure 4
Figure 4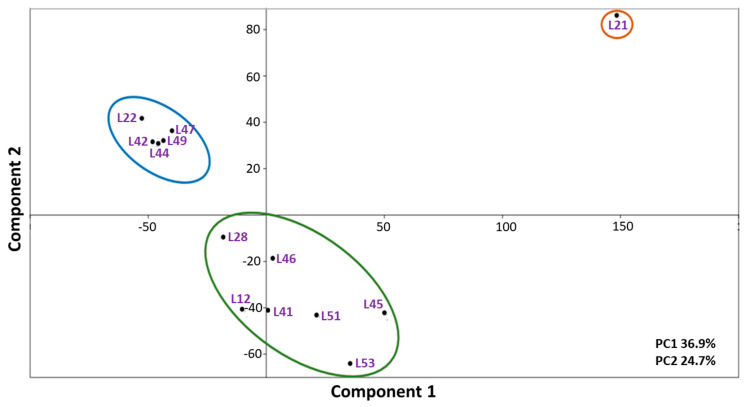 Figure 5
Figure 5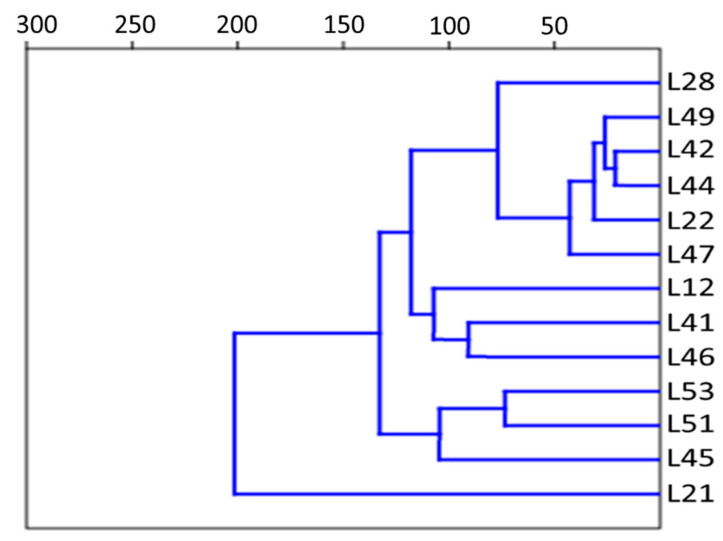 Figure 6
Figure 6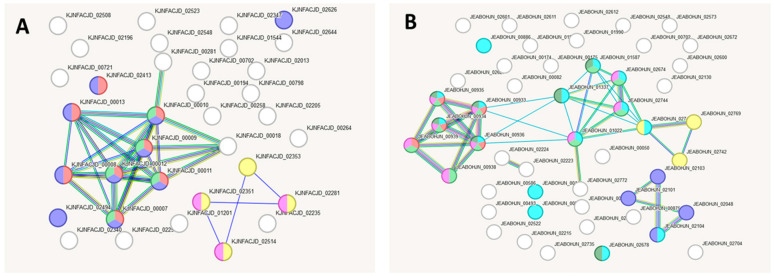 Figure 7
Figure 7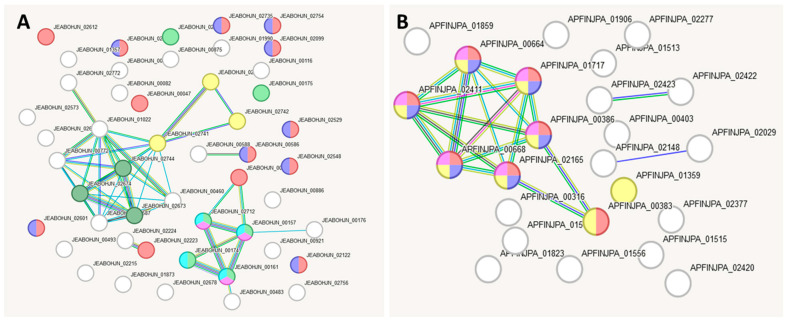 Figure 8
Figure 8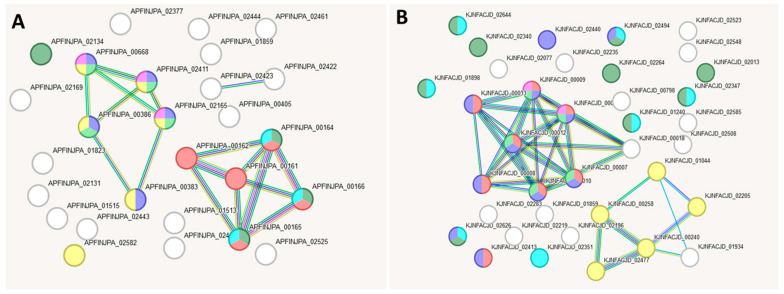 Figure 9
Figure 9- —Ministry of Science and Higher Education
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Milk Quality and Mastitis in Dairy Cows · Biopolymer Synthesis and Applications
1. Introduction
Thousands of industrial starters derived from lactic acid bacteria are used worldwide in the production of fermented dairy products. Private farms often use starters derived from unique local bacterial flora, the microbiological structure of which is currently being actively studied [1,2,3,4,5]. When characterising a recently isolated lactic acid bacteria culture, determining its taxonomic position and microbiological characteristics does not fully reflect its industrial potential. Significant intraspecific diversity of lactic acid bacteria is used to produce products with various flavours, aromas, and textural properties characteristic of fermented milk products in different regions. Metagenomic analysis of the microflora of artisanal lactic acid products revealed general trends in the use of specific microorganism species. According to these data, Lactococcus predominates in cream and solid products obtained by fermentation of raw milk at room temperature, whereas Lactobacillus predominates in beverages. Furthermore, it has been shown that Streptococcus species predominate in food products made from baked or pasteurised milk and fermented at higher temperatures [1]. The main objective of our study was to investigate the strain composition of the bacterial consortium of the Kraft cheese starter culture from the village of Kosh-Agach, Altai Republic, using metagenomic analysis. Metagenomic analysis of the Kraft starter microflora allows us to determine these characteristics. Genome sequencing of individual strains allows us to identify key gene differences and, based on bioinformatics analysis, construct genetic networks involved in shaping the quality of fermented milk products. Typically, the highly polymorphic species Lactococcus lactis is used in cheese starters, which has five subspecies: L. l. cremoris, L. l. hordniae, L. l. lactis, L. l. lactis bv. diacetilactis, and L. l. tructae. Of these, L. l. lactis, L. l. lactis bv. L. diacetilactis, and L. l. cremoris exhibit the closest taxonomic relationship [6]. Traditionally, these bacteria have been differentiated using microbiological methods, since microscopic analysis is often insufficient to distinguish between these subspecies of Lactococcus lactis. For example, microscopic specimens of L. l. lactis and L. l. diacetilactis are virtually indistinguishable. Their cells are spherical or oval, 0.5–1.5 μm, and form pairs or short chains [7]. However, they differ from L. l. cremoris, which typically forms longer chains. At the same time, the enzymatic properties of L. l. lactis and L. l. diacetilactis differ significantly. For example, active strains of L. l. lactis coagulate milk within 4–6 h, forming a smooth, dense curd and reaching maximum acidity (125 °T) after 5–7 days of development in milk. Strains of this subspecies break down arginine to form ammonia and are used in starters for cottage cheese, sour cream, butter, and low-temperature cheeses. Some