Generation of Novel High-Quality Small-Grained Rice Germplasm by Targeting the OsVIN2 Gene
Xi Chen, Yarong Lin, Xiangzhe Xi, Shaohua Yang, Shiyu Wu, Hongge Qian, Mingji Wu, Taijiao Hu, Fating Mei, Mengyan Zheng, Chuanlin Shi, Yiwang Zhu

TL;DR
Scientists used gene editing to create small-grained rice with better quality, including less chalkiness and more protein, without affecting plant growth.
Contribution
A novel method for breeding high-quality small-grained rice by targeting the OsVIN2 gene with CRISPR/Cas9.
Findings
Editing OsVIN2 reduced grain size by 19.9% in length and 15.2% in width.
The edited rice showed 11.0% higher protein and 77.5% less chalkiness.
Plant growth and seed-setting were unaffected by the gene modification.
Abstract
Small-grained rice is valuable for hybrid seed production and food markets, but developing high-quality small-grained varieties is challenging. This study aimed to use the CRISPR/Cas9 gene-editing technique to modify the OsVIN2 gene in rice variety MH86, hoping to reduce grain size while improving quality. We successfully edited OsVIN2, resulting in rice grains that were shorter and narrower than normal. The edited rice also had better quality: less chalkiness, more protein (11.0%), and optimized cooking texture. Importantly, other plant traits like growth and seed-setting remained unaffected. This method provides an effective way to breed high-quality small-grained rice, which can lower hybrid seed production costs and meet consumer demand for better-tasting rice. Small-grained rice varieties are highly valued in hybrid seed production and food markets because of their unique…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
