Effects of TGFBR1 on Proliferation of Dermal Papilla Cells in Fine-Wool Sheep
Tong Xiao, Yu Luo, Chao Yuan, Yufang Song, Jianxiang Tang, Zengkui Lu, Jianbin Liu, Tingting Guo

TL;DR
This study shows that TGFBR1 suppresses the growth of dermal papilla cells in fine-wool sheep, affecting hair follicle development and wool fiber diameter.
Contribution
The novel contribution is identifying TGFBR1 as a negative regulator of dermal papilla cell proliferation through multiple signaling pathways in fine-wool sheep.
Findings
TGFBR1 overexpression suppresses dermal papilla cell proliferation in fine-wool sheep.
TGFBR1 modulates Wnt/β-catenin, BMP, and Notch signaling pathways to influence hair follicle development.
Downregulation of TGFBR1 in fine-wool sheep correlates with reduced wool fiber diameter.
Abstract
This study investigates the regulation of dermal papilla cells proliferation by TGFBR1, a differentially expressed gene identified through single-cell transcriptomic sequencing. The aim is to elucidate the molecular mechanisms through which TGFBR1 governs dermal papilla cells and the role of TGFBR1 in hair follicle development of fine-wool sheep. Single-cell transcriptomic sequencing data revealed significant downregulation of TGFBR1 in dermal papilla cells of ultra-fine-wool sheep, suggesting that this gene serves as a key negative regulator for wool fiber diameter. To validate this hypothesis, we constructed TGFBR1 overexpression plasmids and knockdown vectors for transfection into dermal papilla cells. Combined with functional assays such as EDU and CCK-8, we confirmed TGFBR1 exerts negative control over dermal papilla cell proliferation. Mechanistic studies revealed that TGFBR1…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
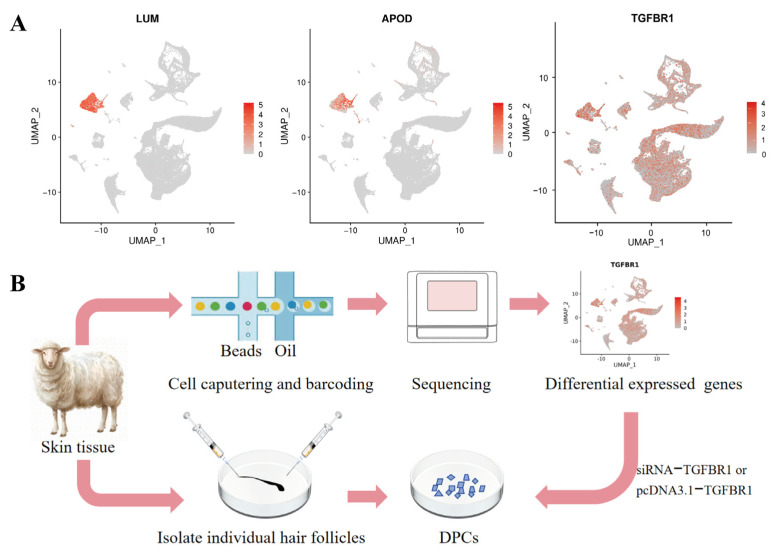 Figure 1
Figure 1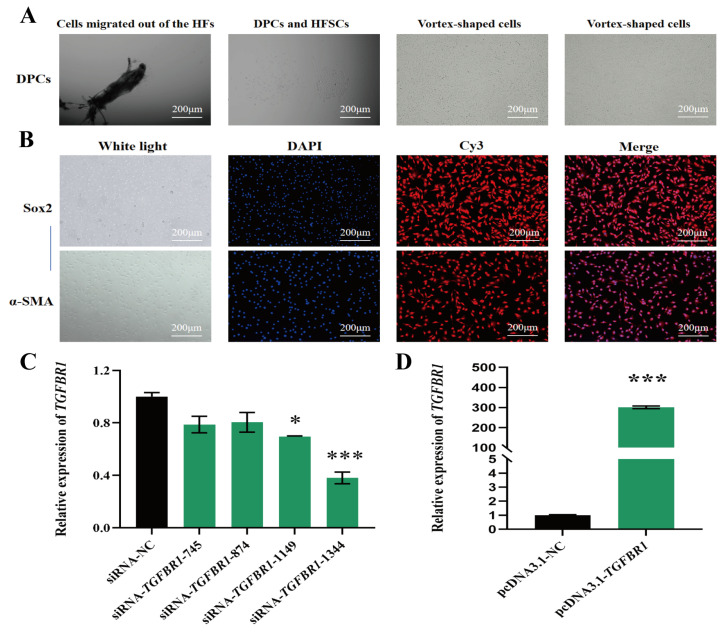 Figure 2
