The Impact of Nanoplastics on the Quality of Fish Sperm: A Review
Hayam Djafar, Saira Naz, Maria Montserrat Rivera Del Alamo, Juan Carlos Balasch, Mariana Teles

TL;DR
This review explores how nanoplastics harm fish sperm quality, affecting motility, viability, and reproduction, with potential long-term impacts on fish populations.
Contribution
The paper provides a comprehensive synthesis of nanoplastic effects on fish sperm, emphasizing gaps in understanding and transgenerational risks.
Findings
Nanoplastics consistently impair sperm motility, viability, and fertilization capacity in fish.
Exposure to nanoplastics causes oxidative stress, DNA damage, and mitochondrial dysfunction in fish sperm.
Nanoplastics can lead to reduced offspring survival and disrupted gene expression in fish.
Abstract
Nanoplastics (NPs) are increasingly present in aquatic environments and are small enough to interact directly with biological cells. Fish sperm are particularly vulnerable because they are released directly into the water during fertilization and have limited protective and repair mechanisms. This review summarizes current evidence on how nanoplastics affect fish sperm quality, including motility, viability, fertilization capacity, and underlying cellular mechanisms such as oxidative stress and mitochondrial dysfunction. It also discusses indirect evidence from gonadal, hormonal, and molecular studies, while highlighting the limitations of extrapolating these effects to sperm function. Emerging findings on multigenerational effects and interactions with other environmental stressors are reviewed, emphasizing areas where data remain scarce. Finally, the review outlines key knowledge gaps…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
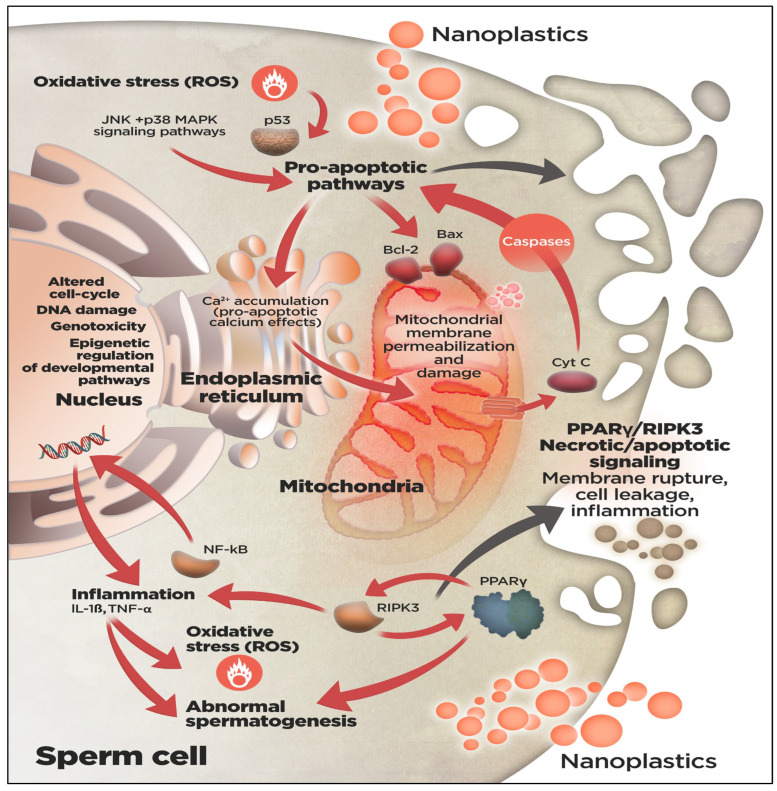 Figure 1
Figure 1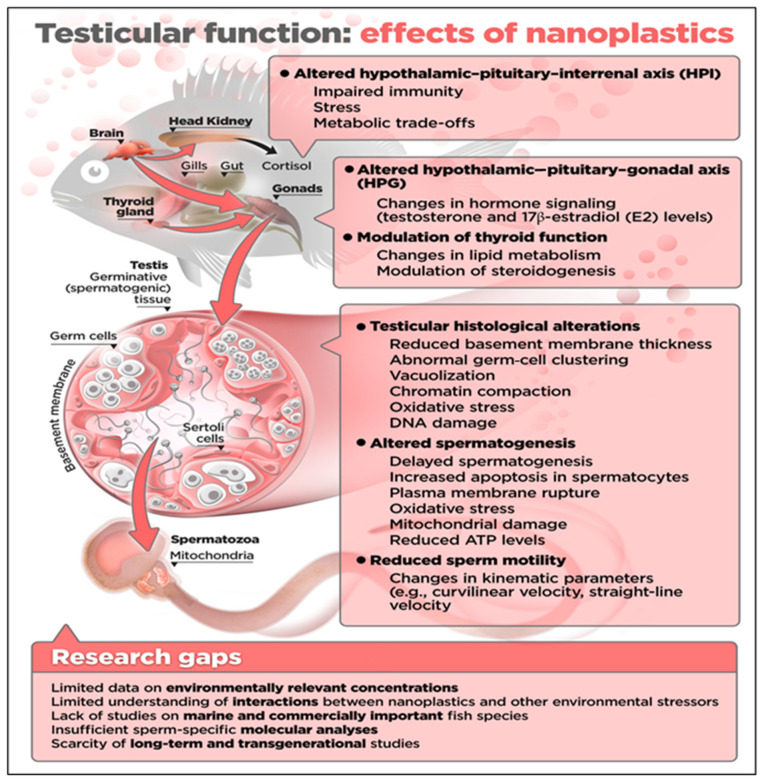 Figure 2
Figure 2- —Plan Nacional de Investigación
- —Ramón y Cajal contract
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicroplastics and Plastic Pollution · biodegradable polymer synthesis and properties · Marine Biology and Environmental Chemistry
1. Introduction
In an ever-changing world of increased plastic contamination, pathogen load, rising temperatures, ocean acidification, and variable marine and atmospheric circulation patterns, the resilience of freshwater and marine species depends critically on the proper activation of the endocrine hypothalamic–pituitary–gonadal axis (HPG). Among the plethora of emergent contaminants described in the last decade, plastic contamination in particular is becoming increasingly severe under changing climatic conditions [1,2]. Extreme weather accelerates the degradation of plastic fragments [3] into microplastics (MPs: