Insights into the Structural and Proteomic Changes in Eimeria tenella Unsporulated Oocysts Treated with Sodium Hypochlorite
Liu-Shu Jia, Qing-Jie Wang, Shun-Hai Zhu, Qi-Ping Zhao, Yu Yu, Hong-Yu Han, Hui Dong

TL;DR
This study reveals how sodium hypochlorite affects the structure and protein composition of Eimeria tenella oocysts, offering new insights into their wall biosynthesis and resilience.
Contribution
The study provides the first proteomic analysis of Eimeria tenella oocysts treated with sodium hypochlorite, identifying key proteins involved in wall structure and stress response.
Findings
Sodium hypochlorite treatment disrupts the oocyst wall bilayer by removing the outer layer and thickening the inner layer.
1345 differentially expressed proteins were identified, with upregulated proteins linked to wall biosynthesis and stress responses.
Twelve proteins, including nine hypothetical ones, were uniquely detected in untreated oocysts, suggesting roles in outer wall formation.
Abstract
Sodium hypochlorite (NaClO) is widely used for oocyst purification, yet the effect of NaClO on oocyst proteomic changes has not been reported. This study investigated the structural and proteomic alterations in Eimeria tenella unsporulated oocysts induced by NaClO treatment. Transmission electron microscopy revealed that NaClO disrupts the oocyst wall bilayer by removing the outer layer and inducing thickening of the inner layer. Label-free quantitative proteomics identified 1344 differentially expressed proteins (DEPs) between the NaClO-treated (Et-T) and the untreated (Et-C) unsporulated oocysts. Functional analysis showed that DEPs are primarily associated with oocyst wall biosynthesis, stress response pathways, outer wall formation, and structural integrity. These findings provide critical insights into the molecular architecture of the oocyst wall and establish a foundation for…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
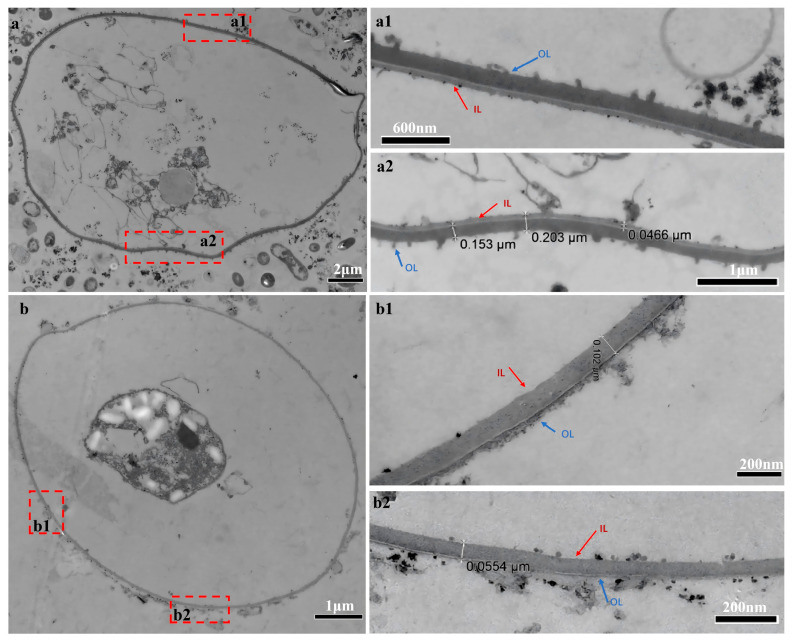 Figure 1
Figure 1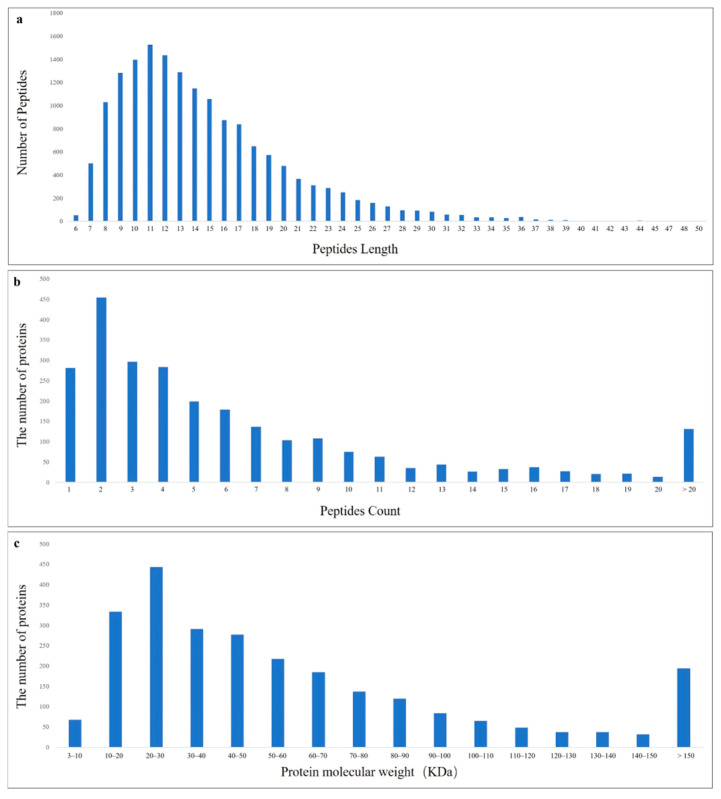 Figure 2
Figure 2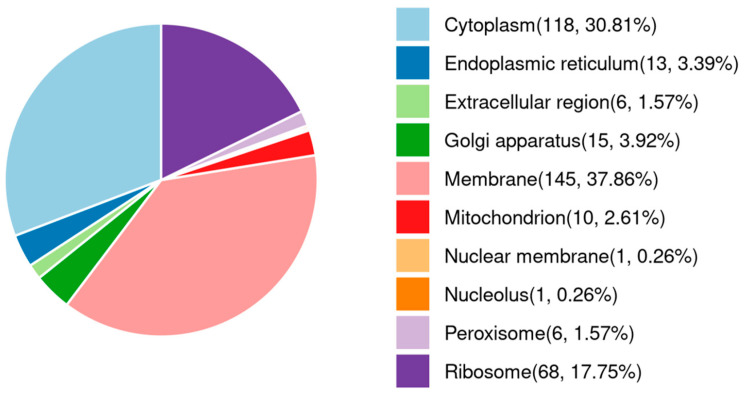 Figure 3
Figure 3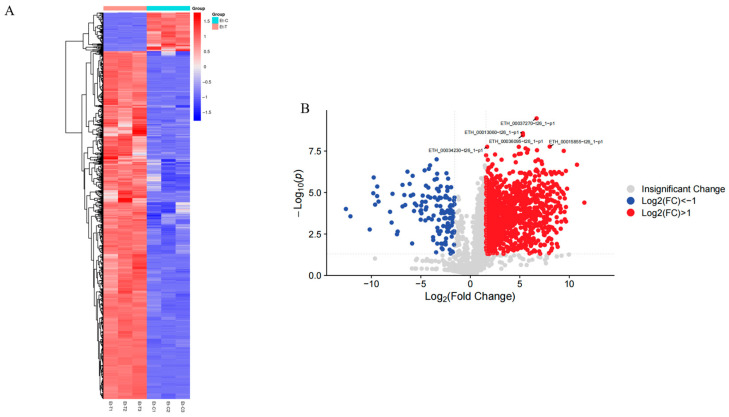 Figure 4
Figure 4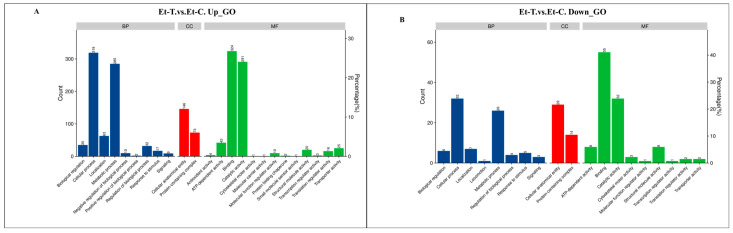 Figure 5
Figure 5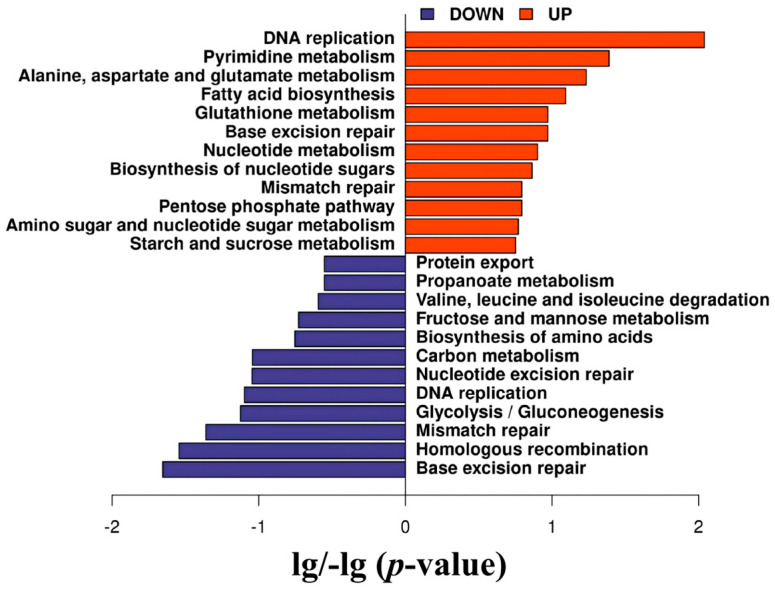 Figure 6
