Phylogenetics and evolution of Digitaria grasses, including cereal crops fonio, raishan and Polish millet
George P Burton, Paolo Ceci, Lorna MacKinnon, Lizo E Masters, Noro Fenitra Harimbao Randrianarimanana, Philippa Ryan, Colin G N Turnbull, Tiziana Ulian, Maria S Vorontsova

TL;DR
This study explores the evolution and phylogeny of Digitaria grasses, including important millet crops like fonio and raishan, to identify wild relatives and improve future breeding efforts.
Contribution
The study provides a comprehensive phylogeny and biogeographic history of Digitaria crops, identifying new close wild relatives and their evolutionary divergence.
Findings
Four distinct evolutionary lineages were identified among Digitaria crops.
South and eastern Africa are proposed as the origin of early Digitaria divergence.
Incomplete domestication traits, such as loss of trichomes, were observed without clear morphological changes in spikelets.
Abstract
Millet crops in the grass genus Digitaria include white and black fonio (D. exilis and D. iburua), raishan (D. compacta) and Polish millet (D. sanguinalis), cultivated across West Africa, India and Europe. Fonio and raishan crops are important in supporting food security and subsistence agricultural systems in rural communities, while D. sanguinalis is no longer cultivated. These crops are resilient to challenging climates. We aim to produce an integrated study of these crops: a phylogeny of the Digitaria genus including all four food species, to identify key crop wild relatives; time-calibrated biogeographic analysis, to investigate the history and evolution of Digitaria; and a morphological study to assess the transition between wild and domesticated species. We use the Angiosperms353 target-enrichment sequencing approach to produce maximum likelihood and coalescent model nuclear…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Fig. 1
Fig. 1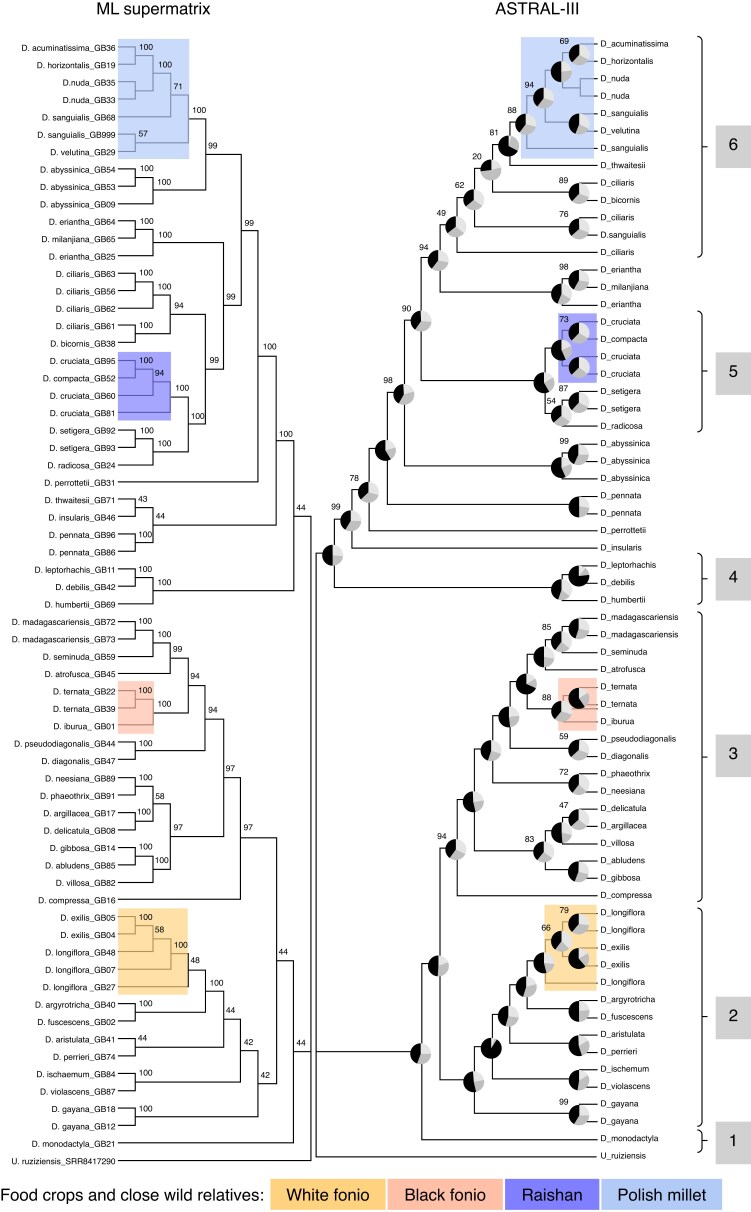 Fig. 2
Fig. 2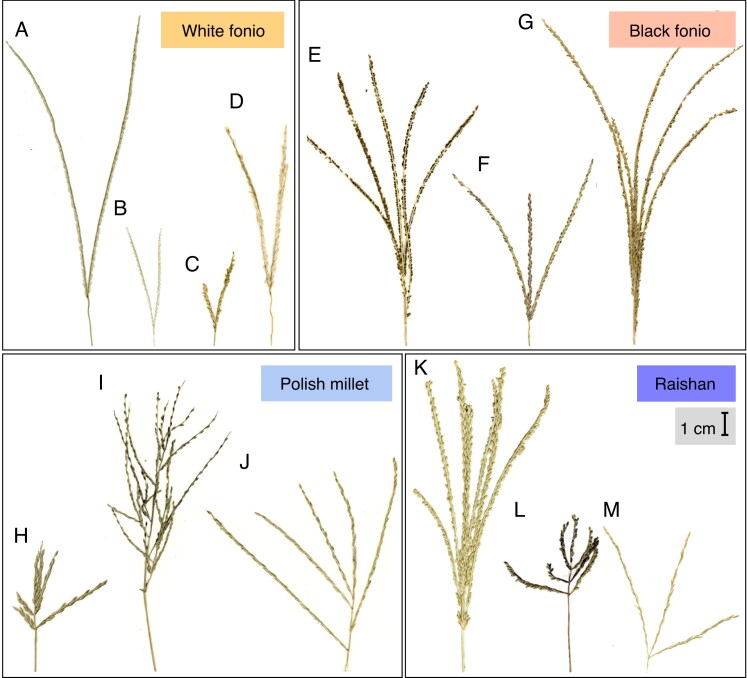 Fig. 3
Fig. 3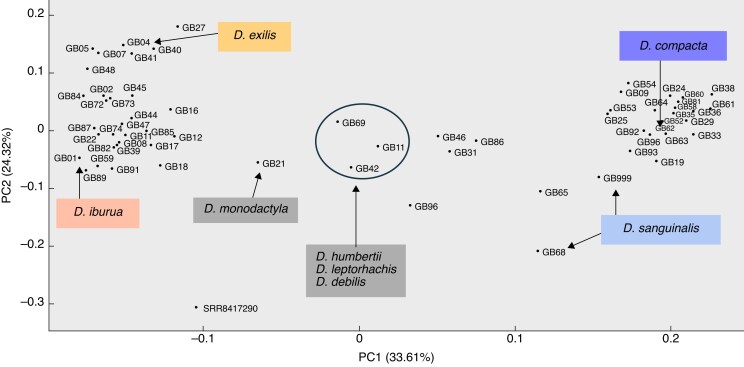 Fig. 4
Fig. 4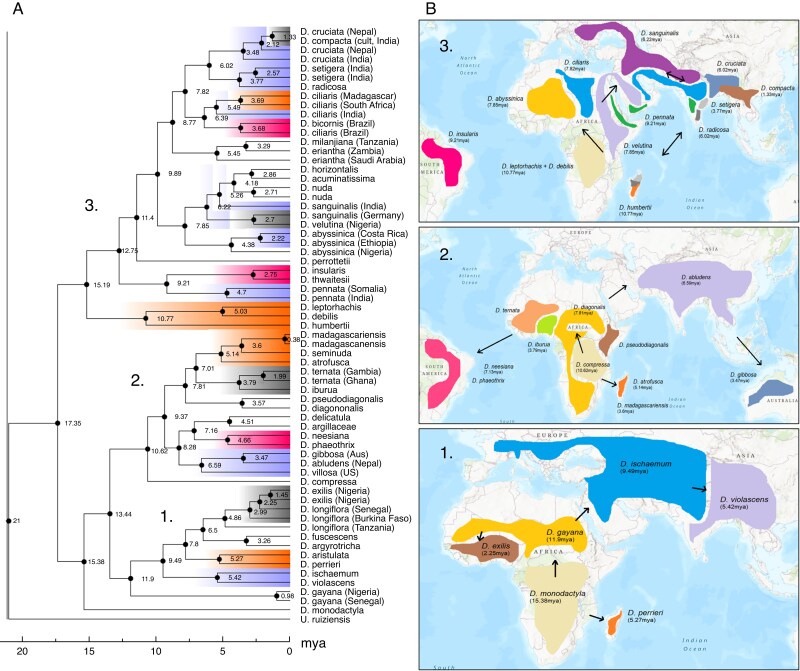 Fig. 5
Fig. 5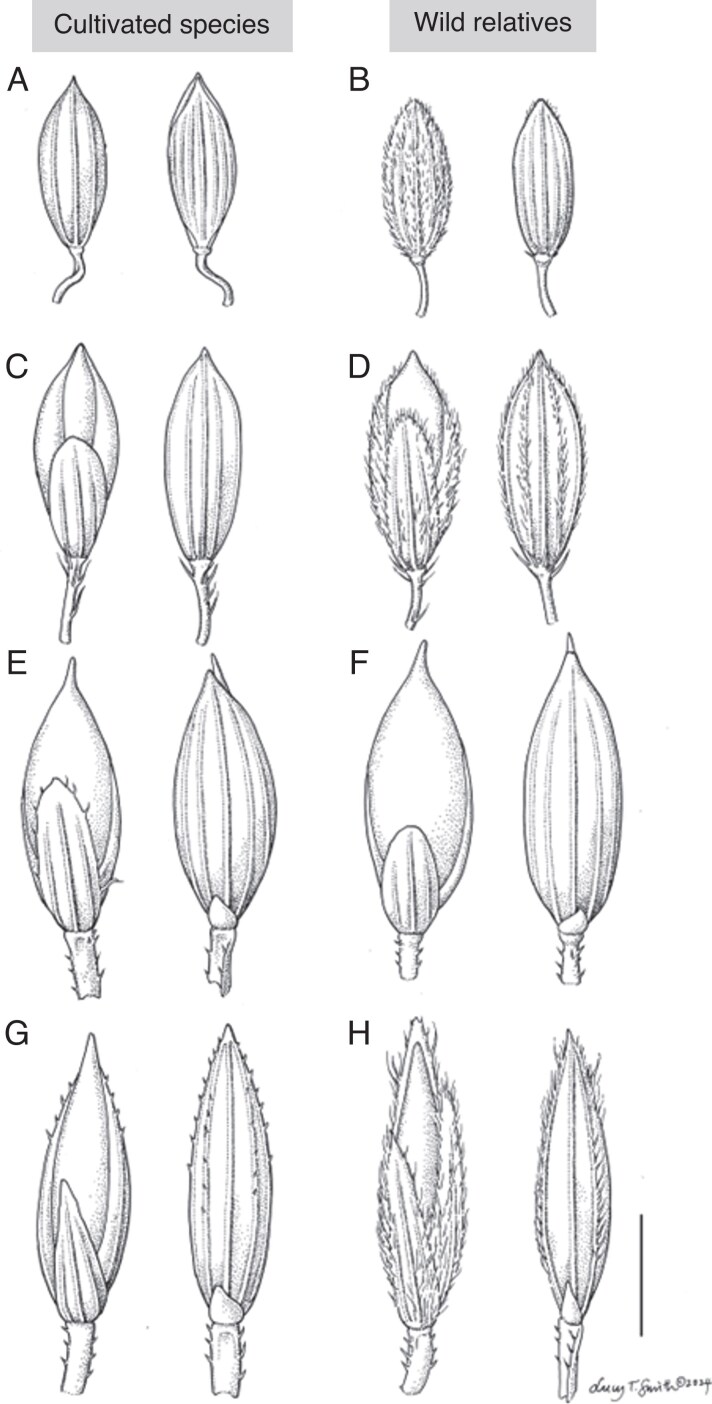 Fig. 6
Fig. 6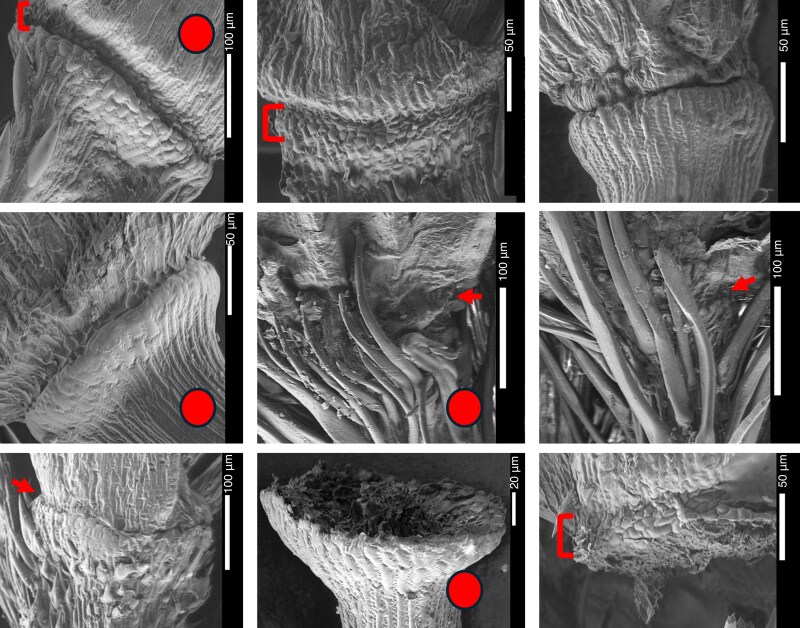 Fig. 7
Fig. 7| Species | Clade | Distribution | Role | Spikelet | Lower glume | Upper glume | Lower lemma | Ploidy | Reference |
|---|---|---|---|---|---|---|---|---|---|
|
| 2 | West Africa | Crop | Ternate | Absent | Full spikelet, glabrous | Glabrous | 4× |
|
|
| 2 | Global | Weed | Ternate | Absent | Full spikelet, trichomes | Trichomes | 2×, 4× |
|
|
| 3 | West Africa | Crop | Ternate | Absent | 1/2 spikelet, glabrous | Glabrous | 4× |
|
|