ranging from 1 μm to 5 mm) and, ultimately, nanoplastics (NPs: less than 1 μm). Unlike MPs, NPs due to their reduced size can cross cellular barriers and nest in gonadal tissues, interfering with the endocrine, metabolic, and stress-related hormonal axis, and, through the production of reactive oxygen species (ROS) that induces mitochondrial and DNA damage, impairing gametogenesis, fertility rates, inducing embryo mortality or malformations, and altering overall growth [4,5,6]. Environmental stressors such as elevated temperatures or high density that may increase the NPs load, may also exacerbate the activation of the endocrine stress axis and impair gonadogenesis and spermatogenesis in fish, biasing the reproductive phenotype towards masculinization [7]. It should be noted that the upper size limit used to define nanoplastics varies across studies, with some authors considering particles <100 nm and others extending the definition up to 1000 nm, which complicates direct comparisons between studies [8,9].
NPs also interact with co-contaminants, such as metals, polycyclic aromatic hydrocarbons, pharmaceuticals, pesticides, antibiotics, plastic additives, and endocrine disruptors that can further degrade the reproductive physiology, and even the behavioral courting in fish [10], inducing oxidative stress, impairing foraging activity, immunity, and lipid metabolism, and modulating steroidogenesis and thyroid function [11,12]. NPs can also be maternally transferred to embryos as described on zebrafish (Danio rerio) and the ovoviviparous species Poecilia reticulata [13], but very few studies have addressed the intergenerational effects of NPs exposure in non-model fish species. Moreover, a recent systematic review highlighted that the effects of plastic pollutants in fish have been studied mainly in adults and females, and, to a lesser degree, in males [14].
In male fish the main physiological variables analyzed in stressful, plastic-ridden environments are spermatogenesis, sperm motility, malformation, and fertility rates, and the histopathology of the testis [15]. Spermatogenesis is a complex process dependent of the species-specific cyclic changes of the HPG axis, and the sequential activation of sex-determining genes [16], or the effect of temperature and pH in species whose sex determination depend on environmental variables acting through epigenetic modification of genes controlling gonadal differentiation and maturation [17]. However, despite increasing concern regarding NPs toxicity, a substantial gap persists in our understanding of its direct impact on fish sperm quality and functionality—a gap this review seeks to address through a critical evaluation of the existing literature.
