Meristems clarify fuzzy floral boundaries: a commentary on ‘Are capitula inflorescences? A reassessment based on flower-like meristem identity and ray flower development’
Paula J Rudall, Richard M Bateman

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
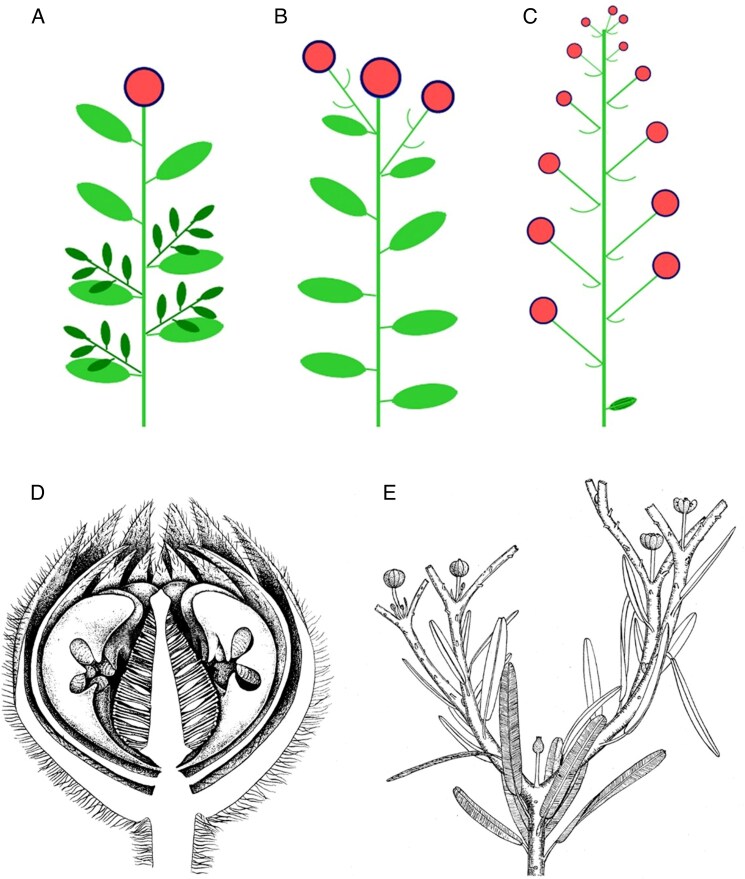 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and animal studies · Flowering Plant Growth and Cultivation · Plant and Fungal Species Descriptions
Some of the best botanical science emerges from studies that address a specific question through observations of a particular well-chosen study group but that ultimately yield results with implications that extend well beyond the original hypothesis. Such is the case with the paper published in this issue of Annals of Botany by Claßen-Bockhoff et al. (2025), entitled ‘Are capitula inflorescences?’, where the authors employ comparative ontogenetic data to document the developmental trajectories of several representative members of the daisy family (Asteraceae). When placed in a developmental–genetic context, the data allow the authors to explore the possible evolutionary origins of the daisy’s characteristic flower-like capitulum, which is composed of numerous, tightly packed flowers that are typically dimorphic: petaloid bilaterally symmetrical ‘ray flowers’ surround non-petaloid radially symmetrical ‘disc-flowers’.
To the eyes of many observers, flowers and inflorescences are strictly non-homologous structures. Capitula are widely regarded as classic examples of pseudanthia, which traditionally have been defined as structures that resemble flowers but are actually judged homologous to inflorescences. However, the pivotal insight achieved by Claßen-Bockhoff and co-workers lies in recognizing the crucial presence and role of a novel category of reproductive meristem, the floral unit meristem (FUM sensu Claßen-Bockhoff and Bull-Hereñu, 2013), which effectively expands the evolutionary potential of reproductive developmental processes. Claßen-Bockhoff et al. (2025) elaborate their concept of three types of reproductive meristem, one of them indeterminate (IMs), the other two determinate (FUMs and flower meristems: FMs). They infer that the daisy capitulum develops from an FUM, which itself evolved through a single developmental shift from an indeterminate to a determinate meristem. Crucially, in their interpretation, this solitary shift replaces the several incremental steps required by traditional hypotheses of pseudanthial origin, as well as necessitating a redefinition of pseudanthia.
