Complete transition from chromosomal to cytoplasmic sex determination during prolonged Wolbachia symbiosis
Takahiro Fukui, Tomohiro Muro, Noriko Matsuda-Imai, Tatsunori Kaneda, Hidetaka Kosako, Hideaki Hiraki, Keisuke Shoji, Takeshi Fujii, Yutaka Suzuki, Atsushi Toyoda, Takehiko Itoh, Takashi Kiuchi, Susumu Katsuma

TL;DR
Wolbachia bacteria, not the host's genes, control sex determination in infected Ostrinia moths through a long-term evolutionary relationship.
Contribution
The study identifies a Wolbachia-encoded mechanism that hijacks the host's feminizing function during prolonged symbiosis.
Findings
OfFem piRNA, a W-linked feminizer, is absent in Wolbachia-infected Ostrinia moths.
Ofznf-2 is essential for masculinization and dosage compensation in the host.
Wolbachia acquires the Oscar protein to hijack the host's feminizing piRNA function.
Abstract
Wolbachia infection causes male-specific death in Ostrinia furnacalis, but its removal from infected strains results in female-specific death instead of restoring 1:1 sex ratio, suggesting that cytoplasmic Wolbachia, not the host genome, primarily determines femaleness in infected strains. This phenomenon is a striking example of the evolutionary outcome of cytoplasmic sex determination, potentially arising from prolonged host-symbiont co-evolution. Although we recently identified Oscar, the Wolbachia-encoded male-killing effector targeting the host masculinizing factor OfMasc in Ostrinia moths, inactivation or loss of the host’s endogenous feminizer remains unknown. Here we identify a W-linked primary feminizer, OfFem piRNA, which targets an mRNA encoding an OfMasc-interacting protein Ofznf-2. We demonstrate that Ofznf-2 is essential for both masculinization and dosage compensation. We…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
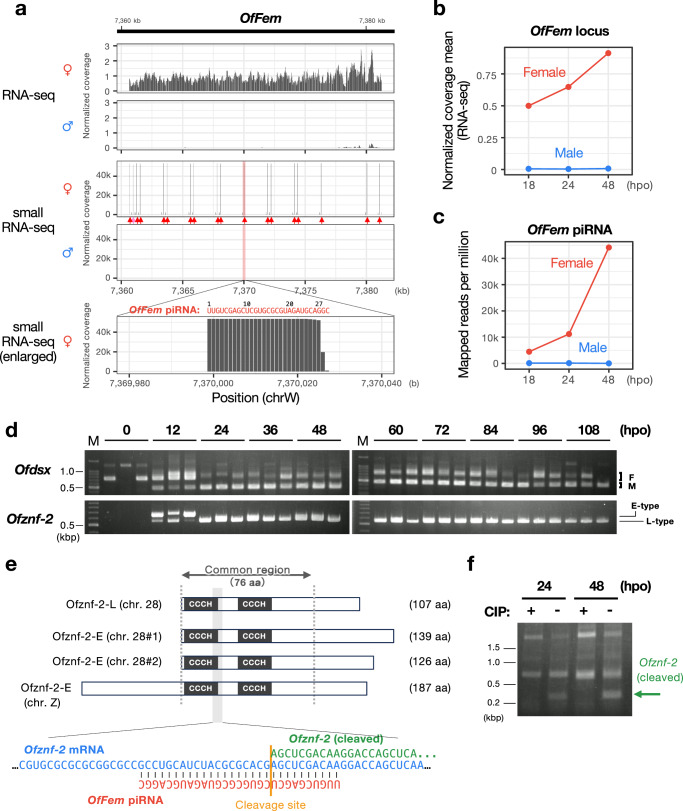 Figure 1
Figure 1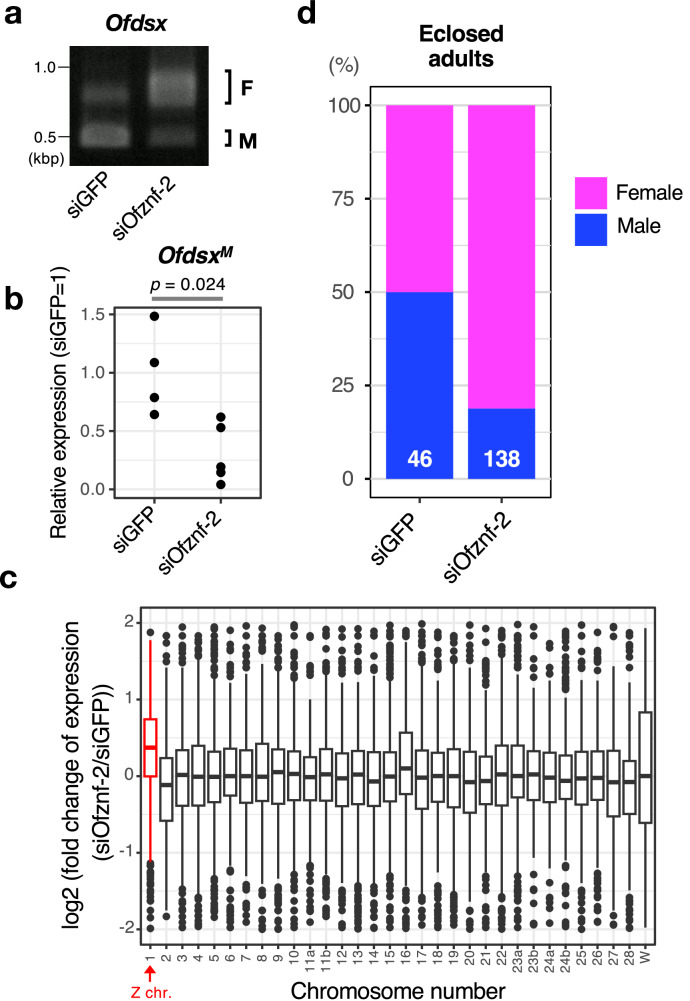 Figure 2
Figure 2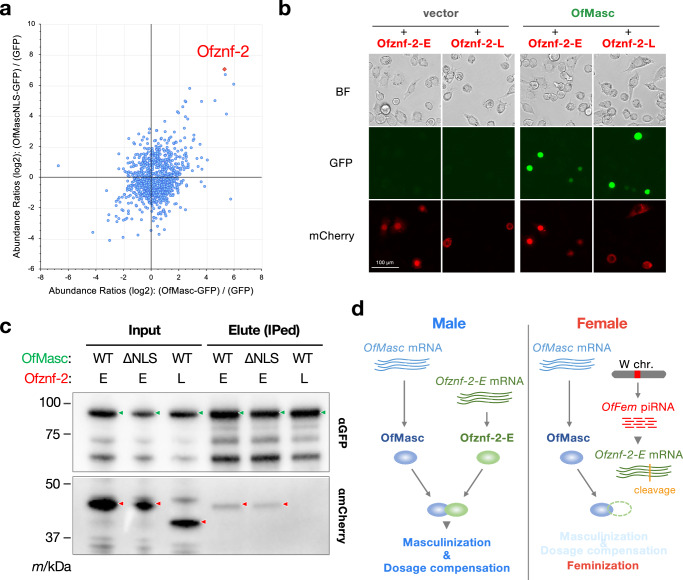 Figure 3
Figure 3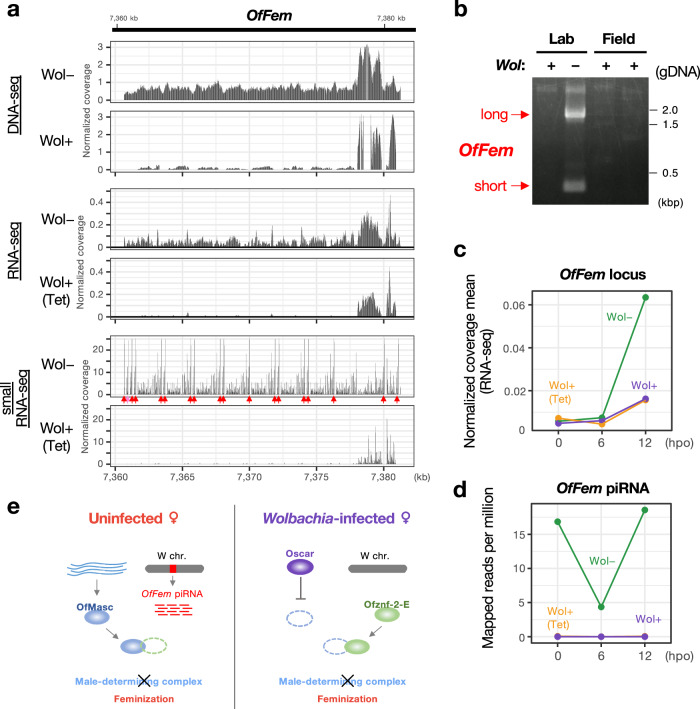 Figure 4
Figure 4- —https://doi.org/10.13039/501100001691MEXT | Japan Society for the Promotion of Science (JSPS)
- —G-7 Scholarship Foundation (no grant number)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect symbiosis and bacterial influences · CO2 Reduction Techniques and Catalysts · Microbial Fuel Cells and Bioremediation
Introduction
Some bacterial endosymbionts, such as Wolbachia species, manipulate the reproductive systems of arthropod hosts through male killing, feminization, cytoplasmic incompatibility, or parthenogenesis^1^. In various host-endosymbiont combinations, male killing causes infected mothers to produce only female progeny by selectively killing the male offspring^2^. In the interaction between Ostrinia species and male-killing Wolbachia strains (wSca or wFur, Fig. S1), an evolutionary phenomenon termed “cytoplasmic sex determination” has been observed, as in a few other arthropod species^3–5^. In wFur-infected Ostrinia furnacalis, the Wolbachia male-killing protein Oscar essentially determines femaleness, as the host’s endogenous feminizing factor is considered to have been inactivated or lost during prolonged symbiosis^6–9^. While several bacterial effectors responsible for reproductive manipulations have recently been identified^9–12^, no molecular evidence has yet demonstrated the inactivation or absence of the host’s endogenous feminizing factor in symbiont-dependent cytoplasmic sex determination systems.
