Generalists or Specialists? Testing Genetic Specificity of Leucocytozoon Lineages and Black Fly Vectors in Thailand
Waraporn Jumpato, Wannachai Wannasingha, Kingkan Sakundet, Chavanut Jaroenchaiwattanachote, Tongjit Thanchomnang, Wanchai Maleewong, Peter H. Adler, Pairot Pramual

TL;DR
This study explores how Leucocytozoon parasites and black flies in Thailand are genetically linked, revealing new parasite lineages and evidence of co-evolution.
Contribution
The study identifies three novel Leucocytozoon lineages and provides evidence of co-speciation between black fly species and parasites.
Findings
12 Leucocytozoon lineages were detected, including three novel and genetically distinct ones.
Phylogenetic analysis showed that closely related black fly species host genetically similar Leucocytozoon lineages.
Cophylogeny analysis revealed significant co-evolution between Leucocytozoon and black fly species.
Abstract
Understanding host–vector–parasite relationships is essential for effective disease control. Black flies are blood-sucking insects that transmit a variety of pathogens causing diseases in humans and other animals. Avian blood parasites of the genus Leucocytozoon are transmitted exclusively by black flies, with only one known exception. In this study, we examined the diversity and genetic specificity between Leucocytozoon parasites and black fly species from Thailand. Six species of black flies were genetically screened for Leucocytozoon infection. In total, 12 Leucocytozoon lineages were detected, three of which are novel and genetically distinct from previously recorded lineages. Phylogenetic analyses revealed evidence of cospeciation between black fly species and Leucocytozoon lineages. Lineages infecting closely related species of black flies were genetically more similar, suggesting…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
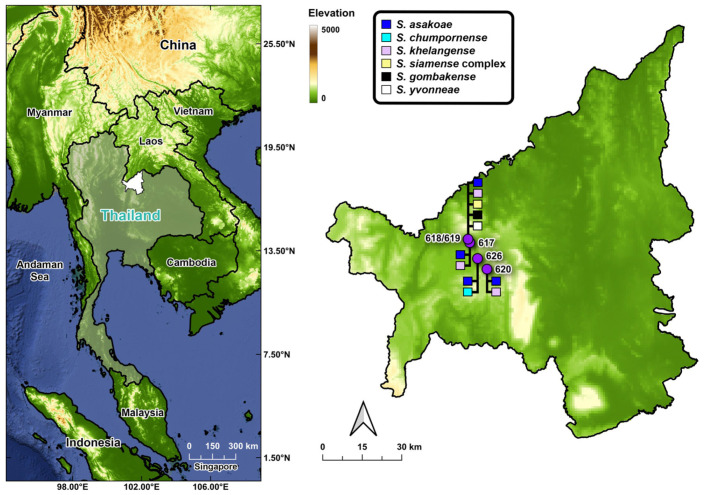 Figure 1
Figure 1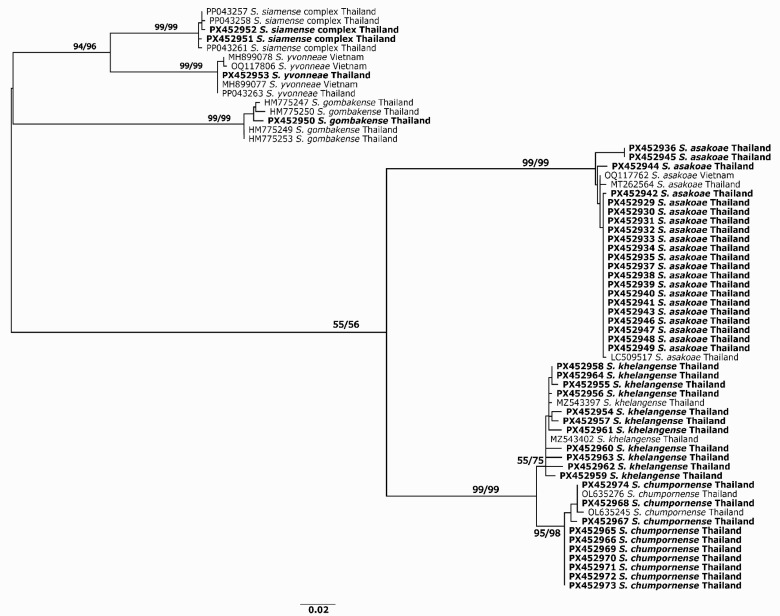 Figure 2
Figure 2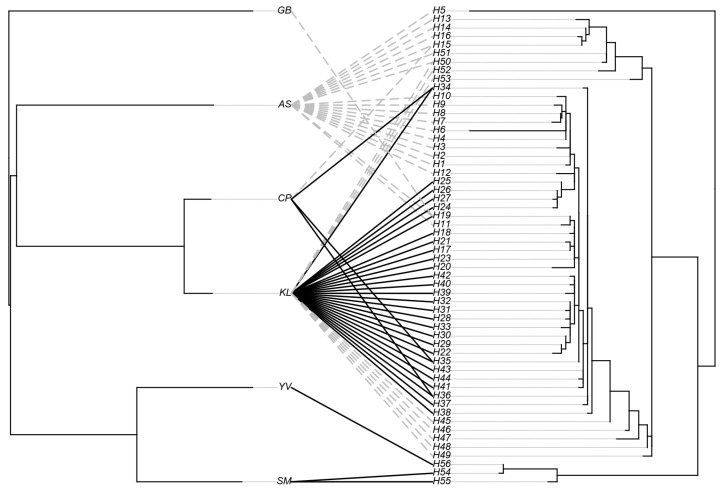 Figure 3
Figure 3- —Mahasarakham University
- —National Research Council of Thailand (NRCT)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBird parasitology and diseases · Coccidia and coccidiosis research · Phytoplasmas and Hemiptera pathogens
1. Introduction
