Facilitative interspecific interactions in marine vertebrates across scales: from individuals to ecosystems
Eduardo Döbber Vontobel, Sophie Smout, Jorge L. Rodrigues Filho, Ronaldo Angelini, Mauricio Cantor, Fábio G. Daura‐Jorge

TL;DR
This paper reviews how beneficial interactions between marine animals, like birds and fish, help ecosystems from individual behavior to whole communities.
Contribution
The paper emphasizes the overlooked role of facilitative interactions in marine ecosystems and suggests integrating them into ecosystem models for better management.
Findings
Facilitative interactions improve foraging success and predator-prey dynamics in marine ecosystems.
Some marine apex predators depend on facilitation for survival and population dynamics.
Including facilitation in ecosystem models could improve fisheries management and conservation efforts.
Abstract
Facilitative interspecific interactions (FIIs) confer benefits to at least one participant without detriment to others. Although often less emphasised than antagonistic interactions in ecological studies, this review highlights the significant ecological role of FIIs across biological scales – from individual behaviours to population, community, and ecosystem‐level effects – with a focus on mobile marine vertebrates such as birds, mammals, and fish. These interactions enhance foraging success, shape predator–prey dynamics and contribute to the structure and function of marine ecosystems. FIIs include diverse associations such as multi‐species aggregations among marine apex predators (e.g. dolphins, seabirds, and surface‐feeding fish), mixed‐species shoals, fish cleaning mutualisms, and cooperative foraging involving predators, including humans. At the population level, FIIs can improve…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Fig. 1
Fig. 1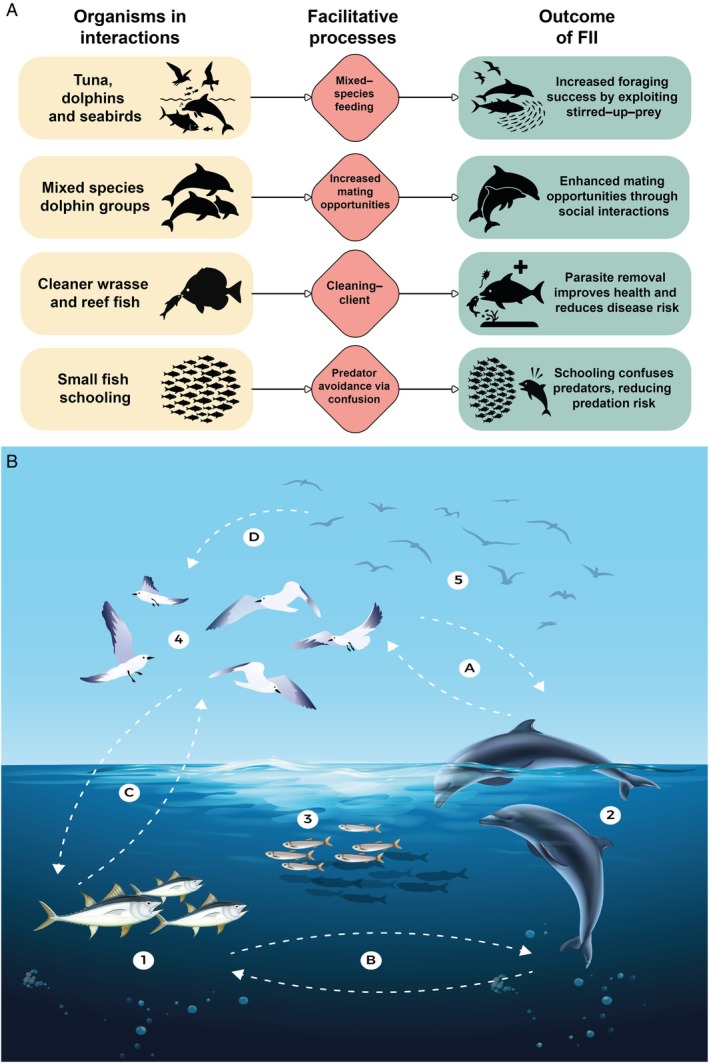 Fig. 2
Fig. 2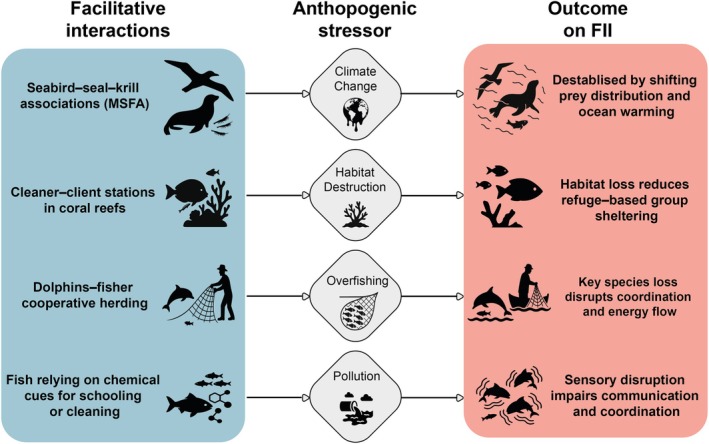 Fig. 3
Fig. 3| Facilitation and ecological interactions | |
| Commensalism | An ecological interaction in which one species benefits while the other experiences no significant positive or negative effect. |
| Facilitative interspecific interaction (FII) | An interaction between species where at least one benefits without causing harm to the other. These interactions can influence community structure, population dynamics, and ecosystem processes. |
| Facultative associations | Ecological interactions where the relationship between species is not essential for their survival or reproduction but still provides some benefit. |
| Facultative mutualism | A mutualistic relationship in which both species benefit but are not entirely dependent on each other for survival. |
| Mutualism | Any ecological interaction between two species in which both individuals experience a net benefit. Mutualistic relationships can involve resource exchange, protection, or other advantages that enhance survival or reproduction. |
| Obligate mutualism | A form of mutualism where both species strictly depend on each other for survival or reproduction. |
| Positive interaction | An ecological interaction where at least one species benefits, including mutualism, commensalism, and facilitation. |
| Strategies and facilitative behaviours | |
| Aggregation | The co‐occurrence of two or more individuals of the same or different species in space and time due to a shared environmental driver, without necessarily involving social or cooperative interactions. |
| Cooperative foraging | A type of cooperative hunting where individuals from the same or different species coordinate their actions to increase hunting success. |
| Follower–nuclear species | A behavioural interaction in which one species (nuclear) is actively followed by another (follower) that benefits from access to resources, information, or protection. |