2. Materials and Methods
A comprehensive literature search was conducted to identify peer-reviewed journal articles examining the toxic effects of NPs on fish sperm. Particular emphasis was placed on outcomes related to sperm motility, viability, proteomics, and transcriptomics. Search was performed in three major electronic databases: Google Scholar, Web of Science, and PubMed, and covered studies published up to August 2025 (Table 1).
The following search terms were applied, both individually and in combination:
“NPs effect on fish sperm viability”
“NPs effect on fish sperm motility”
“NPs effect on fish sperm fertilization capacity”
“NPs effect on fish reproduction”
“NPs effect on fish sperm metabolism”
For each database, the number of results generated by these keywords is summarized in Table 1.
Following the initial search, a filtering process was applied to refine the literature search, such as duplicate records identified across Google Scholar, Web of Science, and PubMed, which were removed manually. We included only original research articles with experimental data on NPs and fish reproduction (Table 2). Titles and abstracts were first screened to exclude non-fish species, review articles, opinion papers, and studies not addressing reproductive or sperm-related endpoints. Full-text screening was then applied to retain only original experimental studies that specifically assessed sperm quality parameters (e.g., motility, viability, fertilization capacity) or closely related reproductive outcomes directly linked to sperm function. Examples include studies focusing on gonadal development, hormone regulation, bioaccumulation, maternal transfer, immune responses, and co-exposure with other toxicants. After excluding such studies, only those articles specifically targeting sperm quality indicators, reproductive performance, or closely related endpoints were retained (Table 2). No formal scoring system was applied; however, studies were evaluated for experimental clarity, relevance to NPs, and the presence of clearly defined exposure conditions and endpoints. The final dataset was narrowed to nine articles because only these met all inclusion criteria, reflecting the current scarcity of studies directly addressing NPs’ effects on fish sperm. The overall literature searches and selection process followed an approach conceptually aligned with PRISMA guidelines, although implemented in a narrative rather than systematic review framework.
Studies from both marine and freshwater systems were considered. Despite the fact that these environments differ substantially in their physicochemical properties, such as salinity, ionic composition, and organic matter content, which in turn can influence NPs aggregation, stability, and bioavailability [32,33], integrating evidence from both systems allowed for a broader and more comprehensive overview of the available data. Further several studies include both microplastics and NPs but, this review focuses specifically on NPs, and results from mixed-size studies are discussed only when NPs-specific effects can be distinguished or when size-dependent differences are explicitly reported. The findings from these studies are synthesized below according to key sperm-related endpoints and biological processes, providing a structured overview of current knowledge and highlighting where gaps remain. While this review focuses on sperm quality and functionality, it is important to distinguish between studies that directly assess sperm-specific endpoints (e.g., motility, viability, fertilization capacity, mitochondrial activity, or DNA integrity) and those that evaluate broader reproductive or gonadal-level responses, such as testicular histopathology, endocrine disruption, or whole-gonad transcriptomics. The latter provide valuable mechanistic and physiological context but represent indirect proxies of sperm function. Consequently, extrapolations from gonadal alterations to sperm quality should be interpreted with caution, particularly when direct sperm measurements are not available.
3. Effects of NPs on the Quality of Sperm in Fish
Fish sperm, released into the surrounding water during external fertilization, are immediately exposed to contaminants, including MPs and NPs [34]. Due to small size, limited cytoplasmic content, and high membrane fluidity, sperm are particularly fragile and highly sensitive to environmental stressors [35,36]. The reviewed studies encompass both single-stressor exposures to NPs and multi-stressor scenarios in which NPs co-occur with other chemical contaminants. These two experimental approaches provide complementary insights, allowing NPs to be evaluated either as primary toxicants or as vectors and modulators of co-contaminant toxicity. Distinguishing between these roles is essential for interpreting reproductive outcomes and for understanding the ecological relevance of NPs’ exposure in complex environmental settings. In single-stressor studies, NPs alone consistently impaired sperm-related and reproductive endpoints, including spermatogenesis, sperm motility, fertilization success, and offspring development, supporting their role as primary reproductive toxicants [18,20,21]. In contrast, multi-stressor studies demonstrated that NPs can exacerbate or modify the toxicity of co-occurring contaminants, such as microcystin-LR, triclosan, or sulfamethazine, often amplifying oxidative stress, endocrine disruption, and germ cell damage [23,28,31]. These findings indicate that NPs may function not only as toxicants per se, but also as vectors or stress amplifiers in chemically complex environments.