Figure 6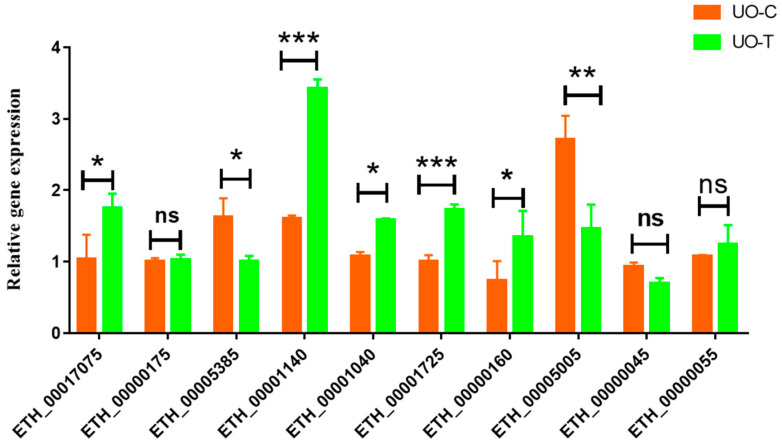 Figure 7
Figure 7- —National Natural Science Foundation of China
- —Jiangxi Provincial Natural Science Youth Fund Project
- —Jiangxi Province Early-career Young Talent Cultivation Program for Science and Technology
- —Key Research and Development of Science and Technology Plan in the Tibet Autonomous Region
- —National Parasite Resource Center
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCoccidia and coccidiosis research · Neuropeptides and Animal Physiology · Echinoderm biology and ecology
1. Introduction
Avian coccidiosis is a prevalent parasitic disease that results from infections by obligate intracellular parasites belonging to the Eimeria genus [1]. Eimeria tenella is recognized as the most pathogenic species associated with coccidiosis. This parasite has a specific site of parasitism, leading to the destruction of cecal/intestinal mucosa, as well as causing inflammation in the intestines and disintegration of epithelial cells [2]. This disease significantly impairs chicken growth and development, causing major economic losses exceeding £10.36 billion annually to the global poultry industry [3].
E. tenella undergoes a complex life cycle within the cecal epithelium of chickens following the ingestion of sporulated oocysts. This process involves three cycles of schizogony (asexual) followed by a subsequent phase of gametogony (sexual), culminating in the shedding of unsporulated oocysts with feces, which then proceed to undergo sporulation [4,5]. The unsporulated oocysts are the endpoint of sexual reproduction, possessing a robust bilayered wall that is fundamental to their environmental resilience. This structural barrier not only maintains oocyst integrity after excretion from the host but also effectively shields the internal undifferentiated protoplasm from a wide range of external physicochemical stressors. By protecting the parasite during its extracellular phase in the environment, this protective envelope ensures the successful completion of sporulation under permissive conditions and underpins the transmission potential and subsequent infection of new hosts [6,7]. To study the biological and molecular characteristics of oocysts, a large number of coccidial