Knowledge of parasite–vector–host interactions is essential for understanding disease epidemiology [1,2,3] and for controlling associated diseases [4]. Key questions in this system are whether parasites are transmitted only by particular vector species (specialists) or by diverse vectors (generalist), and what are the mechanisms that drive such patterns. Several factors can shape parasite–vector relationships, including genetic and physiological compatibility [5], vector-feeding behavior [6], and abiotic factors such as climate [7].
Leucocytozoon Sambon, 1908 (Haemospororida: Plasmodiidae) is a genus of haemosporidian parasites infecting a wide range of avian hosts [8]. Morphologically, at least 45 species have been described [9], but molecular studies have revealed much greater diversity, with more than 1500 lineages recognized (1542 lineages in the MalAvi database) [10], (accessed 31 October 2024). Leucocytozoon infections are generally subclinical; however, some species, such as Leucocytozoon simondi Mathis & Léger, 1910, L. marchouxi Mathis & Léger, 1910, and L. toddi Sambon, 1908, are pathogenic. The disease leucocytozoonosis, caused by Leucocytozoon infections, can reduce growth rate and egg production, and in severe cases can cause host mortality. Symptoms are typically more severe in young birds than in adults and in poultry compared with wild birds [11,12].
All Leucocytozoon species are transmitted by black flies (Diptera: Simuliidae), except Leucocytozoon (Akiba) caulleryi Mathis & Léger, 1909, which is transmitted by biting midges (Diptera: Ceratopogonidae) [13,14]. At least 48 species of black flies serve as vectors of Leucocytozoon [14,15,16,17,18,19]. The majority of Leucocytozoon species can be transmitted by multiple black fly species. For example, the pathogenic species Leucocytozoon simondi is transmitted by at least seven species (Cnephia ornithophilia Davies, Peterson & Wood, 1962; Simulium anatinum Wood, 1963; S. dogieli (Rubtsov, 1956); S. meigeni (Rubtsov & Carlsson, 1965); S. rugglesi Nicholson & Mickel, 1950; S. usovae (Golini, 1987); and the S. venustum Say, 1823 complex). Similarly, L. schoutedeni Rodhain, Pons, Vandenbranden & Bequaert, 1913, a blood protozoan parasite infecting domestic chickens [20], is transmitted by various black fly species [17]. However, some Leucocytozoon lineages exhibit vector specificity, such as the IGRYS1 lineage, which has been found only in S. annulus (Lundström, 1911) [6].