NPs, with their reactive surfaces and ability to cross biological barriers, can adhere to or penetrate sperm cells, inducing oxidative stress [37], particularly those regulated by the nuclear factor erythroid 2–related factor 2 (Nrf2), which mediates antioxidant responses to elevated reactive oxygen species (ROS). Studies show that NPs induce oxidative stress in gonadal tissues, leading to decreased antioxidant enzyme activities and impaired spermatogenesis (Figure 1), as observed in medaka fish exposed to polystyrene NPs [30]. Excessive or prolonged oxidative stress can overwhelm Nrf2 defenses, causing mitochondrial dysfunction, disrupted energy metabolism, DNA damage, and reduced sperm motility and viability [38,39]. Proteomic and transcriptomic analyses reveal that NPs exposure dysregulates pathways related to protein transport, RNA splicing, and mTOR signaling, further impairing sperm function [40]. The limited antioxidant capacity of sperm makes them particularly vulnerable to oxidative damage, which can compromise fertilization and early embryo development [39,41]. Overall, these findings highlight oxidative stress and Nrf2 pathway dysregulation as central mechanisms of NPs-induced reproductive toxicity in fish sperm, emphasizing the need for further research on protective strategies [42].
Such impairments drastically lower fertilization success, and prolonged or chronic exposure may ultimately lead to widespread reproductive failure, threatening the survival and stability of fish populations [43]. The underlying mechanisms by which NPs impair sperm function were explored in several of the included studies, revealing a combination of oxidative stress, hormonal imbalance, and transcriptional reprogramming. In zebrafish, transcriptomic analysis following NPs exposure identified the differential expression of thousands of genes involved in meiosis, DNA repair, and chromatin remodeling [18]. These large-scale molecular changes suggest that NPs interfere with the genetic and epigenetic regulation of spermatogenesis, potentially leading to defective sperm formation. Similarly, Zheng et al. [25] reported alterations in lipid metabolism pathways and abnormal folliculogenesis in zebrafish exposed to size-dependent NPs, linking metabolic disruption to impaired gamete production. Endocrine disruption was another consistent observation.
Sun et al. [20] showed that chronic exposure to NPs altered circulating levels of estradiol, testosterone, and vitellogenin in zebrafish, indicating hypothalamic–pituitary–gonadal (HPG) axis disruption. Zhang et al. [22] confirmed these findings, demonstrating that NPs transferred maternally to offspring caused hormone imbalances and disrupted sex differentiation. At the cellular level, oxidative stress and apoptosis were recurring mechanisms. For instance, Contino et al. [24] documented increased ROS production and mitochondrial dysfunction in mussel sperm, while Sun et al. [20] observed oxidative damage in zebrafish gonads. These molecular and cellular disruptions collectively explain the observed reductions in motility, viability, and reproductive success.
Comparing across species highlights both shared vulnerabilities and species-specific differences in responses to NPs exposure. Freshwater fish models, particularly zebrafish, dominate the literature and consistently show systemic reproductive impairments, including reduced sperm motility, altered gonadal histology, endocrine disruption, and multigenerational effects [18,20,22]. In contrast, European whitefish demonstrated subtler immediate effects on fertilization capacity but significant impacts on offspring development [21]. Marine invertebrates such as mussels and oysters also exhibited acute spermiotoxicity upon direct exposure to functionalized NPs, with effects evident within hours [24,26]. These results highlight the importance of the exposure route: direct sperm exposure tends to reveal immediate impairments in motility and fertilization, whereas chronic waterborne or maternal transfer exposures reveal systemic and generational consequences. The consistent finding across taxa is that NPs impair reproductive success, but the severity and manifestation of effects differ depending on species, NPs properties, and experimental context.