oocysts need to be isolated from Eimeria-infected chicken feces and sterilized and purified with sodium hypochlorite (NaClO). NaClO is widely used as a disinfectant on surfaces in settings such as healthcare facilities and food production plants, and is recognized as a cost-effective method for reducing the burden of waterborne diseases [8,9], and it also has the ability to disrupt biofilms and degrade structural proteins [10]. Many studies have successfully used NaClO solutions to clean fecal debris from oocysts of several species of coccidia [11,12,13]. However, when NaClO is used to treat oocysts, concentration and time must be strictly controlled. Studies have shown that some oocysts appear to break the oocysts wall and sporocysts overflow when treated with NaClO for more than 40 min and 5–10% NaClO solution can strip the outer wall of oocysts, cause damage to the structure of oocyst wall, and then affect the vitality of oocysts [14,15,16].
The effect of NaClO on the ultrastructure and protein profile of coccidian oocysts has not yet been reported. In this study, we systematically investigated structural changes in the wall of unsporulated oocysts of Eimeria tenella and alterations in protein expression following NaClO treatment by integrating transmission electron microscopy (TEM) with Label-free quantitative proteomics. The primary objective was to identify key functional proteins associated with outer oocyst wall formation and inner wall stabilization. Furthermore, quantitative real-time PCR (qPCR) was applied to validate transcriptional changes corresponding to the significantly differentially expressed proteins identified in the proteomic analysis. These findings offer novel insights and establish a theoretical foundation for understanding the protein composition, structural regulation, and biogenesis of the E. tenella oocyst wall.
2. Materials and Methods
2.1. Animals and Parasites
The protocol for the animal experiment received approval from the Animal Care and Use Committee at the Shanghai Veterinary Research Institute, Chinese Academy of Agricultural Sciences (Permit Number: SHVRI-SZ-20230323-4 on 23 March 2023). All procedures followed the approved guidelines to ensure adherence to ethical standards throughout the study.