Figure 2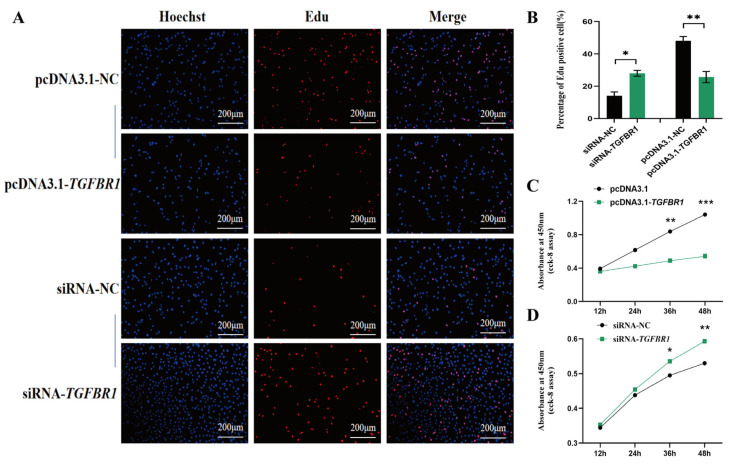 Figure 3
Figure 3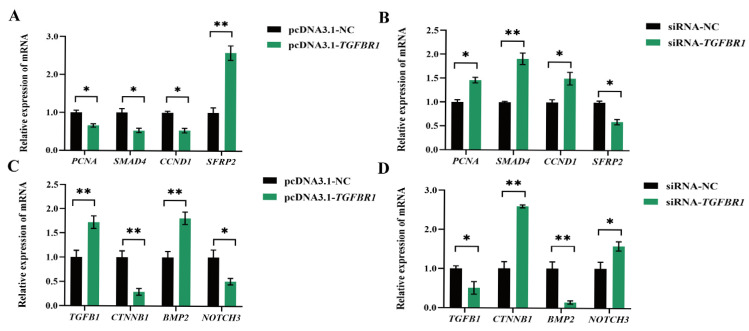 Figure 4
Figure 4- —Science and Technology Innovation 2030-Major Program
- —Natural Science Foundation of Gansu Province
- —China Agriculture Research System
- —Chinese Academy of Agricultural Sciences Technology Innovation Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHair Growth and Disorders · Skin and Cellular Biology Research · Cancer and Skin Lesions
1. Introduction
Wool, as a vital natural textile material, possesses excellent biodegradability and renewability. Its unique biological properties, such as thermal insulation and felting capability, make it highly valued in the textile industry, conferring significant economic value [1]. As DPCs are the biological “production factories” of wool fibers, the structure and functional state of DPCs directly determine key physical properties such as fiber diameter (FD) and curvature. Among the diverse cellular components of the HFs, DPCs specialized mesenchymal cells located at the basal region of HFs play a central role in follicular morphogenesis, cyclic growth, and developmental regulation. They serve as the “signaling hub”, initiating and sustaining the hair cycle, releasing multiple growth factors via paracrine mechanisms, while activating key signaling pathways to precisely regulate follicular development and structural homeostasis [2]. Research indicates that the number and volume of DPCs are closely correlated with the FD, curvature morphology, and cyclical growth characteristics of HFs. When the number of DPCs falls below a critical threshold, even intact epithelial structures cannot initiate new hair growth, leading to follicular dysfunction [3]. Therefore, in-depth analysis of the biological characteristics of DPCs and their molecular regulatory mechanisms is crucial for understanding HF development and wool trait formation.