| Kleptoparasitism | A feeding strategy in which an individual steals food or prey that another individual has captured or is actively hunting. |
| Local enhancement | A behavioural phenomenon in which an individual is attracted to a location based on the presence or activity of another, usually related to foraging or predator avoidance. |
| Mixed‐species feeding association (MSFA) | A specific type of |
| Mixed‐species group (MSG) | Temporary or long‐term group formed by individuals of different species interacting or coexisting within the same ecological community or habitat. These groups may arise for foraging benefits, predator avoidance, social interaction, or mutualistic relationships. |
| Social predation | A form of cooperative hunting where predators work together to locate, capture, and consume prey, improving hunting success and efficiency. |
| Key species and functional roles | |
| Ecosystem engineer species | Species that actively modify their environment, creating, maintaining, or altering habitats in ways that benefit other species. These modifications can be physical (e.g. beavers building dams, corals forming reefs) or biogeochemical (e.g. earthworms altering soil composition). |
| Flagship ecological interaction | A rare, culturally significant, and ecologically relevant interaction between two species, where conserving the interaction itself has broader ecological and social benefits. |
| Keystone facilitator | A species critical in facilitating interactions between other species, significantly influencing community structure and biodiversity. |
| Keystone species | A species that has a disproportionately large impact on its ecosystem relative to its abundance. Keystone species regulate community structure by affecting species interactions such as predation, competition, and facilitation. |
| Structuring species | Species that provide habitat complexity and stability, influencing ecosystem composition and organisation. Unlike ecosystem engineers, structuring species do not necessarily modify their environment but create physical conditions that support other organisms (e.g. kelp forests, mangroves, and mussel beds). |
| Model category | EM examples | EM reference | EM paper | FII description | FII reference |
|---|---|---|---|---|---|
| Mass‐balance | ECOPATH |
| Olson & Watters ( | Tuna, dolphins, and seabirds foraging | Au & Pitman ( |
| Biogeochemical | ATLANTIS |
| Ainsworth | Marine mammals foraging | Bacon |
| Minimum Realistic | MSM | Jurado‐Molina | Jurado‐Molina | Gray whales and seabirds foraging | Grebmeier & Harrison ( |
| MSVPA |
Helgason & Gislason ( Pope ( Sparre ( | Pope & Macer ( | Seabirds foraging | Camphuysen | |
| GADGET |
| Trenkel | Minke whales and seabirds foraging | Anderwald ( | |
| Agent‐based | INVITRO |
| McDonald | Cleaning mutualism | Caves ( |
| Individual‐based | OSMOSE |
| Fu | Marine mammals and seabirds foraging | Pierotti ( |
| Size‐based | MIZER |
| Woodworth‐Jefcoats | Tuna, dolphins and seabirds foraging | Hebshi |
| Bioenergetic | Yodzis & Innes ( |
Yodzis & Innes ( Yodzis ( Koen‐Alonso & Yodzis ( | Koen‐Alonso & Yodzis ( | Pinnipeds and seabirds foraging | Thiebot & Weimerskirch ( |
- —Coordenação de Aperfeiçoamento de Pessoal de Nível Superior10.13039/501100002322
- —National Geographic Society10.13039/100006363
- —Fundação de Amparo à Pesquisa e Inovação do Estado de Santa Catarina10.13039/501100005667
- —Conselho Nacional de Desenvolvimento Científico e Tecnológico10.13039/501100003593
- —Fundação de Amparo à Pesquisa e Inovação de Santa Catarina
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine and fisheries research · Marine animal studies overview · Cephalopods and Marine Biology
INTRODUCTION
I.
