Circular RNAs in Diabetic Foot Ulcers: A Scoping Review of Clinical, Preclinical, and In Silico Evidence on Diagnostic and Therapeutic Potentials
Amir Reza Ghafourian, Masoomeh Hamdi, Atefeh Soltan Mohseni, Maryam Davoudi, Hamid Choobineh, Fariba Nabatchian, Reza Afrisham

TL;DR
This review explores how circular RNAs could help diagnose and treat diabetic foot ulcers by influencing wound healing and inflammation.
Contribution
The study integrates clinical, preclinical, and computational evidence to highlight novel circRNA targets for diabetic foot ulcers.
Findings
Clinical studies link specific circRNAs to wound severity and tissue repair in diabetic foot ulcers.
Preclinical models show circRNA delivery improves wound healing and reduces inflammation.
In silico analysis reveals circRNA-miRNA-mRNA networks involved in inflammation and angiogenesis.
Abstract
Diabetic foot ulcers (DFUs) involve chronic inflammation, impaired angiogenesis, oxidative stress, and disrupted fibroblast–keratinocyte interactions. Circular RNAs (circRNAs), a category of stable non‐coding RNAs, have become essential regulators of these processes; nevertheless, their comprehensive functions in DFUs are still inadequately characterised. This scoping review integrated clinical, preclinical, and in silico evidence on circRNAs in DFUs to assess their diagnostic, mechanistic, and therapeutic potential. Systematic searches of MEDLINE, EMBASE, Web of Science, and Google Scholar were conducted on July 16, 2025, according to the PRISMA‐ScR guidelines. Eligible papers included clinical investigations of circRNAs in the tissues of patients with DFUs, preclinical animal models assessing circRNA‐based therapies, and computational predictions of circRNA‐miRNA‐mRNA networks.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
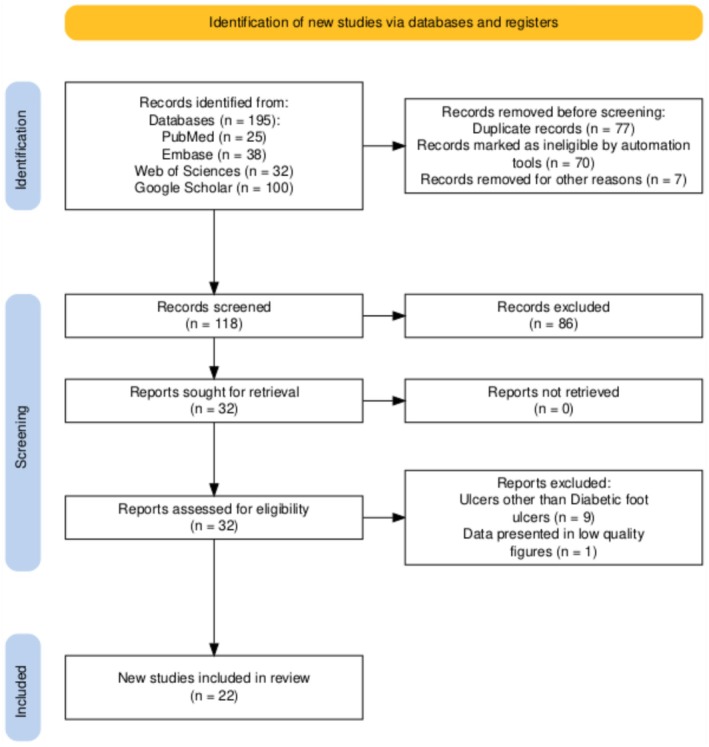 FIGURE 1
FIGURE 1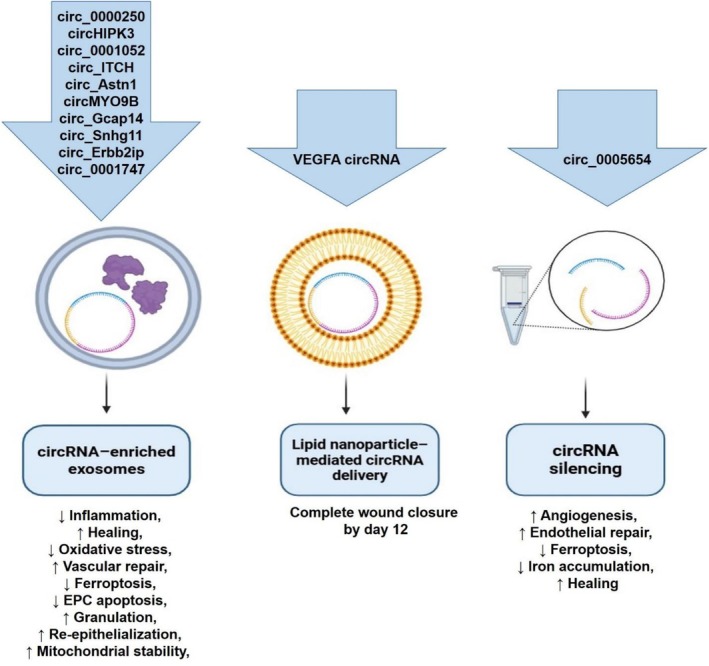 FIGURE 2
FIGURE 2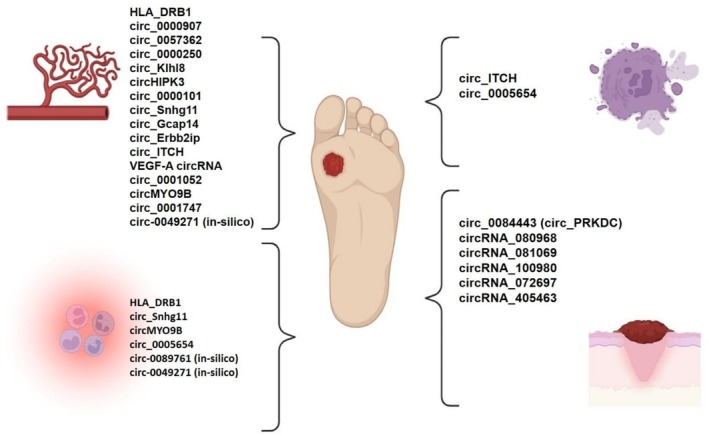 FIGURE 3
FIGURE 3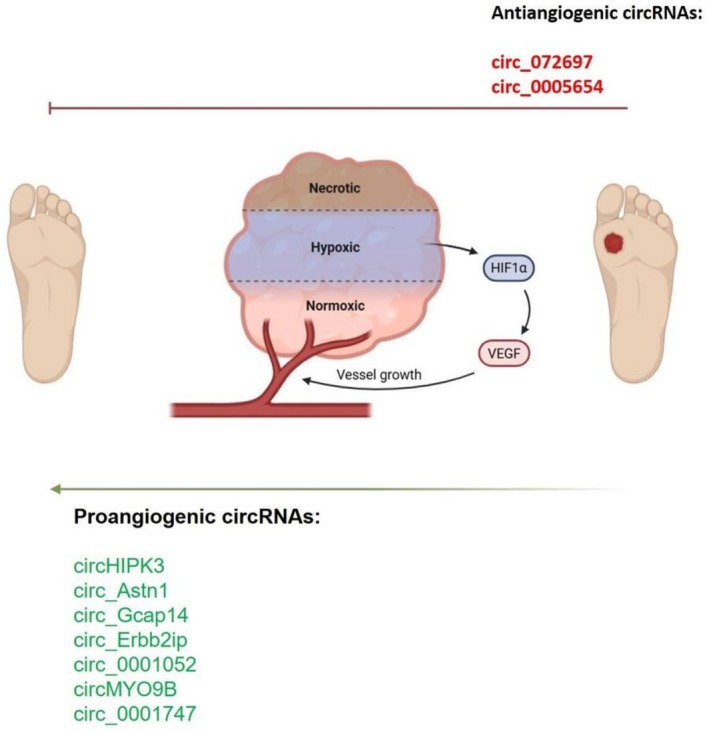 FIGURE 4
FIGURE 4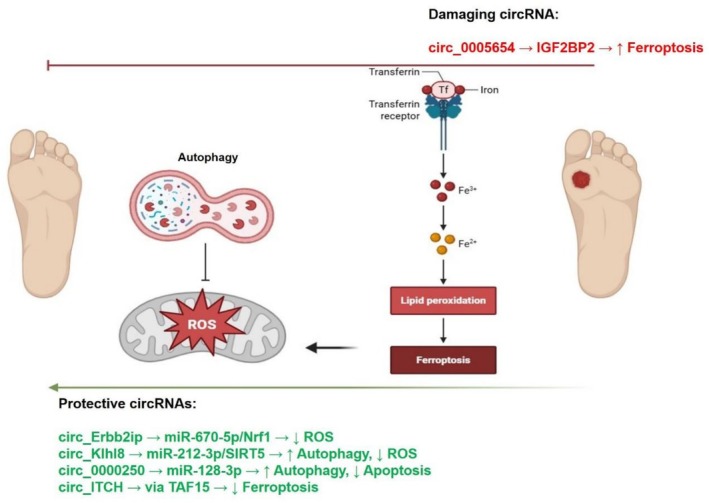 FIGURE 5
FIGURE 5| Database | Search query |
|---|---|
| PubMed | ((((RNA, circular[MeSH Te) OR (circRNA*[Title/Abstract])) OR (Circular RNA*[Title/Abstract])) OR (closed circular RNA*[Title/Abstract])) AND ((((((diabetic foot[MeSH Terms]) OR (diabetic foot ulcer*[Title/Abstract])) OR (diabetic foot[Title/Abstract])) OR (diabetic feet[Title/Abstract])) OR (foot ulcer*[Title/Abstract])) OR (DFUs[Title/Abstract])) |
| ISI Web of Science | ((TS = (circRNA*)) OR TS = (Circular RNA*)) OR TS = (closed circular RNA*) AND ((((TS = (diabetic foot ulcer*)) OR TS = (diabetic foot)) OR TS = (diabetic feet)) OR TS = (foot ulcer*)) OR TS = (DFUs) |
| Embase | ((TITLE‐ABS‐KEY (circrna*) OR TITLE‐ABS‐KEY (circular AND rna*) OR TITLE‐ABS‐KEY (closed AND circular AND rna*))) AND ((TITLE‐ABS‐KEY (foot AND ulcer*) OR TITLE‐ABS‐KEY (dfus) OR TITLE‐ABS‐KEY (diabetic AND foot AND ulcer*) OR TITLE‐ABS‐KEY (diabetic AND feet) OR TITLE‐ABS‐KEY (diabetic AND foot))) |
| No. | Authors (year; country) | Study design/Data source | Study groups (n) | DFU inclusion/Exclusion criteria | circRNA regulation in DFUs | miRNAs | Roles/Mechanistic pathways | Key findings (expression and diagnostic insights) |
|---|---|---|---|---|---|---|---|---|
| 1 | Wang et al. (2020; China) [ | Experimental case–control study with in vitro functional validation | DFU ( |
|
| NA | Regulates JAK/STAT, EGFR, PI3K, ERK, and HIF‐1 pathways; affects keratinocyte proliferation/motility. |