TGFBR1 is a key member of the serine/threonine protein kinase family, playing a vital role in the TGF-β/Smad signaling pathway. Its primary function is to mediate intracellular signal transduction as a transmembrane receptor [4]. By recognizing and binding the TGF-β ligand, this receptor translates extracellular signals into intracellular cascades, thereby extensively regulating key biological processes such as cell proliferation, migration, differentiation, and apoptosis [5,6]. Beyond activating the classical Smad-dependent pathway, TGFBR1 can activate other signaling pathways through non-canonical mechanisms, such as MAPK (mitogen-activated protein kinase) [7], PI3K (phosphatidylinositol-3-kinase) [8], and AKT (A serine/threonine kinase) [9], demonstrating its multifunctionality and complexity within cellular signaling networks. Numerous studies have elucidated TGFBR1’s regulatory functions across different cell types. For instance, all-trans retinoic acid (ATRA) reduces DPC proliferation and induces apoptosis via the TGF-β/Smad pathway, thereby influencing HF development [10]. MIR-22 regulates the downstream of smad3 signaling pathway by targeting TGFBR1, thereby inhibiting myoblast proliferation and promoting differentiation [11]. Knockdown of TGFBR1 in ST2 cells accelerates adipogenic differentiation [12]; Let-7a inhibits cell proliferation by targeting TGFBR1 in cervical carcinoma [13]. These studies collectively demonstrate that TGFBR1, as a key regulatory node, broadly participates in the molecular regulation of diverse cellular fate decisions. However, the specific biological mechanisms of this gene in DPC proliferation and differentiation remain unclear and warrant further systematic investigation.
Based on the aforementioned research background, this study uses fine-wool sheep as experimental subjects to systematically elucidate the biological functions of TGFBR1 in HF development. First, tissue localization analysis clarifies the expression characteristics of TGFBR1 for HF structures in skin. Subsequently, molecular biological techniques such as gene knockdown and overexpression are employed to regulate TGFBR1 expression levels in in vitro cultured DPCs, investigating its effects on key biological processes including DPCs proliferation and activity. Furthermore, by integrating studies on HF development-related signaling pathways, we elucidate the molecular mechanisms through which TGFBR1 participates in regulating DPC proliferation and HF morphogenesis. This research not only provides new experimental evidence for understanding the role of the TGF-β family in HF development but also lays a theoretical foundation for investigating the molecular regulatory networks governing HF development in fine-wool sheep breeding.
2. Materials and Methods
2.1. Animal Tissue Samples
All experimental animals used in this study were sourced from the Fine-Wool Sheep Core Breeding Farm (Gansu Provincial Sheep Breeding Technology Promotion Station, Sunan, Gansu, China). Under identical rearing conditions, six 14-month-old Alpine Merino ewes of the same paternal lineage were selected and divided into three groups of two each based on wool count and FD, an ultra-fine group (FD: <18 μm), a medium-fine group (FD: 18.120.0 μm), and a fine group (FD: 20.121.5 μm), for sampling. Skin tissue samples (0.5 cm × 0.5 cm) were collected from the scapular region of the experimental sheep using a skin biopsy punch (ACUDERM, Fort Lauderdale, FL, USA). Blood was rinsed off the tissue with PBS, and samples were preserved in PBS containing penicillin and streptomycin. Cell isolation experiments were conducted immediately upon arrival at the laboratory. The animal study protocol was approved by the Institutional Ethics Committee of the Institute of Animal Sciences, Chinese Academy of Agricultural Sciences (IAS-CAAS) (permit no. SYXK-2014-0002). Approval date: 13 May 2022.
2.2. ScRNA-Seq Data Analysis
ScRNA-seq data were generated using the 10× Genomics platform with Illumina Nova PE150 sequencing (Illumina, San Diego, CA, USA). Raw data underwent quality control using Cell Ranger (v.3.0), followed by clustering analysis with Seurat (v.5.2.0) and differential gene expression analysis. This study focused on cell lineages harboring LUM-labeled [14] and APOD-labeled [15] DPCs. TGFBR1, the gene selected for validation, was identified as a downregulated gene in the intergroup difference between ultra-fine group and fine group.
2.3. Isolation, Purification, and Immunofluorescence of DPCs
Under aseptic conditions, we collected a 0.5 cm × 0.5 cm skin tissue sample from the posterior margin of the shoulder blade of Alpine Merino sheep. DPCs were isolated using enzymatic digestion and mechanical dissociation. The skin tissue sample was immersed in 75% ethanol for 30 s, then rinsed with 10× PBS and 1× PBS separately for 35 min each. Wool and adipose tissue were trimmed from the skin using sterile surgical scissors. The tissue was then dissected into small pieces along the direction of HF growth and digested in 0.25% trypsin solution at 37 °C for 1.52 h. Once softened, we used a 1 mL syringe to isolate individual HF units. We incised the terminal bulbus pilosus and extruded the dermal papilla. We transferred the dermal papilla to a 48-well plate for culture using a pipette tip. Within 6~7 days, DPCs emerged from the papilla and adhered to the culture surface. Once densely populated, the cells were digested and passaged. Leveraging their adherent nature, DPCs were purified using differential digestion. After three consecutive purifications, DPCs more than 95% pure were obtained.