Classical morphology distinguishes between determinate (‘closed’) and indeterminate (‘open’) inflorescences (Weberling, 1992). In a determinate inflorescence such as a solitary terminal flower (Fig. 1A) or a cyme (Fig. 1B), all of the axes terminate in a flower. In contrast, in an indeterminate inflorescence such as a raceme (Fig. 1C), the meristem forms nodes and internodes by acropetal extension growth; it generates flowers by lateral segregation until it becomes exhausted, never forming a terminal flower. These two developmental trajectories encompass a strikingly diverse spectrum of inflorescence architectures, including panicles and thyrses. In addition to external morphology, the internal cellular arrangements of the apical zones also differ. Those of indeterminate meristems resemble vegetative meristems, focused on a ‘central mother-cell zone’ of self-renewing stem cells that confer indeterminacy. Determinate meristems (including FMs and FUMs) lack a pluripotent central zone and are determinate from inception. Determinate inflorescence meristems develop by expansion and fractionation; both FUMs and FMs possess a flattened or conical receptacle. FUMs resemble FMs except that they can fractionate into secondary FUMs or FMs. Thus, in this concept, daisy capitula equate with FUMs, raising the argument that FUMs could legitimately be equated with pseudanthia per se.
Can the FUM concept be extended to examine the origin and early evolution of inflorescences and flowers in angiosperms? This question is far less simple than it sounds, as is evident from the complex traditional terminology accumulated by botanists in their attempts to accurately describe inflorescences and to clarify their boundaries with true flowers. Evolutionary–developmental genetics has in recent years given us a much deeper understanding of the MADS-box genes that determine the fate and spatial organization of floral organ primordia, epitomized by the pivotal ‘ABC’ model and its derivatives, in which the C-class (AGAMOUS-like) genes not only interact with B-class genes to determine carpel identity but also ensure the determinacy of the floral axis (e.g. Coen and Meyerowitz, 1991). In contrast, the wide spectrum of inflorescence architectures has proved far less tractable to modelling. The key genes that regulate inflorescences are TERMINAL FLOWER1 (TFL1), LEAFY (LFY) and APETALA1 (AP1); differences in the form of closed inflorescences can be explained by evolutionary shifts in the respective expression levels of TFL1-like and AP1-like genes, which interact with other genes to specify FM identity via a series of feedback loops (Ma et al., 2017).
Although the phylogenetic context among angiosperms has changed through recent decades, it is likely that the determinate inflorescence – perhaps a solitary terminal flower – represents the ancestral type, though many transitions have occurred within numerous clades (Rudall and Bateman, 2010). Indeed, FUMs are apparently widespread across the angiosperm phylogeny. For example, Claßen-Bockhoff and co-workers have documented FUMs in detail in several eudicot families in addition to Asteraceae (Claßen-Bockhoff and Bull-Hereñu, 2013; Baczyński et al., 2022). They supplemented their observations with data from other sources, not only for monocots and magnoliids, but also for some of the ANA-grade families that consistently constitute the earliest-diverging lineages in molecular phylogenies.
For example, in Amborella, which is placed as the sister taxon to all other angiosperms in many molecular analyses, the inflorescence is determinate and resembles an FUM; the young flower or inflorescence axis is initially conical but expands laterally to form a flat floral meristem with marginal initiation of appendages (Posluszny and Tomlinson, 2003). Furthermore, reproductive meristems in the ANA-grade family Hydatellaceae (Nymphaeales) cannot readily be interpreted as either flowers or cymose inflorescences, despite using criteria ranging from comparative morphology to gene expression, since LFY proteins are localized in reproductive primordia at several different hierarchical levels (Rudall et al., 2009). In their influential attempt to reconstruct phylogenetically the morphology of the ancestral angiosperm flower, Sauquet et al. (2017) treated pseudanthia in general as inflorescences rather than flowers and chose to score Hydatellaceae as non-interpretable. Rudall et al. (2009) described these ambiguous structures as ‘non-flowers’, perhaps resembling a prefloral condition; they hypothesized iterative hierarchical shifts between flowers and inflorescences, a version of developmental heterochrony that is directly relatable to the FUM concept. Indeed, the highly iterative nature of this transition implies that the FUM was a major evolutionary innovation.
We might then move down the seed-plant phylogeny to consider the probable conditions in candidate groups still competing to be perceived as the closest relatives of angiosperms. Most molecular phylogenies based exclusively on extant taxa are unhelpful, placing angiosperms as sister to all other extant groups of seed-plants (e.g. Magallon et al., 2013). However, comparable but contrasting morphological phylogenies (e.g. Hilton and Bateman, 2006) have persistently placed some intriguing extinct groups known only from fossils, such as Bennettitales and Caytoniales, as potential angiosperm-sisters. In particular, many Bennettitales possessed terminal flower-like structures that were either solitary or arranged in cymose clusters (Rudall and Bateman, 2010). These reproductive structures can be readily envisaged as having developed via a laterally extensive FUM-like meristem, generating the conical/discoid receptacles of Mesozoic bennettites such as Cycadeoidea and Williamsoniella (Fig. 1D, E: structurally a genuine flower by some definitions: Bateman et al., 2006).