↓ in NHW vs. intact skin; ↑ in DFU vs. NHW Potential biomarker for wound chronicity |
| 2 | Fu et al. (2023; China) | Bioinformatics‐integrated clinical experimental study (GEO source: | DFU ( |
|
|
|
|
↑ ↓ migration and ↑ proliferation of keratinocytes |
| 3 | Tian et al. (2023; China) [ | Clinical case–control study with in vitro mechanistic validation (experimental functional study) | DFU ( |
| ↑ |
|
Impacts MAPK pathway | Its silencing restores proliferation/migration in AGEs‐treated HaCaT cells |
| 4 | Tian et al. (2020; China) | Bioinformatics‐integrated clinical experimental study (GEO source: | DFU ( | NA |
|
| Regulates KRAS/MAPK pathway | They proposed as novel DFU biomarkers |
| 5 | Chen et al. (2020; China) | Bioinformatics‐integrated clinical experimental study (GEO source: |
Discovery cohort: 10 pairs of DFU and NHW tissues analysed by qRT‐PCR for validation of bioinformatics results Validation cohort 1: DFU ( Validation cohort 2 (large‐scale diagnostic validation): DFU ( |
|
| NA | Serum circRNAs negatively correlate with ABI and TcPO2 in the patients | Both serum and exosomal |
| 6 | Han et al. (2021; China) [ | In vitro mechanistic validation study (a follow‐ups study for Wang et al.'s findings on the | Healthy volunteers with wounds ( |
| ↓ |
|
| hsa_circ‐PRKDC knockdown → ↓ miR‐31 → ↑ MMP‐2, MMP‐9 → enhanced keratinocyte migration |
| 7 | Bai et al. (2025; China) | In vitro experimental mechanistic study (based on high‐throughput sequencing and functional validation in HUVECs; GEO assigned: | DFU ( |
| ↑ |
|
| Mediates inflammation and angiogenesis |
| No. | Authors (year; country) | Animal model (sex, n) | Ulcer development (wound size) | Induction of diabetes & diagnostic indicators | Intervention & intervention contents | Follow‐up/Sample collection | Main findings/Mechanistic insights |
|---|---|---|---|---|---|---|---|
| 1 | Shi et al. (2020; China) [ | C57BL/6 mice (Male; | 4‐mm full‐thickness dorsal wound | STZ 60 mg/kg, single injection; glucose ≥ 250 mg/dL | Subcutaneous injection of ADSC exosomes (200 μg/100 μL PBS) containing mmu_circ_0000250 vs. PBS | Day 15 | Exosomal |
| 2 | Shang et al. (2021; China) [ | C57BL mice (NA) | 4‐mm full‐thickness dorsal wound | STZ 60 mg/kg; glucose ≥ 250 mg/dL | Local injection with mmu_circ_Klhl8_EPCs or SIRT5 inhibitor (MC3482, 5 mg/kg/day) | Days 0, 7, and 14 |
|
| 3 | Liang et al. (2022; China) [ | BALB/c mice (Male; | Full‐thickness dorsal wound (size: NA) | STZ 200 mg/kg; glucose ≥ 250 mg/dL | UCMSC‐derived exosomes (exo‐circHIPK3) vs. exo‐vector vs. vehicle (DPBS) | Days 0, 3, 7, 14 |
|
| 4 | Wang et al. (2023; China) [ | BALB/c mice (NA) | 4‐mm full‐thickness dorsal wound | STZ 60 mg/kg; glucose ≥ 250 mg/dL | ADSC exosomes overexpressing circ_Astn1 (mmu_circ_0000101) vs. PBS (200 μg/100 μL) | Days 7, 14 |
|
| 5 | Shi et al. (2022; China) [ | C57BL/6 mice (sex: NA; | 4‐mm full‐thickness wound | STZ 60 mg/kg i .p.; glucose ≥ 250 mg/dL | Hypoxia‐pretreated ADSC exosomes (circ‐Snhg11) vs. PBS | Days 7, 14 | Downregulation of |
| 6 | Wang et al. (2021; China) [ | BALB/c mice (NA) | 4‐mm full‐thickness wound | STZ 60 mg/kg; glucose ≥ 250 mg/dL | Hypoxia‐treated ADSCs vs. normoxic ADSCs | 21 days | Hypoxia upregulated |
| 7 | Tang et al. (2024; China) [ | C57BL/6 mice (Male; | 4‐mm full‐thickness wound | STZ 60 mg/kg i .p.; glucose ≥ 250 mg/dL | ADSC‐Exos vs. hypoxic Exos (HExos) ± circRNAs (circ_Erbb2ip, circ_0000613, circ_0001490) | Days 7, 14 |
|
| 8 | Chen et al. (2022; China) [ | BALB/c mice (Sex: NA; | Square dorsal foot wound (1 × 1 cm) | STZ 45 mg/kg × 5 days; glucose > 16.7 mM × 10 days | Diabetic BMSC‐Exos vs. OE‐circ_ITCH‐Exos (10 mg/kg, 100 μL) | Days 3, 7, 13 |