3.1. Sperm Motility
Sperm motility was one of the most consistently assessed parameters across the included studies and appears to be a sensitive indicator of NPs toxicity. In European whitefish (Coregonus lavaretus), direct exposure of sperm to carboxyl-coated PS-NPs (50 nm) caused clear motility impairments in a concentration-dependent manner [21]. At higher particle concentrations, the proportion of motile sperm decreased significantly, and swimming trajectories were disrupted, suggesting that NPs’ interference with sperm surface interactions or flagellar function directly compromises motility. Interestingly, although fertilization rates were not drastically reduced, the offspring derived from exposed sperm exhibited reduced body mass and impaired swimming ability, indicating that sub-lethal effects on sperm may carry consequences into early life stages.
Similar motility impairments were observed in marine invertebrate species, such as mussels and oysters, when sperm were exposed directly to NPs. In Mytilus galloprovincialis, amino-modified 50 nm PS particles significantly reduced sperm motility, increased ROS production, and lowered fertilization rates [24]. Likewise, in Crassostrea gigas, exposure to amine- and carboxyl-functionalized PS-NPs impaired sperm motility and embryogenesis, highlighting that particle surface chemistry is a critical determinant of toxicity [26]. Similarly, in zebrafish both acute [18] and chronic [20] exposures reported motility reductions associated with altered spermatogenesis and gonadal histology. Collectively, these studies provide strong evidence that motility reduction is a primary and early endpoint of NPs toxicity in aquatic organisms, although the severity of impairment varies by species, particle size, functionalization, and exposure conditions. However, the underlying mechanisms of toxicity, the effects of dose–response relationships in normal and abnormal physiology of spermatogenesis, and the potential long-term or intergenerational consequences for fish reproduction and population dynamics remain unassessed.
3.2. Effects on Sperm Viability and Fertilization Capacity
Several studies have focused on the effect of NPs on sperm viability and the ability of gametes to achieve successful fertilization in aquatic species. Yaripour et al. [21] demonstrated that while fertilization rates were not significantly impaired, the resulting offspring showed reduced body mass and lower swimming performance in European whitefish (Coregonus lavaretus). This indicates that even when sperm retain fertilization capacity, NPs may cause subtle cellular or molecular alterations that carry forward into offspring development. In zebrafish, both multigenerational [22] and acute exposure studies such as Pujol et al. [18] reported reduced egg production, delayed hatching, and decreased larval survival, all of which can be linked to compromised sperm or oocyte quality. These results contrast with the effect of NPs in invertebrates such as mussels and oysters, where NPs exposure led to marked decreases in fertilization rates. For example, M. galloprovincialis sperm exposed to increasing concentrations of amino-modified NPs showed not only decreased motility but also reduced viability and mitochondrial function, leading to compromised fertilization success [24]. Similarly, C. gigas sperm exposed to functionalized NPs exhibited dose-dependent decreases in fertilization and abnormal embryogenesis [26]. Taken together, these findings suggest that NPs affect not only the immediate functionality of sperm but also the broader reproductive outcome, including embryo quality and early survival. However, results differ by species and exposure method, indicating a need for standardized testing across taxa.
Rising water temperatures linked to global climate change can modify the toxicity of NPs in aquatic organisms [44]. Although direct experimental evidence remains limited, rising water temperatures associated with global climate change may act as an important modifying factor of NPs toxicity in fish reproduction. However, it has been documented that elevated temperatures can increase metabolic rate, membrane fluidity, and mitochondrial activity in ectothermic organisms, potentially intensifying reactive oxygen species production and reducing cellular stress tolerance [45,46,47]. In sperm cells, which rely on tightly regulated energy metabolism and possess limited antioxidant defenses, thermal stress may therefore exacerbate NPs-induced oxidative damage and functional impairment [48]. These considerations suggest that temperature could influence the magnitude of NPS effects on sperm quality, even if it has not yet been systematically evaluated.