One-day-old Three-yellow chickens were housed in temperature-controlled isolators (coccidia-free) with a constant ambient temperature of 30 ± 1 °C and a relative humidity of 50 ± 10%, and a 16 h:8 h light–dark cycle was maintained. Feed and water were provided ad libitum using a commercially available anticoccidial-free starter diet.
The E. tenella Shanghai strain was isolated from a commercial chicken farm in Shanghai and has been maintained in our laboratory since its original isolation [17]. The parasite was propagated through passage in coccidia-free chickens aged two weeks. Fecal samples were collected from chickens infected with E. tenella at 6–8 days post-infection. The fecal material was sequentially filtered through stainless steel sieves with mesh sizes of 80, 100, and 120 to collect the filtrate and centrifuged at 3200 rpm for 10 min. The pellet containing unsporulated oocysts (UO) was retained following supernatant removal.
2.2. Experimental Design
The omics experiment was divided into the NaClO-treated group (Et-T) and the control group (Et-C). The groups were treated as follows: (1) Et-T: 50% NaClO solution (with 4.5–5.0% available chlorine) was added to unsporulated oocysts, which were then repeatedly agitated on ice for 30 min and centrifuged at 2500 rpm for 10 min to extract the upper layer (i.e., oocysts) as previously described [18]. Subsequently, residual NaClO was removed by extensive washing with water. (2) Et-C: saturated salt flotation was used, according to Jenkins et al. (2017) [19], to obtain partially purified oocysts from fecal samples, serving as the control group. Previous studies have indicated that this treatment does not damage the wall of Eimeria oocysts [20].
Each group of samples was prepared in triplicate. All oocysts were washed thoroughly with sterile PBS and stored in liquid nitrogen.
2.3. Transmission Electron Microscopy
Unsporulated oocysts from the Et-C and Et-T groups were subjected to fixation with 1% glutaraldehyde at ambient temperature for an hour, followed by additional fixation using 1% osmium tetroxide for half an hour. Subsequent to the final wash in buffer and ethanol dehydration, the fixed samples were cleared with propylene oxide and subsequently embedded in a 1:1 mixture of Epon and Araldite (Pelco International, Fresno, CA, USA). Ultrathin sections measuring 40–60 nm were created and stained using uranyl acetate (Sigma-Aldrich, St. Louis, MO, USA) and lead citrate (Aladdin, Shanghai, China). Ultimately, these sections were examined under a transmission electron microscope (CM10, Philips, Amsterdam, The Netherlands) [21].
2.4. Protein Extraction and Protein Digestion
Proteins were isolated from samples utilizing SDT lysis buffer composed of 4% SDS (Bio-Rad, Hercules, CA, USA), 100 mM DTT (Sigma-Aldrich, St. Louis, MO, USA), and 100 mM Tris-HCl at pH 8.0. The samples were homogenized by vortexing with an equal volume of glass beads for 3 to 5 min. Samples were boiled for 5 min, further ultrasonicated, and then boiled for another 5 min. Undissolved cellular debris was removed by centrifugation at 16,000× g for 15 min. For digestion, 200 μg of protein from each sample was processed using the FASP method as outlined by Wiśniewski et al. (2009) [22]. Briefly, the UA buffer was supplemented with detergent, DTT, and IAA (Sigma-Aldrich, St. Louis, MO, USA) to prevent the reduction of cysteine residues. Finally, the protein suspension was digested overnight at 37 °C with trypsin (Promega, Madison, WI, USA) at a 50:1 ratio. Peptides were then collected by centrifugation at 16,000× g for 15 min and desalted using a C18 StageTip (Thermo Fisher Scientific, Waltham, MA, USA) before LC-MS (Thermo Fisher Scientific) analysis. The peptide concentrations were determined at an OD280 using a Nanodrop One device (Thermo Fisher Scientific, Waltham, MA, USA).
2.5. LC–MS/MS Analysis
The MS data were analyzed using MaxQuant software version 2.0.1.0 (MPI of Biochemistry, Martinsried, Germany). MS data were searched against the ToxoDB-59_EtenellaHoughton_AnnotatedProteins.fasta. An initial search was set with a precursor mass window of 6 ppm. The method involved an enzymatic cleavage approach utilizing trypsin KR/P, with a mass tolerance of 20 ppm for fragment ions; up to two missed cleavage sites were permitted. The results from the database search were filtered and subsequently exported, achieving a false discovery rate (FDR) of less than 1% at both the peptide-spectrum-matched level and the protein level. Label-free quantification was performed in MaxQuant, employing an intensity determination and normalization algorithm as outlined in prior studies [23,24,25]. The “LFQ intensity” for each protein across various samples was calculated to provide the most accurate estimate, fulfilling all pairwise peptide comparisons. This LFQ intensity closely matched the aggregated peptide intensities. Protein ratios were weighted and normalized using the median ratio in MaxQuant software. Only those proteins exhibiting a change of ≥1.5-fold along with a p-value of less than 0.05 were deemed to represent significant differential expressions.