Lepidopteran sex chromosomes are female-heterogametic, with female-specific W chromosomes that are enriched in repetitive elements and transposons^13–16^. These elements serve as sources of PIWI-interacting RNAs (piRNAs), a class of small RNAs that silence transposons^17^. In Bombyx mori and Plutella xylostella, female-specific piRNAs originating from the W chromosome, called Feminizer (Fem) piRNAs, determine femaleness^18,19^. Fem piRNAs target an mRNA of Masculinizer (Masc), a lepidopteran-specific CCCH-tandem zinc finger protein gene essential for both masculinization and dosage compensation^18,20–30^. The Fem piRNA-PIWI protein complex cleaves Masc mRNA^18^, inhibiting male-type splicing of doublesex (dsx) and dosage compensation in male embryos^18,31^, making Fem piRNAs the primary determinant of sex in Lepidoptera. In contrast, we discovered that in O. furnacalis, Masc (OfMasc) is not regulated by female-specific piRNAs during sex determination, indicating a unique sex determination system distinct from that of B. mori^32^.
In Japan, the infection rate of Wolbachia in O. furnacalis is relatively low^33^ (~15%), where infected individuals consistently harbor a single strain of wFur (wFur9)^33,34^. Because this infection causes complete male killing, where Wolbachia-infected females can only mate with uninfected males, the autosomal gene pool is shared between uninfected and Wolbachia-infected strains. Unlike male killing observed in other lepidopteran insects^35–39^, depleting wFur from infected moths using antibiotics induces female killing instead of restoring a 1:1 sex ratio^6,8^. This suggests that the primary feminizing factor, which may be located on the matrilineally inherited W chromosome, has been lost or inactivated in the Wolbachia-infected strain.
Recently, we demonstrated that the Wolbachia male-killing factor Oscar degrades OfMasc, thereby inhibiting male-type splicing of O. furnacalis dsx (Ofdsx) and dosage compensation^9,40^. Ectopic overexpression of Masc in females has been shown to cause female-specific death during larval stages^41,42^, as observed in female killing in antibiotics-treated Wolbachia-infected O. furnacalis, suggesting that Oscar directs the female-determining pathway by inactivating OfMasc in infected strains where the primary feminizing factor may be missing or nonfunctional. To unravel the molecular mechanism underlying Wolbachia-dependent cytoplasmic sex determination, identifying the endogenous feminizing factor of O. furnacalis and validating its absence or inactivation in infected females is essential.