Facilitative interspecific interactions (FIIs), or positive interspecific interactions, occur when individuals of one species enhance the overall fitness of at least one participant without causing detriment to the other species (Stachowicz, 2001; Bruno, Stachowicz & Bertness, 2003; Bronstein, 2009). These interactions may significantly impact species distributions, population dynamics (Crowley & Cox, 2011), and community structure (Boucher, James & Keeler, 1982). The spectrum of FII is broad, ranging from casual and facultative associations to commensalism and obligate mutualisms (Bronstein, 1994, 2015; Bruno et al., 2003; Bulleri, 2009; Stachowicz, 2001), with such interactions being widespread across ecosystems. They are observed from the emergence of eukaryotic life forms (Boucher et al., 1982) to intricate relationships between flowers and seed‐dispersing pollinators (Baraza et al., 2007), complex coral reef networks (Stachowicz, 2001), and facilitative foraging among predators (Au & Pitman, 1986). Humans can also be involved, as in the cooperative foraging between honeyguide birds and honey hunters (Spottiswoode, Begg & Begg, 2016) or between wild dolphins and artisanal fishers (Simões‐Lopes, 1998; Cantor, Farine & Daura‐Jorge, 2023).
Historically, ecological research has prioritised antagonistic over facilitative interactions (Bruno et al., 2003). However, in a seminal essay, Gould (1988) shed light on Kropotkin's perspectives on ecological interactions. Drawing from the concept of “mutual aid” pioneered by the German zoologist K. F. Kessler, Kropotkin (1902) emphasised the significance of facilitative interactions, explicitly contrasting them with competitive dynamics. Departing from one of the central tenets of Darwin's The Origin of Species, both Kessler and Kropotkin challenged Darwin's emphasis on competition in the “struggle for existence”, arguing that cooperation could be equally, if not more, important (Todes, 1987). This viewpoint has gained traction with growing interest in the ecological roles of FII across diverse biological scales, from studies of individuals to impacts on whole ecosystems (Bruno et al., 2003; Bulleri, 2009; Ford & Roberts, 2019), encompassing plant–pollinator systems (Gotelli, Graves & Rahbeck, 2010), coral symbioses (Easson et al., 2014), and crustacean–fish partnerships (Vaughan et al., 2017).
These diverse examples highlight not only the ubiquity but also the variety of FIIs in form and function. To understand their ecological consequences, it is useful to distinguish among types of facilitation based on dependency and environmental context. Some species act as ecosystem engineers, physically altering habitats in ways that benefit others (Jones, Lawton & Shachak, 1994). In marine systems, facilitation is often facultative, where one species benefits from another but neither depends on the interaction for survival or reproduction. By contrast, obligate facilitation involves long‐term dependencies, where at least one species relies on the interaction to persist. Facultative interactions are particularly common in dynamic and open environments like the pelagic zone, where species distributions and behaviours often respond to fluctuating resources and conditions. Such associations can be highly context dependent, shifting along a continuum from facilitation to neutral or even antagonistic outcomes depending on ecological pressures (Bruno et al., 2003; Bulleri, 2009; Stachowicz, 2001).
Despite their now‐recognised ecological significance, FIIs have historically received less attention in the scientific literature due to perceptions that they are rare, difficult to demonstrate, or unimportant (Bronstein, 1994, 2009; Bruno et al., 2003). While interest in facilitation has increased, interactions involving highly mobile marine vertebrates remain relatively underexplored. Persistent inconsistencies in defining interactions across the facilitative spectrum (Hoeksema & Bruna, 2015) highlight the need for clear conceptual frameworks and functional glossaries to reduce ambiguity in ecological discourse (e.g. Silknetter et al., 2020; Cram et al., 2022; Crowley & Cox, 2011; Lang & Farine, 2017), particularly in the study of highly mobile organisms in the oceans. Moreover, within the spectrum of FIIs lies a dynamic array of strategies, each characterised by distinct definitions and subtleties. FII can also vary in terms of intentionality, ranging from incidental benefits (i.e. by‐products of behaviour directed at one's own fitness) to behaviours shaped by selection to benefit another species. For example, mobbing behaviour aimed at deterring predators may incidentally benefit multiple prey species, whereas coordinated foraging suggests mutual advantages selected over time. Although this review does not explicitly categorise FII by intentionality, we recognise that such distinctions are relevant to understanding their origins and stability (related terms are clarified in the glossary in Table 1).