|
| 9 | Liu et al. (2024; China) [ | NA (Sex: NA; DFU models with 30 mice and normal mice with 12 ones) | 8‐mm full‐thickness wound | High‐fat/sugar diet × 1 month + STZ i.p.; glucose > 16.7 mmol/L × 2 weeks | Topical drip of | 12 days |
|
| 10 | Liang et al. (2022; China) [ | BALB/c mice (male; | 4‐mm full‐thickness wound | STZ 45 mg/kg + high‐fat diet; glucose ≥ 16.7 mmol/L | ADSC exosomes with | Days 3, 7, 14 |
|
| 11 | Wang et al. (2024; China) [ | C57BLKS/J db/db diabetic mice (male; | 1‐cm full‐thickness wound | STZ + high‐fat diet; glucose ≥ 16.7 mmol/L | MSC‐Exos‐oe‐c | Days 0, 3, 7, 13 |
|
| 12 | Li et al. (2025; China) [ | Sprague–Dawley rats (male; | 2 × 5 mm rectangular dorsal wound | STZ 50 mg/kg × 5 days + high‐fat diet; glucose > 250 mg/dL | Tail‐vein injection of lentivirus (1 × 108 UT/50 μL) for sh‐ | 14 days |
|
| 13 | Wang et al. (2025; China) [ | BALB/c mice (NA) | 4‐mm full‐thickness wound | STZ 60 mg/kg; glucose > 250 mg/dL | ADSC‐Exos vs. HExos vs. | Days 0, 7, 14 |
|
| No. | Authors (year; country) | Dataset(s) | circRNAs/miRNAs | Major findings |
|---|---|---|---|---|
| 1 | Liao et al. (2020; China) [ |
| Identified top DFU biomarkers (AUC > 0.8): | |
| 2 | Zeng et al. (2022; China) [ |
| Key DFU axes: |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCircular RNAs in diseases · Diabetic Foot Ulcer Assessment and Management · Connective Tissue Growth Factor Research
Introduction
1
Diabetic foot ulcers (DFUs) are among the most disabling consequences of diabetes mellitus (DM), defined by non‐healing ulcers that extend into the subdermis and ankle joint [1]. Globally, DFUs affect approximately 19%–34% of diabetic patients, equivalent to nearly 18.6 million people, of whom 50%–60% develop wound infections and approximately 20% result in lower limb amputation [2, 3]. The mortality rate for people with DFUs is 231 deaths per 1000 person‐years, which is much higher than the rate for people with diabetes who do not suffer from DFUs [3]. These numbers reveal that DFUs are a huge public health issue that makes life worse for the sufferers.
These ulcers arise from a complex interaction of factors, including neuropathy, ischemia, metabolic disorders, impaired angiogenesis, and chronic inflammation [1, 4, 5]. Hyperglycemia disrupts endothelial homeostasis, reduces perfusion, and induces ischemic injury through activation of protein kinase C (PKC) [6, 7]. Neuropathy causes sensory and motor impairments in patients, leading to altered foot and wound biomechanics [8]. In addition, hyperglycemia‐induced immune dysfunction delays wound healing by impairing neutrophil activity, inducing advanced glycation end products (AGEs), and slowing extracellular matrix (ECM) remodelling [1, 4, 5]. Although these mechanisms are well documented, the molecular regulatory networks that impair healing in DFUs are still poorly understood. Furthermore, traditional biomarkers, such as inflammatory proteins (procalcitonin, pentraxin 3, interleukins (IL‐6, IL‐18, IL‐20, IL‐22, IL‐24), and C‐reactive protein (CRP)), metabolic markers (arginine, leucine, and isoleucine), and microbial profiles (gram‐positive cocci and anaerobic pathogens), lack sufficient sensitivity or specificity for early diagnosis or monitoring of treatment responses to this disease [9, 10, 11]. Therefore, the identification of novel molecular signatures with diagnostic and therapeutic potential is a key unmet need in the management of DFU.