Here, we identify a single feminizing piRNA derived from the W chromosome of O. furnacalis. Unlike Fem piRNAs in B. mori or P. xylostella, this piRNA targets an mRNA encoding Ofznf-2, a protein that interacts with OfMasc. Knockdown experiments reveal that Ofznf-2 is crucial for male-type dsx splicing and dosage compensation. Additionally, we find that this piRNA is entirely absent in the Wolbachia-infected lineage of O. furnacalis. These findings provide molecular evidence that a male-killing Wolbachia hijacks the role of a W-linked feminizing piRNA by acquiring the Oscar protein during prolonged endosymbiosis.
Results
A single W-linked piRNA targets mRNAs for CCCH-type zinc finger proteins in O. furnacalis
We hypothesized that the feminizing factor in O. furnacalis had been inactivated or lost in the Wolbachia-infected lineage. To identify the primary feminizing factor in O. furnacalis, we sequenced the transcriptomes of molecularly sexed early embryos during the period of sex determination^32^; the sex-specific splicing of Ofdsx is first observed after 12 hours post-oviposition (hpo) and is established at 48 hpo. From the de novo assembled transcriptome, 14 contigs were found to be significantly more highly expressed in females than in males across all embryonic stages (Fig. S2a). Among these, two contigs met the following criteria: (i) higher expression in uninfected embryos and ovaries compared to infected ones, and (ii) higher expression in uninfected ovaries compared to uninfected testes (Fig. S2b). In two in-house assembled whole-genome assemblies (version #1 and version #2, derived from O. furnacalis collected in Japan; Table S1) and the published genome of O. furnacalis^43^ (collected in China), these two contigs were mapped to a single locus on the W chromosomes. Fragments from this locus were PCR-amplified exclusively from female genome samples (Fig. S3a, b). Combined with the observation that its expression was restricted to females, not males, during the sex determination period (Figs. 1a, b, S3c, and S4a, b), this locus emerged as a strong candidate for encoding the feminizing factor. We designated this locus as O. furnacalis Feminizer (OfFem). The OfFem locus consisted of numerous repeat elements (Fig. S5a), a typical feature of piRNA clusters^44–46^. Small RNA sequencing (small RNA-seq) revealed that a single female-specific piRNA (OfFem piRNA) was abundantly produced from the OfFem locus in embryos and gonads (Figs. 1a, c and S4a, c).Fig. 1A single female-specific OfFem piRNA targets Ofznf-2 mRNA in *Ostrinia furnacalis.*a Mapping of RNA-seq and small RNA-seq reads on the OfFem locus in sexed eggs (48 hpo). Arrows indicate the positions of the OfFem piRNA-producing sites. Red arrows denote exact matches to OfFem piRNA, and a pink arrow shows a sequence with a single nucleotide mismatch. b Expression profile of OfFem transcripts in sexed eggs during the sex determination period. c Expression profile of OfFem piRNA in sexed eggs during the sex determination period. d Splicing patterns of Ofdsx mRNA and expression of Ofznf-2 variants during embryogenesis (n = 3). e Structures of Ofznf-2 variants annotated in the version #2 genome (top), with the putative OfFem piRNA-mediated cleavage site indicated by a yellow line (bottom). f Cleavage site identification of Ofznf-2 mRNA through RACE analysis of mRNA fragments without calf intestinal phosphatase (CIP) treatment. Nucleotide sequences of the cloned fragments are detailed in Fig. S11. Similar results were obtained in two technical replicates. Source data are provided as a Source data file.
The most plausible target gene of OfFem piRNA, named Ofznf-2, is a putative homolog of the O. scapulalis zinc finger gene Osznf-2^47,48^. Within the OfFem locus, sequences of 162–197 base pairs (bp) homologous to Ofznf-2 were repeated 17 times (Fig. S5a, b). Six loci containing Ofznf-2 genes were identified across the O. furnacalis genome (version #2) (Fig. S6). Two copies of Ofznf-2 on the W chromosome appeared pseudogenized due to a 6-kbp-long insertion (Fig. S6), so they were excluded from further analysis. The remaining four copies were categorized into two expression types: late type (Ofznf-2-L) and early type (Ofznf-2-E) (Fig. 1d, e). Additionally, Ofznf-2-E contained a specific 75–131 bp-long insertion absent in Ofznf-2-L (Fig. S6). A copy of Ofznf-2-L on the 28th chromosome was expressed throughout embryogenesis after 12 hpo, while two copies on the 28th chromosome and one on the Z chromosome, classified as Ofznf-2-E, were expressed exclusively around 12 hpo, which is the onset of sex determination^32^ (Figs. 1d and S7a). Both the version #1 genome (derived from a different individual from that used for version #2 genome) and the published genome^43^ also encoded at least one copy of Ofznf-2-L and Ofznf-2-E, respectively (Fig. S8, Table S2), suggesting variability in the copy number of Ofznf-2-E among O. furnacalis populations.
Across the genome, OfFem piRNA perfectly matched the antisense orientation of Ofznf-2 sequences (Figs. 1e and S6). All copies of Ofznf-2-E and Ofznf-2-L encoded short proteins (126–187 aa and 107 aa, respectively) with two CCCH-type zinc finger domains (Figs. 1e and S9a). These proteins showed high homology to Bmznf-2, which is required for proper male differentiation in B. mori^49,50^. In a cell line from O. scapulalis, male-type O. scapulalis dsx (Osdsx) was expressed, but transinfection with male-killing Wolbachia (wSca) induced the expression of female-type Osdsx and altered Osznf-2 expression^47,48^. These observations support the role of Ofznf-2 proteins in sex determination. A BLASTp search identified homologs of Ofznf-2 across Lepidoptera, all containing two CCCH-type zinc finger domains similar to Masc homologs (Fig. S10). A rapid amplification of cDNA ends (RACE) experiment confirmed that OfFem piRNA-mediated cleavage of Ofznf-2 mRNA occurred in early embryos (Figs. 1f and S11). At this stage, Ofznf-2 mRNA expression was higher in males than in females (Fig. S7b, c). These findings suggest that OfFem generates a female-specific piRNA that suppresses Ofznf-2 mRNA, which may play a critical role in male-determining functions.