In this review, we explore how FIIs among marine vertebrates manifest and propagate across biological scales, from individual behaviours to population processes, community structure, and ecosystem‐level effects. This cross‐scale framing allows us to consider not only the direct benefits between species but also the broader ecological consequences that emerge over space and time. To contextualise, our synthesis draws from species coexistence theory (Chesson, 2000), mutualism ecology (Bronstein, 1994), and ecological network theory (Kéfi et al., 2016). We focus primarily on direct facilitative interactions – behavioural coordination, spatial association, and resource provisioning (e.g. cooperative foraging and cleaning symbioses) – as these are more readily documented in highly mobile species in marine environments. Indirect effects, such as those mediated through habitat modification or food‐web dynamics (Thomsen et al., 2010; Bruno et al., 2003; Bulleri et al., 2016), lie beyond the scope of this review but warrant future analyses.
ANIMAL GROUPS UNDER ANALYSIS
II.
Despite numerous studies documenting FIIs among marine fish (Strand, 1988; Caves, 2021a,b) and marine apex predators (Au & Pitman, 1986; Veit & Harrison, 2017; Fig. 1), it remains unclear how these interactions impact patterns and processes at various biological scales, including their effects on resultant ecosystem services utilised by humans. Marine fish, for instance, are critical resources (Food and Agriculture Organization, 2022; Holmlund & Hammer, 1999). Understanding how FIIs shape their populations and behaviours is vital, as they provide food, support subsistence livelihoods, and underpin the socio‐economic sustainability of fishing communities. Consequently, they are key targets in Ecosystem‐Based Fisheries Management (EBFM; Link, 2010).
Exemplar facilitative interspecific interactions among marine vertebrates. (A) Cooperative fisheries between net‐casting fishers and Tursiops truncatus gephyreus in Laguna, southern Brazil, where dolphins actively herd fish toward fishers' nets, increasing capture success for both fishers and dolphins (source: F.G. Daura‐Jorge). (B) Procellariiform seabirds foraging in association with subsurface pinnipeds in Tramandaí, southern Brazil (source: G. Frainer), where pinnipeds drive prey towards the surface, making them available to seabirds. (C) A joint school of fish from the genera Lutjanus and Haemulon in Cozumel, Mexican Caribbean, where aggregating in a multispecies school enhances predator avoidance and foraging efficiency. (D) Sphyraena barracuda being cleaned by Elacatinus phthirophagus in Fernando de Noronha, Brazil, where the goby removes ectoparasites and dead tissue, benefiting the barracuda while gaining food (source: S.R. Floeter).
Similarly, marine apex predators also play essential roles in regulating marine food webs, impacting species targeted by industrial fishing (e.g. tunas and sharks; Ferretti et al., 2010) and those important to small‐scale subsistence fisheries [e.g. pinnipeds (Hurtubise, 2016; Jensen, Sheehan & MacLean, 2009)]. Clarifying the role of FIIs in their ecology is therefore critical to understanding food‐web structure and dynamics. Moreover, many apex predators such as seabirds and cetaceans are central to EBFM strategies aligned with international biodiversity agreements, such as the Convention on Migratory Species (CMS; https://www.cms.int) and the Convention on International Trade in Endangered Species (CITES; https://cites.org). Understanding how FII influences their life histories and population dynamics is thus crucial for effective conservation planning. In the following sections, we review FII across biological scales, focusing specifically on marine fish, seabirds, and cetaceans, emphasising apex predators. We conclude by summarising key insights that can guide marine species conservation and by identifying future research avenues to address current knowledge gaps.
INDIVIDUAL‐LEVEL FACILITATIVE INTERSPECIES INTERACTIONS
III.
Facilitation at the individual level underpins broader ecological effects, often emerging through behavioural coordination, local enhancement, or social learning. Drawing on niche theory and species coexistence frameworks, which recognise that interspecific behaviours – from incidental associations to structured cooperation – can influence survival, resource use, and fitness, we review cases where facilitation shapes individual behaviour with potential cascading consequences across populations, communities, and ecosystems.
Numerous studies have documented marine apex predators engaging in potential FII with other marine predators (Fig. 2). For example, Bearzi (2006) described behavioural correlations between California sea lions (Zalophus californianus) and three dolphin species (Tursiops truncatus, Delphinus delphis, and D. capensis) during social predation, including diving, travelling, surface‐feeding, and socialising. These correlations were stronger during aggregation events than in short‐term associations, suggesting foraging advantages for individual sea lions. However, dolphin behaviour ranged from cooperative strategies (Gallo‐Reynoso, 1991) to kleptoparasitism and commensalism (Bearzi, 2006), where dolphins opportunistically steal prey or benefit from other species' foraging without direct impact. This illustrates a gradient of interspecific interactions, from antagonistic to neutral or weakly positive facilitation, depending on the ecological context and mutual benefit.