Identification of DPC marker proteins α-SMA and SOX2 was performed using immunofluorescence staining. Fourth or fifth -generation DPCs were seeded onto plates. When cells reached 60~70% confluence, medium was removed, cells were washed three times with PBS, and cells were fixed with 4% paraformaldehyde for 20 min. After three PBS washes, cells were permeabilized with 0.5% Triton X-100 for 10 min, followed by three PBS washes and blocking with 3% BSA for 1 h. After three PBS washes, we added 1:100 diluted primary antibody specific to the marker protein and incubated the cells overnight at 4 °C. We washed the cells three times with PBS, then added 1:100 diluted Cy3-labeled secondary antibody (goat anti-rabbit) (Biyun Tian, Shanghai, China) and incubated the cells for 1 h at room temperature in the dark. Finally, we washed the plates three times with PBS, stained the cell nuclei with DAPI at room temperature for 10 min, and obtained images using a confocal microscope. All cell cultures were performed at 37 °C in a 5% CO_2_ incubator.
2.4. Construction of Overexpression and Interference Vectors
Based on the TGFBR1 gene coding sequence published in the NCBI database, gene-specific primers containing double restriction sites for HindIII and EcoRI were designed. The products were directionally cloned into the eukaryotic expression vector to construct the pcDNA3.1(+) recombinant plasmid. The negative control vector pcDNA3.1-NC, recombinant plasmid pcDNA3.1-TGFBR1, and siRNA-TGFBR1 were designed and synthesized by GenePharma (Suzhou, China). Sequence information is shown in Table 1.
2.5. Real-Time Fluorescent Quantitative PCR (RT-qPCR)
Total RNA was extracted from DPCs using the Trizol method, with total RNA concentration measured by UV spectrophotometry (IMPLEN, Palo Alto, CA, USA). Total RNA samples extracted from DPCs following successful transfection (Zeta Life, San Francisco, CA, USA) were reverse-transcribed using a Prime Script™ RT Reagent Kit with gDNA Eraser (Vazyme, Nanjing, China) to obtain cDNA fragments. RT-qPCR primers were designed using NCBI Primer-Blast based on gene coding sequences published in the NCBI database. Primers were synthesized by Sangon Biotech Co., Ltd. (Shanghai, China) (Table 2). Reaction reagents were added to RNAse-free Eppendorf tubes, with β-actin serving as the internal control. Reaction protocol was 94 °C pre-denaturation for 30 s, 94 °C denaturation for 10 s, and 60 °C extension for 30 s, repeated for 40 cycles.
2.6. Cell Proliferation and Viability Assays
EDU proliferation assay: DPCs were cultured until they reach a well-established, morphologically stable state. Cell proliferation was assessed using an EDU assay kit (Biyun Tian, China) 24 h after transfection with overexpression plasmids and siRNA. The EDU solution was diluted in medium to a 1:500 ratio. We added equal volumes to 6-well plates in a 1:1 ratio, incubated the plates for 2 h, aspirated the medium, and added 4% paraformaldehyde for 15 min at room temperature. After discarding the fixative, we washed the plates 3 times with 3% BSA-PBS solution for 3~5 min. We added 0.3% Triton X-100 permeabilization solution and incubated the plates for 15 min. The plates were thoroughly washed, then 500 μL Click reaction solution was added to each well. We gently shook the plate to ensure complete mixing of medium and reaction solution. We incubated the plate at room temperature in the dark for 30 min. After incubation, the plate was washed 3 times. We added Hoechst 33,342 reaction solution to the cells for nuclear staining. After 10 min, we observed cell proliferation under a fluorescence microscope and document images.
CCK-8 assay: Transfected DPCs were seeded into 96-well plates with three biological replicates per group. CCK-8 solution was added in the dark. After incubation at 37 °C in a 5% CO_2_ incubator for 2, 12, 24, 36, and 48 h, absorbance was measured at 450 nm.