Circular RNAs (circRNAs) are a recently discovered class of non‐coding RNAs. These molecules form covalently closed loops that make them highly resistant to exonucleases [10, 11]. These molecules generally act as sponges for microRNAs (miRNAs), regulate mRNA stability and transcription, and interact with RNA‐binding proteins (RBPs) to influence post‐transcriptional gene expression. Recent studies have demonstrated their importance in processes involved in wound healing, including angiogenesis, inflammation, oxidative stress, and apoptosis [10, 11]. In addition to diabetes, circRNAs have been implicated in many other multifactorial diseases, such as cancer, cardiovascular disease, and neuropathy. This finding suggests that these genetic molecules can participate in the regulation and manipulation of diseases by forming regulatory circRNA‐miRNA‐mRNA networks [12, 13, 14, 15]. Therefore, they may be good targets for the diagnosis, monitoring, and treatment of diseases, especially since they are highly stable and have tissue‐specific expression [10, 11, 16]. However, despite the large amount of experimental and computational data in this area, we still do not know exactly how circRNAs function as biomarkers and therapeutic targets for DFUs.
Rationale and Objectives
1.1
Because DFUs have a high incidence of morbidity and death, with rates of 5% at 1 year and 42% at 5 years [17], it is very important to provide trustworthy diagnostic tools and effective treatment plans for them. In this context, circRNAs may help us learn more about the molecular causes of DFUs and provide us with chances to diagnose them early and treat them quickly and effectively. Therefore, this scoping review brought together all the information from clinical, preclinical, and in silico investigations on circRNAs in this field. The goal was to (i) find circRNAs that can be used to diagnose or predict diabetes, (ii) explain how they work in diabetic wound healing, and (iii) point out possible circRNA‐based therapy targets for future study.
Methodology
2
Protocol and Registration
2.1
This scoping review was conducted in accordance with the PRISMA Extension for Scoping Reviews (PRISMA‐ScR) [18]. The review protocol followed five key stages: (i) identifying the research question, (ii) identifying relevant studies, (iii) study selection, (iv) charting the data, and (v) collating, summarising, and reporting the results. The study protocol was prospectively registered in the International Prospective Register of Systematic Reviews (PROSPERO) under the registration number of CRD42024586997 (https://www.crd.york.ac.uk/prospero/display_record.php?RecordID=586997).
Research Question
2.2
The present scoping review aimed to investigate the significance of circRNAs as diagnostic/differentiation (in clinical studies), therapeutic targets (in preclinical studies), and candidate discovery (in in silico studies).
Search Strategy and Databases
2.3
A systematic literature search was performed on July 16, 2025, across MEDLINE (via PubMed), EMBASE, ISI Web of Science, and Google Scholar databases. The search was not restricted by publication date to ensure inclusivity of all relevant studies. Both MeSH and non‐MeSH keywords were used in various Boolean combinations. The detailed search strategies for each database are provided in Table 1. To ensure comprehensiveness, the reference lists of all included studies and relevant reviews were manually screened for additional eligible articles. Duplicate records were removed using EndNote 21, and screening was independently performed by two reviewers according to the following inclusion and exclusion criteria.
Eligibility Criteria
2.4
The target population consisted of human patients with DFUs and/or experimental mice or rat models of DFU. Eligible clinical and preclinical studies assessed circRNAs in wound tissues or serum profiling. In silico studies were also included if they reported circRNA‐miRNA‐mRNA regulatory networks. Comparators consisted of healthy or non‐DFUs, untreated or treated animal models of DFU, or computational controls obtained from publicly accessible transcriptome datasets. The outcomes of interest encompassed the diagnostic, mechanistic, and therapeutic functions of circRNAs. Acceptable study designs encompassed observational and experimental clinical studies evaluating circRNA expression or function in DFU patients, preclinical animal studies examining targeted circRNA interventions, and in silico studies delineating regulatory networks pertinent to DFU pathogenesis.
We excluded studies that were irrelevant to both DFUs and circRNAs. Studies were also excluded if they did not include primary data (e.g., reviews, comments, or editorials). This thorough approach ensured that research that may shed light on the function of circRNAs in DFUs was all included.
Data Extraction
2.5
A standardised data extraction form was developed in Microsoft Excel, pilot‐tested on three randomly selected studies, and refined before final data collection. The following data were extracted:
- − Bibliographic details: First author, year of publication, country of origin.
- − CircRNA characteristics: corresponding miRNAs, expression trends, molecular pathways, and functional roles.
- − Clinical study parameters: Study design, patient population, sample size, DFU grade, and inclusion/exclusion criteria.
- −In vivo experimental details: Animal model characteristics, diabetes induction method, ulcer creation protocol, therapeutic interventions, and wound‐healing outcomes.
- − In silico dataset features: GEO accession numbers, dataset types, differential expression analyses, and predicted molecular interaction networks.
Data Charting and Synthesis
2.6
Data were charted independently by three authors using a standardised template. Any disparities were resolved by consulting with a supervisor. No meta‐analysis or bias assessment was performed due to methodological heterogeneity. To ensure methodological clarity, clinical, preclinical, and in silico data were extracted and summarised separately in Tables 2, 3, 4. Studies that initially evaluated circRNA expression using the Gene Expression Omnibus (GEO) datasets and subsequently performed validation in human clinical samples were presented in the clinical section (Section 3.2). In contrast, studies limited to bioinformatic analyses without experimental or clinical verification were reported separately in the in silico section (Section 3.4).
Results
3
Systematic Search Results
3.1
A total of 195 records were retrieved from PubMed, Scopus, Embase, and Web of Science. After removing 77 duplicates, 118 studies were screened by title and abstract, of which 32 were subjected to full‐text assessment. Following the inclusion and exclusion criteria, 22 studies were included in this scoping review: 7 clinical, 13 preclinical, and 2 in silico. Details of the screening and selection process are provided in Figure 1.
A graphical illustration detailing the processes involved in selecting literature.
Clinical Findings (Diagnostic and Differentiation Roles of circRNAs)
3.2
Seven clinical trials published from 2020 to 2025 were identified (Table 2) [19, 20, 21, 22, 23, 24, 25]. We aimed to characterise circRNAs in diverse biological samples, including serum, tissue biopsy, and exosomal samples. Different studies used a variety of profiling techniques, including high‐throughput sequencing, microarray screening, and quantitative RT‐PCR validation. Studies varied in terms of inclusion criteria and control group selection, and most compared DFU tissues with healthy or non‐DFU diabetic wound tissues [19, 20, 21, 22, 23, 24, 25].