Ofznf-2 is required for male determination in O. furnacalis
To test whether Ofznf-2 is essential for male determination, we performed embryonic knockdown experiments using small interfering RNA (siRNA) designed to target the conserved sequence shared across all copies of Ofznf-2. Microinjection of Ofznf-2 siRNA inhibited the male-type splicing of Ofdsx (Fig. 2a, b) and led to the upregulation of Z-linked genes in egg masses (Fig. 2c). These findings indicate that Ofznf-2 is necessary for both male-type splicing of Ofdsx and dosage compensation. Furthermore, embryonic knockdown of Ofznf-2 produced adult moths with a female-biased sex ratio (Fig. 2d), suggesting that Ofznf-2 plays a critical role in male viability. All observed phenotypes mirrored those resulting from the knockdown or degradation of OfMasc^8,9,21,32,40^. Additionally, two of the 26 male adults that emerged from eggs injected with Ofznf-2 siRNA displayed female-like wing morphology (Fig. S12), further supporting the role of Ofznf-2 in masculinization.Fig. 2Ofznf-2 is required for male-type splicing of Ofdsx and dosage compensation in *Ostrinia furnacalis.*a Splicing patterns of Ofdsx mRNA in egg masses injected with GFP or Ofznf-2 siRNA at 48 hpo. Similar results were obtained in three independent experiments. b Expression levels of Ofdsx^M^ in egg masses injected with GFP (n = 4) or Ofznf-2 (n = 5) siRNA at 48 hpo. A two-sided Welch’s t-test indicated a significant difference between the two groups, t(5.1141) = 3.19, p = 0.024, Cohen’s d = 2.24, 95% CI = [0.138, 1.25]. c Chromosomal distribution of differentially expressed transcripts in egg masses injected with Ofznf-2 siRNA at 48 hpo. The box shows the interquartile range (IQR = Q3−Q1), with a line at the median. The whiskers extend from the edge of the box to the most extreme values within 1.5× IQR. Only values in the range −2 to 2 are shown. The numbers of transcripts analyzed per chromosome (ordered left to right in the panel) are: 675, 331, 607, 747, 759, 546, 486, 587, 539, 673, 380, 583, 578, 579, 410, 792, 539, 650, 545, 622, 469, 445, 642, 560, 231, 276, 387, 576, 387, 279, 343, and 632. d Sex ratio of adult moths emerging from embryos injected with GFP- or Ofznf-2 siRNA, with sample sizes indicated for each group. Source data are provided as a Source data file.
Ofznf-2 cooperates with OfMasc in the masculinizing pathway in O. furnacalis
Given that OfMasc and Ofznf-2 are both required for male determination in O. furnacalis, we hypothesized that these proteins physically interact. To test this, we used a cell line derived from O. furnacalis embryos^9^. Expression of GFP-tagged OfMasc or OfMascNLS (a derivative of OfMasc lacking the active nuclear localization signal but retaining masculinizing activity^51^) in the O. furnacalis cell line, followed by immunoprecipitation (IP) and LC-MS/MS analysis, identified Ofznf-2 as an interacting partner for both OfMasc and OfMascNLS (Fig. 3a). When mCherry-tagged Ofznf-2-E or Ofznf-2-L was co-expressed with OfMasc-GFP or OfMascNLS-GFP in B. mori BmN-4 cells, only Ofznf-2-E-mCherry co-localized with OfMasc-GFP or OfMascNLS-GFP in the nucleus or cytoplasm (Figs. 3b, S9b and S13). A co-IP experiment further confirmed the protein interaction between OfMasc and Ofznf-2-E, but not between OfMasc and Ofznf-2-L (Fig. 3c). These results suggest that Ofznf-2-E, rather than Ofznf-2-L, physically associates with OfMasc to facilitate male-specific splicing of Ofdsx and dosage compensation in males. In females, these processes are inhibited through OfFem piRNA-mediated cleavage of Ofznf-2 mRNA (Fig. 3d).Fig. 3. Ofznf-2-E physically interacts with OfMasc.a LC-MS/MS analysis of proteins co-immunoprecipitated with anti-GFP nanobody from cells transfected with GFP, OfMasc-GFP, or OfMascNLS-GFP. Dots represent O. furnacalis proteins quantified by label-free precursor ion analysis. b Fluorescence microscopy of BmN-4 cells co-transfected with OfMasc-GFP and Ofznf-2-mCherry cDNAs. Similar results were obtained in two independent experiments. c Co-immunoprecipitation experiments using BmN-4 cells co-transfected with OfMasc-GFP or OfMascNLS-GFP and Ofznf-2-mCherry cDNAs. The immunoprecipitates with anti-GFP nanobody beads were immunoblotted using anti-mCherry antibody. GFP- and mCherry-fused protein bands are highlighted with green and red arrowheads, respectively. Similar results were obtained in two independent experiments. d Proposed model of the sex determination mechanism involving Ofznf-2 and OfMasc in O. furnacalis. Source data are provided as a Source data file.
A Wolbachia hijacks the host feminizing piRNA function in O. furnacalis
Previous studies suggested that the endogenous feminizing factor is either absent or inactivated in the Wolbachia-infected lineage of Ostrinia species^6–8^. In this lineage, the Wolbachia male-killing protein Oscar inactivates OfMasc protein, effectively substituting for the host feminizing factor^9,40^. Whole-genome sequencing with short reads from a Wolbachia-infected female revealed that almost no reads mapped to the OfFem locus (Fig. 4a). In the draft assembly of Wolbachia-infected O. furnacalis, the contig tig00004611 contained a single 222 bp-long region homologous to the OfFem piRNA-producing region (80.2% identity), but it lacked the repeat structure homologous to Ofznf-2 (Fig. S14). Additionally, fragments of the OfFem locus could not be PCR-amplified from field-collected Wolbachia-infected Ostrinia individuals (Fig. 4b). Consistently, the expression levels of OfFem and OfFem-derived piRNA were nearly undetectable in the Wolbachia-infected lineage (Figs. 4a, c, d and S15). These findings indicate that the OfFem locus is absent from the W chromosome in the Wolbachia-infected lineage. As a result, this W chromosome lacks feminizing ability through the downregulation of Ofznf-2 mRNA. This absence explains the sex-reversal phenomenon in O. furnacalis infected with male-killing Wolbachia, where the depletion of Wolbachia from infected mothers leads to female killing (Fig. S16).Fig. 4OfFem piRNA is absent from the W chromosome of Wolbachia-infected *Ostrinia furnacalis.*a Comparative mapping of DNA-seq, RNA-seq, and small RNA-seq reads onto the OfFem locus in Wolbachia-infected (Wol+), uninfected (Wol−), and tetracycline-treated Wolbachia-infected (Wol+ (Tet)) O. furnacalis egg masses at 12 hpo. Arrows indicate the positions of the OfFem piRNA-producing sites. Red arrows denote exact matches to OfFem piRNA, and a pink arrow shows a sequence with a single nucleotide mismatch. b PCR amplification of the OfFem locus from the genomes of field-collected O. furnacalis moths infected with male-killing Wolbachia (n = 2). Expression profile of the OfFem locus (c) and OfFem piRNA (d) in uninfected, Wolbachia-infected, and tetracycline-treated Wolbachia-infected egg masses. e Proposed model illustrating the sex determination system hijacked by male-killing Wolbachia in O. furnacalis. Source data are provided as a Source data file.