Diversity and complexity of facilitative interspecific interactions (FIIs) in marine ecosystems. (A) Summary of different types of facilitative interactions, showing example taxa, interaction types, and ecological outcomes. These include feeding, mating, cleaning, and predator avoidance, each contributing to survival, reproduction, and ecosystem balance. (B) Visual representation of facilitative foraging interactions among pelagic vertebrates. Numbers represent functional groups: (1) predatory fish (e.g. tuna); (2) marine mammals (e.g. dolphins); (3) forage fish (e.g. sardines); (4) predatory seabirds (e.g. terns); (5) kleptoparasitic seabirds (e.g. frigatebirds). Letters indicate interactions: (A) dolphins and seabirds; (B) dolphins and tuna; (C) seabirds and tuna –which may range from mutualism to competition; (D) local enhancement/follower–nuclear relationships.
Many studies have also evaluated the impact of FII on cetaceans within mixed‐species groups (MSGs) (see Stensland, Angerbjörn & Berggren, 2003; Fig. 2A). Such groups typically consist of individuals seeking advantages through facilitative interactions, potentially improving foraging success or predation protection. Additional benefits include increased chances of reproduction and survival, as well as opportunities for mating practice, playful behaviour, and alloparental care. MSGs may offer advantages over single‐species groups by reducing breeding competition (Dunbar, 1993; Buchanan‐Smith, 1999). These overlapping benefits (Carlson et al., 2023; Goodale, Beauchamp & Ruxton, 2017; Stensland et al., 2003) exemplify FII. For instance, Herzing & Johnson (1997) and Elliser & Herzing (2016) documented affiliative and aggressive behaviours between Atlantic spotted dolphins (Stenella frontalis) and bottlenose dolphins (Tursiops truncatus) in MSGs in the Bahamas. Similarly, Syme, Kiszka & Parra (2021) observed common socialising and travelling in MSGs of Australian humpback dolphins (Sousa sahulensis) and Indo‐Pacific bottlenose dolphins (Tursiops aduncus), with foraging observed in approximately 11% of sightings. They proposed a primary social function for these MSGs, providing social learning and mating opportunities, with antipredator behaviour being secondary. Other studies have documented the frequency of MSGs compared to single‐species groups (Syme, Kiszka & Parra, 2023), highlighting their importance for certain species (Acevedo‐Gutiérrez, 1999; Thompson, 2010; Kiszka et al., 2011).
Cetaceans also frequently interact with seabirds (Evans, 1982), often near predatory fish (Ballance, Pitman & Fiedler, 2006; Clua & Grosvalet, 2001; Veit & Harrison, 2017; Rogan & Mackey, 2007; Guse et al., 2013; Goyert, Manne & Veit, 2014). These interactions typically involve a triadic dynamic where subsurface predators (tunas, dolphins, and whales) corral fish towards the surface, forming prey balls that aid capture for all involved (Au & Pitman, 1988; Veit & Harrison, 2017; Fig. 2). Documented cases include sharks trailing tunas (Au, 1991) and numerous seabird–tuna interactions (Au & Pitman, 1988; Hebshi, Duffy & Hyrenbach, 2008). These range from antagonistic competition and parasitism to facilitative scenarios (Veit & Harrison, 2017; Bronstein, 1994; Stachowicz, 2001) (Fig. 2). Total dependence occurs in some cases, such as between Parkinson's petrels (Procellaria parkinsoni) and melon‐headed (Peponocephala electra) and false killer whales (Pseudorca crassidens) (Pitman & Ballance, 1992). Evans (1982) also details seabird–dolphin associational behaviours, including mass attacks (Clua & Grosvalet, 2001) and prey disruption (Thiebault et al., 2016), which may alleviate foraging challenges and constitute FII (Ashmole & Ashmole, 1967; Au & Pitman, 1986; Ballance et al., 2006; Guse et al., 2013). These temporary or obligate interspecific associations are thought to enhance individual predator fitness (Pitman & Ballance, 1992). However, studying these complex interactions faces methodological challenges inherent in underwater research (Norris & Dohl, 1980; Scott et al., 2012), although recent advancements in acoustics (Ceyrac et al., 2018; McInnes et al., 2024) and biologging (Beltran et al., 2024; Clayton et al., 2023; Hessing et al., 2024) have facilitated progress.