strains of L. lactis subsp. lactis produce the bacteriocin nisin, which exhibits antagonistic activity against most Gram-positive bacteria [8]. Lactococcus lactis subsp. lactis bv. diacetilactis, unlike L. l. lactis, is an aromatic streptococcus with relatively weak acid-forming activity. Its strains coagulate milk within 16–18 h, with maximum acidity not exceeding 70–100 °T. The resulting curd often contains gas bubbles (CO_2_) and has a pleasant, characteristic aroma due to the accumulation of diacetyl, as L. l. Lactococcus lactis subsp. cremoris breaks down lactose and citrates, forming diacetyl, carbon dioxide, and acetoin [9]. These strains are widely used in starters for most fermented dairy products [7]. Lactococcus lactis subsp. cremoris is characterised as a cream-type streptococcus. Its strains coagulate milk relatively quickly, within 6–8 h, forming a dense curd with a slightly viscous or creamy consistency, which is explained by the ability of L. l. cremoris to synthesise polysaccharides. However, the maximum acidity of milk does not exceed 110–115 °T. Unlike L. l. lactis, L. l. strains of cremoris do not metabolise arginine, although some can produce diacetyl depending on the concentration of citric acid in the medium [10]. These strains are used in starters for sour cream, butter, and other fermented dairy products. Some strains of this subspecies can also synthesise the bacteriocin lactococcin [11]. This study examines the microbiological and molecular genetic characteristics of lactic acid bacteria belonging to the species Lactococcus lactis, isolated from a farmer’s cheese starter culture, designated as the lakt1p sample. The functional properties of the lakt1p consortium are also discussed.
2. Results and Discussion
2.1. Microbiological Characterisation of Pure Cultures Isolated from the lakt1p Starter Culture
Pure cultures were isolated from the lakt1p cheese starter culture sample from a private farm in the Kosh-Agach district of the Altai Republic by sequential subculturing. Seventeen colonies exhibiting distinct morphological characteristics on differential media were selected from these. On Reddy’s differential agar, some colonies exhibited a cream colour, corresponding microbiologically to L. l. subsp. lactis and L. l. subsp. diacetilactis, while others were yellow, corresponding to L. l. subsp. cremoris (Figure 1A).
The yellow colouration observed in L. l. subsp. cremoris colonies on Reddy’s differential agar is associated with acidification of the medium, as the bromocresol purple indicator acts as a pH-sensitive dye, turning yellow at pH values below 5.2 and purple at pH values above 6.8.
For further differentiation, we used test media based on modified Reddy agar. On these media, L. lactis subsp. diacetilactis hydrolyses citrate, forming gas bubbles and turning colonies purple (Figure 1B—L51, L12, L42), whereas L. cremoris colours colonies yellow (Figure 1B—L1, L21, L52). However, as shown in Figure 1B, none of the cultures tested or any colony in the consortia inoculum (Figure 1A) produced gas bubbles on modified Reddy’s agar. This finding indicates that the cultures under analysis cannot utilise citrate, a trait uncharacteristic of L. lactis subsp. lactis bv. diacetilactis.
Citrate utilisation ability was assessed by performing deep inoculations on a medium containing milk hydrolysate and citrate. However, again, none of the cultures demonstrated this ability.