Wang et al. [19] found that hsa_circ_0084443 (circ_PRKDC) was a crucial transcript that was expressed differently in patients with DFUs. Functionally, circ_PRKDC was discovered to influence keratinocyte proliferation and migration by modulating key signalling pathways, including Janus kinase (JAK)/Signal transducer and activator of transcription (STAT), Epidermal growth factor receptor (EGFR), Phosphatidylinositol 3‐kinase (PI3K), Extracellular signal‐regulated kinase (ERK), and HIF‐1. Expression of circ_PRKDC was lower in normal wounds compared with intact skin but significantly elevated in DFUs. This data indicated its involvement in compromised wound healing and persistent ulceration [19]. Similarly, Han et al. [24] also validated the regulatory function of circ_PRKDC in epidermal regeneration. Thus, inhibiting circ_PRKDC facilitated wound healing by reducing miR‐31 levels, which subsequently elevated the production of Matrix metalloproteinases 2 & 9 (MMP‐2 and MMP‐9). This mechanism made keratinocytes migrate better. Furthermore, miR‐31 was shown to target Fibrillin 1 (FBN1). In fact, overexpressing FBN1 largely undid the effects of knocking down circ_PRKDC. The dynamic modulation of circ_PRKDC throughout the phases of wound healing (downregulated on Days 1 and 7 relative to undamaged skin) indicated its role in the temporal coordination of wound closure [24].
Tian et al. [21, 22] conducted two complementary studies to demonstrate increased levels of circRNA_072697 in DFU tissues. The findings indicate that circ_072697 acted as a molecular sponge for miR‐3150a‐3p. This event led to the overexpression of lysine demethylase 2A (KDM2A) to stop keratinocyte growth and movement. Their previous bioinformatics study (2020; GSE114248 dataset) had already linked this mechanism to the Kirsten rat sarcoma viral oncogene homologue (KRAS)/mitogen‐activated protein kinase (MAPK) signalling cascade [21, 22]. Fu et al. [20] looked at the same GEO dataset again and showed that circRNA_080968, circRNA_081069, and circRNA_100,980 were all higher in DFU tissues than in NHWs and non‐DFU diabetic controls. Functional predictions indicated that circRNA_080968 interacted with miR‐326 and miR‐766‐3p. This function, in turn, reduced keratinocyte migration while stimulating cell proliferation [20].
Chen et al. [23] also combined clinical validation and data mining of the GEO dataset to evaluate circulating circRNAs in DFUs. Two transcripts, hsa_circ‐0000907 and hsa_circ‐0057362, exhibited considerable dysregulation in DFU tissues and serum. Interestingly, they demonstrated negative relationships with the ankle‐brachial index (ABI) and transcutaneous oxygen tension (TcPO_2_), both clinical indicators of tissue perfusion. These results underscored their clinical diagnostic significance and viability as serum or exosomal biomarkers for the noninvasive evaluation of DFUs [23]. Bai et al. [25] recently identified hsa_HLA_DRB1 as a DFU‐associated circRNA by several prediction pipelines, including Find_circ, CircRNA_finder, CIRCexplorer2, and CIRI2. They hypothesized that this circRNA modulated the expression of miR‐12,118/FLT‐1 (VEGFR‐1), which was associated with inflammation and angiogenesis [25].
Preclinical Studies (Therapeutic Target Identification)
3.3
A total of thirteen preclinical studies using mouse and rat models of DFUs were identified (Table 3) [26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38]. Most of the studies induced diabetes via an intraperitoneal injection of streptozotocin (STZ; 60 mg/kg, citrate buffer, pH 4.5) and confirmed hyperglycemia as a tail vein glucose level more than 250 mg/dL. Diabetic foot ulcers were typically created using 4‐mm full‐thickness punch biopsies of the hind paw or hindlimb. The reviewed studies investigated a variety of circRNA‐based interventions, broadly categorised as (i) exosome‐mediated circRNA delivery, (ii) hypoxia‐preconditioned circRNA‐enriched exosomes, (iii) nanoparticle‐assisted circRNA delivery, and (iv) circRNA silencing approaches (Figure 2).
This figure summarises circRNAs identified as therapeutic or pathogenic targets in animal models of diabetic foot ulcers. Protective circRNAs delivered via exosomes, hypoxia‐preconditioned exosomes, or lipid nanoparticles promote angiogenesis, autophagy, and oxidative stress resistance. In contrast, silencing of pathogenic circRNAs such as circ_0005654 mitigates ferroptosis and inflammation.
Exosome‐Mediated circRNA Delivery
3.3.1
A lot of research has been done on the idea that stem cell exosomes with certain circRNAs may accelerate tissue restoration in DFU models. For example, the exosomal mmu_circ_0000250 reduced local inflammation by sequestering the pro‐inflammatory miR‐128‐3p. This led to the activation of Sirtuin‐1 (SIRT1) and LC3, thereby facilitating autophagy and angiogenesis [26]. Similarly, circHIPK3‐enriched exosomes inhibited miR‐20b‐5p. This resulted in the mRNA upregulation of nuclear factor erythroid 2–related factor 2 (Nrf2), vascular endothelial growth factor A (VEGF‐A), and transforming growth factor beta 1 (TGF‐β1). Mechanism‐wise, it effectively attenuated oxidative stress while promoting vascular repair and wound closure [28]. Moreover, adipose‐derived stem cell (ADSC)‐derived exosomes overexpressing mmu_circ_0001052 demonstrated a strong pro‐healing phenotype by sequestering miR‐106a‐5p. This mechanism activated the fibroblast growth factor 4 (FGF4)/VEGF‐A/p‐p38 signalling cascade [35]. Simultaneously, the downregulation of mmu_circ‐ITCH was reinstated in DFU tissues using exosomal delivery. This intervention improved angiogenesis in human umbilical vein endothelial cells (HUVECs) via activating the Nrf2 and TAF15 signalling pathways [33]. Additionally, circ‐Astn1‐overexpressing ADSC exosomes protected endothelial progenitor cells (EPCs) from apoptosis under hyperglycemic conditions. This was attributed to the inhibition of miR‐138‐5p. This was able to upregulate SIRT1 and Forkhead box O1 (FOXO1), thereby facilitating angiogenesis and epidermal regeneration [29]. Mesenchymal stem cell (MSC)‐derived exosomes containing circMYO9B dramatically improved wound healing by inhibiting CBL and increasing Lysine‐specific demethylase 1A (KDM1A) and VEGF‐A levels. This allowed new blood vessels to grow, granulation tissue to develop, and the epithelium to heal. In contrast, inhibiting circMYO9B negated these beneficial effects [36].