2.7. Data Statistics and Analysis
RT-PCR results were analyzed for relative quantitative analysis by 2^−ΔΔCt^ method. One-way ANOVA conducted with SPSS 22.0 (IBM, Armonk, NY, USA) software was used for statistical analysis. Experiments were designed with at least three biological replicates. All results are expressed as mean ± standard deviation (mean ± SD). * p < 0.05, ** p < 0.01, and *** p < 0.001 indicate statistical significance. All bar charts were generated using GraphPad Prism 10.0.
3. Results
3.1. Differentially Expressed Genes from DPCs in Ultra-Fine Group and Fine Group of Fine-Wool Sheep
ScRNA-seq analysis revealed 147 significantly differentially expressed genes between the two DPC populations, comprising 71 significantly upregulated genes and 76 significantly downregulated genes (Table S1). UMAP visualization revealed that cell regions highly expressing LUM and APOD could be identified as DPCs (Figure 1A). Notably, among the differentially expressed genes, TGFBR1 exhibited widespread expression in the hair papilla region, suggesting its role in regulating DPC function.
3.2. Isolation, Purification, and Identification of DPCs
This study successfully isolated primary DPCs from fine-wool sheep using a combination of mechanical separation and enzymatic digestion, establishing a stable in vitro culture system (Figure 1B). Between 6 and 7 days post-follicle isolation, DPCs were observed migrating from the dermal papilla and adhering to the bottom of the culture dish. Their initial morphology was predominantly triangular, with plump cell bodies and abundant cytoplasm. As culture duration increased, cell morphology gradually changed, losing the initial triangular shape. The first passage was performed after around 10 days of culture. In the primary cell culture, in addition to the target DPCs, there were mixed populations of hair follicle stem cells (HFSCs) with a pebble-like morphology and long spindle-shaped fibroblast-like cells. After morphological stabilization, differential digestion ultimately yielded DPCs exhibiting stable growth characteristics, irregular spindle-shaped morphology, and characteristic vortex-like arrangement (Figure 2A). Immunofluorescence results revealed the significant positive expression of both markers in the cultured cells, thereby further confirming that the isolated cells were DPCs that were suitable for subsequent functional validation experiments (Figure 2B).
3.3. TGFBR1 Inhibits Proliferation of DPCs
RT-qPCR results showed that compared with the control and the empty vector control groups, TGFBR1 mRNA expression levels were significantly elevated in the overexpression group, confirming successful establishment of the overexpression system (Figure 2D). Among the four screened siRNA interference fragments, each siRNA exhibited varying degrees of reduced expression. However, TGFBR1-siRNA-1344 demonstrated the most pronounced knockdown efficiency, significantly downregulating TGFBR1 transcription levels. Consequently, TGFBR1-siRNA-1344 was selected as the effective interference tool for subsequent functional experiments (Figure 2C).
EDU assay results revealed that TGFBR1 overexpression significantly reduced the EDU-positive cell ratio, indicating marked inhibition of DPCs proliferation. Consistent with this, CCK-8 assays showed lower absorbance values in the overexpression group at all time points compared to the control group, further confirming TGFBR1’s inhibitory effect on DPC proliferation (Figure 3A–C). Conversely, after TGFBR1 knockdown, the EDU-positive cell ratio significantly increased, and CCK-8 assay results similarly demonstrated enhanced cell proliferation activity (Figure 3A,B,D).
3.4. TGFBR1 Regulates Genes Associated with HF Development
Experimental results demonstrated that TGFBR1 overexpression in DPCs significantly downregulated PCNA and CCND1 transcription levels. Concurrently, mRNA expressions of CTNNB1, NOTCH3, and SMAD4 were markedly suppressed. Correspondingly, the gene expressions of SFRP2, BMP2, and TGFB1 were significantly upregulated (Figure 4A,C). This expression pattern was perfectly validated in the reverse experiment: upon TGFBR1 knockdown, the suppressed genes (PCNA, CCND1, CTNNB1, NOTCH3, SMAD4) showed significantly increased expression, while the activated genes (SFRP2, BMP2, TGFB1) exhibited markedly decreased expression (Figure 4B,D).