Viewed from a more process-based perspective, the developmental model envisaged for the Asteraceae capitulum is appealing because it confers important yet complementary roles on heterochrony, spatial/geometric constraints and auxin clines. Clines in auxin across the capitulum prompt competition among peripheral bracts, ray florets and disc florets that in extreme cases can suppress one of these organ categories (Zhang et al., 2021). Similarly for Fibonacci spirals, the popular idea that the near-perfect packing of the disc flowers within sunflower pseudanthia is pre-programmed gives way to a more pragmatic hypothesis based not on mathematical perfection but rather on transient local auxin maxima generating crowded flower primordia (FMs) that are forced to jostle for position until the underpinning FUM is exhausted. In other words, the impressively precise floral patterning of Asteraceae is the emergent product of spatial constraint acting upon meristematic behaviour that in turn reflects transient auxin profiles. Thus, the morphological end-point is separated from any underlying change(s) in gene expression by a crucial series of epigenetic filters.
Such developmental systems are vulnerable to threshold effects that could reflect even subtle changes in developmental context, leading to one or more heterochronic shifts in the timing of transitions from indeterminacy to determinacy. Assuming heritability, such shifts would have the potential to alter any or all aspects of development downstream of the initial change (i.e. could cause extensive pleiotropy and so greatly modify the resultant phenotype). Such auxin-mediated transformations could potentially emerge across a single generation and, at least initially, without the need for strong directional or disruptive selection (Bateman and DiMichele, 2002). A high diversity of inflorescences would easily and simply evolve, helping to explain the frequency of evolutionary transitions to pseudanthia implied by the molecular phylogenies of angiosperms (e.g. Baczyński and Claßen-Bockhoff, 2023, fig. 3).
Classical terminology can sometimes help to describe morphological diversity, but it can also merely end-run rather than resolve questions of inflorescence evolution. The multiplicity of meristem categories involved in inflorescence ontogeny together constitute a less overt developmental system than that governing simple bona fide flowers – one where architecture, shape and size interact to allow a greater role for epigenetic processes, limiting some evolutionary possibilities while simultaneously opening the door to others. Developmental morphologists raised on the ABC model of flower development, formulated under the influence of only a single category of meristem (FM), are now successfully grasping the nettle of inflorescence ontogeny (e.g. Ma et al., 2017). Nonetheless, they may be obliged by the FUM hypothesis to widen their horizons still further, with regard to both homology assessment and modes of gene expression.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baczyński J, Celep F, Spalik K, Claßen-Bockhoff R. 2022. Flower-like meristem conditions and spatial constraints shape architecture of floral pseudanthia in Apioideae. Evo Devo 13: 19. doi:10.1186/s 13227-022-00204-636536450 PMC 9764545 · doi ↗ · pubmed ↗
- 2Baczyński J, Claßen-Bockhoff R. 2023. Pseudanthia in angiosperms: a review. Annals of Botany 132: 179–202. doi:10.1093/aob/mcad 10337478306 PMC 10583202 · doi ↗ · pubmed ↗
- 3Bateman RM, Di Michele WA. 2002. Generating and filtering major phenotypic novelties: neo Goldschmidtian saltation revisited. In: Cronk QCB, Bateman RM, Hawkins JA. eds. Developmental genetics and plant evolution. London: Taylor and Francis, 109–159.
- 4Bateman RM, Hilton J, Rudall PJ. 2006. Morphological and molecular phylogenetic context of the angiosperms: contrasting the ‘top-down’ and ‘bottom-up’ approaches to inferring the likely characteristics of the first flowers. Journal of Experimental Botany 57: 3471–3503. doi:10.1093/jxb/erl 12817056677 · doi ↗ · pubmed ↗
- 5Claßen-Bockhoff R, Bull-Hereñu K. 2013. Towards an ontogenetic understanding of inflorescence diversity. Annals of Botany 112: 1523–1542. doi:10.1093/aob/mct 00923445936 PMC 3828942 · doi ↗ · pubmed ↗
- 6Claßen-Bockhoff R, Baczyński J, Hanke V, Henkes SS, Ferdinand N. 2026. Are capitula inflorescences? A reassessment based on flower-like meristem identity and ray flower development. Annals of Botany 137: 47–77. doi:10.1093/aob/mcaf 144PMC 1278408340632900 · doi ↗ · pubmed ↗
- 7Coen ES, Meyerowitz EM. 1991. The war of the whorls: genetic interactions controlling flower development. Nature 353: 31–37. doi:10.1038/353031 a 01715520 · doi ↗ · pubmed ↗
- 8Crane PR . 1985. Phylogenetic analysis of seed plants and the origin of angiosperms. Annals of the Missouri Botanical Garden 72: 716–793. doi:10.2307/2399221 · doi ↗