In Thailand, the prevalence of Leucocytozoon infections in vertebrate hosts varies from 2–8% in wild birds and 29% in captive birds [21] up to 89% in domestic chickens [22]. At least three black fly species (Simulium asakoae Takaoka & Davies, 1995, S. chumpornense Takaoka & Kuvangkadilok, 2000, and S. khelangense Takaoka, Srisuka & Saeung, 2022) are candidate vectors of Leucocytozoon among birds, especially domestic chickens in Thailand [16,23]. Previous studies also found genetic associations between Leucocytozoon lineages and black fly species, particularly S. asakoae vs. S. khelangense/S. chumpornense [23]. Jumpato et al. [23] proposed that these Leucocytozoon–simuliid associations result from parasite–vector coadaptation in response to seasonal temperature, which influences the phenology of the black flies. To further investigate this hypothesis, we examined Leucocytozoon infections in co-occurring black fly species and tested the genetic differentiation and cophylogeny between the parasites and their vectors.
2. Materials and Methods
2.1. Collection and Identification of Black Flies
Five collections of black flies were made in villages, around animal pens at the forest edge, and in the forest at three sites in Phu Ruea District, Loei Province, in northeastern Thailand on 5 March 2022 and 4–5 February 2023 (Figure 1 and Table 1). Adult black flies were collected by sweeping aerial nets in a figure-8 motion 0.5–2.0 m above ground. Specimens were fixed in 80% ethanol and stored at −20 °C until processed. Black flies were morphologically identified using a key to the black flies of Thailand [24]. Because females of some closely related species (Simulium siamense Takaoka & Suzuki, 1984 complex and S. yvonneae Takaoka & Low, 2018; S. chumpornense and S. khelangense) are difficult to identify morphologically, DNA barcoding was also used.
2.2. DNA Barcoding of Black Flies
The GF-1 Nucleic Acid DNA extraction kit (Vivantis Technologies Sdn Bhd, Subang Jaya, Malaysia) was used to extract DNA from individuals, following the manufacturer’s protocol. The primers LCO1490 and HCO2198 [25] were used to amplify an approximately 650-bp fragment of the COI gene. The PCR reaction conditions followed those of Tangkawanit et al. [26]. The PCR products were dyed with Novel Juice (GeneDireX, Taoyuan, Taiwan, Republic of China) and examined using 1% agarose gel electrophoresis. Successful amplifications were subsequently purified using the PureDirex PCR CleanUp & Gel Extraction Kit (Bio-Helix, New Taipei City, Taiwan, Republic of China), following the manufacturer’s instructions. The purified PCR products were sequenced at ATCG Company Limited (Thailand Science Park, Pathumthani, Thailand) with the same primers used for PCR.
2.3. Molecular Detection of Leucocytozoon
DNA was extracted from individual black flies, using the same method as in the COI gene study. The nested PCR method and primer described by Hellgren et al. [27], with the PCR reaction conditions of Jumpato et al. [16], were used to amplify an approximately 500-bp fragment of the cyt b gene. PCR products were checked, purified, and sequenced using the same procedure as for the COI gene but with the specific primers for Leucocytozoon.
2.4. Data Analysis
COI sequences from black flies (accession nos. PX452929–PX452974) and cyt b from Leucocytozoon (accession nos. PX511765–PX511810) were checked for quality using the “Edit/View Sequencer Files” option in MEGA X [28]. The COI sequences (650 bp) of the black flies were compared with those in the NCBI GenBank, and conspecific sequences were retrieved for phylogenetic analyses. Phylogenetic relationships of black flies were examined using neighbor joining (NJ) and maximum likelihood (ML) methods. The NJ tree was inferred in MEGA X using the Kimura 2-parameter model and bootstrapping with 1000 replications for testing branch support. The ML tree was also inferred in MEGA X using the GTR+I+G model. Branch support was calculated using bootstrapping with 1000 replications. The output trees were visualized in FigTree v.1.4.3 (https://tree.bio.ed.ac.uk/software/figtree/) (accessed on 30 July 2025).
Leucocytozoon lineages were identified using the BLAST search option in the MalAvi database [10] (accessed on 27 March 2025). The population pairwise FST was used to test genetic differentiation of Leucocytozoon in different populations and black fly species. To test the hypothesis that Leucocytozoon lineages in black flies from different populations differed genetically, cyt b sequences of these protozoa from the same black fly collection were treated as a population regardless of species. Similarly, to test the genetic differentiation between Leucocytozoon lineages in different black fly species, all cyt b sequences of these protozoa from the same black fly species, regardless of sampling location, were treated as the same population. The pairwise FST analysis was calculated in Arlequin ver 3.5 [29] with 1023 permutations for statistical tests. The Bonferroni correction was used to adjust the significance level for multiple tests. We examined whether Leucocytozoon lineage assemblages differed between sampling locations and between black fly species, using analysis of similarity (ANOSIM) [30]. The ANOSIM analysis was performed in PAST version 1.81 [31].