Taxonomic affiliation was determined through a microbiological characterisation of the cultures. This process involved the evaluation of the pure strain activity by measuring the time required for pasteurised milk to curdle after inoculation with 5% of a pure culture and assessing acid-forming activity and curd characteristics. The results showed that the cultures coagulated pasteurised milk within 18–25 h. The acidity of all the samples ranged from 40 °T (pale pink colour, samples L1, L21, L34, and L52) to 80 °T (bright pink colour, samples L42, L22, and L28) (Figure 2A), which corresponds to L. lactis subsp. lactis bv. diacetilactis. However, in all the samples, the curds had a similar texture: they were dense, without gas bubbles and had a consistency of thick sour cream, and upon heating, the curds did not rise but sank to the bottom, indicating the absence of carbon dioxide production (Figure 2B), which is uncharacteristic for L. lactis subsp. lactis bv. diacetilactis. Moreover, none of the cultures produced diacetyl, the main aroma compound of L. l. subsp. diacetilactis. The presence of a pink hue within 10 min is indicative of diacetyl within the starter. In all analysed cultures, the time for the pink colour to appear varied from 52 min to 1.5 h (Figure 2C). Consequently, none of the starter cultures analysed contained diacetyl.
Fluorescent microscopy of the consortium and pure cultures demonstrated that the samples contained spherical cells occurring in groups of 2–4, occasionally forming short chains, approximately 1–1.5 µm in diameter, non-motile and non-spore-forming (Figure 3), which is morphologically consistent with L. lactis and L. lactis subsp. diacetilactis. Most cells were stained with DAPI and exhibited intense fluorescence at 460 nm.
2.2. Analysis of the Metagenome of the Microbiological Consortium in Sample lakt1p
Metagenomic analysis of sample lakt1p was performed to identify the microbial consortium present in the lakt1p cheese starter culture and to facilitate the subsequent isolation of pure cultures. Analysis of the DNA sequencing data obtained from lakt1p revealed that the contig sequences belonged to the genus Lactococcus. At the same time, it proved challenging to identify the taxonomic classification using 16S rRNA within the metagenomic data set. Table 1 shows the results of mapping the reads from this sample to reference genomes.
In total, 85.9% of reads were mapped to reference genomes of L. lactis subsp. lactis bv. diacetilactis, with 13% assigned to L. lactis subsp. cremoris and approximately 1% to L. lactis subsp. lactis.
It should be noted that L. lactis subsp. lactis and L. lactis subsp. lactis bv. diacetilactis are closely related strains of the same species and therefore have very similar genomes. When using short-read metagenomic sequencing, the low density of genetic differences between these strains makes them difficult to distinguish. Likewise, the contig-binning software cannot distinguish L. lactis from L. lactis bv. diacetilactis. Given the above, only two rather than three distinct genomes were recovered, corresponding to L. cremoris subsp. cremoris and L. lactis subsp. lactis. It cannot be excluded that the lakt1p sample contains only two strains, L. cremoris and L. lactis subsp. Lactis, which differ from the specific reference genomes chosen (L. lactis subsp. lactis and L. lactis subsp. lactis bv. diacetilactis).
2.3. Bioinformatic Analysis of Sequenced Genomes
Seventeen genomic assemblies were obtained and analysed from individual cultures of the lakt1p consortium. Clustering allowed the identification of three clusters based on the rpoB gene, one cluster based on the gyrB gene, and two clusters according to 16S rRNA. The pairwise ANI comparisons demonstrated 99.9% similarity between assemblies within the same rpoB cluster. However, assemblies from different clusters had ANI values ranging from 99.03% to 99.37%. When compared with reference genomes for all the assemblies, the following ANI values were determined: 87.3–87.5% relative to L. cremoris subsp. cremoris KW2 (GCF_000468955.1); 98–98.3% relative to L. lactis subsp. lactis strain 14B4 (GCF_003176835.1); and 99.2–99.6% relative to L. lactis subsp. lactis bv. diacetilactis strain S50 (GCF_003627395.2). These findings indicate that the 17 genome assemblies form three rpoB-based clusters (Table 2), each representing a distinct strain of L. lactis subsp. lactis bv. diacetilactis.
2.4. Mass Spectrometric Analysis of Strains After Separation of the lakt1p Consortium
Mass spectrometric profiling of 13 cultures from the lakt1p consortium enabled their separation into three groups: (1) sample L21; (2) a group of five samples—L22, L42, L44, L47 and L49; (3) a group of seven samples—L12, L28, L41, L45, L46, L51, and L53. The averaged spectra of three experimental repeats for samples L21, L44 and L51 are presented, respectively, in Figure 4A–C.
Samples L22, L42, L44, L47 and L49 (Figure 4B) exhibited similar compositions and intensities of m/z peaks at 3868, 5184, 6063, 6532, 8426 and 9547. The group comprising L12, L28, L41, L45, L46, L51, and L53 contained several intense peaks shared with the previous group, as well as peaks characteristic of this group alone (m/z 3857, 4772, 7861, 9538). Sample L21 contained several additional intense m/z signals, including 3372, 4372, 5794, and 6951. An analysis of the mass spectrograms of the lakt1p consortium cultures, based on the m/z values of all identified mass peaks and their intensities, using principal component analysis, is presented in Figure 5. The analysed samples are clearly divided into three distinct groups.