Hypoxia‐Preconditioned circRNA–Enriched Exosomes
3.3.2
Preconditioning stem cells with hypoxia has been shown to be a significant enhancer of the effectiveness of circRNA‐mediated exosomal therapies. For instance, mmu_circ‐Gcap14, which was increased in ADSCs during hypoxia, was shown to block miR‐18a‐5p. This process augmented hypoxia‐inducible factor 1 alpha (HIF‐1α) and VEGF levels, which together promote angiogenesis in DFU models [31]. Similarly, mmu_circ‐Snhg11, which was downregulated under hyperglycemia, was restored by hypoxic exosome treatment. This restoration increased HIF‐1α and induced M2 macrophage polarisation through inhibition of miR‐144‐3p, facilitating VEGF‐mediated angiogenesis [30]. It was demonstrated that hypoxia exposure modified exosomal circRNA profiles by diminishing the expression of mmu_circ‐0000495 while augmenting mmu‐circ‐0000495, mmu‐circ‐0000613, and mmu‐circ‐0001490. They collectively inhibited miR‐670‐5p and enhanced Nrf1 expression, resulting in enhanced oxidative resistance and tissue regeneration [32]. Similarly, mmu_circ‐0001747 was found to be highly expressed in hypoxia‐preconditioned ADSC‐derived exosomes and promoted wound healing through miR‐199a‐5p/HIF‐1α signalling, leading to reduced oxidative stress, apoptosis, and improved angiogenesis [38].
Lipid Nanoparticle (LNP)–Mediated circRNA Delivery
3.3.3
Targeted molecular delivery via nanoparticles has also been studied. For instance, it was examined by Liu et al. [34] how VEFG‐A circRNA transported by lipid nanoparticles performed therapeutically. Topically applied VEFG‐A circRNA‐modified (A‐LNP) or unmodified (U‐LNP) nanoparticles were used by the researchers to treat animal models of DFUs. All wounds in the group treated with A‐LNP had completely healed by the 12th day [34].
circRNA Silencing Approaches
3.3.4
Several studies have also shown pathogenic circRNAs that, when blocked, help wound healing. For example, overexpressing mmu_circ‐Klhl8 in EPCs stopped miR‐212‐3p, which in turn increased SIRT5, started autophagy, and encouraged angiogenesis [27]. However, silencing mmu_circ_0005654, a pathogenic circRNA, markedly decreased iron buildup, inflammatory infiltration, and oxidative damage. This mechanism was accomplished by downregulating insulin‐like growth factor 2 mRNA‐binding protein 2 (IGF2BP2). Thanks to this alternation, the expressions of glutathione peroxidase 4 (GPX4) and solute carrier family 7 member 11 (SLC7A11), markers of ferroptosis resistance, were restored, while inflammatory cytokines (IL‐1β, TNF‐α, IL‐6) were suppressed. The overexpression of IGF2BP2 counteracted these effects, therefore validating the regulatory role of the circ_0005654/IGF2BP2 axis in the aetiology of DFUs [37].
In‐Silico Studies (Candidate Discovery)
3.4
To further understand the pathophysiology of DFU, in silico investigations have shed light on the regulatory interaction networks underlying circRNA, miRNA, and mRNA. According to Table 4, two important bioinformatics research studies were found in this regard [39, 40].
Liao et al. [39] merged circRNA (GSE114248) and miRNA (GSE84971) expression datasets with diabetes‐related miRNAs from the Human MicroRNA Disease Database (HMDD v3.0). Their differential expression revealed hsa_circ‐0089761 as a key circRNA anticipated to control inflammation‐associated miR‐146a‐5p. Following Receiver operating characteristic (ROC) and Gene Ontology (GO) semantic similarity studies, BCL2, CCND1, IRAK4, SMAD4, SP1, and SUFU were identified as promising biomarkers with good discriminating power (AUC > 0.8). SMAD4, SUFU, and IRAK4 showed 100% sensitivity and specificity in distinguishing DFU diabetic tissues from non‐DFU diabetic tissues. SMAD4 was the best biomarker associated with the hsa_circ‐0089761/miR‐146a‐5p regulatory axis, implicating this pathway in DFU‐associated inflammatory and apoptotic responses [39].
Zeng et al. [40] evaluated the GSE114248, GSE84971, GSE68185, and GSE80178 transcriptome datasets to investigate circRNA, miRNA, and mRNA co‐expression networks in DFU, diabetic (non‐DFU) tissues, and healthy tissues. They found that hsa‐circ‐0049271 and hsa‐circ‐0074559 were linked to the growth of DFUs. Functional enrichment tests demonstrated that the VEGF receptor and T cell signalling pathways exhibited differences in DFU tissues compared to other groups. Particularly, the hsa‐circ‐0049271/hsa‐miR‐24‐3p/JUNB axis regulated infectious inflammation and angiogenesis during diabetic wound development [40].
Discussion
4
This study comprehensively examines the relationship between clinical, preclinical, and computational findings on the regulatory role of circRNAs in DFUs. By constructing circRNA‐miRNA‐mRNA interaction networks, we identified specific groups of circRNAs that modulate four relevant and essential biological processes for wound healing: angiogenesis, inflammation, oxidative stress response, and fibroblast‐keratinocyte communication (Figure 3). The expression levels of circRNAs, whether increased or decreased, suggest that these molecules may have both detrimental and beneficial effects on the development of DFUss.
This figure depicts circRNAs identified in clinical, preclinical, and in silico studies of diabetic foot ulcers, categorised based on their role in four key biological processes: Angiogenesis, inflammation, ferroptosis/oxidative stress, and fibroblast‐keratinocyte communication/re‐epithelialization.
Angiogenesis and circRNA Regulation
4.1
Impaired angiogenesis is implicated in the development of DFUs, particularly through pathways involving HIF‐1α and VEGF (Figure 4). Hypoxia makes HIF‐1α more unstable and rapidly degraded by actions that are dependent on the prolyl hydroxylase (PHD) domain [41]. Increased methylglyoxal (MGO) in diabetes makes it more difficult for HIF‐1α to dimerize and simultaneously bind to the co‐activator (p300) [41, 42, 43]. This is due to its inhibition of the carboxyl‐terminus of Hsp70‐interacting protein (CHIP)‐mediated ubiquitination of HIF‐1α [42]. This reduces the expression of HIF‐1α target genes, including VEGF, GLUT1, and EPO, which slows wound healing and inhibits the growth of new blood vessels [41].
CircRNAs that either promote or inhibit angiogenesis in DFUs through regulation of HIF‐1α and VEGF‐related pathways.
Our study identified several proangiogenic circRNAs that work possibly through this mechanism. They include circHIPK3, circ_Astn1, circ_Gcap14, circ_Erbb2ip, circ_0001052, circMYO9B, and circ_0001747 [28, 29, 31, 35, 36, 38]. These molecules help endothelial cells grow, move, and form tubes. They also restore angiogenesis through the miR‐20b‐5p/VEGF‐A, miR‐18a‐5p/HIF‐1α, and miR‐670‐5p/Nrf1 pathways [28, 31, 32]. Circ_Astn1 and circMYO9B increased VEGF‐A by activating SIRT1 and KDM1A, which in turn accelerated wound healing and increased blood flow in diabetic mice [29, 36].