3.3. Transgenerational and Developmental Effects
Evidence for transgenerational effects of NPs in fish is currently limited to a small number of studies and is primarily restricted to parental (F0) exposure with measurable effects in the first filial generation (F1). While these studies provide clear evidence of multigenerational responses, NPs-induced reproductive toxicity has been demonstrated to extend beyond the directly exposed generation, raising hypotheses regarding possible population-level impacts. Zhang et al. [22] reported maternal transfer of NPs in zebrafish, with F1 offspring displaying disrupted gene expression in sex differentiation pathways, altered hormone levels, and impaired gonadal development. This study demonstrated that NPs can accumulate in parental tissues and be transferred to gametes and embryos, resulting in persistent developmental and reproductive alterations in offspring. Similarly, Pujol et al. [18] showed that NPs-exposed zebrafish parents produced larvae with delayed hatching, reduced survival, and abnormal cardiac function. Zheng et al. [25] also observed impaired gametogenesis and disrupted lipid metabolism that persisted in the next generation, further supporting the occurrence of multigenerational effects. Collectively, these findings indicate that NPs exposure is not limited to acute, individual-level toxicity; however, extrapolation to long-term population-level consequences remains speculative and requires further multi-generational and ecologically relevant studies.
4. Knowledge Gaps
Despite growing evidence, substantial gaps remain in our understanding of NPs’ effects on fish-sperm quality and reproduction (Figure 2). As in other studies of ecotoxicology in fish, there is a clear bias toward freshwater model species such as zebrafish, with marine fish species largely underrepresented despite their ecological and commercial importance. Few studies have specifically examined the effect of plastic pollution on spermatogenesis through the lens of proteomic and transcriptomic analysis in fish, and most investigations focus on whole gonadal tissue, which makes it difficult to isolate sperm-specific pathways of toxicity. Notably, the majority of studies did not directly expose sperm to NPs under conditions simulating the aquatic environment, even though sperm in natural waters are highly sensitive and fragile. Direct exposure of sperm to NPs may be critical because gametes in aquatic environments are often the first point of contact for pollutants and may respond differently than intact gonadal tissue or systemic exposure.
A major limitation across the available literature is the frequent use of high or acute NPs exposure concentrations, often in the mg/L range, which may exceed currently reported environmental levels. Field measurements generally indicate that NPs occur at much lower concentrations, typically in the ng/L to μg/L range in surface waters [49,50], although substantial uncertainty remains due to analytical limitations and the lack of standardized detection methods [51,52]. Consequently, many experimental exposure scenarios likely represent worst-case conditions rather than average environmental exposure. Another limitation of the current literature is the reliance on gonadal, hormonal, or transcriptomic endpoints in the absence of direct sperm-specific measurements, limiting conclusions on functional sperm quality and fertilization potential. Further, relatively few studies directly compare single- and multi-stressor exposure scenarios using standardized sperm-specific endpoints, limiting the ability to disentangle whether NPs act predominantly as primary toxicants or as modulators of co-contaminant effects under environmentally realistic conditions. Further from the discussion, it can be said that temperature may modulate NPs toxicity, and empirical studies explicitly examining interactions between NPs and thermal stress on sperm quality are currently lacking. Future research should prioritize multi-stressor experiments incorporating environmentally relevant temperature scenarios to better reflect climate-driven changes in aquatic systems.
Fish reproductive sensitivity to NPs is influenced by diverse life-history traits, but this area remains underexplored. Species with external fertilization and broadcast spawning release sperm directly into water, exposing sperm immediately to NPs during activation and motility, increasing vulnerability to NPs toxicity [15,53]. In contrast, species with internal fertilization, nest-guarding, or benthic spawning may have reduced or altered exposure routes and durations, potentially lowering direct sperm–NP interactions [54]. Additionally, interspecific differences in sperm longevity from seconds to minutes or longer affect the window of susceptibility to NPs, especially during the critical fertilization period [19,53]. NPs can accumulate in gonadal tissues, disrupt the hypothalamic–pituitary–gonadal axis, induce oxidative stress, and impair spermatogenesis and oogenesis, leading to reduced fecundity and abnormal offspring across fish species [15,25,53]. These findings highlight the need for targeted research on species-specific reproductive vulnerabilities to NPs, considering reproductive strategies, sperm traits, and spawning environments to better assess ecological risks and inform aquaculture management [53]. Finally, standardized protocols for assessing sperm endpoints, including motility, viability, fertilization success, and molecular markers are needed to enable cross-study comparisons and meta-analyses.