| 3 | Global | Weed | Ternate | Absent | 1/2–2/3 spikelet, trichomes | Trichomes | 4× |
|
|
| 5 | Asia | Crop | Binate | Small | 1/3 spikelet, trichomes | Glabrous | 2×, 4× |
|
|
| 5 | Asia | Weed | Binate | Small | 1/3 spikelet, trichomes | Glabrous | 2×, 4× |
|
|
| 6 | Global | Weed | Binate | Small | 1/2 spikelet, trichomes | Trichomes | 4×, 6× |
|
|
| 6 | Global | Weed, crop | Binate | Small | 1/3 spikelet, trichomes | Trichomes | 4×, 6× |
|
- —Natural Environment Research Council10.13039/501100000270
- —Christiane Morgan at the
- —Imperial College London10.13039/501100000761
- —Botanic Gardens10.13039/100006347
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Taxonomy and Phylogenetics · Wheat and Barley Genetics and Pathology · Pasture and Agricultural Systems
INTRODUCTION
Cereal crops provide the majority of the world’s food, yet are likely to be disrupted heavily by future and present climate change (FAO 2016; Fatima et al., 2020). Many underutilized or ‘orphan’ crops are likely to be more tolerant to this change, and can provide resilience in vulnerable food systems that will otherwise be degraded (Mabhaudhi et al., 2019; Ulian et al., 2020; Santhoshini et al., 2025). A selection of these crops have been presented as neglected and underutilized species (NUS; Ulian et al., 2020; Animasaun et al., 2023), which have experienced far less research and commercial development than major crops, despite their climate-resilient traits and importance to indigenous identity and rural communities. Many of these are millet grasses, including white (Digitaria exilis) and black (D. iburua) fonio from West Africa. Under-improved crops (including fonio) and crop wild relatives (CWRs) are known to retain allelic diversity which may have disappeared from more heavily domesticated species that experience genetic bottlenecks, and hold the key to preventing diseases, increasing nutrition and providing climate resilience in vulnerable areas (Warburton et al., 2006; Hajjar and Hodgkin, 2007; Dempewolf et al., 2017; Kashyap et al., 2022). The involvement of wild species in crop breeding programmes has been utilized extensively for rice (Stein et al., 2018; Zheng et al., 2024), wheat (Charmet, 2011) and maize (Warburton et al., 2006; Abdoul-Raouf et al., 2017). However, for the improved utilization of orphan millets like fonio to be possible and effective, scientists first require a detailed understanding of the taxonomy, evolutionary history and geographic niche of the study species. Knowledge of how grasses have evolved and adapted to climatic conditions in the past can indicate how and why they might respond to challenging future climates.