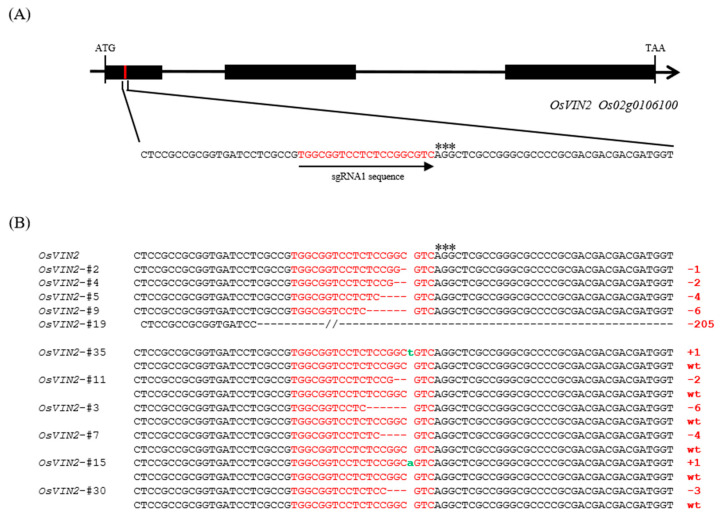 Figure 1
Figure 1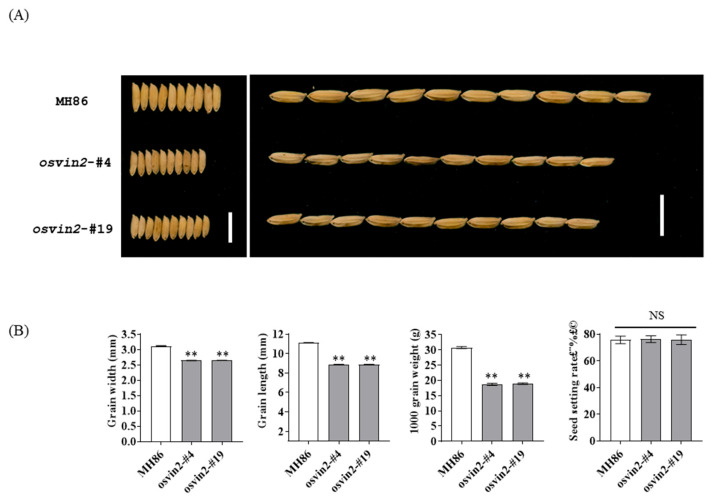 Figure 2
Figure 2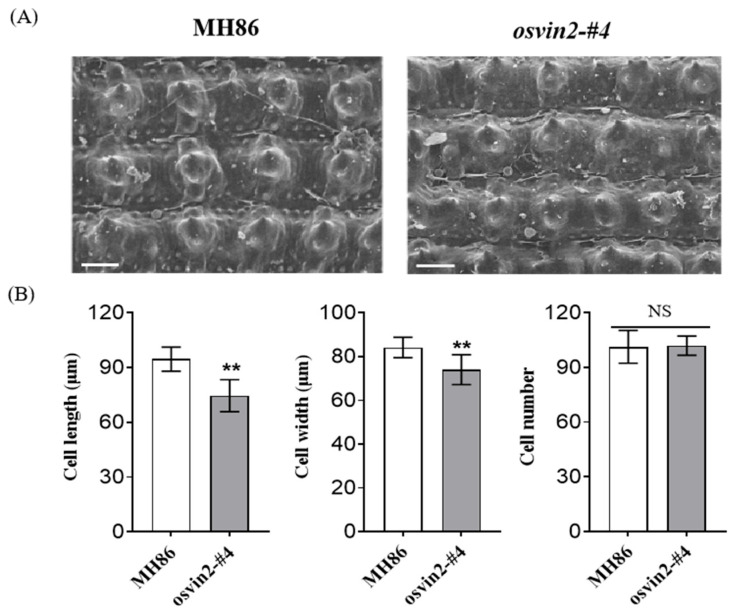 Figure 3
Figure 3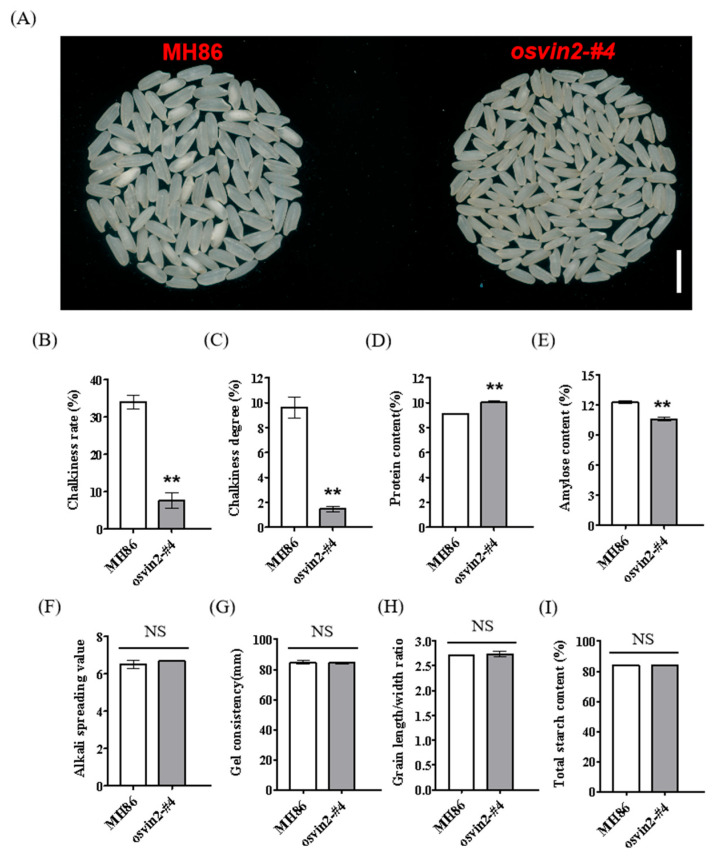 Figure 4
Figure 4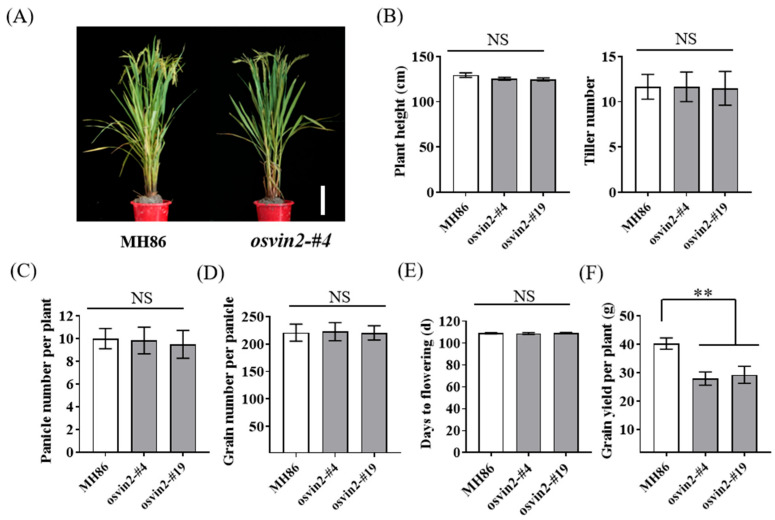 Figure 5
Figure 5- —the National Natural Science Foundation of China
- —the Excellent Youth Foundation of Fujian Academy of Agricultural Sciences
- —the Extended Research Project of National Natural Science Foundation of China
- —‘5511’ Collaborative Innovation Project of Fujian Academy of Agricultural Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant tissue culture and regeneration · GABA and Rice Research · Genetic Mapping and Diversity in Plants and Animals
1. Introduction
Rice grain size, a key agronomic trait, not only determines grain yield but also influences grain quality and application scenarios [1,2]. Among diverse grain size variants, small-grained rice germplasm represents a crucial component of rice genetic diversity. It provides valuable genetic resources for dissecting the molecular regulatory network of grain shape and also has significant practical value in hybrid rice seed production [1,2,3]. Specifically, small-grained male sterile lines can reduce the male-to-female seed ratio during mechanical mixing, lower seed production costs, and improve seed propagation efficiency [4]. Additionally, small-grained rice meets diversified market demands, as it is preferred for traditional snacks and high-quality congee in some regions [5,6]. Thus, developing efficient strategies to create high-quality small-grained rice germplasm is needed in rice breeding.
In recent years, CRISPR/Cas9-mediated gene editing technology has revolutionized crop germplasm innovation due to its high efficiency, precision, and simplicity [7]. This technology has been widely applied to modify grain size-related traits in rice. For example, targeted mutation of GS3 (a major negative regulator of grain length) or GW2 (a negative regulator of grain width) via CRISPR/Cas9 increased grain length or width effectively in many rice varieties [8,9,10], confirming that CRISPR/Cas9 is a powerful tool for grain size modification.
To date, numerous genes regulating rice grain size have been identified, but their application in small-grained germplasm creation remains limited. This is mainly due to two constraints. On the one hand, a large proportion of the identified grain size-related genes are negative regulators [11]. Their loss-of-function mutations typically increase grain size, rendering them unsuitable for creating small-grained varieties via gene editing. On the other hand, the few positive regulators that can reduce grain size when edited often exert adverse effects on other agronomic traits. For instance, the mutation of the OsCSLD4 gene reduced grain size, but it led to side effects on agronomic traits such as poor plant growth, reduced plant yield, and decreased bacterial blight resistance [12]. Therefore, the key challenge lies in selecting suitable target genes that can synchronously optimize grain size and quality without adverse effects on other agronomic traits.