Further conclusions regarding the similarities and differences between the studied cultures will be made after evaluating the results using other methods (microbiological and genetic).
Figure 6 presents a dendrogram based on all identified m/z values, determined using the UPGMA method (Unweighted Pair Group Method with Arithmetic Mean). The analysis of the sample has revealed three clear groupings. The cophenetic correlation coefficient was high (0.9691), indicating that the dendrogram accurately reflects the similarity between the samples.
Hence, mass spectrometric analysis revealed three distinct bacterial groups within the lakt1p consortium.
Table 3 summarises the results of the microbiological, genetic, and mass spectrometric analyses of 17 pure cultures derived from the lact1p consortium.
As shown, microbiological analysis on Reddy’s differential medium, along with genomic and mass spectrometric analyses, consistently divided the lact1p pure culture isolates into three clusters. According to phenotypic characteristics based on colony colouration, Cluster 1 was assigned to L. lactis cremoris, Cluster 2 to L. lactis lactis, and Cluster 3 to L. lactis diacetilactis. However, genetic analysis indicated that the three clusters defined by the rpoB gene represent distinct strains of L. lactis diacetilactis. Microscopic analysis also placed all the strains within L. lactis diacetilactis, as the cells were arranged predominantly in pairs (diplococci), virtually without forming chains. Likewise, based on the rate of milk coagulation, the nature of the curd, and final acidity of the samples during milk fermentation, these strains were found to exhibit the traits of L. lactis diacetilactis. However, none of the strains, including those identified as L. lactis diacetilactis on Reddy differential medium, produced the key aroma-forming compounds characteristic of L. lactis diacetilactis, such as diacetyl and CO_2_.
Given the uncertainty regarding the precise taxonomic assignment of the lact1p strains, we sought to identify distinguishing genomic features of the lact1p metagenome. In particular, it is worth performing a comparison with the reference genome of Lactococcus lactis subsp. lactis and with a synthetic metagenome composed of reference sequences from L. lactis cremoris, L. lactis lactis and L. lactis diacetilactis. Special attention should be paid to genes involved in the synthesis of enzymes, pheromones, exopolysaccharides and other compounds responsible for the flavour, aroma and textural properties of the final product in the lakt1p consortium strains.
Furthermore, the division of the lakt1p consortium into three clusters, based on genomic, proteomic, and phenotypic characteristics observed in differential media, necessitates supplementary comparative bioinformatic analyses of genomic traits among the strains within these clusters. The following sections of the article address these questions in detail.
2.5. Comparison of Exopolysaccharide Synthesis Genes in the Metagenome of the lakt1p Starter Culture and the Synthetic Metagenome
As noted above, L. lactis subsp. cremoris is characterised as a “creamy” streptococcus, owing to its higher capacity for exopolysaccharide (EPS) synthesis and secretion compared with other L. lactis subspecies. Microbiological tests of individual colonies from the lakt1p starter on Reddy’s differential agar exhibited a yellow colouration, which, according to microbiological characteristics, is typical of L. l. cremoris (Figure 1A), suggesting that this subspecies may indeed be present in the lakt1p culture. This assumption is also consistent with the metagenomic analysis of lakt1p.
Exopolysaccharide synthesis in lactic acid bacteria is facilitated by an extensive pool of glycosyltransferases with a broad spectrum of sugar and linkage specificities. Exopolysaccharides secreted by lactic acid bacteria can be directly applied to achieve the desired viscosity, texture, and mouthfeel in food production, particularly due to their water-binding capacity, gel formation, flow behaviour and overall rheological performance of the final product [12]. Exopolysaccharides exhibit great structural diversity, and some provide specific biological activity and physiological functions, such as antioxidant activity [13], antimicrobial effects [14], and immunostimulatory effects [15], among others.
We compared the set of glycosyltransferases identified in the metagenome of the lakt1p sample and found that, when compared with the genome of L. lactis subsp. lactis, the glycosyltransferase repertoire was identical. However, comparison with a synthetic metagenome composed of reference sequences from L. l. cremoris, L. l. lactis and L. l. diacetilactis revealed that the lakt1p sample contains genes encoding glucosyltransferase, oligosaccharide 4-α-D-glucosyltransferase and poly(ribitol-phosphate) β-glucosyltransferase, which are absent from the annotated synthetic metagenome. This indicates certain genetic differences between the strains in the lakt1p consortium and the reference genomes of L. lactis cremoris and L. lactis diacetilactis.
2.6. Analysis of the Peptidase Spectrum in the lakt1p Metagenome and in the Isolated Cultures
Given the strong correlation between high acidification capacity of starter strains and the proteolytic activity of their proteinases [16,17], consideration should be given to the fact that lactic acid bacterial strains may differ substantially in both the spectrum and activity of their proteinases. Consequently, we analysed the peptidase repertoire in the lakt1p sample and in the reference genomes. The results are summarised in Table 4 and Table 5.
As shown in Table 4, the lakt1p metagenome shares 11 peptidases with the L. l. lactis reference genome. It also contains five peptidases that are not annotated in the reference genome, whereas the reference genome encodes eight peptidases that were not annotated in the lakt1p metagenome. Table 5 shows that the lakt1p metagenome shares 14 peptidases with the synthetic metagenome. The lakt1p sample also contains four peptidases absent from the annotations of L. l. cremoris, L. l. lactis and L. l. diacetilactis, while three reference species collectively possess ten peptidases that were not annotated in the lakt1p metagenome.