Discussion
We recently identified the Wolbachia protein Oscar and demonstrated its feminizing function through the inhibition of OfMasc activity^9,40^. In this study, we identified the host feminizing factor OfFem and showed its absence in Wolbachia-infected females. These findings provide molecular evidence for sex-determination hijacking: Wolbachia’s Oscar has fully taken over the role of the host’s feminizing piRNA in O. furnacalis during endosymbiosis (Fig. 4e). Beyond male killing in Ostrinia species, cytoplasmic sex determination, where an endosymbiont entirely hijacks the host’s sex-determination factor, has been observed in two other female-heterogametic hosts: the terrestrial isopod Armadillidium vulgare (pill bug) and the butterfly Eurema mandarina, both infected with feminizing Wolbachia^3,4^. In E. mandarina, Wolbachia depletion results in all-male offspring, suggesting that Wolbachia hijacks the role of the host’s feminizing factor. Given that this feminizing Wolbachia encodes an Oscar homolog^48^ and the W chromosome is entirely absent from infected strains of E. mandariana^4,5^, it is likely that the loss of the feminizing piRNA and its replacement by Oscar has occurred in this host-endosymbiont system.
Most lepidopteran species infected with male-killing bacteria retain their original feminization system, as indicated by the restoration of a 1:1 sex ratio following antibiotic-induced bacterial depletion^35–39^. A recent study identified Oscar homologs in the genomes of male-killing Wolbachia infecting several lepidopteran hosts, supporting the feminizing activity of Oscar homologs^48^. This suggests that both Oscar and the host feminizing factor coexist in these host-endosymbiont combinations, and that the presence of redundant feminizing factors does not always lead to the endosymbiont’s complete hijack of the host’s sex determination system. On the other hand, in species where sex is determined by the number of Z chromosomes rather than a W chromosome-encoded feminizing factor^52^, the feminizing system cannot be hijacked cytoplasmically. Considering the evolutionary lability of piRNA clusters^46,53^, species with feminizing piRNA encoded on their W chromosomes may be more vulnerable to sex determination hijacking. Further investigations into the feminizing systems of lepidopteran insects infected with male-killing symbionts could help validate this hypothesis. Additionally, although a loss-of-function mutation in a W-linked feminizing factor is unlikely to provide any selective advantage, why has such a mutation become fixed in a Japanese population? We propose that founder effects are responsible for this. In O. furnacalis, nine Wolbachia strains have been identified in China^54^, but only one of them (wFur9) has been observed in Japan^33,34^. This suggests that after a small number of wFur9-infected individuals carrying the OfFem-lacking W chromosome have been introduced from China to Japan, an increase in Wolbachia prevalence due to male killing has facilitated the rapid spread of the OfFem-lacking W chromosome, akin to the spread of neo-W chromosomes hitchhiking via male-killing Spiroplasma^55^. Investigating the presence of the OfFem locus in Chinese populations could clarify whether the OfFem-lacking W chromosome is unique to the Japanese population.
Bombyx mori has two homologs of znf-2, Bmznf-2 (KWMTBOMO15133) and Bmznf-2L (KWMTBOMO15134), located tandemly on the 25th chromosome (Fig. S17a). In BmN-4 cells, both protein homologs displayed nuclear localization, and co-expression with BmMascNLS (a derivative of B. mori Masc lacking an active nuclear localization signal^56^) induced the nuclear localization of BmMascNLS, indicating their association with BmMasc (Fig. S17b–e). A co-IP experiment further confirmed the physical association of Bmznf-2 and Bmznf-2L with BmMasc (Fig. S17f, g). As previously reported^49^, Bmznf-2 showed distinct masculinizing activity in BmN-4 cells (Fig. S17h), whereas Bmznf-2L exhibited a small positive effect (Fig. S17h). Notably, when co-expressed with BmMasc, both homologs enhanced BmMasc-dependent masculinizing activity (Fig. S17h). Together with evidence of the functional interaction between OfMasc and Ofznf-2, these findings suggest that a complex comprising two or more CCCH-tandem zinc finger proteins is likely a conserved regulatory element for male-determining functions in Lepidoptera. Recently, van’t Hof et al. proposed that a heterozygotic combination of Masc alleles is essential for male development in Bicyclus anynana butterflies^29^. This strongly supports the hypothesis that heterodimerization of CCCH-tandem zinc finger proteins is crucial for forming a functional male-determining complex, with the composition of CCCH-tandem zinc finger proteins in this complex potentially varying across species.
In this study, we identified OfFem, the feminizing factor on the W chromosome, in uninfected O. furnacalis. This marks the fourth instance of a primary sex determiner being molecularly identified in Lepidoptera^18,19,30^. OfFem acts as a precursor to a single female-specific piRNA, which targets Ofznf-2 mRNA. Functional Ofznf-2 genes were identified at multiple genomic loci and categorized into two types: Ofznf-2-E and Ofznf-2-L. While Ofznf-2-E is expressed early in embryogenesis, Ofznf-2-L is expressed throughout embryogenesis. Knockdown experiments revealed that either or both Ofznf-2 types are essential for two male-specific biological processes: male-type dsx splicing and dosage compensation. In two other lepidopteran species, B. mori and P. xylostella, the male-specific processes are inactivated by feminizing piRNAs targeting Masc mRNA^18,19^. The homologs of Masc in these species perform conserved roles in these processes. Therefore, all feminizing factors identified so far are female-specific piRNAs that target genes essential for male-determining processes. Given the physical interaction between OfMasc and Ofznf-2-E in O. furnacalis, other genes encoding components of the male-determining complex are plausible targets for W-linked feminizing piRNAs. Future studies may uncover the diversity of feminizing piRNA targets, emphasizing the role of the repetitive W chromosome as the origin of feminizing piRNAs. Additionally, we found that OfFem piRNA is derived from the tandem, degenerate copies of Ofznf-2 on the W chromosome. Similarly, feminizing piRNAs in P. xylostella originate from the retrocopies of Masc on the W chromosome^19^. These findings strongly suggest that the transposition of genes with male-determining functions to the W chromosomes represents the initial step in the evolution of feminizing piRNAs.