Digitaria species uses and systematics
In the genus Digitaria (Panicoideae: Poaceae) there are four crop species that are or have been important to agriculture systems and providing food for people: white fonio (D. exilis), black fonio (D. iburua), raishan (D. compacta) and Polish millet (D. sanguinalis) (Portères, 1955, 1957, 1976; Singh and Arora, 1972; Cruz et al., 2016). The genus includes the cultivated pasture grass D. eriantha (Pangola grass), common in South Africa, the Americas and Southeast Asia (Parsons, 1972; Tikam et al., 2013). Digitaria grasses can also have negative economic impacts on agriculture, as the invasive, spreading habit of some wild species (including wild D. sanguinalis, D. ciliaris and D. humbertii) makes them difficult to manage in farm and horticultural settings (Jones et al., 2021; Touafchia et al., 2023; Randrianarimanana et al., 2024), and Digitaria grasses are challenging to identify to species level. Although there have been several genomic and phylogenetic studies of Digitaria, described below, there has been no integrated approach that considers phylogenetic, morphological and biogeographical analysis of the group through history, or study that includes all four domesticated crop species and close wild relatives. By producing this analysis, we can build knowledge of the close wild relatives of Digitaria crops, to facilitate future researchers in conducting research on this genus.
The genus Digitaria was first described by Haller (1768), and now contains around 250 species globally (Henrard, 1950; Touafchia et al., 2023; POWO, 2025), occupying both temperate and tropical niches. Studies in Digitaria systematics include work by Stapf (1915), Henrard (1950) and Vega et al. (2009) in morphological classifications. Key traits that have been used to identify species include the shape and size of the upper glume, trichomes on spikelet veins, branching patterns of the inflorescence (technically a synflorescence (Kellogg, 2015), but referred to in this study as an inflorescence for ease of terminology), number of racemes, and number of spikelets in a group (binate = two spikelets, ternate = three or more). These characters are illustrated in Supplementary Data Fig. S1.
Studies in molecular phylogenetics have utilized the combination of ITS sequences and morphology (Lo Medico et al., 2017), plastid sequences (Morrone et al., 2012) and a mix of Sanger (ITS, matK, rbcL, ndhF, atpB and trnL-F) and next-generation sequencing (NGS) target-enrichment sequence capture of 353 genes (using the Angiosperms353 bait kit) within a supertree matrix approach (Touafchia et al., 2023), to untangle the complex relationships between Digitaria grasses, though not wide-scale phylogeny using purely NGS data. Regional taxonomic treatments of Digitaria have been conducted in Pakistan and Central Asia, Southeast Asia and Madagascar by Gilani et al. (2003), Boonsuk et al. (2016) and MacKinnon et al. (In Press), respectively. Minoji and Sakai (2024), in the assembly of a chromosome-level genome for the Asian species D. radicosa, also included several of the major clades within a phylogenomic analysis. Throughout these studies several key patterns and traits have remained consistent: the topology includes several major clades distinguished by ternate versus binate spikelet morphology, combined with the presence of spikelets with complex or absent versus simple trichomes (complex trichome morphologies including capitate, verrucose, clavate or hooked, as categorized for Digitaria by Touafchia et al., 2023). The genus contains many polyploid species, including both fonio species, which were found to be likely allopolyploids, which makes genetic study of the group particularly difficult (Abrouk et al., 2020; Minoji and Sakai, 2024). The Digitaria clade has also given rise to other morphologically divergent grass genera, including Anthephora and Chlorocalymma (Touafchia et al., 2023).
Food crops in Digitaria
White fonio (D. exilis) is commonly grown in West Africa, where it is especially important for both subsistence agriculture in rural areas (Portères, 1976; Cruz et al., 2016; Burton et al., 2024), and as an increasingly popular export crop (CBI, 2022; Tridge, 2023). It is recognized for its arid climate tolerance, high nutritional value and socio-economic and cultural importance, but also its small grains (1–2 mm in length) and seed husk, which pose post-harvest processing challenges (Portères, 1976; Cruz et al., 2016; Animasaun et al., 2023). Black fonio (D. iburua) is cultivated in small areas of Benin, Togo and Nigeria. It is far less popular than white fonio, and there are few detailed studies available on this rare crop, though Portères (1955) describes the likely domestication of D. iburua in the Aïr mountains of Niger before being transported south through to Nigeria, and then to northern Benin and Togo. White and black fonio plants are shown in Fig. 1.
White (left) and black (right) fonio (Digitaria exilis and D. iburua), cultivated from seed originating in Guinea and Nigeria, respectively, in the Jodrell Glasshouses, Kew. Photo credit and copyright: George P. Burton.
A third Digitaria food crop, raishan (D. compacta), is a millet cultivated in the Khasi Hills near Shillong (Meghalaya, India), a wet highland environment. Hooker (1897) describes its cultivation in nearby Assam and the Khasi Hills, and Portères reports that in the 1950s it was still under cultivation on ∼100 acres of land in nearby Assam (Portères, 1957). The most detailed published source of information for this crop is a study by Singh and Arora (1972), which includes a theory of how the crop was selected and domesticated from wild D. cruciata, including photographs of the crop’s long, unbranching racemes and providing details of its cultivation in tribal communities. This study makes no mention of cultivation in Assam, and since this study there have been no formal published reports of cultivation in Meghalaya. The name Digitaria cruciata var. esculenta Bor., previously used for the cultivated variety, is now an accepted synonym of D. compacta (POWO, 2025), a species with occurrence overlapping wild D. cruciata across central and south-eastern Asia. A photograph of raishan grown in the Khasi hills is reproduced from Singh and Arora (1972) in Supplementary Data Fig. S2.
Polish millet (D. sanguinalis) is thought to have been cultivated during the 1500–1600 s in Eastern Europe, first intentionally cultivated in a monastery garden in Croatia or Albania, before moving north and expanding into Germany, Hungary and later Poland and Ukraine (Portères, 1955, 1957) before its decline in the mid- to late 1800s. Portères suggests that the first record of this cereal is from 1561, and that it was reported to still be cultivated in Poland in the 1890s; Netolitzky (1914) says that ‘blood millet’ was cultivated in Germany in the 16th century. He mentions no difference between cultivated and wild grains. Henrard (1950) also cites a specimen of the cultivated form of this crop (D. sanguinalis var. esculenta) at the Paris herbarium, which is described to have longer inflorescence racemes and a more robust, erect habit than common D. sanguinalis; however, no specimens or detailed records could be found for this study.
Fonio genetic studies
One of the earliest attempts to investigate the relationship between white and black fonio and their wild relatives using phylogenetic methods was by Hitu et al. (1997), who used random amplified polymorphic DNA (RAPD) markers to support that the phylogenetically closest wild relative of white fonio is D. longiflora and that of black fonio is D. ternata. This relationship was tested and confirmed in later analyses by Adoukonou-Sagbadja et al. (2010), Nyam et al. (2017) and Ngom et al. (2019). Adoukonou-Sagbadja et al. (2010) supports a genetic affinity of >92 % between fonio crops and closest wild relatives, and a similar close ancestry between D. sanguinalis and the common weed D. ciliaris. The production of a chromosome-level annotated genome for D. exilis by Abrouk et al. (2020) also confirmed the relationship between white fonio and D. longiflora, and shared useful insights into synteny shared with other cereal crops.
Domestication of grass species
Domestication of cereal crops involves phenotypic development or transition based on traits that have been selected by farmers and by cultivation practices (Meyer et al., 2012; Fuller et al., 2023). Through this ‘accelerated evolution’, wild plants experience selection through many generations of seed collection and re-sowing (Harlan et al., 1973). Estimates suggest that transition to early stages of domestication can take as long as 2000 years of active cultivation, but crops display different thresholds of domestication phenotypes over time, dependent on plant pollination and reproductive strategies, and introgression, crossing and cultivation methods (Brush et al., 1995; Glémin and Bataillon, 2009; Allaby et al., 2017, 2022).
The reduction of seed shattering is a key phenotypic change of cereal domestication: this is a mechanistic change involving the devolution of an abscission zone in the inflorescence, which prevents the grain from easily separating from the rest of the plant before it has reached full maturity (Yu and Kellogg, 2018; Yu et al., 2020). This increases the chances of being harvested and cultivated, unlike wild plants which benefit from seed dispersal (Woodhouse and Hufford, 2019). Another phenotype is the reduction of trichomes on the spikelet; in wild plants the presence of trichomes allows the seed or spikelet to be dispersed by the wind and by animals, and helps to establish the seed in the ground. Larger seed size is also a common sign of domestication, reflecting improved seed vigour for successful germination, especially for grasses in arid or difficult environments, and selection for higher yield and seed nutritional content (Fuller et al., 2023). By identifying the range of domestication traits shown by a cereal crop like fonio, researchers can strategize routes to trait improvement through breeding and gene editing.