While some facilitative interactions involve large, dynamic aggregations, others are characterised by more direct and structured cooperation, where individuals modulate behaviour for shared goals or mutual benefits (Chase, 1980; Clutton‐Brock, 2009; Dugatkin, 2002; Bronstein & Sridhar, 2024). Such cooperative behaviours include group hunting (Dugatkin, 2002; Bailey, Myatt & Wilson, 2013), cooperative kinship (Smith, 2014; Dugatkin, 2002), cooperative breeding (Hatchwell, 2009), and joint defence (Dugatkin, 2002; Hager & Jones, 2009). A fascinating example is cooperative fishing involving net‐casting fishers and dolphins (Simões‐Lopes, 1998) coordinating to access mullet (Mugil spp.) (Cantor et al., 2023; Fig. 1A). Lang & Farine (2017) analysed similar social predation cases, proposing a multidimensional framework for examining this form of interaction considering communication, sociality, and level of dependency, highlighting the intricate nature of FIIs among marine apex predators, including dolphins (Hebshi et al., 2008; Au & Pitman, 1988) and fish (Bshary et al., 2006).
A common manifestation of these dynamics is local enhancement, where apex predators adaptively use information from conspecifics or other species for foraging (Pöysä, 1992; Buckley, 1996; Fig. 1B), often fitting the follower–nuclear species perspective (Inagaki et al., 2020). This behaviour has been observed across various marine taxa: seabirds [including Procellariiformes (Thiebault et al., 2014) and penguins (Silverman et al., 2004)] as well as cetaceans (Veit & Harrison, 2017), pinnipeds (Bearzi, 2006) and fish [including bony fish (Strand, 1988) and sharks (White et al., 2022)], all of which have been seen following other species to locate prey. These interactions often involve a nuclear species (leader) and follower species exploiting social information for foraging. While followers clearly benefit, nuclear species may also gain advantages like improved prey capture through mass attacks or group foraging disruption. Yet, assessing the magnitude and direction of these benefits remains challenging.
A crucial conceptual challenge is distinguishing true facilitation from basic functional complementarity or accidental co‐occurrence. For instance, dolphin, tuna, and seabird aggregations targeting schooling prey may result from independent responses to shared cues like prey density, lacking active interspecific coordination. Our framework defines facilitation requiring at least one species consistently benefiting from the presence or actions of another beyond chance. This benefit may manifest as improved prey accessibility, reduced search time, or enhanced predator avoidance. While some multi‐species foraging groups may arise from incidental convergence (i.e. non‐intentional co‐occurrence) that leads to facilitative outcomes, others show behavioural adjustments or learning that qualify as truly facilitative.
This continuum is also evident in interspecific mobbing, where one species assists others in deterring predators. For example, humpback whales (Megaptera novaeangliae) defend conspecifics and other species like gray whales (Eschrichtius robustus), Weddell seals (Leptonychotes weddellii), and sea lions from killer whale (Orcinus orca) attacks, an antipredator strategy (Pitman et al., 2017) The motivations behind these interspecific responses remain puzzling, possibly reflecting predator harassment culture or generalised protective behaviour triggered by distress cues. Pitman et al. (2017) suggest that future research should explore whether these responses arise from social learning or innate tendencies shaped by cultural or natural selection. Other examples of marine predator mobbing include Galápagos fur seals (Arctocephalus galapagoensis), Galápagos sea lions (Zalophus wollebaeki), and Australian fur seals (Arctocephalus pusillus doriferus) mobbing sharks (Barlow, 1972; Trillmich, 1996; Dickie, 2005), Steller sea lions mobbing killer whales (Heise et al., 2003), and dolphins mobbing sharks (Essapian, 1953; Connor, 2000).
While the previous examples highlighted interspecific coordination in predator deterrence, similar dynamics occur in foraging. Traditionally, fish behavioural studies focused on single‐species shoals (Norris & Schilt, 1988; Pitcher, 2012). Recent research, however, has examined multi‐species shoals, particularly among coral‐reef species (Lukoschek & McCormick, 2000; Strand, 1988; Fig. 1C). Ormond (1980) categorised four potentially facilitative multi‐species foraging strategies: (1) following and scavenging, where large carnivorous fish are trailed by other carnivores and opportunistic feeders; (2) hunting by riding, where a predator positions itself near another species to conserve energy; (3) interspecific joint hunting, involving cooperation between predators; and (4) aggressive mimicry, where a predator mimics a harmless species. By contrast, non‐predatory species such as scarids and acanthurids, which primarily graze on algae or bite sessile corals, appear to benefit from group foraging through social facilitation or local enhancement. Studies have shown that bite rates in these non‐predatory species correlate with shoal size (Baird, 1993), and individuals in larger shoals feed for longer periods (Foster, 1985; Wolf, 1987). These benefits often arise through mechanisms such as local enhancement from a nuclear species (Lukoschek & McCormick, 2000) and social facilitation among attendants is also well documented, as seen between the puddingwife wrasse (Halichoeres radiatus) as a nuclear species and the bar jack (Caranx ruber) as an attendant (Baird, 1993).