In contrast, some circRNAs were found to inhibit angiogenesis or promote cell death. For example, circ_072697 suppresses endothelial activity through the miR‐3150a‐3p/KRAS–MAPK axis [21, 22], while circ_0005654 induces ferroptosis, a type of cell death dependent on iron accumulation and inflammation, through mechanisms mediated by IGF2BP2 [37]. Taken together, DFUs result in impaired angiogenesis by loss of proangiogenic circRNAs and overexpression of inhibitory circRNAs. Therapeutically, increasing proangiogenic circRNAs or blocking ferroptosis‐related circRNAs may help restore vascular homeostasis and promote wound healing.
Oxidative Stress and Ferroptosis
4.2
Chronic hyperglycemia increases the production of reactive oxygen species (ROS) and mitochondrial dysfunction, leading to ferroptosis cell death (Figure 5) [37]. However, several circRNAs help cells deal with oxidative stress. For example, circ_Erbb2ip and circ_Klhl8 enhance mitochondrial stability and antioxidant defence via the miR‐670‐5p/Nrf1 and miR‐212‐3p/SIRT5 pathways, respectively [27]. Circ_0000250 also stimulates autophagy via miR‐128‐3p, which accelerates angiogenesis and decelerates apoptosis [26]. In contrast, circ_0005654, which is more active in diabetic tissues, increases oxidative damage and ferroptosis more severely [37]. Exosomal circ_ITCH, however, which is sustained by TAF15, inhibits these effects by preventing ferroptosis and facilitating the regeneration of blood vessels [33]. These findings indicate that the exosome‐mediated production of beneficial circRNAs may represent an innovative approach to mitigate oxidative damage in this condition [26, 28, 29, 30, 31, 32, 33, 35, 36, 38].
CircRNAs involved in the regulation of oxidative stress and ferroptosis in diabetic foot ulcers. Protective circRNAs increase mitochondrial stability, enhance autophagy, reduce reactive oxygen species (ROS), inhibit ferroptosis, and support angiogenesis. In contrast, circ_0005654 acts as a deleterious circRNA by promoting ferroptosis and iron accumulation, exacerbating oxidative damage and impairing wound healing.
Nrf2, which regulates antioxidant enzymes, is a crucial link between oxidative stress and angiogenesis [44]. In DFUs, Nrf2 levels are significantly reduced, partly due to aberrant histone deacetylase (HDAC) signalling and altered DNA methylation [45]. Decreased Nrf2, VEGF, HIF‐1α, and SDF‐1α, along with increased TSP‐2, indicate a disruption in the redox‐angiogenic balance. Furthermore, differences in HDAC and SIRT expression (with HDAC4 showing an inverse correlation with Nrf2) indicate epigenetic remodelling of DFU tissue. This HDAC4‐Nrf2‐angiogenesis axis may be a critical target for future therapies [45].
Iron overload exacerbates oxidative stress and inflammation, worsening tissue necrosis and vascular damage [46, 47]. Iron accumulation may also cause diabetic peripheral neuropathy, which can cause loss of sensation in DFUs [6]. Ferroptosis also induces axonal degeneration and nerve dysfunction in diabetic animal models [48]. However, therapy with liproxstatin‐1 sped up wound healing by stopping ferroptosis and inflammation [37]. These results show that circRNA‐controlled ferroptosis, oxidative stress, and angiogenesis are all closely related. This forms a molecular trio that might lead to new ways to treat persistent wounds that won't heal.
Inflammation‐Associated circRNA Networks
4.3
Inflammation plays a dual role in DFU, initially protective but becoming detrimental when it becomes chronic [49]. CircRNAs help fine‐tune this balance. For example, circ_HLA‐DRB1 is higher in DFU and helps the immune system work better and change the shape of blood vessels via the miR‐12,118/FLT‐1 axis [25]. FLT‐1, a receptor for VEGF, is very important for angiogenesis when there is inflammation. Clinical studies show that patients with DFU have higher levels of sFlt‐1, advanced oxidation protein products (AOPPs), malondialdehyde (MDA), TNF‐α, and VEGF, indicating a disruption in the angiogenesis‐inflammation interface [50]. Under these conditions, neutrophils are the main producers of VEGF and induce FLT‐1 expression, suggesting that hyperglycemia disrupts the VEGF/FLT‐1 balance [51]. Similarly, mast cell activation and increased FLT‐1 have been associated with impaired angiogenesis and tissue viability in diabetic mice [52].
Meanwhile, circ_Snhg11 increases HIF‐1α expression and promotes macrophage polarisation from proinflammatory (M1) to repair (M2), supporting inflammation resolution and matrix remodelling [30]. In contrast, circ_0089761 and circ_0049271 may worsen inflammation by silencing miR‐146a‐5p and miR‐24‐3p and disrupting genes such as SMAD4, BCL2, CCND1, IRAK4, and JUNB [39]. Among these, SMAD4, a central mediator of the TGF‐β/Smad pathway, drives M2 macrophage activation, ECM synthesis, and wound closure [53]. Thus, the circRNA‐miRNA‐TGF‐β axis emerges as a critical mechanism in regulating inflammation and tissue remodelling in DFU.
Fibroblast–Keratinocyte Communication and Re‐Epithelialization
4.4
For re‐epithelialization to occur properly, fibroblasts and keratinocytes must work together well. In diabetic wounds, disruption of this interaction leads to impaired wound closure [54]. Recent studies suggest that circRNAs have a regulatory function in this process. In DFUs, increased levels of hsa_circ_PRKDC and hsa_circRNA_080968 can increase keratinocyte activity through the miR‐31/FBN1/MMP2/MMP9 and miR‐326/miR‐766‐3p pathways [19, 20, 24]. This also activates downstream signalling pathways such as JAK/STAT, EGFR, PI3K/ERK, and HIF‐1. This leads to successful re‐epithelialization and matrix remodelling [19, 20, 24]. The ECM protein fibronectin is also crucial in this process [55]. Its continuous presence supports the entry of latent TGF‐β binding protein 1 (LTBP‐1) and latent TGF‐β into the ECM and enables fibroblast‐to‐myofibroblast differentiation through TGF‐β1‐dependent signalling [56]. The EDA domain in FN enhances LTBP‐1 binding and is essential for the acquisition of the contractile phenotype of myofibroblasts, which is essential for tissue regeneration [57, 58]. However, chronic hyperglycemia in diabetes induces excessive production of ROS, which leads to fragmentation of FN and the formation of AGEs [55, 59]. These changes not only disrupt fibroblast adhesion and morphology, but also induce sustained inflammation by stimulating neutrophil recruitment and macrophage activation [60]. The formation of FN fragments is induced in diabetic conditions, which, unlike intact FN, increases the release of MMPs and the production of inflammatory cytokines [61]. As a result, the ECM in diabetic wounds is not properly organised, making it difficult for cells to move and for blood vessels to deform [57, 58].