2.6. Bioinformatics Analysis
Analyses of bioinformatics data were carried out using Perseus software (version 1.6.10.50) [26], Microsoft Excel, and R statistical computing software (version 3.5.3). Hierarchical clustering analysis was conducted utilizing the heatmap package, wherein the Euclidean distance served as the designated distance metric and complete linkage was employed as the chosen agglomeration method. Data was retrieved from sources including UniProtKB/Swiss-Prot [27], the Kyoto Encyclopedia of Genes and Genomes (KEGG) [28], and Gene Ontology (GO) [29] for the purpose of annotating the sequences. Enrichment analyses for GO and KEGG were performed using Fisher’s exact test, with false discovery rate (FDR) correction conducted to account for multiple testing. The GO terms were categorized into three groups: biological process (BP), molecular function (MF), and cellular component (CC) [30]. The enriched GO and KEGG pathways achieved nominal statistical significance at the p < 0.05 threshold.
2.7. Quantitative Real-Time PCR Assay
Ten genes were selected from the differently expressed unsporulated oocyst proteins to investigate transcription levels using qPCR [31]. Total RNA was extracted from unsporulated oocysts of Et-C and Et-T using TRIzol reagent according to the manufacturer’s instructions (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA) and reverse transcribed into cDNA using the HiScript III RT SuperMix for qPCR (+gDNA wiper) (Vazyme, Nanjing, China). qPCR analysis was performed on complementary DNA using the QuantiNova SYBR Green PCR Kit (Qiagen, Hilden, Germany) in combination with custom-designed primers. The primers, which were specific to the target genes, were synthesized by Sangon Biotechnology Co. (Shanghai, China) and are listed in Table 1. The 18S ribosomal RNA (rRNA) of E. tenella was simultaneously amplified as an internal reference gene [32]. The relative expression levels were determined using the 2^−ΔΔCt^ method [33], with each sample analyzed in three replicates.
2.8. Statistical Analysis
The relative gene expression levels in the Et-C and Et-T groups are presented as the mean ± standard deviation (SD). The statistical significance between samples was evaluated by performing Student’s t-test, with p-values less than 0.05 regarded as statistically significant, p < 0.01 indicated highly significant difference, p < 0.001 indicated extremely significant difference.
3. Results
3.1. Ultrastructural Changes in Oocyst Walls After Treatment with NaClO
Oocyst walls from control group remained an intact bilayer structure. The outer layer appeared electrodense with a roughened appearance. The outer surface had a thickness of 0.153 μm–0.161 μm. The electron-lucent inner layer appeared much thinner and had a thickness of 0.0466 μm–0.0784 μm (Figure 1(a1,a2)). However, in NaClO-treated samples, the bilayer structure of the oocyst wall was destroyed. The outer wall was stripped away; generally, only the inner layer was seen. The texture of the inner wall was loose, and the intermolecular gaps were increased. Its thickness was 0.0871 μm–0.102 μm. The inner wall was visually noticeably thickened (Figure 1(b1,b2)).
3.2. Proteins Detected in Unsporulated Oocysts of E. tenella
A total of 16,401 peptides (Table S1) and 2422 proteins (Table S2) were identified in unsporulated oocysts of E. tenella using the label-free proteomic approach. The lengths of the peptides varied between 7 and 20 amino acids, with 95% of the identified peptides measuring under 32 amino acids (Figure 2a). Analysis of the unique peptide counts for the identified proteins revealed that the majority were characterized by 1 to 20 peptides, and most of these proteins had fewer than 10 peptide segments (Figure 2b). Regarding the distribution of protein masses, extensive coverage was achieved across a broad spectrum of molecular weights for proteins under 160 kD (Figure 2c).
Of these proteins, 2389 co-existed in both the Et-T and Et-C groups, which grouped according to their subcellular locations as follows: 145 proteins were expressed in membranes (37.86%), 118 proteins were located at the cytoplasm (30.81%), and 68 proteins (17.75%) were expressed in ribosomes (Figure 3). The top 50 proteins with the most abundance in unsporulated oocysts of E. tenella are listed in Table 2. The 56 kDa gametocyte antigen, elongation factor 1-α, actin, equisetin synthetase, protein disulfide isomerase, heat shock protein 90, heat shock protein 70, glycogen phosphorylase family protein, fructose-bisphosphate aldolase, and enolase 2 were the top 10 most abundant proteins.