To determine the congruence between black fly hosts and Leucocytozoon phylogenies, we used two global fit methods: Procrustean Approach to Cophylogeny (PACo) [32] and ParaFit [33]. The ML trees inferred from COI haplotypes (650 bp) (Supplementary Materials S1) of representative black fly species (6 species) and cyt b (473 bp) Leucocytozoon lineages (n = 56) (Supplementary Materials S2) reported in black flies from Thailand [16,23], including those found in this study, were used as input trees. The ML trees of black fly species (Supplementary Materials S3) and Leucocytozoon lineages (Supplementary Materials S4) were inferred in IQ-TREE [34,35] web server version (http://iqtree.cibiv.univie.ac.at/) (accessed on 19 April 2025). The best-fit model for the ML trees of black fly species and Leucocytozoon lineages based on the Bayesian information criterion were GTR+F+G4 and K3Pu+F+R2, respectively. The ultrafast bootstrap method with 1000 replications was used to calculate branch support for black fly species and Leucocytozoon ML tree analyses.
For the PACo analysis, significant associations of parasite and vector phylogenies were determined using the global goodness of fit statistic (residual sum of squares in Procrustes superimposition or m2xy) based on 100,000 permutations. PACo analysis was implemented in R 4.5.0, using the ape ver. 5.8.1 [36] and vegan ver. 2.8-0 [37] packages. For the ParaFit analysis, both global and individual host-parasite links were tested based on statistical significance values estimated from 999 permutations with the Cailliez correction for negative eigenvalues. The ParaFit analysis was performed in the software package ape in R 4.5.0.
3. Results
3.1. Leucocytozoon Prevalence and Diversity
We collected a total of 1081 adult black flies. Morphological identification supplemented with DNA barcoding revealed six species, all in the subgenus Gomphostilbia Enderlein, 1921: Simulium asakoae, S. chumpornense, S. khelangense, S. siamense complex, S. gombakense Takaoka and Davies, 1995 and S. yvonneae (Table 1, Figure 2). Two species, S. khelangense (n = 733) and S. asakoae (n = 256), represented the majority of specimens (Table 1). A total of 410 flies was used for molecular detection of Leucocytozoon, including representatives of S. asakoae (n = 150), S. khelangense (n = 220), S. chumpornense (n = 30), and all adults of the S. siamense complex (n = 6), S. gombakense (n = 2) and S. yvonneae (n = 2) (Table 1). In total, 46 individuals (11.2%) of six black fly species were positive for Leucocytozoon (Table 1), mostly S. asakoae (45.7%), S. khelangense (23.9%), and S. chumpornense (21.7%). Two individuals of the S. siamense complex and one each of S. gombakense and S. yvonneae also were positive. These latter three species are reported here for the first time as possible vectors of Leucocytozoon.
Twelve haplotypes (H1–H12) were identified among 46 cyt b sequences of Leucocytozoon. Comparisons of these cyt b haplotypes with those in GenBank indicated that most (8 of 12) are of an unidentified species (Leucocytozoon sp.) reported in chickens, with sequence similarity of >99%. One haplotype (H8) was identical to Leucocytozoon schoutedeni (accession no. DQ676823) in a chicken (Gallus gallus (L., 1758)) from Uganda. Haplotype H9 in Simulium asakoae was genetically highly similar (99%) to Leucocytozoon recorded in a raptor (accession no. MT281505). Three remaining haplotypes, two in the S. siamense complex (H10 and H11) and one in S. yvonneae (H12), were genetically distinct from all remaining Leucocytozoon in our study. The top blast hit for H10 was Leucocytozoon sp. (accession no. KX223864) in a Sandhill crane (Grus canadensis (L., 1758)) from the USA, with sequence similarity of 92%. H11 was most similar (92%) to Leucocytozoon sp. in the village weaver Ploceus cucullatus (Müller, 1776) (accession no. MT761629). H12 was genetically closest (92%) to Leucocytozoon sp. (accession no. LC782814) in wild birds from Japan.