Methods
Insect rearing and tissue collection
The founder O. furnacalis moths of the laboratory strain were collected during the early summer seasons of 2014–2021 in Japan. The laboratory strains were maintained according to previously described methods^32^. Egg masses were harvested within a 2-h time window at designated time points for further experiments. Genomic DNA and total RNA were extracted using the DNeasy Blood & Tissue Kit (QIAGEN) and TRI reagent (Molecular Research Center), respectively.
Genomic PCR and reverse transcription-PCR (RT-PCR)
The OfFem locus was amplified using KOD One DNA polymerase (TOYOBO, Japan) with the primers designed to match the homologous repeats to Ofznf-2 in the OfFem locus. The sex-specific splicing pattern of Ofdsx was examined by RT-PCR using KOD FX–neo DNA polymerase (TOYOBO, Japan) as previously described^32^. Quantitative RT-PCR (RT-qPCR) was performed with the KAPA SYBR FAST qPCR kit (Kapa Biosystems). Expression levels of Ofdsx^M^ were normalized to Ofdsx^C^, the common region of the male- and female-type Ofdsx. RT-qPCR primers specifically targeting Ofznf-2-E are detailed in Fig. S6, while all primers used in this study are listed in Table S3.
Modified rapid amplification of cDNA ends (RACE)
Modified RACE experiments followed a previously established protocol with minor adjustments^18,57^. The GeneRacer Kit (Invitrogen) was employed according to the manufacturer’s instructions, excluding calf intestine phosphatase (CIP) treatment. Omitting this step enabled adapter oligonucleotide ligation to both the capped and 5′-OH mRNA molecules for comprehensive 5′-end analysis.
RNA-seq and small RNA-seq
Total RNA samples were isolated from individual eggs at 18, 24, and 48 hpo and were molecularly sexed following established protocols^32^. Sexed egg samples were either pooled by sex or analyzed individually. RNA-seq libraries were constructed from these samples using the Stranded mRNA Prep, Ligation Kit (Illumina), and indexed with IDT for Illumina RNA UD Indexes SetA. The libraries were sequenced on the NovaSeq 6000 Sequencing System (Illumina, San Diego, CA, USA) with 100-bp paired-end reads. Small RNA-seq was performed on samples depending on the experimental setup. For sex-pooled egg samples, RNA fractions of 18–30 nucleotides in length were isolated and used to construct small RNA-seq libraries. Sequencing of these libraries was conducted on the DNBSEQ platform, with library preparation and sequencing services provided by BGI (Shenzhen, China). For gonad and egg mass samples, total RNA was isolated from the testes of eight males, the ovaries of two to seven females, and egg masses collected from five females at each time point. Small RNA fractions (20–30 nucleotides in length) were isolated for these samples as described previously^32^. Small RNA-seq libraries were constructed and sequenced using the Illumina HiSeq 2500 platform. For RNA-seq analysis of egg mass samples, RNA-seq libraries were constructed using the Ribo-Zero rRNA Removal Epidemiology Kit and Illumina TruSeq RNA Prep Kit, which are non-strand-specific and omit poly(A) selection. These libraries were sequenced on the Illumina HiSeq 2500 platform.
Identification and expression analysis of OfFem locus and OfFem piRNA
The de novo transcriptome assembly was performed using the Trinity software package^58^. Contig expression levels were quantified using Salmon^59^, and the output files were processed with the R package Wasabi (https://github.com/COMBINE-lab/wasabi). Sex-biased contigs with statistically significant expression differences between male and female samples (q value < 0.05) were identified using the Sleuth software tool^60^. Two contigs that met several criteria (see Results) were mapped on the single OfFem locus identified from an in-house assembled whole-genome assembly (version #2), which is used for further analysis.
RNA-seq data were mapped to the OfFem locus using HISAT2^61^. Coverage depth at the locus was calculated with SAMtools^62^ and BEDTools^63^. Small RNA-seq data were mapped to the OfFem locus using Bowtie^64^, allowing up to two nucleotide mismatches and permitting multi-maps. OfFem piRNA reads were specifically quantified by mapping small RNA-seq data onto a single representative repeat of the Ofznf-2 sequence within the OfFem locus using BWA^65^ with no mismatches allowed. To infer the structure of the repeat elements in the OfFem locus, a custom repeat library was constructed de novo from the O. furnacalis whole-genome sequence (version #2) using RepeatModeler2^66^. The genome was subsequently annotated with RepeatMasker^67^, employing this custom repeat library. Homologous repeats to the Ofznf-2 sequence within the OfFem locus were identified by performing a BLASTn search with the nucleotide sequence of Ofznf-2-L as the query.
Phylogenetic analysis
Homologs of Ofznf-2, limited to the top hit from each species, were identified through a BLASTp search against the NCBI non-redundant protein database (Table S4). BLASTp search was performed via the NCBI BLAST homepage (https://blast.ncbi.nlm.nih.gov/) with default parameters, using as the query the partial protein sequence of Ofznf-2 (“MCKDWVRGTCARGAACIYAHELDKDQLKGVYRFCRDFENDRCERQVCYFVHATTFEKEHFFRTAFLPPHALHHLKT”). Phylogenetic analysis of Ofznf-2 homologs was conducted alongside previously published Masc protein sequences (Table S4). Amino acid sequences were aligned using MAFFT^68^ with default settings, and the alignments were trimmed with trimAl^69^ in “automated1” mode. A maximum likelihood tree was constructed via the IQ-TREE web server^70^, with node support calculated from 1000 ultrafast bootstrap replicates^71^.
Analysis of dosage compensation
RNA-seq libraries were prepared from total RNA using the Illumina platform protocols and sequenced as stranded libraries on the NovaSeq 6000 Sequencing System (Illumina, San Diego, CA, USA) with 150-bp paired-end reads. Library preparation and sequencing were performed by Novogene (Beijing, China). RNA-seq data were mapped to the annotated O. furnacalis genome (version #2) using HISAT2^61^. Gene expression levels were quantified with featureCounts^72^. Counts per million values and fold-change metrics were calculated using the edgeR package, employing normalization by the trimmed mean of M-values method^73^.