Aims and scope
Improving our understanding of the evolution of Digitaria species will provide valuable knowledge to future cultivation and biodiversity projects. This study aims to identify CRWs within Digitaria beyond D. longiflora and D. ternata, and resolve when, where and how separate crop lineages within Digitaria may have evolved.
We construct a well-resolved phylogeny of the genus using target-enrichment sequencing and maximum likelihood (ML) and coalescent-based tree-building methods, with sampling taking into account geography, and clades containing crop domestications. We record and discuss the development of distinct and shared morphologies across the group, including domestication traits. This provides an integrated history of the genus, involving the spread and growth of wild species as an important part of this crop history.
MATERIALS AND METHODS
Species sampling and material
Ninety-six samples of 46 species were chosen for DNA extraction and sequencing based on the cladistics and monography presented by Henrard (1950) and Stapf (1915), Lo Medico et al. (2017), Vega et al. (2009), and Touafchia et al. (2023). To approach questions about the evolution of crop species, sample selection was targeted on close relationship and geographic affiliation with key crop species Digitaria exilis, D. iburua, D. compacta and D. sanguinalis. Urochloa ruziziensis was chosen as the outgroup taxon, within Paniceae. Details of collections involved in the final trees are included in Supplementary Data Table S1.
Samples were primarily collected from dried herbarium specimens available from the K herbarium, with few others from P herbarium (the herbarium codes used are from Index Herbariorum; Thiers, 2025), and others were extracted from silica-dried material from recent fieldwork expeditions. Leaf and inflorescence material was sampled for DNA extraction. Specimens were identified and selected by comparison against Henrard (1950) and against the K herbarium collections, with a focus on type specimens. Type specimens of D. iburua and D. cruciata var*. esculenta* (to represent D. compacta) were sampled at the K herbarium, from native cultivation regions in Nigeria and India, respectively. For material most likely to represent D. sanguinalis in its cultivation zone in Europe, a germplasm accession was acquired from the Leibniz Seedbank (IPK) in Germany, which was previously stored and grown annually during the summer in the Alter Botanischer Garten, Gottingen, described as a fast-growing invasive and a descendent of the cultivated variety. This material was grown and sequenced as part of the NERC Environmental Omics Facility (NEOF) de novo project NEOF1392 2021, using WGS (included in this study as sample GB999), and will be assembled and published as part of a future ongoing study.
Sixty-four DNA samples and corresponding morphological traits from specimens were recovered and analysed for 46 species: 22 binate, 23 ternate and one with solitary spikelets. Samples were most commonly taken from specimens collected in Nigeria (eight), Madagascar (seven), India (nine) and Ghana (seven). Of all species’ native distributions, 34 occur in Africa, 16 in Asia, 4 in the Americas and 3 in Europe.
DNA extraction and library preparation
DNA was extracted and isolated from samples using an adapted CTAB method (Doyle and Doyle, 1987), using CTAB, isopropanol and chloroform for purification and magnetic beads for cleaning steps. Target capture sequencing was conducted using the Angiosperms353 target capture probe kit (Johnson et al., 2019; Baker et al., 2021). Libraries were prepared using indexes from the NEBNext Ultra II DNA Library Prep Kit, and NEBNext Multiplex Oligos from Illumina. Pooled libraries were hybridized with baits from the Angiosperms353 probe kit for 24 h at 65 °C and 12 cycles of PCR. Samples were sent to Macrogen Inc. in Seoul, South Korea, for sequencing using an Illumina HiSeqX platform, for 2 × 150 bp paired-end reads. TapeStation (Agilent Technologies 4200 TapeStation) High Sensitivity D1000 ScreenTapes, gel electrophoresis and Quantus Fluorometer (Promega, USA) measurements were used for validating DNA integrity and quantity throughout the extraction and library preparation process.
DNA sequence assembly
Raw data reads were trimmed of their adaptors and organized into paired-end and unpaired sequences, and filtered for quality using Trimmomatic 0.39 (Bolger et al., 2014). Locus assembly was completed using HybPiper 2.1.2 (Johnson et al., 2016) using a DNA target fasta file to capture reads from the Angiosperms353 genes and WGS, using BLASTX v2.5.0 (Camacho et al., 2009). The gene coding regions were extracted and distributed into reads per 353 genes, per sample, using HybPiper. Average gene recovery statistics were provided by the hybpiper_stats.py function in HybPiper. Sequences were aligned using MAFFT v7.476 (Katoh and Standley, 2013), then filtered to remove sequences with >30 % missing data within alignments using PhyUtility 2.2.6 (Smith and Dunn, 2008).
Phylogenetic tree-building
Cleaned gene alignments were (1) used to build a concatenated supermatrix using the -t1 flag ML method, with a GTR + G + I model, in IQ-TREE v2.1.2 (Minh et al., 2020), and (2) analysed with IQ-TREE to produce individual ML gene trees based on BIC scores; low support values <0.3 were again collapsed using Newick Utilities v1.6 (Junier and Zdobnov, 2010), taxa with long branches (often due to contamination errors, misalignment or long-branch attraction) removed using TreeShrink v1.3.9 (Mai and Mirarab, 2018), and finally a coalescent species tree was inferred using ASTRAL-III v5.7.7 (Zhang et al., 2018). Bootstrap values for ML trees were calculated using 1000 ultrafast replicates in the IQ-TREE concatenated tree, and LPP and quartet scores (QSs) using the -t2 flag in ASTRAL-III for the coalescent species tree.
Visualizations of trees were produced in the R environment v4.4.0 (R Core Team, 2024), using the ggplot2 (Wickham, 2016), ggtree (Yu et al., 2017), ape (Paradis and Schliep, 2019), phytools (Revell, 2012) and treeio (Wang et al., 2020) packages.
A principal component analysis (PCA) of sequence genetic distance was produced, using AMAS 1.0 (Borowiec, 2016) to concatenate gene alignments from MAFFT, and then a distance matrix of sequences was produced using the dist.alignment function in seqinR (Charif and Lobry, 2007) to calculate a pairwise distance matrix from DNA sequence alignments.
Time-scaled tree calibration and biogeography
Alignments were input to Sortadate (v2020-12-05; Smith et al., 2018), to rank gene clock-likeliness based on tip-to-root variation, bipartition and tree lengths, using the ‘gene-shopping’ method, to avoid computational issues caused by inputting large gene datasets in BEAST analysis. Alignments were concatenated and then split by gene partition using AMAS, to fill missing taxa per alignment. The top 15 ‘best’ genes were input as 15 separate partitions on BEAUti2 (https://www.beast2.org/beauti/).
An uncorrelated lognormal relaxed clock model was used to allow for independent evolutionary rates across taxa. A GTR site model (same as the ML tree) was used, with a γ category of 4. Topology (the starting tree) was constrained to the rooted ML tree. A Yule model prior was set, with U. ruziziensis set as an outgroup using a mean divergence time of 21 mya (Pessoa-Filho et al., 2017; Hackel et al., 2018) and a σ of 2 (σ representing the standard deviation and uncertainty around the mean). The MCMC run was set to 500 000 000 with a burn-in of 50 000 000 (10 %), sampling every 100 000. The.xml control file from BEAUti2 was then run using BEAST2 (Bouckaert, et al., 2014). Two independent chains were run and combined. The log files from these BEAST2 runs were analysed using Tracer (Rambaut et al., 2018), and tree files with an ESS >200 combined using LogCombiner (https://beast.community/logcombiner). A final maximum clade credibility (MCC) tree with mean node heights was produced using TreeAnnotator (https://beast.community/treeannotator).
The MCC tree from BEAST analysis and a set of binarized species distributions set into five main regions (Americas, West Africa, South and East Africa, Europe and Asia) were used with the program BioGeoBEARS (Matzke, 2013) in R to estimate and illustrate ancestral geographic ranges. Native species distribution data were collected from POWO, GBIF and specimens at K herbarium. The models DEC and DIVALIKE (and +J) were used, and the best iterations were determined using LnL values.
Ploidy estimation
Ploidy level for samples was estimated using nQuire (v2018-05-05; Weiß et al., 2018). Sorted BAM files from the HybPiper gene extractions were input to nQuire, and analysed using the lrdmodel function. Delta log-likelihood values were calculated and used to interpret ploidy (either diploid, triploid, or tetraploid and above). Reported ploidy levels for Digitaria species were also accessed from the Chromosome Count Database (http://ccdb.tau.ac.il/), and found in other literature.
Morphological traits
Morphological traits were extracted from Plants of the World Online (POWO, https://powo.science.kew.org/) and GrassBase (Clayton et al., 2016) and verified against specimens and type collections at K herbarium. Spikelet traits were observed using dissection microscopes. Traits collected included growth form (mat-forming, rhizomes or stolons), growth habit (erect, spreading), whether spikelets were binate or ternate, presence and type of trichomes present on spikelets (glabrous, complex or simple trichomes), number of racemes, length of racemes, and length and width of spikelets.