4. Discussion
As vital skin appendages, HFs serve as the fundamental biological basis for regulating hair formation and its key economic traits (e.g., diameter, curvature). Their morphogenesis and cyclical growth depend on intricate collaboration among multiple cell types, including DPCs, HFSCs, and matrix cells (Mxs) [16]. During embryonic development, HF formation relies on a series of interactions between the epidermal basal layer and dermal mesenchyme. Postnatally, localized mesenchymal cell clusters gradually differentiate into DPCs, which are subsequently enveloped by Mxs. These cells persist as DPCs, continuously influencing HF growth and development [17]. Furthermore, this process is tightly regulated by multiple signaling pathways, including the Wnt/β-catenin pathway, which promotes HF initiation and growth [18,19]; the BMP signaling pathway, involved in cell fate determination [20,21]; the SHH signaling pathway, which regulates morphogenesis [22]; the Notch signaling pathway, affecting stem cell maintenance and differentiation [23,24]; and the transforming growth factor TGF-β family, which is extensively involved in regulating cellular behavior [25]. These pathways form a complex regulatory network through paracrine and autocrine mechanisms, collectively determining the developmental progression and homeostasis of HFs. TGFBR1, as the specific receptor for the TGF-β signaling pathway, plays crucial roles in diverse tissues and cells. Typically, TGF-β ligand binding to its receptor activates downstream Smad proteins, thereby regulating target gene expression and influencing cellular processes including proliferation, differentiation, and apoptosis [26]. Studies indicate that TGFBR1 mediates Smad phosphorylation and downstream signaling to influence myoblast proliferation and differentiation [27]. Furthermore, microRNA-98 suppresses cardiac fibroblast differentiation by targeting TGFBR1, highlighting the receptor’s broad role in cell fate determination [28]. However, although the functions of TGFBR1 in various biological systems are increasingly well-defined, its specific mechanism of action in DPCs in fine-wool sheep remains to be systematically elucidated.
This study functionally validated TGFBR1 at the cellular level, which is a differentially downregulated gene in DPCs that was identified through scRNA-seq. The proliferative capacity of DPCs is fundamental to maintaining their population size and performing their functions, being crucial for the normal growth and development of HFs as well as their cyclical growth [29]. EDU and CCK-8 assays demonstrated that TGFBR1 expression directly influences DPCs proliferation capacity: overexpression significantly suppressed DPC proliferation, manifested as reduced EDU-positive cell proportions and decreased CCK-8 absorbance values. Conversely, knocking down TGFBR1 expression markedly enhanced cellular proliferation, suggesting TGFBR1 acts as a negative regulator of DPC proliferation. This further supports the critical role of the TGF-β signaling pathway in initiating the HF regression phase or maintaining the quiescent state of DPCs during the resting phase. Furthermore, the skin samples used for scRNA-seq in this study were collected in August, when the HFs of fine-wool sheep were in a vigorous growth phase. The differential downregulation of TGFBR1 in DPCs from ultra-fine-wool sheep aligns with its proliferation-inhibiting effect observed in functional validation experiments, providing additional evidence for the gene’s negative regulatory role in DPC growth from a natural expression pattern perspective.
RT-qPCR results for HF-development-related genes revealed a potential mechanism through which TGFBR1 influences DPC proliferation. PCNA, a classic marker assessing cell proliferation capacity [30], exhibited significantly reduced mRNA expression following TGFBR1 overexpression, providing molecular evidence for TGFBR1’s inhibitory effect on DPCs proliferation. The Wnt/β-catenin signaling pathway plays a crucial role in DPC proliferation and differentiation [31]. CNND1 and CTNNB1, the core downstream effectors promoting HF formation and development [32], both exhibited significantly downregulated mRNA expression following TGFBR1 overexpression, suggesting TGFBR1 influences Wnt/β-catenin signaling by suppressing downstream gene expression, thereby affecting DPC proliferation. SFRP2, a regulator of the Wnt pathway, plays a role in suppressing HF growth, degeneration, and remodeling by inhibiting keratinocyte proliferation [33]. Its upregulation following TGFBR1 overexpression provides further compelling evidence that TGFBR1 influences the Wnt/β-catenin signaling pathway. TGFB1, belonging to the same transforming growth factor β family as TGFBR1, inhibits HF epithelial cell growth and induces cell degeneration [34]. Its expression increases upon TGFBR1 overexpression, consistent with TGFB1’s known role in suppressing HF epithelial cell proliferation, further reinforcing TGFBR1’s proliferative inhibitory function. The BMP signaling pathway regulates cell proliferation, differentiation, and apoptosis in multiple organs, including skin [35]. In cashmere goat HFs, BMP2 is highly expressed during the resting phase but is barely detected during the active phase [36]. This finding further supports the hypothesis that TGFBR1 promotes the transition of DPCs to a quiescent state. SMAD4 is a key regulator of HF formation and development, synergistically modulating epidermal homeostasis via the TGF-β pathway [37], maintaining the morphological and structural integrity of HFSCs [38], and promoting DPC proliferation [39]. The Notch signaling pathway suppresses cell differentiation to promote proliferation in HFs, thereby maintaining stem cell characteristics, as a key component of this pathway. The NOTCH3 gene likely plays a crucial role in this process [40]. The significant reduction in SMAD4 and NOTCH3 following TGFBR1 overexpression further supports TGFBR1’s negative regulatory role in DPCs. Collectively, these findings indicate that TGFBR1 coordinates multiple signaling pathways, including Wnt/β-catenin, BMP, and Notch, to form a complex molecular network that finely regulates DPC proliferation capacity.