In addition, the researchers found that circ_072697, one of the downregulated circRNAs, suppresses keratinocyte growth by exploiting the miR‐3150a‐3p/KDM2A/KRAS pathway [21, 22]. If this regulatory circuit is disrupted, keratinocyte growth can become more difficult, and the wound matrix may not be sufficiently stable [21, 22]. The KDM2A gene encodes an F‐box protein that acts as a histone demethylase. This means that it plays a role in managing metabolism and inflammation [21]. Tanaka et al. [62] showed that metformin activates KDM2A through AMPK signalling, which prevents rapid cell growth. KDM2A is also involved in the inflammatory responses of keratinocytes in psoriasis [63]. This means that if it does not work properly, it may worsen inflammation in DFU and make it more difficult for epithelial cells to regrow [21]. However, the precise molecular mechanisms of KDM2A‐mediated circRNA regulation in diabetic wounds require further elucidation.
Methodological Limitations
4.5
Despite the growing interest, current evidence remains limited by several factors. First, small sample sizes and the lack of longitudinal designs in clinical studies limit generalizability and causal inference. Second, methodological heterogeneity, such as varying biological sample sources in terms of age, sex, and race, normalisation pipelines, and validation platforms, complicate cross‐study comparisons. Third, no randomised clinical trials (RCTs) evaluating circRNA‐based interventions were identified, emphasising the early stage of development of this field. Integrating a multi‐omics approach is essential to elucidate broader regulatory networks involving circRNAs.
Preclinical studies have shown promising results for the therapeutic application of exosome‐ or nanoparticle‐mediated circRNA delivery. However, several translational challenges persist. They include immunogenicity concerns, difficulties in large‐scale production, and the absence of Good Manufacturing Practice (GMP)‐grade circRNA formulations suitable for clinical use.
In silico network analyses have complemented experimental data by identifying key regulatory circuits; nonetheless, a considerable gap remains between computational predictions and experimental validation. Most bioinformatic predictions of circRNA‐miRNA interactions have yet to be validated in vivo. To fill this gap, future studies should use RNA immunoprecipitation (RIP) methods and dual‐luciferase reporter assays in DFU tissue models to validate the predicted interactions.
Future Perspectives
4.6
The goal of future research should be to make the ways that circRNA is isolated and profiled more consistent and standardised. This helps improve the reproducibility and repeatability of results. When organising future clinical trials, it is highly crucial to carefully consider the genetic, clinical, and social aspects of the persons who will be researched. This strategy makes it easier to find biomarkers that are effective for many people.
Advanced bioinformatics, machine learning algorithms, and deep neural networks could also help build accurate and customizable models that can help doctors diagnose DFUs early, track their progression, and monitor patient response. These kinds of advances could usher in a new era of guided tissue regeneration and precision medicine.
Conclusion
5
This review provides significant evidence for the essential role of circRNAs in the aetiology of diabetic foot ulcers. In silico, preclinical, and clinical studies have identified various circRNAs as potential biomarkers for disease severity and therapeutic targets, showing encouraging results in modulating angiogenesis, inflammation, oxidative stress, and fibroblast‐keratinocyte communication through circRNA‐based therapies (Figure 3). Three innovative approaches to targeting DFUs include the use of circRNA‐based nanoparticles, circRNA‐enriched extracellular vesicles, and gene silencing techniques (Figure 2), which provide a summary of circRNAs that have been used for therapeutic purposes in animal models so far. However, further research is required to convert these molecular findings into a clinically viable approach.
Author Contributions
A.R.G.: writing – original draft, investigation, methodology. M.H.: writing – original draft; A.S.M.: searching, visualisation; M.D.: editing the manuscript, data curation; H.C.: writing – review and editing, conceptualization; F.N.: writing – review and editing, supervision, conceptualization. R.A.: writing – review and editing, supervision, conceptualization and methodology.
Funding
The authors have nothing to report.
Ethics Statement
The authors have nothing to report.
Consent
The authors have nothing to report.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1J. M. Raja , M. A. Maturana , S. Kayali , A. Khouzam , and N. Efeovbokhan , “Diabetic Foot Ulcer: A Comprehensive Review of Pathophysiology and Management Modalities,” World Journal of Clinical Cases 11, no. 8 (2023): 1684–1693.36970004 10.12998/wjcc.v 11.i 8.1684 PMC 10037283 · doi ↗ · pubmed ↗
- 2D. G. Armstrong , T. W. Tan , A. J. M. Boulton , and S. A. Bus , “Diabetic Foot Ulcers: A Review,” Journal of the American Medical Association 330, no. 1 (2023): 62–75.37395769 10.1001/jama.2023.10578 PMC 10723802 · doi ↗ · pubmed ↗
- 3K. Mc Dermott , M. Fang , A. J. M. Boulton , E. Selvin , and C. W. Hicks , “Etiology, Epidemiology, and Disparities in the Burden of Diabetic Foot Ulcers,” Diabetes Care 46, no. 1 (2023): 209–221.36548709 10.2337/dci 22-0043 PMC 9797649 · doi ↗ · pubmed ↗
- 4J. Kim , “The Pathophysiology of Diabetic Foot: A Narrative Review,” Journal of Yeungnam Medical Science 40, no. 4 (2023): 328–334.37797951 10.12701/jyms.2023.00731 PMC 10626291 · doi ↗ · pubmed ↗
- 5W. D. Aumiller and H. A. Dollahite , “Pathogenesis and Management of Diabetic Foot Ulcers,” Jaapa 28, no. 5 (2015): 28–34.10.1097/01.JAA.0000464276.44117.b 125853673 · doi ↗ · pubmed ↗
- 6D. Yousif , Z. Yousif , and P. Joseph , “Diabetic Foot Ulcer Neuropathy, Impaired Vasculature, and Immune Responses,” in Diabetic Foot Ulcers‐Pathogenesis, Innovative Treatments and AI Applications (Intech Open, 2024).
- 7Z. Liu , L. Hu , T. Zhang , et al., “PKCβ Increases ROS Levels Leading to Vascular Endothelial Injury in Diabetic Foot Ulcers,” American Journal of Translational Research 12, no. 10 (2020): 6409.33194039 PMC 7653566 · pubmed ↗
- 8K. Parveen , M. A. Hussain , S. Anwar , H. M. Elagib , and M. A. Kausar , “Comprehensive Review on Diabetic Foot Ulcers and Neuropathy: Treatment, Prevention and Management,” World Journal of Diabetes 16, no. 3 (2025): 100329.40093290 10.4239/wjd.v 16.i 3.100329 PMC 11885961 · doi ↗ · pubmed ↗