Imaging
Images of whole plants including inflorescences and roots were taken using an Epson 10000XL scanner at 800 dpi and processed using Adobe Photoshop. Spikelets were imaged using a Leica Z-stacking microscope. Digitaria spikelets were drawn to 1.25 mm scale at ×24 magnification by Lucy T. Smith.
SEM was performed using two methods, the first using platinum coating at the Jodrell Laboratory in Kew Gardens and the second using VP-SEM with uncoated spikelets at the Natural History Museum, London. Spikelets were samples from herbarium sheets at K. For SEM the samples were placed on metal stubs and coated with platinum, using a quorum-Q150t ES series sputter-coater. A Hitachi 8230 with a 5 kV electron beam was used to process images. For VP-SEM, uncoated spikelets were placed on metal stubs and observed using a FEI Quanta 650 FEG scanning electron microscope.
RESULTS
DNA sequence recovery
DNA recovery statistics from the 353 target genes resulted in capturing an average of 345 genes per sample. Full recovery statistics from HybPiper are provided in Supplementary Data Table S2. Morphological traits, reported ploidy and clade group numbers of key cultivated and wild species are shown in Table 1 and are mapped across phylogenetic trees in Supplementary Data Figs S3 and S4.
Ploidy estimation
Twenty-eight samples were estimated to be diploid, 22 triploid and 13 tetraploid or above, shown in Supplementary Data Fig. S4. Tetraploid or above samples include D. iburua, D. exilis, D. sanguinalis, D. monodactyla, D. gayana, and D. leptorhachis. Diploid samples include D. abyssinica, D. cruciata, D. compacta, D. ternata, and D. longiflora. Samples of D. ciliaris and D. sanguinalis were estimated as both triploid and tetraploid.
Digitaria species topology
There is similar topology for Digitaria species presented by the ML and coalescent method trees produced by IQ-TREE and ASTRAL-III, respectively, shown in Fig. 2. In both trees, there is a split into two large clades: clades 1–3 with almost exclusively ternate species (D. monodactyla in clade 1 being the only binate species) and clades 4–6 with exclusively binate species. Species inflorescence morphology for each food crop clade is shown in Fig. 3.
Maximum likelihood supermatrix (left) and ASTRAL-III coalescent (right) phylogenetic trees of Digitaria species. Bootstrap scores are displayed on the ML tree, and QSs and pies show primary (black), second (medium grey) and third (light grey) gene tree topology agreement. Species with cultivated food species and close wild relatives are highlighted: fonio species in orange, and raishan and Polish millet species in blue. Clades are numbered 1–6 for discussion: 2–3 are ternate species and 4–6 are binate. Not all species are included within numbered clades due to discrepancy between trees.
Photo scans of Digitaria inflorescences from herbarium specimens, representing the four main cultivated food crops and closely related wild relatives. Species shown are: (A) D. exilis (DP 1001), (B) D. longiflora (R.A. Farrow 112a), (C) D. fuscescens (J.B. Hall 3304), (D) D. argyrotricha (Greenway 10819), (E) D. iburua (DP 1010a), (F) D. ternata (Tuley 1547), (G) D. atrofusca (A. Falannigui 25), (H) D. sanguinalis (Stewart 26651), (I) D. velutina (Hatch 4296), (J) D. ciliaris (Gledhill 200), (K) D. compacta (Hooker & Thomson s.n.), (L) D. cruciata (Stainton, Sykes & Williams 3989) and (M) D. radicosa (Vorontsova 2116). Scale bar = 1 cm; all images are to same scale.
Clade 1: Digitaria monodactyla
Digitaria monodactyla is the first-diverging species in the wider ternate group, though it is itself binate. The location of this primary divergence is the same on both trees with high ASTRAL-III support values >99 % and gene tree congruence for first topology.
Clades 2 and 3: white and black fonio and relatives
Within clades 2 and 3 the two different fonio crop species emerge into separate well-supported clades in both trees: clade 2 with white fonio (D. exilis) and relatives D. longiflora, D. argyrotricha and D. fuscescens (this sister clade supported by 100 % ML bootstrap and QS), and clade 3 with black fonio (D. iburua) and relatives D. ternata, D. atrofusca and D. madagascariensis (sister clade supported at 94 % ML and 100 % QS). On the ASTRAL-III tree, low gene tree congruence in clade 2 separates multiple samples of D. longiflora and D. exilis (58 % ML, 66 % QS). One D. longiflora sample from Tanzania (GB27) is observed to be outside the clade containing West African fonio and other D. longiflora samples (48 % ML, 100 % QS). Support for the separation between D. iburua and D. ternata in clade 3 is strong at 100 % ML, and 88 % QS.
Clade 4: Digitaria leptorhachis, D. debilis and D. humbertii
A clade with D. leptorhachis, D. debilis and D. humbertii contains the first-diverging species among the binate species (not including D. monodactyla). This is congruent in both trees, with 100 % ML and QS support, and especially strong gene tree agreement in the ASTRAL tree for the relationship between D. debilis and D*. leptorhachis*.
Clade 5: raishan and relatives
Both trees place the Indian crop raishan (D. compacta) in a well-resolved clade with relatives D. cruciata, D. setigera and D. radicosa, with identical topology between trees and high ASTRAL-III support for the difference between D. cruciata samples and relatives D. setigera and D. radicosa (100 %) and for the whole clade (90 %). This is similarly high in the ML tree, with 94–100 % support values for the whole clade and within.
Clade 6: Polish millet, D. ciliaris and relatives
The placement of D. sanguinalis samples is poorly supported in the ML tree (57–71 %), but more confidently in ASTRAL-III (84–94 %), where two samples from India (GB68) and Germany (GB999) appear more closely related to D. nuda and D. velutina, respectively.
The most discordant relationship between trees is the placement of species D. abyssinica, D. eriantha and D. ciliaris. While the ML tree presents clade 5 and D. ciliaris as well-supported sister groups (99 %), with samples of D. ciliaris from India, Brazil and Africa forming a single clade in the ML tree (94 %), the ASTRAL-III tree splits the D. ciliaris samples with low support (20–62 %), placing them between the D. eriantha and D. sanguinalis clades. Digitaria eriantha itself is well supported as a clade with D. milanjiana at 99–100 % ML, and 94–100 % QS, and appears to be closely related to D. ciliaris in both trees. A clade of D. abyssinica contains samples from across Africa and South America, all supported in the same clade together in both trees (99–100 %); however, it appears to be more closely related to D. sanguinalis in the ML tree (99 %), while its placement outside of both D. compacta and D. sanguinalis clades in the ASTRAL-III tree is similarly well supported (98 %).
Genetic distance matrix
A PCA of genetic distance between concatenated sequence alignments is shown in Fig. 4; the species D. monodactyla, D. debilis, D. humbertii and D. leptorhachis appear to be the closest to the outgroup U. ruziziensis, and the most separate and distant from either major binate or ternate clades. These distinct divergences are well supported in both ML and ASTRAL-III trees, which share the same main topologies in gene congruence.
PCA plot of genetic distance between Digitaria sample alignments. Colours correspond to crop clades shown in Fig. 2; key species are indicated in the labels, including early-diverging Digitaria ternate species D. monodactyla, and the binate group is circled. Sample SRR8417290 is the outgroup species Urochloa ruziziensis.
Evolution and biogeography
The results of the BEAST2 Bayesian analysis are shown in Fig. 5. The earliest-diverging extant Digitaria are African natives D. monodactyla, D. compressa and D. gayana from mostly ternate clades 1–3, and D. debilis, D. leptorhachis and D. humbertii in binate clade 4. Both D. exilis and D. iburua are estimated to have diverged from wild relatives around 2–3 mya. Digitaria sanguinalis diverged from D. velutina (north Africa) around 2.7 mya, though an earlier divergence into India is dated at 6.22 mya. The specimen of D. compacta, cultivated in India, diverged from wild D. cruciata more recently at 1.33 mya, and both species from the clade containing D. radicosa and D. setigera around 6 mya.
Bayesian (BEAST2) time-calibrated phylogeny of Digitaria species and likely dispersal routes. (A) Maximum clade credibility (MCC) tree, where clades are coloured by dispersals to Madagascar (orange) and the Americas (pink). Cultivated food species and relatives are highlighted in grey. (B) Likely routes of dispersal, corresponding to clades numbered in the MCC tree. Maps are based on overlapping native extant species distributions, from POWO (https://powo.science.kew.org/).