Comparisons with haplotypes of Leucocytozoon in the MalAvi database [10] (accessed on 31 March 2025) showed that five haplotypes (H4–H8) were genetically identical with four existing lineages (GALLUS06, GALLUS17, GALLUS35, and GALLUS46). H6 and H7 matched 100% with GALLUS35 but were treated here as different haplotypes because they had single nucleotide differences in the final 20-bp of the cyt b sequences, which were longer than those in the MalAvi database. The remaining seven haplotypes, therefore, were new Leucocytozoon lineages.
3.2. Association of Leucocytozoon and Black Fly Species
Among 12 Leucocytozoon haplotypes, only two were shared by two or more black fly species (Table 2). H4 (Leucocytozoon sp.) and H8 (L. schoutedeni) were shared by Simulium asakoae, S. khelangense, and S. gombakense and by S. asakoae and S. chumpornense, respectively. Leucocytozoon–vector associations, based on Parafit tests, revealed significant co-phylogeny between black fly species and Leucocytozoon lineages (ParaFitGlobal value = 0.0171, p = 0.0024). Significant co-phylogeny was also supported by PACo analysis (m^2^xy = 0.475, p < 0.001; R^2^ = 0.525). Among 63 individual parasite–vector links, 34 exhibited significant p-values (<0.05) with ParaFit1 and ParaFit2 (Figure 3).
ANOSIM analysis revealed no significant difference between Leucocytozoon assemblages in different populations (R = −0.4375, p = 0.9728). However, Leucocytozoon assemblages in different black fly species differed significantly (R = 0.6406, p = 0.0284). The FST values indicated that only one pair of comparisons between Leucocytozoon from different populations differed significantly (Table 3). In contrast, all comparisons between Leucocytozoon lineages from different black fly species were significantly different except between the Simulium siamense complex and S. khelangense and between S. asakoae and S. chumpornense (Table 4).
4. Discussion
We report Leucocytozoon infections in Simulium gombakense, the S. siamense complex, and S. yvonneae for the first time. These black flies are in the subgenus Gomphostilbia and possess claws with a large basal tooth, an adaptation for feeding on birds [38]. Thus, the detection of avian haemosporidia in these species is not unexpected. The Leucocytozoon lineage in S. gombakense, which was also detected in S. asakoae and S. khelangense, corresponds to GALLUS17 in the MalAvi database [10], previously recorded from domestic chickens. Although the blood hosts of S. gombakense are unknown, the presence of this parasite suggests that S. gombakense feeds on chickens.
A Leucocytozoon lineage previously identified in raptors was detected in Simulium asakoae, providing the first evidence that this species probably feeds on raptors. Leucocytozoon lineages in the S. siamense complex and S. yvonneae, with only 92% sequence similarity to existing records from diverse hosts (e.g., Sandhill crane, village weaver, and unidentified wild birds from Japan), indicated that they are possible novel species [39]. These results highlight the need for further surveys of Leucocytozoon diversity in wild birds of Thailand, as well as blood-meal analysis of the S. siamense complex and S. yvonneae to identify the parasite sources.
The ParaFit test shows significant co-phylogeny between four black fly species and Leucocytozoon lineages. Simulium khelangense has the strongest signal, with 27 co-phylogenetic lineages, while its close relative S. chumpornense is significantly associated with three of these. However, both species also have multiple non-significant parasite associations, indicating they are generalist vectors. Similarly, S. asakoae transmits a broad diversity of Leucocytozoon lineages, consistent with its generalist feeding behavior.
Vector specificity in haemosporidian parasites could arise either as a by-product of host blood preference—where parasites are specific to avian hosts and vectors acquire them through blood feeding—or by direct vector–parasite coevolution. Such associations have been demonstrated across parasite genera (e.g., Plasmodium–Culicidae, Haemoproteus–Ceratopogonidae, and Leucocytozoon–Simuliidae) [40]. At broader taxonomic levels, however, both specialist and generalist relationships occur [41]. To date, at least 48 species of black flies have been incriminated as vectors or candidate vectors of Leucocytozoon [14,15,16,17,18,19], with most species exhibiting low specificity. Exceptions include S. annulus, which is specifically associated with lineage IGRYS1 due to its apparently restricted host associations with only certain groups of birds, notably cranes [6] and loons (family Gaviidae) [42].