3.3. Differentially Expressed Proteins (DEPs) in NaClO-treated Unsporulated Oocysts of E. tenella
Although identical total protein concentrations were utilized in samples from Et-T and Et-C unsporulated oocysts, we found considerable differences in individual protein levels between Et-T and Et-C (Table S3). The results from screening the DEPs have been visualized in hierarchical cluster analysis and volcano plots. Hierarchical cluster analysis was performed for all the DEPs for the Et-T vs. Et-C group (Figure 4A). The intra-group variation was minimal, and the inter-group comparability was high. In Figure 4B, upregulated protein expression is displayed in red, whereas downregulated expression is shown in blue. Compared with the untreated group, 1344 proteins displayed statistical changes in their expression levels (p < 0.05) in the NaClO-treated unsporulated oocysts, with 1210 upregulated and 134 downregulated. Additionally, 1045 proteins were not significantly changed between them.
The detailed upregulated data of 611 previously described proteins and 599 hypothetical proteins were observed in NaClO-treated unsporulated oocysts of E. tenella, of which 35 protein kinases, 17 proteasomes, 17 ubiquitins, 15 zinc finger proteins, 13 ribosomal proteins, and 6 RNA binding proteins displayed over a 500-fold change compared with the untreated unsporulated oocyst group (Table S4). Additionally, 16 proteins were associated with oocyst wall biosynthesis (Table 3), 15 proteins were related to the translation process of proteins (Table 4), 17 proteins were associated with response to stimuli of the E. tenella unsporulated oocysts (Table 5), and 25 proteins included splicing factors, DnaJ domain-containing proteins, and SAGs (Table 6).
The downregulated DEPs included 67 proteins that had been previously reported and 67 proteins with unknown functions. The top 35 proteins with the highest abundance in the outer wall of unsporulated oocysts are listed in Table 7. Histone H2A, tubulin beta chain, ATP synthase alpha chain, histone H4, tubulin alpha chain, and histone H2B were the most abundant proteins. Additionally, 18 proteins of interest were identified, such as microneme proteins, elongation factor G, and several enzymes (Table 8). Interestingly, 12 proteins specific for group Et-C were identified, including nine hypothetical proteins, acid phosphatase, adenylyl cyclase, and microneme protein 2 (Table 9).
3.4. GO Annotations and KEGG Pathway Analysis of DEPs
To further understand the changes observed in unsporulated oocysts of E. tenella upon NaClO treatment, DEPs were categorized into three functional categories—biological process, molecular function, and cellular component—based on the Gene Ontology (GO) classification system and the UniProt database. In the 1210 upregulated DEPs, the most prevalent biological processes were cellular processes (319 proteins) and metabolic processes (285). The most prevalent cellular component was the cellular anatomical entity (146) and protein-containing complex (73). The predominant molecular functions were binding (324), catalytic activity (291), ATP-dependent activity (42), and transporter activity (25) (Figure 5A). Similarly, among the 134 downregulated DEPs, the dominant components of the biological processes and cellular components were the same as the upregulated DEPs. The most prevalent molecular functions included structural molecule activity (8) and binding, catalytic activity, and ATP-dependent activity (Figure 5B).
The biological functions of these 1344 differentially expressed unsporulated oocyst proteins were further analyzed with the KEGG database, which mapped them to 40 pathways (Table S5). According to the p-value, Figure 6 presents the pathways with the top 12 enrichment significance. The upregulated DEPs were involved in DNA replication, pyrimidine metabolism, alanine-aspartate and glutamate metabolism, and fatty acid biosynthesis. In addition, the downregulated DEPs were associated with base excision repair, homologous recombination, mismatch repair, and glycolysis/gluconeogenesis.
3.5. Validation of Label-Free Proteomic Results with qPCR
Ten proteins were selected through qPCR to confirm the reliability of the proteomic results. These included four upregulated, three downregulated, and three non-significant.
The mRNA expression levels detected with qPCR were consistent with those obtained by proteomics for seven proteins, including acid phosphatase (ETH_00005385), AGC kinase (ETH_00001040), aspartyl proteinase (Eimepsin) (ETH_00001725), acetyltransferase domain-containing protein (ETH_00000160), prolyl-tRNA synthetase (ETH_00000045), and two hypothetical proteins (ETH_00000175 and ETH_00000055). There was 70% agreement between the qRT-PCR and proteomic results. Results for three proteins did not agree with the proteomic data: adenylyl cyclase (ETH_00017075), hypothetical protein (ETH_00001140), and ATP-binding cassette sub-family F member 1 (ETH_00005005) (Table 10 and Figure 7). These results indicate that most proteins were regulated directly at the transcriptional level. Nevertheless, there were instances where the levels of gene transcripts did not align with those of the respective proteins. This discrepancy may imply that protein abundance is not solely reliant on transcript levels but may also be influenced by post-translational modifications [34].