Biogeographical analysis by BioGeoBEARS is shown in Supplementary Data Fig. S5, which supports an ancestor of Digitaria in south-eastern Africa (DEC model LnL = −167.38, DEC + J LnL = −162.34). This corroborates the BEAST results, where the earliest-diverging Digitaria (including D. monodactyla, D. gayana, D. compressa, D. leptorhachis and D. debilis) all have native distributions that overlap in south and eastern Africa.
Domestication
Illustrations of Digitaria crop and wild species spikelets are shown in Fig. 6. There appears to be a loss of trichomes for cultivated fonio species, though it is not fully complete. Digitaria longiflora has trichomes present on the spikelet and pedicel that are not present on D. exilis. Trichomes are present on the spikelet and pedicel of D. ternata, and on D. iburua there are no trichomes on the spikelet but they are still present on the pedicel. Trichomes are present on the pedicel of both D. compacta and D. cruciata, and only slightly discernible on some specimens on the spikelet. On D. sanguinalis complex trichomes are very noticeable across the whole spikelet and pedicel. Close relatives D. ciliaris and D. velutina (not pictured) are similarly pubescent.
Illustrations of Digitaria spikelets, representing the four main cultivated food crops and CWRs. (A) D. exilis (Clarke s.n), (B) D. longiflora, (C) D. iburua (Lamb 54), (D) D. ternata (Schimper 76), (E) D. compacta (Hooker & Thomson s.n.), (F) D. cruciata (Royle s.n), (G) D. sanguinalis (Stewart 26651), (H) D. ciliaris (Lindeman & Haas 459). Drawn by Lucy Smith at Kew Gardens. Scale bar = 1.25 mm at ×24 magnification.
Regarding the size of the spikelet, from average measurements provided by GrassBase (Clayton et al., 2016), D. exilis has an average spikelet length of 1.7–2 mm versus 1.2–2 mm for D. longiflora, and D. iburua has ∼2 mm versus 1.8–2.7 mm in D. ternata. Although there are very few samples of cultivated D. compacta available, the average spikelet length is recorded as ∼ 2 mm versus 2–3.5 mm in D. cruciata and 2.5–3 mm in D. radicosa. Digitaria sanguinalis has an average of 2.5–3.5 mm, which is smaller than 1.5–2.1 mm in D. velutina, but similar to 2.5–4 mm for D. ciliaris. Visually, the crop species do not appear any larger than wild species; thorough future morphometric studies should be conducted on a wide range of material to confirm this.
The abscission zones for Digitaria species are shown in Fig. 7 (cultivated and CWRs) and Supplementary Data Fig. S6 (wild species). The abscission zone is evident below the whole spikelet, shown by comparison of the complete spikelet and removal, where an empty pedicel is shown. On the abscission zone itself, a layer of specialized cell structures appears to be arranged horizontally in squares, perpendicular to the vertical, long striations in the pedicel likely representing lignification patterns. For small millets, looking at cell arrangements is a good alternative for smooth versus rough appearances in archaeological studies. The same abscission zone cell patterns are seen in both wild D. cruciata and cultivated D. compacta, as is the ‘clean’ line pattern in both D. longiflora and D. exilis, and in D. iburua and D. ternata. The ‘horizontal cell’ pattern is only observed for binate species in D. sanguinalis, though this may be due to inconsistency of age between specimens. There appears to be no easily noticeable or consistent phenotypic difference between the wild and cultivated species. In Supplementary Data Fig. S6 an array of wild Digitaria species is shown for comparison, presenting the concave and convex sides of the pedicel base and abscission zone layer, which follows the same patterns as seen in cultivated and CWR species.
Scanning electron microscope photos of Digitaria abscission zones in cultivated and close wild relatives, captured at Kew Gardens and the Natural History Museum (UK). Red circles indicate cultivated species. Arrows and brackets indicate the abscission zone area. Species from top left to bottom right are D. compacta (Hooker & Thompson s.n.), D. cruciata (Royle s.n), D. longiflora (Merklinger 65), D. exilis (Burton 15), D. iburua (Lamb 54, type), D. ternata (Hall 636), D. sanguinalis (Santos 1844), D. exilis (Thomas 1485) and D. sanguinalis (Peterson 23883).
DISCUSSION
Digitaria species topology
The structure of the nuclear trees and general clade topologies concur with those of the trees presented by Touafchia et al. (2023) and Minoji and Sakai (2024). The topology is also supportive of the systematics proposed by the morphometrics of Vega et al. (2009) and the Sanger phylogenies of Lo Medico et al. (2017) and Morrone et al. (2012). However, these trees are the first presentation of Digitaria using a high number of NGS low-copy target nuclear genes, a wide range of species sampling both geographically and across all major clades, and including all four known species domesticated for food cultivation, and their close wild relatives. Touafchia et al. (2023), for example, only used 17 Digitaria NGS sequences, and did not include white or black fonio or raishan samples in their phylogeny.
Clade 2: white fonio and relatives
The closest relative of D. exilis is D. longiflora, following previous studies including Adoukonou-Sagbadja et al. (2010) and Abrouk et al. (2020). A sample of D. longiflora from Tanzania, the only one sampled from outside of West Africa, is also seen to be less closely related to D. exilis in both trees, supporting that D. exilis diverged from D. longiflora in West Africa, and is more closely related to populations there. The two species are similar in morphology and often grown in close proximity in cultivated fields across West Africa (Burton et al., 2024), though D. exilis has lost its trichomes, has increased raceme number and length, and has abandoned the rhizomatous growth habit.
The next set of closely related species are D. fuscescens and D. argyrotricha, the former of which is native to West Africa, and the latter to the coast of East Africa. All four species have maintained a strong v-shaped finger inflorescence shape with no or rare auxiliary branching (Fig. 2). Digitaria exilis is the only species that commonly has more than two racemes, though this varies between landraces, and the racemes are usually still connected at a single node. The morphology of D. fuscescens is similar to that of D. longiflora in the short racemes, and has similar ‘weedy’ invasive traits, such as a spreading habit, stoloniferous roots and small inflorescences. Digitaria argyrotricha has longer racemes with larger spikelets, and these are covered with a dense coat of trichomes. Digitaria gayana is the earliest-divergent ancestor in this lineage, an African native that varies between a v-shaped and a sometimes more branched inflorescence, and spikelets also with dense trichomes.
White fonio has not become completely indistinguishable from its close relatives; its racemes are longer, but it still experiences lodging and shattering. Its lineage in invasive and drought-tolerant African grasses is likely to offer unexploited potential, though it otherwise suffers from noticeably incomplete domestication.
Samples of the two fonio crops D. exilis and D. iburua are estimated to be tetraploid (2n = 36), corroborating the previous study by Adoukonou-Sagbadja et al. (2007). Close relatives D. longiflora and D. ternata were estimated to be diploid, contrary to previous studies that suggest both are tetraploid (Adoukonou-Sagbadja et al., 2007; Abrouk et al., 2020). Only one diploid accession of D. longiflora (2n = 18) has been reported, by Hoshino and Davidse (1988). Close relative D. fuscescens is estimated here as tetraploid, and has been reported as both diploid and tetraploid in Gould and Soderstrom (1967, 1974).
Clade 3: black fonio and relatives
The close relatedness of D. iburua and D. ternata has been reported since Hitu et al. (1997), and is supported again here. A sister clade containing the common African weed D. atrofusca is also presented. All three species share a common morphology: multiple secondary branches from the main rachis and black upper florets, also shared with their earliest-diverging species in the clade, African D. compressa. Between wild species and D. iburua we observe the development of thick culms and robust racemes, and the loss of spikelet trichomes. Digitaria ternata and D. atrofusca overlap with D. iburua in its restricted cultivation area in Nigeria, and also both occur across West and south-eastern Africa. It is likely from this, their shared morphologies and phylogenetic similarity that there were several speciation events within this cluster of species in West Africa as early as 8 mya.
The documentation of black fonio cultivation and subsequent research on its traits and genetic history is particularly scarce, and recent research aimed at improving knowledge about D. iburua often rely on germplasm provided by seed banks in the USA, Togo, Benin and Nigeria. However, due to low germplasm quantities stored at these seed banks, very limited germplasm could be obtained for this and other related studies, from a single accession collected in the 1960s. The authors were fortunately able to collaborate with Dr Abubakar Bello and Ishaq Muawiyya from Umaru Musa Yar'adua University, Nigeria, and early surveys from their work confirm that black fonio is still being cultivated in the Jos Plateau region. The scarcity of available germplasm material for black fonio is concerning, and more collections should be made to enable future research and conservation. Future research with this crop will benefit from including material not only from D. ternata but also D. atrofusca in their studies. Although D. iburua also suffers from apparent incomplete domestication in its small shattering grain, its robust growth habit and multiple racemes make it maybe even more attractive than D. exilis for breeding programmes, if it can be conserved, and promoted alongside improved access to dehusking and processing machinery.