Many associations, driven by foraging, also reduce predation risk. Mixed‐species shoals may enhance collective vigilance, as individuals respond to others' alarm cues or escape behaviours (Fig. 1C). Confusion effects also arise from different morphologies and escape tactics. Sazima et al. (2007) describe the goatfish (Pseudupeneus maculatus) as a nuclear species in multi‐species foraging groups, attracting jacks and other followers. Primarily foraging‐centred, these aggregations also offer antipredator advantages by increasing detection and reducing individual risk, suggesting that multi‐species shoals are flexible strategies balancing feeding efficiency and predator avoidance. Foraging benefits enhancing fitness can also arise from socially acquired information (Gil et al., 2017), evident in collective behaviours across taxa, including ontogenetic changes in fish within subtropical inshore habitats. Juvenile fish exchange information in MSGs, using behavioural cues from species they will later compete with as adults. This highlights the role of FIIs in shaping ontogenetic development, suggesting potential commensal or mutualistic dynamics within juvenile fish MSGs (Haak et al., 2020). Such intricate foraging associations are not limited to juveniles. A notable example in adults is the cooperative hunting between grouper (Plectropomus pessuliferus) and giant moray eel (Gymnothorax javanicus), where visual signals from the grouper coordinate behaviour, enhancing foraging success over solitary hunting (Bshary et al., 2006).
Beyond foraging‐related facilitations, fish engage in other notable FIIs such as cleaning interactions where cleaner fish remove ectoparasites and debris from larger ‘client’ fish, which benefit from parasite reduction (Beebe, 1928) (Figs 1D and 2A). Widely cited as a ‘textbook’ mutualism (Caves, 2021a) and sometimes cooperative (Limbaugh, 1961; Feder, 1966), these interactions involve clients modifying behaviour based on cleaning needs (Caves, 2021a) and signalling for coordination. Importantly, cleaning mutualisms are among the few marine interspecific interactions with experimentally demonstrated fitness benefits (Grutter, 1999; Bshary, 2003), showing increased client recruitment, survival, and local biodiversity. Soares et al. (2012) also found that clients adjust cleaning behaviour based on prior interactions, suggesting a learned component.
However, cleaning interactions also illustrate the context‐dependent nature of interspecific outcomes, transitioning from mutualistic to parasitic based on partner behaviour, environmental pressures, or individual decisions, a classic mutualism–parasitism continuum. While clients typically benefit from parasite removal, cleaners sometimes cheat by removing scales, mucus, or tissue (Limbaugh, 1961; Grutter, 1997; Soares et al., 2008a,b; Quimbayo et al., 2023). This imposes costs on clients and introduces conflict. Such variability highlights the complexity of cleaning interactions, where net outcomes are shaped by partner behaviour, ecological conditions, and individual decision‐making (Bronstein, 1994). Although generally facultative, marine cleaning mutualisms are mediated by signalling cues that influence engagement. Despite decades of research, key questions remain about abiotic, ecological, social, behavioural, and genetic factors affecting their dynamics and stability (Soares et al., 2008a,b; Caves, 2021a,b). Thus, cleaner stations remain valuable model systems for understanding the costs, benefits, and plasticity of interspecific facilitation in marine ecosystems.
POPULATION‐LEVEL CONSEQUENCES OF FACILITATIVE INTERSPECIES INTERACTIONS
IV.
Facilitation between species can enhance individual fitness by improving vital rates such as survival and fecundity, thus boosting population growth rates (Bronstein, 1994; Bruno et al., 2003; Crowley & Cox, 2011; Svenning et al., 2014). However, traditional population ecology has largely concentrated on antagonistic, density‐dependent factors like resource competition, disease transmission, and predation (Bronstein, 1994), often neglecting the vital role of facilitative interactions in marine ecosystems (Bruno et al., 2003) and their evolutionary implications (Stensland et al., 2003; Goodale et al., 2017).
Seabirds, once considered transient participants in ephemeral feeding aggregations with minimal population and community impact (Munn & Terborgh, 1979; Sridhar, Beauchamp & Shanker, 2009), are now recognised for their significant roles in interspecific foraging associations (Veit & Harrison, 2017), with clear population‐level consequences of FII. For instance, species such as the sooty tern (Sterna fuscata) and great frigatebird (Fregata minor) rely heavily on tuna and dolphins to access surface prey (Brewer & Hertel, 2007). Other aerial predators, such as frigatebirds themselves, even exploit prey like flying fish that evolved aerial escape tactics to avoid marine predators, only to become vulnerable to capture in the air. This dependence on the facilitator implies that fluctuations in the abundance or behaviour of these predators can cascade through seabird foraging efficiency, breeding success, and ultimately, population dynamics.