5. Conclusions
This study demonstrates that TGFBR1 is expressed in DPCs and inhibits DPC proliferation. Further mechanistic analysis revealed that TGFBR1 exerts this effect by modulating key signaling pathways, including Wnt/β-catenin, BMP, and Notch. These results identify TGFBR1 as a key negative regulator of DPC proliferation and provide new insights into the molecular mechanisms of hair follicle development. The findings offer potential implications for the genetic improvement of wool traits.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Szatkowski P. The Influence of Various Chemical Modifications of Sheep Wool Fibers on the Long-Term Mechanical Properties of Sheep Wool/PLA Biocomposites Materials 202518305610.3390/ma 1813305640649546 PMC 12251111 · doi ↗ · pubmed ↗
- 2Li J. Zhao B. Yao S. Dai Y. Zhang X. Yang N. Bao Z. Cai J. Chen Y. Wu X. Dermal Papilla Cell-Derived Exosomes Regulate Hair Follicle Stem Cell Proliferation via LEF 1Int. J. Mol. Sci.202324396110.3390/ijms 2404396136835374 PMC 9964005 · doi ↗ · pubmed ↗
- 3Chi W. Wu E. Morgan B.A. Dermal papilla cell number specifies hair size, shape and cycling and its reduction causes follicular decline Development 20131401676168310.1242/dev.09066223487317 PMC 3621486 · doi ↗ · pubmed ↗
- 4Loboda A. Sobczak M. Jozkowicz A. Dulak J. TGF-β1/Smads and mi R-21 in Renal Fibrosis and Inflammation Mediat. Inflamm.2016201611210.1155/2016/8319283 PMC 500560427610006 · doi ↗ · pubmed ↗
- 5Zieba J. Forlenza K.N. Khatra J.S. Sarukhanov A. Duran I. Rigueur D. Lyons K.M. Cohn D.H. Merrill A.E. Krakow D. TGFβ and BMP Dependent Cell Fate Changes Due to Loss of Filamin B Produces Disc Degeneration and Progressive Vertebral Fusions P Lo S Genet.201612 e 100593610.1371/journal.pgen.100593627019229 PMC 4809497 · doi ↗ · pubmed ↗
- 6Zimowska M. Duchesnay A. Dragun P. Oberbek A. Moraczewski J. Martelly I. Immunoneutralization of TG Fbeta 1 Improves Skeletal Muscle Regeneration: Effects on Myoblast Differentiation and Glycosaminoglycan Content Int. J. Cell Biol.2009200965937210.1155/2009/65937220111627 PMC 2809363 · doi ↗ · pubmed ↗
- 7Mu D. Cambier S. Fjellbirkeland L. Baron J.L. Munger J.S. Kawakatsu H. Sheppard D. Broaddus V.C. Nishimura S.L. The integrin αvβ8 mediates epithelial homeostasis through MT 1-MMP–dependent activation of TGF-β1J. Cell Biol.200215749350710.1083/jcb.20010910011970960 PMC 2173277 · doi ↗ · pubmed ↗
- 8Vogelmann R. Nguyen-Tat M.D. Giehl K. Adler G. Wedlich D. Menke A. TGF beta-induced downregulation of E-cadherin-based cell-cell adhesion depends on PI 3-kinase and PTENJ. Cell Sci.20051184901491210.1242/jcs.0259416219695 · doi ↗ · pubmed ↗