Clade 5: raishan and relatives
Digitaria compacta is confirmed to be closely related to the wild D. cruciata, with a sister clade including D. radicosa and D. setigera, all Asian grasses. Morphology-based cladistics of Digitaria from Pakistan and Central Asia in a study by Gilani et al. (2003) corroborate these findings, predicting a close relationship between D. cruciata and D. sanguinalis, followed by D. ciliaris, D. setigera and D. radicosa. There is a stark development from D. cruciata to the cultivated D. compacta: though they have the same inflorescence structure with opposite secondary branchings from the same rachis node (in a star shape), the cultivated form has long racemes with more grains, and a robust root-to-culm growth form. The inflorescence structure of the sister clade which includes D. setigera and D. radicosa are also similar in their parallel inflorescence branchings. Although all three species are common from Afghanistan eastwards, D. radicosa and D. setigera also occur in Madagascar and other areas of south-eastern Africa, across which dispersal events may have taken place.
The recently published chromosome-level analysis of relative D. radicosa in Minoji and Sakai (2024) included an analysis of synteny between D. radicosa and other crops, finding strongly similar syntenic blocks between D. radicosa and D. exilis, and with rice (Oryza sativa), including a diverse range of NLR (resistance gene analogue RGA5) genes to protect from blast fungi. Digitaria radicosa is likely to be a useful CWR species in future research.
Both D. cruciata and D. compacta are estimated to be diploid in this study, and have been previously reported as diploid and tetraploid (Mehra, 1982; Löve, 1985; Kumar and Subramaniam, 1987; Koul and Gohil, 1991). Close relative D. radicosa is estimated to be tetraploid in this study, but was found to be exclusively diploid in a chromosome-level genome assembly and survey by Minoji and Sakai (2024) .
Raishan has been far more neglected in terms of academic study, commercialization and policy initiatives than the African fonio millets. As part of this study, the authors were able to contact and collaborate with researchers at the Botanical Study of India, and with Dr Meera Das and Fullmerries Puwein at the North-Eastern Hill University, in the Shillong region, Meghalaya, India. Communication with rural farmers in the Khasi Hills has confirmed that the crop was still being cultivated in early 2024, though it is rare and under threat, and being replaced by varieties of higher-yielding finger millet (Eleusine coracana). Raishan has a host of traits that may be beneficial and interesting for further study, including its large grains and long, numerous racemes, and may provide useful information to breeding programmes to benefit food security in both West Africa and India, if international effort is made to protect this species. Recent Indian government initiatives, including the UN’s International Year of Millets 2023, made numerous references to raishan in advertising material and policy documents, but there has so far been little effort to accelerate research or develop initiatives for its conservation and promotion. Seed-banking led by local indigenous people would be a particularly effective method for its protection and conservation.
Clade 6: Polish millet, D. ciliaris and relatives
There is noticeable discrepancy in placement of D. ciliaris and relatedness to D. sanguinalis and D. compacta. Henrard (1950) makes observations about the blurred and overlapping suite of characters for these species, and their overlapping occurrences throughout Asia, Europe and northern Africa. Previous study of these species has supported two discrete, separate species as D. sanguinalis and D. ciliaris, but with closely shared ancestry (Adoukonou-Sagbadja et al., 2010).
Digitaria sanguinalis is estimated as both triploid and tetraploid in this study, corroborating other literature (Šmarda et al., 2014), and also reported as hexaploid and above elsewhere (2n = 54; Xue et al., 1992; Bennett et al., 1998). Digitaria ciliaris is similarly estimated to be triploid and tetraploid here, and as tetraploid and hexaploid in the literature (Carnahan and Hill, 1961; Trouin, 1972), and so both are polyploid species.
The closely related species D. sanguinalis, D. ciliaris, D. nuda and D. horizontalis are known to frequently occupy fields together (da Silva et al., 2024), and so there may be ongoing or historical hybridization. It may be useful to consider clade 6 containing these species in the ASTRAL-III tree as a broad taxonomic complex. Somewhere in this complex, a landrace precursor in populations selected as Polish millet would have been foraged and cultivated in Europe, likely a hybrid between several of the species in this clade. This contrasts with the well-resolved and distinct lineages of the other Digitaria crops. As D. sanguinalis is no longer cultivated as a food crop in Europe, it is difficult to untangle this complex.
Polyploidy
Inconsistencies between chromosome count studies in the literature and some of the ploidy estimations in this study (including around D. exilis wild relatives and D. radicosa) show how challenging it can be to conduct phylogenetic studies of polyploid species using NGS target-capture sequencing data (Griffin et al., 2011; Dufresne et al., 2014; Kyriakidou et al., 2018). The accuracy of estimations made by nQuire analysis has also been questioned (T. Sakai, Japan, Kyoto University, pers. comm.). There is certainly scope for future research to reassess the complex interactions of Digitaria species topologies, crop domestication and polyploidy in panicoid grasses, using recently developed methods such as that of Masters et al. (2024) for Brachiaria grasses using allele phasing, and phylogenetic network analysis using tools including PhyloNetworks to detect possible reticulation and hybridization events (Solís-Lemus and Ané, 2016; Solís-Lemus et al., 2017).
Evolution and biogeography
From species distributions and BEAST node age estimation it is predicted that early groups of Digitaria diverged at least 15–17 mya in central or south-eastern Africa, during the Miocene period. This corroborates with estimated divergences of Anthephorinae (the subtribe within which Digitaria belongs) dated to a mean age of 18.43 mya by Gallaher et al. (2022), with a likelihood of ∼21 % having originated in the Afrotropics region. The origin of the tribe Paniceae is estimated to 29.96 mya with a 38 % likelihood of origin in the Afrotropics, in the same study. Hackel et al. (2018) estimates the divergence of Paniceae at around 22 mya, and the split between binate and ternate Digitaria at around 15 mya (similar to 15–17 mya in this study). Africa has generally been suggested as an area of early divergences for many grass lineages (Gallaher et al., 2022; Peterson and Soreng, 2022), especially C4 grasses, as well as for Poaceae as a whole (Bouchenak-Khelladi et al., 2010), which corroborates our results here.
In the phylogenetic trees, the PCA the and biogeographic and time analyses, it appears that D. monodactyla, D. debilis, D. leptorhachis, D. humbertii and D. compressa are the earliest-diverging and most genetically dissimilar to other later-diverging Digitaria, and are all African native species. Digitaria monodactyla is one of the only binate species within the ternate group, and has uncommonly single, spicate racemes. Digitaria monodactyla and D. compressa also have fire-adaptive culm crowns, adapted to fire-regime lifestyles in seasonal savannah open grasslands in south-eastern Africa (see photographs of growth forms in Supplementary Data Fig. S7). The mid- to late Miocene saw aridification of parts of south-eastern Africa that shaped the evolution of plant species (Osborne, 2008; Pokorny et al., 2015), and in the later period allowed the expansion of savannah and grasslands (Sepulchre et al., 2006). The formation of these grasslands saw the dominance of C4 grassland species in these drier climates between 20 and 10 mya (Edwards et al., 2010; Peppe et al., 2023), which is thought to have coincided with ‘enhanced fire activity’ (Hoetzel et al., 2013; Karp et al., 2018). This expansion of C4 grasses, an increasingly arid climate and fire activity would have both shaped and supported the diversification of Digitaria grasses, including ancestors of the early-diverging extant species D. monodactyla and D. compressa, and the spread of both the ternate fonio lineage to the west and the binate raishan and Polish millet lineages north to Europe and Asia.
In other phylogenetic studies including Digitaria, several species included in other genera are embedded exclusively within the Digitaria binate clade, and not among the ternate species clades (Hackel et al., 2018; Touafchia et al., 2023; Grass Phylogeny Working Group III, 2025), including Chaetopoa taylorii, Chlorocalymma cryptacanthum, Taeniorhachis repens, Anthephora pubescens, A. cristata, Tarigidia aequiglumis, Stereochlaena caespitosa and Baptorhachis foliacea. These species are native to dryland African grasslands (Bouchenak-Khelladi et al., 2010; Grass Phylogeny Working Group III, 2025; POWO, 2025). Chaetopoa taylorii, C. cryptacanthum, B foliacea and A. pubescens also all have single racemes (similar to the spicate inflorescence of D. monodactyla). These species provide an ‘African grassland’ core to the otherwise quite widely distributed binate species, and possibly represent a closer relationship to the earlier-diverging Digitaria in clades 1 and 4 from south-eastern Africa, with close morphological similarity and ecological habitat.