Genomic DNA analysis of 17 individual strains from the lakt1p consortium revealed that all 17 genomes encoded peptidases: Neutral endopeptidase (pepO_1, pepO_2), Dipeptidase A (pepDA_1, pepDA_2), Beta-Ala-Xaa dipeptidase (pepV), and Xaa-Pro dipeptidase (pepQ). Notably, only genomes from the third cluster (based on genomic analysis) encoded the peptidase PepF1 (EC 3.4.24, oligoendopeptidase F), which appears to be plasmid-borne. Differences observed between the isolated strains may be explained by plasmid loss in some strains, which could underlie the morphological variation.
Pheromones, exopolysaccharides and other compounds contribute to the flavour, aroma, and textural properties of the final fermented product. The presence of genes encoding these traits in the lakt1p lactococcal strains is summarised in Table 6. Shown are clusters identified through genome analysis.
The genomic analysis presented in Table 6 demonstrates that the individual strains of the lakt1p consortium have genes for most enzymes responsible for lactose utilisation, synthesis of exopolysaccharide precursors with diverse sugars, including glucose, galactose, and rhamnose. Only a single bacteriocin-encoding gene was identified: the lcnB gene for lactococcin B, which is present in clusters 2 and 3 and absent from cluster 1 strains. It has been reported that lactococcin B can be produced by some strains of Lactococcus lactis subsp. lactis [18] as well as by L. lactis subsp. cremoris [19], and thus does not allow subspecies identity of the lakt1p clusters to be determined unambiguously.
The exopolysaccharide synthesis gene cluster contains only the epsL gene. In Bacillus subtilis, this gene produces a phosphoglycosyltransferase. This enzyme initiates exopolysaccharide unit synthesis, utilising UDP-di-N-acetylbacillosamine as a phosphosaccharide donor [20]. These findings suggest that a larger EPS gene cluster may be located on plasmids.
2.7. Bioinformatic Analysis of Gene Interaction Networks in Strains L21, L42 and L51 from the lakt1p Consortium
As demonstrated above, genomic analysis of the rpoB gene (Table 2) and mass spectrometry (Figure 6) reveal that strains L21, L42, and L51, obtained from the lakt1p consortium, belong to different clusters. Furthermore, on Reddy’s differential agar, the colonies of these strains exhibited differential staining, indicative of their belonging to distinct subspecies of Lactococcus lactis. According to differential colouration, L21 corresponds to cluster 1 (L. lactis cremoris), L51 to cluster 2 (L. lactis lactis) and L42 to cluster 3 (L. lactis diacetilactis) (Table 3). These strains were therefore selected as representative genomes for each cluster.
To determine the gene groups responsible for the aforementioned metabolic characteristics in these strains, we performed a comparative genomic analysis and compiled unique gene lists for each strain through pairwise comparisons (Appendix A, Table A1, Table A2 and Table A3). Based on the data obtained, a pairwise comparison of the gene networks of strains L21, L42, and L52 was conducted using the string-db resource. Below, three pairs of gene interaction networks are presented: L21 vs. L42, L42 vs. L51, and L21 vs. L51, each characterised by specific Gene Ontology (GO) categories. For each comparison, the most reliable GO category (based on the number of genes involved) is reported. Figure 7 shows the reconstructed interaction networks based on genes differentiating strains L21 and L42. One network includes 37 genes present in L21 but absent from L42 (Table A1; Figure 7A), while the other comprises 44 genes present in L42 but absent from L21 (Table A1; Figure 7B).
Table 7 and Table 8 provide the data on the gene networks specific to strains L21 and L42, with a focus on their GO characteristics and how they differ from each other.
The protein-coding genes present in strain L42 but absent in L21 are involved in membrane transport processes (Figure 8B, Table 8), including the transport of peptides and metal ions. This indicates a potentially greater capacity of strain L42 to maintain intracellular homeostasis and to utilise protein derivatives. At the same time, the gene network of strain L21 compared with L42 (Table 7) contains GO categories associated with oxo-acid lyase activity, which suggests a potential ability of L21 to metabolise oxo-acids, including citric acid.
Figure 8 shows the reconstructed gene interaction networks based on genes distinguishing strains L42 and L51 from each other. One network comprises 44 genes present in strain L42 but absent in L51 (Table A2; Figure 8A), while the other comprises 27 genes present in L51 but absent in L42 (Table A2; Figure 8B).
According to the data presented in Figure 8 and Table 9 and Table 10, the protein-coding genes present in strain L42 but absent in strain L51 are involved in the biosynthesis of polysaccharides, including pathways using galactose as a precursor. In turn, the protein-coding genes found in strain L51 but absent in L42 are implicated in the metabolism of sugars—including galactose—and polysaccharides. These findings suggest that the differences between strains L42 and L51 are minor and are potentially associated with the involvement of the products of their unique genes in the same polysaccharide biosynthesis processes, albeit at different stages of their implementation.
Figure 9 shows the reconstructed gene interaction networks based on genes distinguishing strains L51 and L21 from each other, comprising 28 genes present in strain L51 but absent in L21 (Table A3; Figure 9A) and 37 genes present in strain L21 but absent in L51 (Table A3; Figure 9B).