Cell culture and transfection
The DNA fragments encoding OfMasc-GFP, Ofznf-2, and their derivatives were cloned into the pIZ/V5-His-g3 vector^51^. B. mori BmN-4 cells were cultured at 26 °C in IPL-41 medium (Applichem, Germany) supplemented with 10% fetal bovine serum (FBS; Gibco, USA). BmN-4 cells (4.0 × 10^5^ cells per 35-mm diameter dish) were transfected with two plasmid DNAs (1 µg each) using FuGENE HD transfection reagent (Promega, USA). The localization of GFP-fused and mCherry-fused proteins was examined at 72 h post-transfection using either a FLoid^TM^ cell imaging station or an EVOS M5000 cell imaging station (Thermo Fisher Scientific). O. furnacalis OfT1C/tet cells, derived from Wolbachia-infected O. furnacalis embryos^9^, were cultured at 26 °C in Express Five^TM^ SFM medium (Gibco, USA) supplemented with 18 mM L-glutamine, 10% FBS, and 3 μg/mL tetracycline. We isolated three RT-PCR clones of Ofznf-2-E from O. furnacalis embryonic cDNA. These clones encoded proteins with slight differences in amino acid sequences, but all displayed similar localization patterns in BmN-4 cells (Fig. S9). Of these, we used Ofznf-2-E_clone #1 as the representative clone for all subsequent cell-based experiments.
LC-MS/MS-based identification of OfMasc-interacting proteins
OfMasc-GFP, OfMascNLS-GFP, or GFP were transiently expressed in OfT1C/tet cells (3.2 × 10^6^ cells) seeded in 10 cm-diameter culture dishes by transfection using FuGene HD (Promega). OfMasc-interacting proteins were co-immunoprecipitated with OfMasc-GFP using GFP-Trap magnetic agarose beads (ChromoTek, Germany). Proteins bound to the beads were digested by Trypsin/Lys-C mix (Promega). LC-MS/MS analysis of the resultant peptides was performed using an EASY-nLC 1200 UHPLC connected to an Orbitrap Fusion mass spectrometer equipped with a nanoelectrospray ion source (Thermo Fisher Scientific). The peptides were separated on a 75 μm inner diameter × 150 mm C18 reversed-phase column (Nikkyo Technos, Japan) with a linear 4–32% acetonitrile gradient for 0–100 min followed by an increase to 80% acetonitrile for 10 min. The mass spectrometer was operated in a data-dependent acquisition mode with a maximum duty cycle of 3 s. MS1 spectra were measured with a resolution of 120,000, an automatic gain control (AGC) target of 4e5, and a mass range from 375 to 1500 m/z. Higher-energy collisional dissociation MS/MS spectra were acquired in the linear ion trap with an AGC target of 1e4, an isolation window of 1.6 m/z, a maximum injection time of 35 ms, and a normalized collision energy of 30. Dynamic exclusion was set to 20 s. The mass spectrometry experiments were performed once for each sample. The raw LC-MS/MS data were directly analyzed against O. furnacalis protein data (GCF_004193835.1_ASM419383v1_protein.faa) downloaded from NCBI, supplemented with sequences for OfMasc-GFP and GFP-Trap. Analyses were performed using Proteome Discoverer version 2.5 software (Thermo Fisher Scientific) with Sequest HT search engine. The search parameters were as follows: (a) trypsin as an enzyme with up to two missed cleavages; (b) precursor mass tolerance of 10 ppm; (c) fragment mass tolerance of 0.6 Da; (d) carbamidomethylation of cysteine as a fixed modification; and (e) acetylation of protein N-terminus and oxidation of methionine as variable modifications. Peptides and proteins were filtered at a false discovery rate (FDR) of 1% using the percolator node and the protein FDR validator node, respectively. Label-free precursor ion quantification was performed using the precursor ions quantifier node, and normalization was performed such that the total sum of abundance values for each sample over all peptides was the same.
Western blotting and immunoprecipitation
Western blotting and immunoprecipitation experiments were performed as previously described^9^. Lysates from BmN-4 cells expressing GFP-tagged proteins were incubated with GFP-Trap magnetic agarose beads. After binding, the beads were washed three times and boiled to elute the bound proteins. The eluted proteins were separated by SDS-PAGE and transferred onto PVDF membranes. For immunodetection, an anti-GFP antibody (1:5000 dilution, MBL, 598, Japan) or an anti-RFP antibody (1:5000 dilution, MBL, PM005, Japan) was used as the primary antibody.
Microinjection of siRNA
Microinjection of siRNA was carried out as previously described^32^. Specifically, 100 μM siRNA solutions were injected into O. furnacalis embryos within 2 hpo. For Figs. 2d and S12, however, 50 µM siRNA was used. Egg masses injected with siRNA were collected at 48 hpo for analysis or allowed to hatch. Hatched larvae were raised to the pupal stage and sexed based on pupal terminal abdominal morphology. siRNAs were purchased from FASMAC Corp. (Atsugi, Japan), prepared in annealing buffer (100 mM potassium acetate, 2 mM magnesium acetate, 30 mM HEPES-KOH, pH 7.4) at concentrations of 100–500 μM, and stored at −80 °C. Nucleotide sequences for the Ofznf-2 siRNA used in this study are listed in Table S3.
DNA extraction and genome sequencing
Whole-genome shotgun sequencing was performed using PacBio and Illumina sequencing platforms. Genomic DNA was extracted from female larvae of laboratory strains (T1-A and T1-C1) of O. furnacalis using the Blood & Cell Culture DNA Midi Kit and a Genomic-tip Kit, respectively (QIAGEN, Hilden, Germany). DNA quality and concentration were assessed using a Qubit 4 Fluorometer (Thermo Fisher Scientific, MA, USA) and a Pippin Pulse electrophoresis system (Agilent Technologies, CA, USA).