From the few native species sequenced as part of this study, Digitaria appears to have migrated to Madagascar from as late as 10–15 mya, with the separation of D. humbertii (endemic to Madagascar) from D. debilis and D. leptorhachis (occurring in mainland Africa, and D. debilis also occurring in Madagascar). Digitaria humbertii is a grazing-adapted grassland species, with rhizomes, spreading and mat-forming; its sister species D. debilis and D. leptorhachis occupy a similar ecological niche, but are more erect in their growth habit. In a study by Randrianarimanana et al. (2024), ethnobotanic interviews with farmers across three regions in central and eastern Madagascar cited D. humbertii as the most troublesome grass weed in agricultural settings, followed by D. longiflora, which also has a similar growth habit to D. humbertii. Other Digitaria species, including D. ternata, D. ciliaris, D. sanguinalis and D. debilis, were also reported. This ‘weedy’ habit is seen in early- and late-diverging species on mainland Africa and Madagascar, and this phenotype was shown by Touafchia et al. (2023) to not be specific to any clade across the genus, and to have instead evolved convergently many times. Hackel et al. (2018), in investigating the origin and diversity of C3 and C4 grass evolution in Madagascar, comments that endemic C4 panicoid grasses are likely to have appeared during the Miocene, supporting the formation of the D. humbertii–D. debilis clade during this period.
Fonio domestication
Despite a long history of evolution, Digitaria crops do not show significant developments that make them overly distinct from their wild relatives: there is no distinct abscission zone change to suggest a strongly evolved non-shattering phenotype, the grain sizes overlap, and there is only complete loss of trichomes in D. exilis (the most commonly cultivated species). In a study of 203 global food crops by Meyer et al. (2012), the term semi-domesticated’ is used to describe crops that have achieved only partial differentiation from wild species, and seems relevant to apply to Digitaria crops.
The only study to consider fonio abscission zones (a key mechanism for regulating shattering) in depth is Patterson et al. (2016), where an SEM figure of just D. exilis is shown. It shows that the abscission zone for Digitaria can be found at the base of the spikelet, similar to other closely related millet crops. This can be seen in our results, with both the shattered and complete spikelets. Abscission zones are known to form differently across the grasses, with differential cell size and lignification being two strategies for formation, regulated by genes including sh1, sh4, qSH3 and qShi1 (Yu and Kellogg, 2018; Yu et al., 2020; Mascher et al., 2024). Differential cell shapes appear to be important to abscission zone formation in Digitaria, though there is no obvious difference in formation between wild and cultivated species. In the assembly of the fonio genome (with surveys for domestication signatures), Abrouk et al. (2020) find that only 37 % of accessions of fonio across its whole cultivated range have a deletion in the sh1 gene, and that plants with the deletion only show a 7 % chance of reduced shattering. Although this deletion has not been identified in the wild relative D. longiflora, this is a low expression of non-shattering phenotypes even if the gene is under selection.
In a survey of major plant domestication pathways (Fuller et al., 2023), the domestication centre of white fonio is placed in the Inner Niger Basin, Mali, with African rice, pearl millet and cowpea, but only fonio is described as having been domesticated through a ‘segetal pathway’ versus a ‘grain pathway’. Segetal is defined as a ‘former weedy species that grew in agricultural contexts that were added to the crop repertoire’. These segetal domesticates, which grew as weeds and were adopted into mainstream cultivation through convenience, were thought to be domesticated much later, and more quickly than others, likely around 2000–3000 years ago. Other ‘segetal pathway’ crops, including rye and oats, probably grew as weeds in fields of wheat and barley before selection. For fonio, a suggestion is that these harbour crops would have been African rice and pearl millet in the Mali region. Pearl millet was domesticated much earlier in Mali, and was very popular until African rice, fonio and sorghum were later incorporated into local food selection (Champion et al., 2021). This explains why fonio may have poorly developed domestication traits. A genetic resources and breeding programme study of fonio in Nigeria and Benin (Kanlindogbe et al., 2020) agrees that across West Africa there has been little conscious, targeted selection of fonio cultivars, describing cultivated varieties as ‘ecotypes’. In Burton et al. (2024), farmers interviewed in Guinea explained that landraces were only selected for suitability to local microclimates, which changed in rotations every few years, and not for specific uses, suggesting a low domestication pressure for morphological traits, and explaining why traits like shattering have not been significantly improved. Another factor may be the co-existence of D. exilis and D. longiflora often in the same fields, noted in Guinea, with overlapping occurrence ranges across West Africa (Abrouk et al., 2020; POWO, 2025). Though low (around 2 %; Barnaud et al., 2017) outcrossing in D. exilis plants has been observed, introgressions between crop and wild species are still likely, effectively slowing the rate of domestication pressure. This adds weight to the theory that cereal domestication can be a long, slow, ongoing landscape-level process, with no single epicentre for some species (Allaby et al., 2017, 2022).
Conclusions
This study presents a comprehensive, widely sampled phylogeny of the grass genus Digitaria, including all four lineages domesticated as food crops (and their wild relatives) throughout history. Tree topologies are largely congruent with cladistics presented by previous studies, and three domesticated crops (white and black fonio and raishan) appear as distinct, monophyletic lineages, while the European Polish millet appears to be part of a more complicated taxonomic complex. Type specimens of the rare crops Indian-endemic raishan and West African black fonio are sequenced and presented here in phylogenetic analyses for the first time. The closest wild relatives to raishan are found to be wild Asian native species D. cruciata, D. radicosa and D. setigera. New close relatives for both fonio species are identified in D. atrofusca (for black fonio) and D. fuscescens and D. argyrotricha (for white fonio), all African native species. We also confirm through communication with specialists in India and Nigeria that D. iburua and D. compacta were still cultivated in 2025.
As the degradation of African grasslands, including large areas of millet cultivation, accelerates due to human activity and climate change, causing erratic weather patterns and increased fire activity, it will be vital to invest in crops that are resilient to fire and extreme climates (Osborne et al., 2018; Joseph et al., 2024; Knowles et al., 2025). Digitaria species, not only white and black fonio, have the evolutionary history and genetics likely to hold potential for improving food security for vulnerable areas in the future.
Future priorities for research should include collections of fresh and dried plant and seed material for both raishan and black fonio with ethnobotanic data, as seed-banking and conservation of traditional crops and knowledge alongside indigenous peoples are vital tools that will help to prevent the ongoing loss of these rare traditional crops.
Supplementary Material
mcaf212_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abdoul-Raouf SM, Ju Q, Jianyu M, Zhizhai L. 2017. Utilization of wild relatives for maize (Zea mays L.) improvement. African Journal of Plant Science 11: 105–113. doi:10.5897/AJPS 2017.1521 · doi ↗
- 2Abrouk M, Ahmed HI, Cubry P, et al 2020. Fonio millet genome unlocks African orphan crop diversity for agriculture in a changing climate. Nature Communications 11: 4488. doi:10.1038/s 41467-020-18329-4PMC 747961932901040 · doi ↗ · pubmed ↗
- 3Adoukonou-Sagbadja H, Wagner C, Dansi A, et al 2007. Genetic diversity and population differentiation of traditional fonio millet (Digitaria spp.) landraces from different agro-ecological zones of West Africa. Theoretical and Applied Genetics 115: 917–931. doi:10.1007/s 00122-007-0618-x 17726599 · doi ↗ · pubmed ↗
- 4Adoukonou-Sagbadja H, Wagner C, Ordon F, Friedt W. 2010. Reproductive system and molecular phylogenetic relationships of fonio millets (Digitaria spp., Poaceae) with some polyploid wild relatives. Tropical Plant Biology 3: 240–251. doi:10.1007/s 12042-010-9063-0 · doi ↗
- 5Allaby RG, Stevens CJ, Kistler L, Fuller DQ. 2022. Emerging evidence of plant domestication as a landscape-level process. Trends in Ecology & Evolution 37: 268–279. doi:10.1016/j.tree.2021.11.00234863580 · doi ↗ · pubmed ↗
- 6Allaby RG, Stevens C, Lucas L, Maeda O, Fuller DQ. 2017. Geographic mosaics and changing rates of cereal domestication. Philosophical Transactions of the Royal Society B 372: 20160429. doi:10.1098/rstb.2016.0429 PMC 566581629061901 · doi ↗ · pubmed ↗
- 7Animasaun DA, Adedibu PA, Olawepo GK, Oyedeji S. 2023. Fonio millets: an underutilized crop with potential as a future smart cereal. In: Farooq M, Siddique KHM. eds. Neglected and Underutilized Crops. Elsevier, 201–219.
- 8Baker WJ, Dodsworth S, Forest F, et al 2021. Exploring Angiosperms 353: an open, community toolkit for collaborative phylogenomic research on flowering plants. American Journal of Botany 108: 1059–1065. doi:10.1002/ajb 2.170334293179 · doi ↗ · pubmed ↗