OsVIN2 could be an optimal candidate target gene for creating small-grained rice germplasm. Two independent studies have demonstrated that loss of OsVIN2 function can effectively reduce grain size [13,14]. OsVIN2 encodes a vacuolar invertase, which influences sugar composition, transport, and starch accumulation [13,14,15]. Specifically, in OsVIN2 loss-of-function mutants, starch composition is altered significantly [13]. Since starch accumulation is a key determinant of rice eating quality and affects various traits such as amylose content (AC), gelatinization temperature (GT), gel consistency (GC), chalkiness rate (CR), and chalkiness degree (CD) [16,17,18], it is worth further investigating whether editing OsVIN2 impacts rice eating quality.
MH86 is a widely used indica restorer line in hybrid rice production, with high combining ability, strong disease resistance, and good grain quality [19,20]. However, its grain size is larger than the ideal small-grained type for mechanical seed production. Notably, gene function in rice often shows genetic background dependence. For example, knockout of QSOXL1 significantly reduces seed dormancy levels in japonica varieties, Nanjing7, and Nanjing35, but has no effect on seed dormancy in N22 (an Aus rice variety) [21]. Whether OsVIN2 editing can modify grain size and quality in MH86 remains untested, and thus experimental validation is needed to clarify its efficacy in this genetic background.
In this study, we aim to edit the rice gene OsVIN2 using CRISPR/Cas9 technology to develop new quality-improved germplasm resources of small-grain rice. We hypothesize that precision modification of OsVIN2 can simultaneously reduce grain size and improve quality traits, such as amylose content. This research provides a new approach for breeding small-grain varieties, holding important implications for broader crop improvement strategies.
2. Materials and Methods
2.1. Plant Material and Growth Conditions
The elite indica rice variety Minghui86 (MH86) was used as the wild-type (WT) control and transformation recipient. All WT and transgenic plants were grown in the experimental field of Fuzhou Agricultural Station (26.08° N, 119.28° E), Fujian Province, under natural conditions. The growing season spanned from late April to early October, with standard agronomic management including regular irrigation, fertilization, and pest control. For trait stabilization, 15 T_2_ homozygous mutants were selected for phenotypic analysis to avoid segregating effects.
2.2. sgRNA Design and Vector Construction
The OsVIN2 gene (Os02g0106100) sequence of MH86 was retrieved from the Rice Genome Annotation Project (RGAP, http://rice.plantbiology.msu.edu/, accessed on 28 December 2025). A 20-bp sgRNA target sequence (5′-TGGCGGTCCTCTCCGGCGTC-3′) was designed in the first exon using CRISPR-P 2.0 (http://skl.scau.edu.cn/targetdesign/, accessed on 28 December 2025) [22], ensuring high specificity and minimal off-target potential. Complementary oligonucleotides (Table S1) were synthesized, annealed to form dimers, and cloned into the CRISPR/Cas9 vector pYLCRISPR/Cas9 [23], which contains the Cas9 gene driven by the maize Ubiquitin promoter and the sgRNA under the rice U6 promoter.
2.3. Agrobacterium-Mediated Rice Transformation
The recombinant vector was introduced into Agrobacterium tumefaciens strain EHA105 via electroporation. Calli induced from mature embryos of MH86 were infected with the positive Agrobacterium cultures. Transformation procedures, including callus induction, co-cultivation, selection (hygromycin 50 mg/L), and plant regeneration, followed the protocol described by Hiei et al. [24]. Regenerated T_0_ plants were transplanted to the field and selfed to produce T_1_ and T_2_ generations.
2.4. Genotyping and Off-Target Analysis
Genomic DNA was extracted from young leaves using the CTAB method [25]. The target region of OsVIN2 was amplified with specific primers (F: 5′-CGCTTCCCTACTCCTACTCG-3′, R: 5′-CATCAGCCTCACCATCTCCT-3′) (Table S1) under the following conditions: 94 °C for 5 min; 35 cycles of 94 °C for 30 s, 58 °C for 30 s, 72 °C for 30 s; and 72 °C for 10 min. PCR products were Sanger-sequenced, and mutation types were identified by comparing with WT sequences using DNAMAN 9.0. For off-target detection, 6 potential sites predicted by CRISPR-P were amplified and sequenced in homozygous mutants.