According to the data provided in Figure 9 and Table 11, the protein-coding genes present in strain L51 but absent in strain L21 are involved in transport processes, signal transduction pathways and carbohydrate metabolism. By contrast, the principal distinction between strain L21 and strain L51 (see Table 12) is the presence in L21 of a citrate utilisation gene cluster, which enables this strain to metabolise citrate.
Thus, the comparative analysis indicates that the sets of protein-coding genes underlying the differences between strains L51 and L42 exhibit only minor variation in metabolic potential, primarily in the repertoire of enzymes involved in carbohydrate metabolism and polysaccharide biosynthesis. In contrast, the genome of strain L21 contains genes encoding the key enzyme of citrate metabolism, a feature characteristic of the subspecies L. lactis diacetilactis [9], although certain strains of L. lactis cremoris may also possess this capability [10].
Since none of the lact1p strains, including L21, can synthesise diacetyl or CO_2_ from citrate, we hypothesise that strain L21 may have lost this ability. The reason for this loss remains unclear. When considering potential causes of this loss, it is important to acknowledge that the activity of the citrate lyase complex depends on the activity of citrate permease, a transport protein required for citrate uptake from the medium. Notably, the gene encoding this citrate-specific transporter in L. lactis diacetilactis is plasmid-borne [9]. Since the current analysis was limited to chromosomal genomic sequences, it proved impossible to determine if strain L21 possesses the plasmid-encoded citP gene, which codes for citrate permease.
3. Materials and Methods
In this study, we examined a cow’s milk cheese starter culture from a farm in the village of Kosh-Agach (Altai Republic). The starter culture (named lakt1p) was aseptically collected into a sterile container. Metagenomic DNA was extracted from the starter culture to analyse the taxonomic affiliation of the cultures in the consortium. The lakt1p starter culture was also separated into individual cultures by sequential subculture on milk agar, MRS and M17 media (Condalab, Madrid, Spain), and Reddy’s differential agar at 30 °C. Seventeen individual colonies were selected for further analysis.
3.1. Microbiological Methods
Microbiological characterisation of the pure cultures was performed by inoculating them onto test media. The Reddy’s differential agar medium was prepared with the following components (%): Tryptone, 0.5; CaCO_3_, 0.3; carboxymethyl cellulose (CMC), 0.6; K_2_HPO_4_, 0.1; yeast extract, 0.5; l-arginine hydrochloride, 0.5; agar-agar, 1.5, 5 mL sterile skim milk; and 0.1% bromocresol purple indicator solution, 2 mL (pH 6.3). On this medium, L. lactis subsp. cremoris, which does not hydrolyse arginine, forms yellow colonies, while L. lactis subsp. lactis and L. lactis subsp. diacetilactis form white colonies.
A modified Reddy’s differential agar was also used, containing (g/L): CMC, 5; sodium citrate, 10; peptone, 7.5; yeast extract, 7.5; meat extract, 5; lactose, 1.5; L-Arginine hydrochloride, 1.5; bromocresol purple, 0.002; and agar-agar, 15 (pH 6.0).
Citrate utilisation was assessed using a medium comprising milk hydrolysate, 2.5% yeast autolysate, 2.0% agar-agar, 2 mL of 0.1% bromcresol purple indicator solution, and 1% sodium citrate. After 18–24 h of cultivation at 30 °C in this enrichment medium, the bacterial suspension was plated using the deep-pour method onto the solidified medium of the same composition. Colonies of L. l. diacetilactis were identified by the gas bubble formation resulting from citrate metabolism.
The acidification activity of pure strains was evaluated by measuring the time required for pasteurised milk to coagulate after inoculation with 5% of the pure culture starter. Titratable acidity was measured by adding 10 mL of the starter culture, 20 mL of distilled water, and three drops of phenolphthalein to a 100 mL flask, then titrating with a 0.1 N NaOH solution until a pink colour appeared. The volume of NaOH (in mL) multiplied by 10 corresponds to the acidity of the sample, expressed in Turner degrees (°T).
Carbon dioxide and four-carbon compounds are metabolic products of the aroma-forming bacterium L. l. diacetilactis. Carbon dioxide production was quantified by thoroughly homogenising the starter culture and pipetting 20 cm^3^ into a tube (15 mm diameter). The initial level of the culture was marked, and the tube was placed in a water bath containing cold water. The water was then heated to 90 °C, and the elevation of the curd was recorded without removing the tube. In the presence of carbon dioxide, the curd becomes spongy and rises 0.6–3 cm or more above the whey. In the absence of carbon dioxide, the curd typically does not rise and may even sink to the bottom.
Determination of diacetyl and acetoin was performed by mixing the starter culture and filtering it through a paper filter. To 200 µL of the filtrate, 200 µL of a 40% KOH solution was added, followed by thorough mixing. The time required for the development of a pink colour was recorded. If the colour appeared within 10 min, the presence of diacetyl in the starter culture was inferred.
3.2. Sample Morphology Analysis by Fluorescence Microscopy
Cells grown on a solid medium were used for microscopy. For each sample, at least three preparations were examined, with a minimum of 10 fields of view counted in each preparation. Fixed preparations were prepared from each sample and subsequently stained with DAPI in PBS buffer. Morphological analysis was performed using Karl Zeiss (Jena, Germany) microscopes: Axio Imager.M2 and Axio Imager.A2 (microscopic analysis was carried out at the Multiple-access Center for Microscopy of Biological Objects (Institute of Cytology and Genetics, Novosibirsk, Russia)) and Axio Scope.1 (Laboratory of Molecular Biotechnology, IC&G SB RAS, Akademgorodok, Russia).