For PacBio sequencing, a continuous long read (CLR) library was constructed from the T1-A sample using the SMRTbell Express Template Prep Kit 2.0 (Pacific Biosciences, CA, USA), followed by size selection with the Blue Pippin system (Saga Science, MA, USA) using a 30 kb cutoff. The library was sequenced on the PacBio Sequel II system with a Binding Kit 2.0 and Sequencing Kit 2.0 (Pacific Biosciences, CA, USA) using 30-h movies. Raw sequencing data from a single 8M SMRT cell were processed using the PacBio SMRT Link v11.0.0.144466 software. For Illumina sequencing, a paired-end library was generated using the Illumina DNA PCR-Free Library Prep Tagmentation Kit (Illumina, CA, USA) and sequenced on the NovaSeq 6000 platform (Illumina, CA, USA) with a 2 × 150 bp read length.
For High Fidelity (HiFi) sequencing, genomic DNA from the T1-C1 sample was fragmented into 15–20 kb sizes using a g-tube device (Covaris Inc., MA, USA). A HiFi library was prepared with the SMRTbell Prep Kit 3.0 (Pacific Biosciences, CA, USA) and subsequently size-selected with 35% AMPure PB beads (Pacific Biosciences, CA, USA). Sequencing was conducted on the PacBio Sequel II system using the Binding Kit 3.2 and Sequencing Kit 2.0 (Pacific Biosciences, CA, USA) with 30-h movies. HiFi reads were generated and processed from one 8M SMRT cell using the DeepConsensus v1.2 program under the default parameters^74^. Genomic DNA was also fragmented to an average size of 500 bp with the focused-ultrasonicator M220 (Covaris Inc., MA, USA). A paired-end library was prepared with the TruSeq DNA PCR-Free Library Prep Kit (Illumina, CA, USA) and size-selected using an agarose gel and the Zymoclean Large Fragment DNA Recovery Kit (Zymo Research, CA, USA). Sequencing library was then conducted on the NovaSeq 6000 system (Illumina, CA, USA) with a read length of 2 × 150 bp.
Hi-C library preparation and sequencing
The Hi-C library, derived from the T1-C8 larval body sample, was prepared following the Omni-C Kit protocol (Dovetail, CA, USA). Briefly, chromatin was fixed in the nucleus using disuccinimidyl glutarate and formaldehyde, then digested with DNase I. Extracted chromatin DNA fragments were end-repaired and ligated to a biotinylated bridge adapter. After proximity ligation, crosslinks were reversed, and DNA was purified. Sequencing library construction involved Illumina-compatible adapters, with biotin-labeled fragments enriched via streptavidin magnetic beads and PCR amplification. The final library was sequenced on the Illumina NovaSeq 6000 system with 2 × 150 bp paired-end reads, yielding a total of 98.3 Gb of sequence data.
Chromosome-level genome assembly
Two versions of the O. furnacalis genome assembly (version #1 and version #2) were constructed using PacBio CLR reads and HiFi reads, respectively. For version #1, PacBio CLR reads were assembled with Canu v2.1.1^75^, with the parameters “genome_size=400m corOutCoverage=200 batOptions="-dg 3 -db 3 -dr 1 -ca 500 -cp 50 " -pacbio-raw.” The resulting contigs were polished with Pilon v1.4.0^76^ using Illumina reads. To address redundancy caused by the independent assembly of haplotypes, haplotigs were removed with Purge_Dups v1.2.5^77^. Hi-C scaffolding was then performed using an Omni-C dataset to produce a chromosome-level assembly.
Before Hi-C scaffolding, 11 W chromosome-derived contigs, initially removed by the Purge_Dups process, were rescued using Hi-C contact information. Hi-C scaffolding was conducted with YaHS v1.2a^78^, utilizing Omni-C read mapping information generated by HiLine v0.2.4 (https://github.com/wtsi-hpag/HiLine). The Hi-C contact map was visualized with Juicebox v2.16.00^79^, underwent detailed manual curation to resolve assembly errors and mis-scaffoldings.
For version #2, the genome assembly was generated using Hifiasm v0.19.6^80^ with the parameter “--primary -s 0.30,” based on PacBio HiFi reads. The “-s” parameter was optimized by comparing assembly statistics from several trial runs, with the primary goal of reducing redundancy caused by unresolved haplotypes in the primary assembly. Obtained contigs were polished with NextPolish2 v0.2.0^81^ using PacBio HiFi reads and Illumina reads. Subsequent Hi-C scaffolding and manual curation were performed on the primary contigs (p_ctg) using the same procedures as for version #1.
Both assemblies demonstrated robust results, with N50 values greater than 16 Mbp and demonstrating >99% completeness of conserved genes, as evaluated by BUSCO^82^. Detailed assembly statistics for both versions are summarized in Table S1. The chromosome numbers were assigned based on synteny with the chromosomes of B. mori (Table S5).
Statistics and reproducibility
All statistical tests were two-sided. The significance threshold and tests used are indicated in each figure legend. Experiments were independently replicated as follows: Fig. 2a, three independent biological replicates; Fig. 3b, c, two independent biological replicates; Fig. 1f, two technical replicates; Figs. 1d (n = 3) and 4b (n = 2), one technical replicate.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Supplementary information
Supplementary Information Description of Additional Supplementary File Supplementary Data 1 Reporting Summary Transparent Peer Review file
Source data
Source data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hirota, K., Matsuda-Imai, N., Kiuchi, T. & Katsuma, S. Characterization of nuclear localization signal in Ostrinia furnacalis Masculinizer protein. Arch. Insect Biochem. Physiol. 106, e 21768 (2021).10.1002/arch.2176833644912 · doi ↗ · pubmed ↗
- 2Smit, A. F., Hubley, R. & Green, P. Repeat Masker Open-4.0. http://www.repeatmasker.org (2015).
- 3Zhou, C., Mc Carthy, S. A. & Durbin, R. Ya HS: yet another Hi-C scaffolding tool. Bioinformatics 39, btac 808 (2023).10.1093/bioinformatics/btac 808PMC 984805336525368 · doi ↗ · pubmed ↗
- 4Hu, J. et al. Next Polish 2: a repeat-aware polishing tool for genomes assembled using Hi Fi long reads. Genom. Proteom. Bioinform.22, qzad 009 (2024).10.1093/gpbjnl/qzad 009PMC 1201603638862426 · doi ↗ · pubmed ↗
- 5Manni, M., Berkeley, M. R., Seppey, M. & Zdobnov, E. M. BUSCO: assessing genomic data quality and beyond. Curr. Protoc. 1, e 323 (2021).10.1002/cpz 1.32334936221 · doi ↗ · pubmed ↗