4. Discussion
Oocysts are the most resistant stage of the Eimeria life cycle. Oocysts may maintain their infectivity following exposure to disinfectants such as bleach, free chlorine, chlorine dioxide, and chloramine, when applied at concentrations and for durations commonly used in domestic or industrial settings [35,36,37]. In sporulated oocysts, the bilayered oocyst and sporocyst walls act as robust, almost hermetic barriers that shield the sporozoites from the harmful impacts of diverse environmental stressors, including both physical and chemical agents [38]. In unsporulated oocysts, the protoplasmic mass is protected by the double-layered oocyst wall, allowing for successful sporulation in a suitable environment. Structural damage to the oocyst wall, compromises the barrier function of the oocyst or sporocyst, leading to cytoplasmic leakage, premature excystation, or exposure of internal stages, which reduces infectivity. Oocysts can also activate compensatory mechanisms, including resistance from tyrosine-rich structural proteins, upregulation of stress-response [39] and redox pathways [40], and stage-specific resource reallocation for repair or development [41]. Although NaClO disrupts wall integrity and impairs viability [14,15], differences in stage susceptibility and intrinsic adaptation reveal the complexity of oocyst persistence in diverse environments.
According to the TEM results of the present study, the non-treated oocysts in the Et-C group retained their typical double-layered wall (observed thickness ~200 nm); that is, the inner layer (observed thickness ~60 nm) and the outer layer (observed thickness ~150 nm). In contrast, the outer layer was absent when oocysts were treated with 50% NaClO on ice for 30 min, with only the inner layer (observed thickness ~90 nm) remaining. In some instances, slight remnants of the outer layer persisted. The oocyst wall thickness was consistent with those reported in the literature (90 nm for the inner layer) [42]. After treatment with NaClO, it was observed that the inner layer was thickened and had become looser and sparser. This phenomenon may be attributed to the absent outer layer, leading to enlarged intermolecular gaps within the inner layer [43]. Ultrastructural changes in the oocyst wall may alter the composition of proteins.
Previous studies have demonstrated that both the functional activity and expression levels of proteins exhibit variability under different experimental conditions, such as alterations in temperature, nutrient availability, oxidative stress, hypoxic environments, and exposure to various pharmacological agents or toxic compounds [44,45,46]. The rapid changes in the protein profile observed following sodium hypochlorite treatment are unlikely to result from de novo protein synthesis, but instead likely arise from oxidative modifications of existing proteins, altered protein solubility and extraction efficiency due to disruption of the oocyst wall structure, and the physical loss of proteins associated with the degraded outer oocyst layers. These findings suggest that the parasite may modulate protein expression as a response to external stress [47,48,49,50]. In this study, NaClO treatment upregulated 17 proteins, including DnaJ domain-containing protein (ETH_00006810), 3,5-cyclic-nucleotide phosphodiesterase (ETH_00011905), phosphatidylinositol 3-kinase (ETH_00030035), and serine/threonine protein phosphatase (ETH_00043830). The upregulated proteins likely contribute to oocyst wall integrity and stress adaptation. The DnaJ domain-containing protein may refold or stabilize oxidative stress-damaged wall-associated proteins [51]. 3′,5′-cyclic-nucleotide phosphodiesterase may regulate intracellular signaling, influencing cytoskeletal reorganization or membrane remodeling needed for structural maintenance [52,53]. Phosphatidylinositol 3-kinase supports membrane trafficking and lipid signaling [54], potentially aiding repair of the inner oocyst wall [55]. Serine/threonine protein kinase may phosphorylate key structural or regulatory proteins, enabling rapid post-translational responses after wall damage [56]. Additionally, 16 proteins involved in the biosynthesis of the oocyst wall, 15 proteins involved in protein transcription and translation, and some functional proteins were identified. These proteins showed similar identification results in the proteomics of E. tenella oocyst wall and Toxoplasma gondii oocyst wall [14,57]. The upregulation of these functional proteins may indicate a compensatory response to the downregulation of proteins related to the stability of the oocyst wall structure, or it may potentially signify one of the adaptive responses of parasites to external stimuli [43,58,59]. Furthermore, we found two types of proteins—cathepsin L-like thiol proteinase (ETH_00033530) and OTU-like cysteine protease (ETH_00040555)—to be involved in the synthesis of the oocyst wall protein and may participate in the sporulation process of unsporulated oocysts [60,61].
Research has shown that NaClO can effectively remove the outer layer of the oocyst [43,62], resulting in the downregulation or disappearance of specific proteins. In this study, the downregulated DEPs mainly included histones, ATP synthase, phosphoglycerate kinase, PAN domain-containing protein, GPI transamidase subunit PIG-U, and glycerol-3-phosphate dehydrogenase. Additionally, functional proteins such as 3-hydroxyisobutyryl-CoA hydrolase, microneme protein MIC4, fatty acid hydroxylase, and elongator complex protein 3 were also identified. These proteins were also identified in the proteomic analyses of sporozoites and merozoites [63]. These findings suggest that these proteins may play a role in maintaining the structural integrity of the oocyst wall, as well as contributing to the biological processes associated with parasite growth, survival, and virulence [64,65,66,67]. Interestingly, in the present study identified 12 proteins specifically found in the Et-C group, including nine hypothetical proteins, acid phosphatase (ETH_00005385), adenylate cyclase (ETH_00017075), and microneme protein 2 (ETH_00026625). This may be related to the removal of the outer wall. Three known proteins have been previously documented to localize on the cell membrane or participate in its biogenesis [49,68,69,70]. However, the functions of the nine hypothetical proteins remain unknown, and further in-depth studies are needed to reveal their potential mechanisms in maintaining the structural stability of the oocyst outer wall and in the process of biosynthesis.