Avian host associations of the black flies provide further context for understanding parasite–vector associations. Simulium khelangense and S. chumpornense primarily feed on chickens [43], but S. khelangense also feeds on turkeys [43] and owls [23]. Simulium asakoae feeds predominantly on chickens and occasionally on humans [43], and based on our study, it is also linked to raptors. These broad host ranges explain the high genetic diversity of Leucocytozoon lineages in these species. In contrast, the blood hosts of the S. siamense complex and S. yvonneae remain unknown, making it unclear whether their observed parasite associations result from host preferences or direct parasite–vector coevolution.
Although some of the black flies in our study are generalists, significant genetic structuring of Leucocytozoon lineages exists between coexisting black fly species, whereas differentiation among geographic populations is relatively low. An exception occurred in Ban Pla Ba, where lineages were predominantly restricted to S. chumpornense. Previous studies reported genetic differentiation of Leucocytozoon between S. asakoae and S. khelangense [23], with hypothesized temperature-driven coadaptation. However, our data do not support this explanation, as samples were collected under the same environmental conditions. Moreover, given that S. asakoae, S. chumpornense, and S. khelangense all feed primarily on chickens, genetic structuring is unlikely to result from host preference.
A more plausible explanation for genetic structuring of Leucocytozoon lineages between coexisting black flies is vector–parasite compatibility. Parasite development success differs significantly among vector–parasite combinations [44,45]. Vectors can evolve resistance mechanisms to limit parasite infections that reduce fitness, while parasites evolve immune-evasion strategies to complete their development [9]. Closely related black fly species are expected to share similar immune responses [46], favoring infection by genetically related Leucocytozoon lineages. Our findings support this idea: lineages associated with S. chumpornense are genetically similar to those in S. khelangense, with three of the four co-phylogenetic lineages shared between them.
5. Conclusions
Our study suggests that vector–parasite compatibility contributes to the observed associations between black fly species and Leucocytozoon lineages. Genetically similar Leucocytozoon lineages are either exclusively (e.g., Simulium siamense complex and S. yvonneae) or predominantly (e.g., S. chumpornense and S. khelangense) associated with closely related black flies. This pattern indicates that parasite adaptation to intrinsic vector traits, such as immunity [46,47] or microbiome composition [48], might play an important role in shaping vector specificity. Further research into these mechanisms is needed to elucidate the dynamics of Leucocytozoon–simuliid interactions. Our results for some species are based on limited sample sizes (S. siamense complex: n = 6; S. gombakense: n = 2; S. yvonneae: n = 2). Additional studies with larger samples are needed before firm conclusions can be drawn.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Penczykowski R.M. Laine A.L. Koskella B. Understanding the ecology and evolution of host–parasite interactions across scales Evol. Appl.20169375210.1111/eva.1229427087838 PMC 4780374 · doi ↗ · pubmed ↗
- 2Shaw W.R. Catteruccia F. Vector biology meets disease control: Using basic research to fight vector-borne diseases Nat. Microbiol.20194203410.1038/s 41564-018-0214-730150735 PMC 6437764 · doi ↗ · pubmed ↗
- 3Cecílio P. Cordeiro-da-Silva A. Oliveira F. Sand flies: Basic information on the vectors of leishmaniasis and their interactions with Leishmania parasites Commun. Biol.2022530510.1038/s 42003-022-03240-z 35379881 PMC 8979968 · doi ↗ · pubmed ↗
- 4Agosta S.J. Janz N. Brooks D.R. How specialists can be generalists: Resolving the” parasite paradox” and implications for emerging infectious disease Zoologia 20102715116210.1590/S 1984-46702010000200001 · doi ↗
- 5Beerntsen B.T. James A.A. Christensen B.M. Genetics of mosquito vector competence Microbiol. Mol. Biol. Rev.20006411513710.1128/MMBR.64.1.115-137.200010704476 PMC 98988 · doi ↗ · pubmed ↗
- 6Hellgren O. Bensch S. Malmqvist B. Bird hosts, blood parasites and their vectors—Associations uncovered by molecular analyses of blackfly blood meals Mol. Ecol.2008171605161310.1111/j.1365-294X.2007.03680.x 18266623 · doi ↗ · pubmed ↗
- 7Chapa-Vargas L. Matta N.E. Merino S. Effects of ecological gradients on tropical avian hemoparasites Avian Malaria and Related Parasites in the Tropics Santiago-Alarcon D. Marzal A. Springer Cham, Switzerland 2020349377
- 8Valkiūnas G. Avian Malaria Parasites and Other Haemosporidia CRC Boca Raton, FL, USA 2005