3.3. DNA Extraction, Library Preparation, Sequencing
Genomic DNA was extracted from pure microbial cultures. The cells were resuspended in TE buffer (pH 8), and genomic DNA was isolated using the phenol–chloroform method. Plasmid DNA was not sequenced separately. The quality of the extracted DNA was assessed spectrophotometrically (Epoch, BioTek Instruments, Winooski, VT, USA) and using a Qubit fluorometer (Quant-iT™ dsDNA Assay Kit, High Sensitivity (HS), Thermo Fisher, Waltham, MA, USA). Library preparation and sequencing were performed at the Sequencing Centre of IC&G SB RAS using an Illumina MiSeq platform (version 4.1.0).
3.4. Bioinformatic Analysis of Sequencing Data
Quality check of metagenomic and individual strain sequencing raw reads was performed using FastQC (version 0.11.8), followed by trimming with Cutadapt (version 1.18). Metagenomic reads were aligned to the following reference genomes available in the NCBI database: Lactococcus cremoris subsp. cremoris KW2 (GCF_000468955.1), Lactococcus lactis subsp. lactis strain 14B4 (GCF_003176835.1), and Lactococcus lactis subsp. lactis bv. diacetilactis strain S50 (GCF_003627395.2), using Bowtie2 (version 2.3.5). Genome and metagenome assemblies were generated using SPAdes (version 4.0.0). The assembled metagenomic contigs were classified using MetaWRAP (version 1.2.2, classify_bins option), and the proportion of contigs assigned to each taxonomic unit was calculated. Genome and metagenome annotation was performed using PROKKA (version 1.14.6). Annotated genomes of individual samples were screened for the 16S, rpoB, and gyrB genes to determine the taxonomic identity of the strains. Clustering of the identified sequences was performed using CD-HIT (version 4.7). Average nucleotide identity (ANI) values for the assemblies were calculated using fastANI (version 1.34). A synthetic metagenome consisting of L. l. cremoris, L. l. diacetilactis, and L. l. lactis was constructed by combining the sequences of the corresponding reference genomes (GCF_000468955.1, GCF_003627395.2, and GCF_003176835.1) into a single file.
Seventeen genome assemblies from individual cultures of the lakt1p consortium were obtained and analysed. Sequence similarity clustering was performed using rpoB, and a genome from each cluster was selected for further bioinformatics analysis (L51, L42, L21).
3.5. Mass Spectrometric Analysis of Strain Proteomes
Proteomic analysis was performed using matrix-assisted laser desorption/ionisation time-of-flight mass spectrometry (MALDI-TOF MS).
3.6. Preparation of Bacterial Cell Lysates for MALDI-TOF MS Measurement
For each colony analysed, approximately 50 µL of culture was collected. The cells were resuspended in 300 µL of deionised water, followed by the addition of 900 µL of 96% ethanol for inactivation. The cells were pelleted by centrifugation for 2 min at 15,600× g. The supernatant was removed, and the pellet was dried for 5 min in an Eppendorf vacuum concentrator. Bacterial cell walls were disrupted by adding 50 µL of 70% formic acid. Proteins were extracted with 50 µL of acetonitrile. The mixture was centrifuged for 2 min at 15,600× g, and the supernatant was transferred to a clean tube for mass spectrometric analysis.
3.7. Conditions for MALDI-TOF MS Measurement
For mass spectrometric analysis, 1 µL of the protein extract was applied to a stainless-steel plate and allowed to dry at room temperature. Subsequently, 1 µL of matrix solution (6 mg/mL α-cyano-4-hydroxycinnamic acid in acetonitrile/water/trifluoroacetic acid, 50:47.5:2.5, v/v) was overlaid onto the sample.
Measurements were performed using an Ultraflex III MALDI-TOF/TOF mass spectrometer (Bruker Daltonics, Billerica, MA, USA). Spectra were acquired in linear positive mode with a laser frequency of 200 Hz over a mass range of 2000–20,000 Da. The accelerating voltage was set to 20 kV, IS2 voltage to 18.66 kV, and lens voltage to 6 kV, with extraction delay disabled. The obtained spectra represent the dependence of the mass of the sample proteins (m/z) and their intensity.
For each colony, the spectra were obtained by summing 2000 laser shots (10 × 200 shots from different target spot positions). External calibration was performed using the accurate mass values of known Escherichia coli proteins: RL36—4365.3 Da, RS22—5096.8 Da, RL34—5381.4 Da, RL32—6315.0 Da, RL29—7274.5 Da, and RS19—10,300.1 Da.
The acquired spectra were exported to mMass. A baseline subtraction procedure was applied to all spectra using the MALDI-TOF Proteins 5–20 kDa preset with default settings. Peak picking was performed using the following parameters: signal-to-noise threshold—2.5, absolute intensity threshold—0, relative intensity threshold—1%, and picking height—100%.
3.8. Statistical Analysis of Mass Spectra
Multivariate statistical analysis was performed using Past 5 software, version 5.0 [21]. Hierarchical cluster analysis was used to visualise the differences between the strains analysed. Multivariate approaches, particularly principal component analysis (PCA), were applied for a more comprehensive and in-depth evaluation. All m/z values and their intensities were used as variables for the principal component analysis (PCA).