2.5. Agronomic Trait Measurement
At maturity, 15 T_2_ plants from each mutant line (osvin2-#4, #19) and WT were randomly selected for trait analysis. Plant height (distance between the ground surface and the tip of the main panicle), tiller number(number of tillers at maturity), panicle length(distance between the rachis and the tip of the main panicle), and grain number per panicle were recorded (15 plants). Grain length and width were measured using a digital caliper (n = 300 grains per line). 1000-grain weight was determined using three replicates of fully filled grains. All data are presented as mean ± standard deviation (SD), and statistical significance was determined using Student’s t-test (p < 0.01).
3. Results
3.1. CRISPR/Cas9 Target Site Selection for the OsVIN2 Gene
Firstly, sequence analysis revealed that the OsVIN2 gene contains three exons. Considering that the first exon of OsVIN2 encodes a conserved domain critical for vacuolar invertase activity, we envision designing a target site within this region. Based on analysis from the prediction website, a sgRNA with high specificity was selected (Figure 1A). To ensure the suitability of this sgRNA in our selected material (MH86, which is an elite indica rice variety with high yield potential and moderate grain quality), we then amplified the sequence flanking this sgRNA in MH86. Sequence alignment results indicated that the sequence at this site in MH86 is identical to that in reference genome Nipponbare (Figure S1). Therefore, this sgRNA was prioritized for subsequent vector construction.
3.2. CRISPR/Cas9-Mediated Mutagenesis of OsVIN2
The CRISPR/Cas9 expression vector harboring sgRNA was introduced into MH86 calli via Agrobacterium tumefaciens-mediated transformation, and a total of 30 independent T0 transgenic plants were regenerated. Genomic DNA extracted from young leaves was used to amplify and identify the mutations of OsVIN2 gene. Sequencing results showed that 18 plants (60.0% efficiency) carried mutations in OsVIN2, including 5 homozygous mutants, 6 biallelic mutants, and 7 heterozygous mutants (Table 1). Most mutations were small deletions (1–20 bp), accounting for 74.2% of edited alleles, while one line exhibited a large-fragment editing with a 205-bp deletion (Figure 1B; Table S2). These findings demonstrated efficient editing of OsVIN2 using this sgRNA via the CRISPR/Cas9 system.
3.3. Putative Off-Target Analysis
Off-target effects have long been a major concern in the CRISPR/Cas9 system. Thus, evaluating the off-target effects by detecting mutations in potential off-target sequences remains an essential step. Therefore, we used the online tool OffTarget (http://skl.scau.edu.cn/offtarget/, accessed on 28 December 2025) to predict potential off-target sites of sgRNA-OsVIN2. Subsequently, 5 homozygous mutants with distinct mutation (osvin2-#2, #4, #5, #19, #9) were selected to assess the predicted off-target sequences. PCR identification results showed that no mutations were detected in the transgene-free mutant lines (Table 2, Figure S2). These results indicated that the designed sgRNA exhibits high specificity in inducing mutations in the OsVIN2 gene, thereby ensuring the reliability of subsequent phenotypic analyses.
3.4. Phenotypic Analysis of Grain Size in OsVIN2 Mutants
To evaluate the effect of OsVIN2 mutants on grain size, we measured grain dimensions of homozygous transgene-free T_2_ mutants (osvin2-#4, #19) and wild-type MH86. Compared with wild type, the mutant seeds showed a significant reduction in both length (19.9%) and width (15.2%), resulting in a 39.2% decrease in 1000-grain weight (Figure 2). Notably, the seed setting rate of OsVIN2 mutants remained unaffected (Figure 2), indicating that the mutation specifically targeted grain size without influencing reproductive output. Scanning electron microscopy of the lemma and palea revealed that the outer epidermal cells of mutants were smaller than those of wild-type plants, with no significant change in cell number (Figure 3). For cell counting, 10 randomly selected regions per lemma were analyzed, and 20 grains per line were measured. This indicated that the reduced grain size in osvin2 mutants was primarily caused by suppressed cell expansion rather than altered cell division—consistent with the role of vacuolar invertases in regulating cell turgor pressure [26]. These findings confirmed that OsVIN2 effectively regulated cell expansion to determine grain size in rice.