5. Conclusions
The evidence synthesized from the selected studies demonstrates that NPs pose a significant threat to fish reproductive health, with sperm quality emerging as a particularly sensitive target. Across multiple species, NPs exposure consistently impaired sperm motility, viability, and fertilization capacity, while also inducing molecular and cellular alterations, including oxidative stress, apoptosis, and endocrine disruption. These effects were observed not only in freshwater fish species but also in marine invertebrates such as mussels and oysters, highlighting the broad ecological relevance of NPs toxicity. Importantly, several studies revealed transgenerational consequences, showing that NPs exposure in parental generations can disrupt offspring development, hormone regulation, and gametogenesis. This underscores the long-term population-level implications of NPs pollution in aquatic environments.
The findings summarized in this review also have important implications for aquaculture and hatchery-based reproduction. In cultured species, sperm quality is a key determinant of fertilization success, larval performance, and stock sustainability. Evidence that NPs impair sperm motility, viability, and fertilization capacity suggests that male broodstock exposed to NPs either through water, feed, or contaminated rearing systems may experience reduced reproductive efficiency. Moreover, studies involving direct sperm exposure indicate that NPs present in activation water or hatchery environments could directly compromise assisted reproduction procedures, including in vitro fertilization and sperm handling protocols. These observations highlight the need for monitoring and mitigating NPs contamination in aquaculture systems, as well as for incorporating male reproductive endpoints into broodstock health assessments and risk evaluation frameworks.
Despite these findings, critical knowledge gaps remain. Most notably, very few studies directly exposed sperm to NPs under conditions reflecting the aquatic environment, even though sperm are highly fragile and may respond differently than whole gonadal tissue or systemic exposures. Additionally, there is limited information on marine fish species, environmentally relevant NPs concentrations, and the combined effects of multiple stressors. Therefore, it is recommended that further studies are conducted to investigate direct sperm exposure, mechanistic pathways, and multi-generational effects in ecologically realistic scenarios. Addressing these gaps is essential for accurately assessing the risks of NPs to fish populations and for developing strategies to mitigate their impact on aquatic ecosystems.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wilkinson J. Hooda P.S. Barker J. Barton S. Swinden J. Occurrence, fate and transformation of emerging contaminants in water: An overarching review of the field Environ. Pollut.201723195497010.1016/j.envpol.2017.08.03228888213 · doi ↗ · pubmed ↗
- 2Zhu X. Yang F. Pang Q. Peng F. Xu B. Wang L. Xie L. Zhang W. Tian L. Hou J. Fluvial dissolved organic matter quality modulates microbial nitrate transformation: Enhanced denitrification under low carbon-to-nitrate ratio Environ. Sci. Technol.202559234562346510.1021/acs.est.5c 0710441117426 · doi ↗ · pubmed ↗
- 3Noyes P.D. Miranda D. Oliveira de Carvalho G. Perfetti-Bolaño A. Guida Y. Barbosa Machado Torres F. Barra R.O. Climate change drives persistent organic pollutant dynamics in marine environments Commun. Earth Environ.2025636310.1038/s 43247-025-02348-4 · doi ↗
- 4Thushari G.G.N. Senevirathna J.D.M. Plastic pollution in the marine environment Heliyon 20206 e 0470910.1016/j.heliyon.2020.e 0470932923712 PMC 7475234 · doi ↗ · pubmed ↗
- 5Tekman M.B. Walther B. Peter C. Gutow L. Bergmann M. Impacts of Plastic Pollution in the Oceans on Marine Species, Biodiversity and Ecosystems WWW Germany Berlin, Germany 2022
- 6Gutierrez M.F. Andrade V.S. Ale A. Monserrat J.M. Roa-Fuentes C.A. Herrera-Martínez Y. Wiegand C. Responses of freshwater organisms to multiple stressors in a climate change scenario: A review on small-scale experiments Environ. Sci. Pollut. Res.2025324431444410.1007/s 11356-025-36034-x 39903437 · doi ↗ · pubmed ↗
- 7Wang H.P. Piferrer F. Chen S. Shen Z.G. Sex Control in Aquaculture John Wiley & Sons Chichester, UK 201810.1002/9781119127291 · doi ↗
- 8Gigault J. Ter Halle A. Baudrimont M. Pascal P.Y. Gauffre F. Phi T.L. El Hadri H. Grassl B. Reynaud S. Current opinion: What is a nanoplastic?Environ. Pollut.20182351030103410.1016/j.envpol.2018.01.02429370948 · doi ↗ · pubmed ↗