3.9. Bioinformatic Analysis: Gene Network Construction
A comparative analysis of gene networks constructed from genes unique to each strain was performed to identify specific metabolic features of strains L21, L42, and L51 isolated from the lact1p sample. The STRING database application (https://string-db.org/, accessed on 23 December 2025) [22] was used to identify associated gene networks.
For each clone (L51, L42, L21), genome protein annotations were uploaded to STRING using the corresponding identifiers: L51—STRG0A09PVY, L42—STRG0A14NDF, and L21—STRG0A47PZJ.
The string-db.org application was configured with the following parameters: (1) meaning of network edges—evidence; (2) types of evidence used—text mining, experiments, databases, co-expression, neighbourhood, gene fusion, co-occurrence; (3) minimum required interaction score—minimum confidence 0.41; and (4) max number of interactions to show—no more than 10 interactions.
When describing the strain comparisons, the most reliable Gene Ontology (GO) categories (based on the number of genes represented) were selected.
4. Conclusions
This study characterised the microbiological and molecular genetic properties of lactic acid bacteria belonging to the species Lactococcus lactis isolated from a farm cheese starter culture (sample lakt1p). Pure cultures were obtained from this sample by sequential subculturing, and 17 colonies exhibiting different characteristics on differential media were selected. Genetic analysis based on the rpoB gene grouped these colonies into three clusters, each corresponding to a distinct strain of L. lactis subsp. diacetilactis. Microbiological and microscopic analyses further confirmed that all the strains could be assigned to L. lactis subsp. diacetilactis. However, none of the studied strains produced the characteristic aroma compounds of L. l. diacetilactis, such as diacetyl and CO_2_. Phenotypically, based on colony colouration on Reddy’s differential media, the strains were also divided into three clusters.
The functional characteristics of the lakt1p consortium strains were further explored using the STRING database. Pairwise comparisons of gene sets unique to strains L21, L42, and L52, which were assigned to different clusters based on genomic, proteomic, and phenotypic features, were performed. Gene network analysis revealed that the unique protein-coding genes distinguishing strains L51 and L42 conferred only minor differences in metabolic potential, primarily in the repertoire of enzymes involved in sugar metabolism and polysaccharide biosynthesis. In contrast, the gene network of unique protein-coding genes in strain L21 included a cluster of genes from the citrate lyase cluster, a feature characteristic of L. lactis subsp. diacetilactis [9]. Nevertheless, the inability of the lakt1p strains, including L21, to synthesise diacetyl and CO_2_ due to citrate utilisation suggests that at least L21 may have once possessed this ability but subsequently lost it. This loss could be associated with the absence of a functional citrate permease, which may be encoded on a plasmid.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kochetkova T.V. Grabarnik I.P. Klyukina A.A. Zayulina K.S. Elizarov I.M. Shestakova O.O. Gavirova L.A. Malysheva A.D. Shcherbakova P.A. Barkhutova D.D. Microbial Communities of Artisanal Fermented Milk Products from Russia Microorganisms 202210214010.3390/microorganisms 1011214036363732 PMC 9697859 · doi ↗ · pubmed ↗
- 2Elcheninov A.G. Zayulina K.S. Klyukina A.A. Kremneva M.K. Kublanov I.V. Kochetkova T.V. Metagenomic Insights into the Taxonomic and Functional Features of Traditional Fermented Milk Products from Russia Microorganisms 2023121610.3390/microorganisms 1201001638276185 PMC 10819033 · doi ↗ · pubmed ↗
- 3Shangpliang H.N.J. Tamang J.P. Metagenome-assembled genomes for biomarkers of bio-functionalities in Laal dahi, an Indian ethnic fermented milk product Int. J. Food Microbiol.202340211030010.1016/j.ijfoodmicro.2023.11030037364321 · doi ↗ · pubmed ↗
- 4Kuerman M. Wang R. Zhou Y. Tian X. Cui Q. Yi H. Gong P. Zhang Z. Lin K. Liu T. Metagenomic insights into bacterial communities and functional genes associated with texture characteristics of Kazakh artisanal fermented milk Ayran in Xinjiang, China Food Res. Int.202316411241410.1016/j.foodres.2022.11241436737993 · doi ↗ · pubmed ↗
- 5Bilal Z. Akhmetsadykova S. Baubekova A. Tormo H. Faye B. Konuspayeva G. The Main Features and Microbiota Diversity of Fermented Camel Milk Foods 202413198510.3390/foods 1313198538998490 PMC 11240983 · doi ↗ · pubmed ↗
- 6Litvina L.A. Gorskikh V.G. Anfilofyeva I.Y. Milk Microbiology: A Teaching Aid. Novosibirsk State Agrarian University, Institute of Ecological and Food Biotechnology 3rd ed.Suppl. and Updated Publishing House of NSAU Novosibirsk, Russia 2024105
- 7Krasnikova L.V. Gunkova P.I. Markelova V.V. Microbiology of Milk and Dairy Products: Laboratory Practical Training: Textbook National Research University ITMO St. Petersburg, Russia Institute of Chemical Technology and Biotechnology St. Petersburg, Russia 201385
- 8Khelissa S. Chihib N.E. Gharsallaoui A. Conditions of nisin production by Lactococcus lactis subsp. lactis and its main uses as a food preservative Arch. Microbiol.202120346548010.1007/s 00203-020-02054-z 33001222 · doi ↗ · pubmed ↗