3.5. osvin2 Mutants Confer Superior Grain Quality
With growing consumer demand, rice quality has become a primary focus in rice breeding. This complex trait encompasses multiple dimensions including grain appearance, cooking properties, eating quality, and nutritional value. Accordingly, we analyzed key quality traits of polished grains from osvin2 T_2_ mutants to evaluate improvements in grain quality. The results showed that compared with the wild type, osvin2 mutant grains exhibited superior grain appearance, characterized by significantly reduced chalkiness rate and chalkiness degree (Figure 4A–C). Furthermore, the protein content of osvin2 mutants was 10.1%, higher than the 9.1% in the wild type (Figure 4D). Amylose content (AC) is a critical factor determining rice’s cooking and eating quality. In general, high AC results in distinct, firmer grains, while lower AC yields a softer, stickier texture. Expectedly, the amylose content in osvin2 mutants was noticeably lower than that in wild type (Figure 4E). However, no significant differences were observed in alkali spreading value, gel consistency, grain length/width ratio, or total starch content between osvin2 mutants and the wild type (Figure 4F–I). Collectively, these analyses demonstrate that osvin2 mutants exhibit improved rice quality, specifically manifested in enhanced visual appearance, increased protein content, and optimized amylose content.
3.6. Agronomic Traits of osvin2 Mutants
To assess the impact of OsVIN2 on other agronomic traits, we investigated the field performance of T_2_ mutants. The results showed that the plant height and tiller number of osvin2 mutants remained unchanged (Figure 5A,B). Additionally, panicle number per plant and grain number per panicle of osvin2 mutant lines were comparable to those of the wild type (Figure 5C,D). Importantly, critical growth stages such as heading date and maturity were unaffected in osvin2 mutants (Figure 5E), ensuring their adaptability to local cropping systems. Nevertheless, due to the reduced grain size and decreased 1000-grain weight of the osvin2 mutants, this results in a significant reduction (27.3–30.3%) in grain yield per plant (Figure 5F). These findings indicated that osvin2 mutants had minimal impact on overall plant architecture. The observed yield adjustment was a specific consequence of the targeted grain size modification, rather than a broad disruption of plant growth.
4. Discussion
The development of small-grained rice varieties holds significant practical value for advancing mechanized hybrid rice seed production. Their reduced grain size facilitates precise mechanical sorting during seed production, which can simplify large-scale cultivation, standardize seed quality, and reduce labor costs associated with traditional manual sorting [3,4]. Although this study did not directly measure mechanization efficiency, the size difference between mutant and WT grains (19.9% shorter and 15.2% narrower) provides a theoretical basis for mechanical separation. This study demonstrates that targeted editing of OsVIN2 can simultaneously improve grain quality, including decreased amylose content and palatability, highlighting the multifaceted advantages of this approach.
Previous research cloned a completely recessive grain size QTL gene, TGW5^FH212^, which is a weak allele of the known D1 gene. The introduction of TGW5^FH212^ converts conventional male sterile lines into small-grained sterile lines, increasing grain number while reducing grain size, thus providing a solution for low-cost, high efficiency mechanized hybrid rice seed production. However, it also results in a dwarf phenotype in NIL^FH212^ [4]. In contrast, the osvin2 mutants in this study only showed a significant improvement in grain shape (Figure 2 and Figure 3), without affecting other key agronomic traits such as tiller number, seed-setting rate, and flowering time (Figure 5). This minimal impact on overall plant architecture offers a feasible strategy for enhancing the efficiency of hybrid rice seed production.