5. Conclusions
This study represents the first report of the abundance and differences in protein composition of unsporulated oocysts of E. tenella before and after household bleach (NaClO) treatment. The DEPs identified may have pivotal implications for the survival of E. tenella oocysts, as well as for wall formation. Our findings validate the resilience of the inner oocyst wall to household bleach [37] and underscore its unforeseen role as a protective barrier. Additionally, 12 specific proteins were discerned in the Et-C group, holding significant promise for early-stage oocyst formation and biosynthesis research on oocyst inner/outer walls. These findings contribute to a deeper understanding of the molecular architecture and stress response mechanisms in Eimeria spp. oocysts. The identified DEPs, particularly those associated with oocyst wall integrity and oxidative stress response, provide valuable insights that may guide the development of novel anti-coccidial therapeutics or targeted disinfection approaches aimed at interrupting parasite transmission. It should be acknowledged, however, that the present study was restricted to unsporulated oocysts; the responses of sporulated oocysts—the infectious stage—may differ due to their unique structural and metabolic properties. Moreover, while Label-free proteomics enables a comprehensive characterization of the proteome, further functional validation is required to definitively elucidate the biological roles of candidate proteins in oocyst wall formation and resistance to NaClO treatment. Future studies incorporating stage-specific comparative analyses and functional assays will be essential to fully unravel the mechanisms underlying oocyst environmental persistence.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Attree E. Sanchez-Arsuaga G. Jones M. Xia D. Marugan-Hernandez V. Blake D. Tomley F. Controlling the causative agents of coccidiosis in domestic chickens; an eye on the past and considerations for the future CABI Agric. Biosci.202123710.1186/s 43170-021-00056-534604790 PMC 8475900 · doi ↗ · pubmed ↗
- 2Zhang Y. Zuo R. Song X. Gong J. Wang J. Lin M. Yang F. Cheng X. Gao X. Peng L. Optimization of maduramicin ammonium-loaded nanostructured lipid carriers using box-behnken design for enhanced anticoccidial effect against Eimeria tenella in broiler chickens Pharmaceutics 202214133010.3390/pharmaceutics 1407133035890226 PMC 9323563 · doi ↗ · pubmed ↗
- 3Adams D.S. Kulkarni R.R. Mohammed J.P. Crespo R. A flow cytometric method for enumeration and speciation of coccidia affecting broiler chickens Vet. Parasitol.202230110963410.1016/j.vetpar.2021.10963434864364 · doi ↗ · pubmed ↗
- 4Hu D. Tang X. Ben Mamoun C. Wang C. Wang S. Gu X. Duan C. Zhang S. Suo J. Deng M. Efficient single-gene and gene family editing in the apicomplexan parasite Eimeria tenella using CRISPR-Cas 9Front. Bioeng. Biotechnol.2020812810.3389/fbioe.2020.0012832158750 PMC 7052334 · doi ↗ · pubmed ↗
- 5Nash T.J. Morris K.M. Mabbott N.A. Vervelde L. Inside-out chicken enteroids with leukocyte component as a model to study host-pathogen interactions Commun. Biol.2021437710.1038/s 42003-021-01901-z 33742093 PMC 7979936 · doi ↗ · pubmed ↗
- 6Kong P. Lehmann M.J. Helms J.B. Brouwers J.F. Gupta N. Lipid analysis of Eimeria sporozoites reveals exclusive phospholipids, a phylogenetic mosaic of endogenous synthesis, and a host-independent lifestyle Cell Discov.201842410.1038/s 41421-018-0023-429844921 PMC 5964319 · doi ↗ · pubmed ↗
- 7Ndao O. Puech P.H. Bérard C. Limozin L. Rabhi S. Azas N. Dubey J.P. Dumètre A. Dynamics of Toxoplasma gondii oocyst phagocytosis by macrophages Front. Cell Infect. Microbiol.20201020710.3389/fcimb.2020.0020732509593 PMC 7248298 · doi ↗ · pubmed ↗
- 8Clasen T. Edmondson P. Sodium dichloroisocyanurate (Na DCC) tablets as an alternative to sodium hypochlorite for the routine treatment of drinking water at the household level Int. J. Hyg. Environ. Health 200620917318110.1016/j.ijheh.2005.11.00416387550 · doi ↗ · pubmed ↗