The identification of OsVIN2 as a positive regulator of grain weight provides a new molecular target for grain size regulation. Frameshift mutations in the first exon stably produce a small-grain phenotype, demonstrating that disrupting this vacuolar invertase gene is a feasible and actionable methodological approach for reducing grain weight without major pleiotropic effects. Unlike traditional breeding methods that often introduce linked undesirable traits, our approach enables targeted modification of grain size while improving rice quality without significantly compromising important agronomic characteristics such as plant type. The OsVIN2-edited lines developed in this study can be integrated into elite male sterile backgrounds to breed small-grained sterile lines suitable for mechanized hybrid seed production.
5. Conclusions
In summary, this study successfully demonstrated that CRISPR/Cas9-mediated targeted editing of the OsVIN2 gene in the elite indica restorer line MH86 is an effective strategy for creating novel small-grained rice germplasm with superior quality attributes. By precisely modifying the first exon of OsVIN2, we achieved a significant reduction in grain size (19.9% shorter and 15.2% narrower) without compromising key agronomic traits such as tiller number, seed-setting rate, heading date, and plant architecture. Furthermore, the edited lines exhibited substantial improvements in grain quality, including increased protein content, and optimized amylose content. The high specificity of the designed sgRNA, confirmed by the absence of off-target mutations, ensures the reliability and safety of the edited germplasm for future breeding applications. This research not only identifies OsVIN2 as a robust molecular target for synchronously regulating rice grain size and quality but also provides a practical framework for accelerating the development of small-grained rice varieties suitable for mechanized hybrid seed production.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gong D. Zhang X. He F. Chen Y. Li R. Yao J. Zhang M. Zheng W. Yu G. Genetic Improvements in Rice Grain Quality: A Review of Elite Genes and Their Applications in Molecular Breeding Agronomy 202313137510.3390/agronomy 13051375 · doi ↗
- 2Ren D. Ding C. Qian Q. Molecular bases of rice grain size and quality for optimized productivity Sci. Bull.20236831435010.1016/j.scib.2023.01.02636710151 · doi ↗ · pubmed ↗
- 3Lei B. Shao J. Zhang F. Wang J. Xiao Y. Cheng Z. Tang W. Wan J. Genetic analysis and fine mapping of a grain size QTL in the small-grain sterile rice line Zhuo 201SJ. Integr. Agric.2023232155216310.1016/j.jia.2023.07.026 · doi ↗
- 4Ying J. Qin Y. Zhang F. Duan L. Cheng P. Yin M. Wang Y. Tong X. Huang J. Li Z. A weak allele of TGW 5 enables greater seed propagation and efficient size-based seed sorting for hybrid rice production Plant Commun.2024510081110.1016/j.xplc.2024.10081138213029 PMC 11009153 · doi ↗ · pubmed ↗
- 5Zhou H. Xia D. He Y. Rice grain quality—Traditional traits for high quality rice and health-plus substances Mol. Breed.201940110.1007/s 11032-019-1080-6 · doi ↗
- 6Ibarrola-Rivas M.J. Nonhebel S. Regional food preferences influence environmental impacts of diets Food Secur.2022141063108310.1007/s 12571-022-01270-3 · doi ↗
- 7Guo Y. Zhao G. Gao X. Zhang L. Zhang Y. Cai X. Yuan X. Guo X. CRISPR/Cas 9 gene editing technology: A precise and efficient tool for crop quality improvement Planta 20232583610.1007/s 00425-023-04187-z 37395789 · doi ↗ · pubmed ↗
- 8Huang J. Gao L. Luo S. Liu K. Qing D. Pan Y. Dai G. Deng G. Zhu C. The genetic editing of GS 3 via CRISPR/Cas 9 accelerates the breeding of three-line hybrid rice with superior yield and grain quality Mol. Breed.2022422210.1007/s 11032-022-01290-z 37309462 PMC 10248666 · doi ↗ · pubmed ↗