Although FIIs among seabirds are prevalent, with benefits generally outweighing antagonistic costs (Veit & Harrison, 2017), the underlying mechanisms driving these interactions remain elusive. A study on Aride Island in the Seychelles documented that roseate tern (Sterna dougallii) reproductive success improved through associations with predatory fish, which lowered breeding failure rates by increasing chick prey availability, demonstrating significant benefits from facilitative interactions with typical competitors (Ramos, 2000). Alongside other predators like tuna, dolphins, marlins, and billfish, seabirds use multidimensional predatory strategies to disrupt fish schools, boosting prey capture efficiency and thereby enhancing their foraging success and population resilience (Lett et al., 2014; Thiebault et al., 2016).
FIIs can directly influence population dynamics and individual variation within species. In southern Brazil, Bezamat et al. (2018) examined how cooperation between Lahille's dolphins (T. truncatus gephyreus) and net‐casting fishers was linked to individual variation and population dynamics, providing a framework to assess the demographic consequences of FII. While fishers benefit through increased catch rates (Simões‐Lopes, 1998; Cantor et al., 2023), long‐term effects on dolphins also appear positive, with higher survival rates for cooperating individuals (Bezamat et al., 2018). Similarly, Syme et al. (2023) explored how MSGs involving Australian humpback dolphins (Sousa sahulensis) and Indo‐Pacific bottlenose dolphins (Tursiops aduncus) in Northwest Cape, Western Australia, serve social functions, such as refining social skills and providing alloparental care, alongside antipredator benefits. Conversely, population density and composition (e.g. relative abundance of adults and juveniles) can also shape FIIs by influencing how individuals engage with conspecifics and heterospecifics across space and time. This highlights how population characteristics can mediate facilitative interactions. For instance, Elliser & Herzing (2016) demonstrated that population characteristics influenced the frequency and nature of mixed‐species encounters between Atlantic spotted dolphins (Stenella frontalis) and Atlantic bottlenose dolphins (Tursiop truncatus) in the Bahamas, suggesting a role of demographic structure in maintaining interspecific associations and overall population stability.
These population‐level consequences of FIIs are not limited to seabirds and marine mammals. Similar patterns emerge in other taxa, including fish, where facilitation influences survival and population dynamics during critical life stages. Juvenile fish use social information in FIIs, including visual and chemical cues about predation risk and food availability (Haak et al., 2020). In mixed‐species prey guilds of similarly sized individuals, this information sharing enhances collective vigilance, coordinated escape responses, and detection of food patches (Paijmans, Reardon & Ward, 2020; Mukherjee & Bhat, 2023; Goodale et al., 2010). These mechanisms improve survival and foraging efficiency during vulnerable early developmental stages, even though many species later become competitors as adults (Lukoschek & McCormick, 2000). Such facilitative dynamics can also contribute to the stability of prey communities (Mukherjee & Bhat, 2023). Beyond prey guilds, empirical studies of cleaning interactions on coral reefs further underscore the importance of FIIs. For example, Waldie et al. (2011) demonstrated that removing cleaner fish led to declines in species abundance, growth, survival, and recruitment, highlighting the foundational role of such interactions in maintaining ecological balance.
However, capturing these complex dynamics in predictive models remains a significant challenge. Although mutualistic population dynamic models, such as those adapting the Lotka–Volterra framework by Gause & Witt (1935), exist, they often face mathematical instability, leading to unrealistic, unbounded growth – a problem identified in the work of Gause & Witt (1935) and addressed by Holland, DeAngelis & Bronstein (2002). Stable coexistence in these models is only possible under specific conditions, such as when mutualistic benefits saturate at high densities or are offset by strong self‐limitation (e.g. density dependence; Holland et al., 2002). Despite these challenges in modelling, recent advances by Hart (2023) are integrating facilitative interactions into classical models, utilising coexistence theory to refine our understanding of mutualistic and commensal relationships. Hart (2023) categorised facilitation within these models, highlighting a shift in coexistence theory from competition to facilitative interactions and stressing the importance of reciprocal interactions and density dependence in ecological outcomes. Integrating empirical and theoretical work is essential for a deeper understanding of the role of facilitation in population to ecosystem dynamics, and for predicting responses to future changes.
IMPLICATIONS OF FACILITATIVE INTERSPECIES INTERACTIONS FOR COMMUNITIES AND ECOSYSTEMS
V.
Compelling evidence indicates that facilitators – often mediated by foundation species, keystone structures, and ecosystem engineers (see Table 1) – can trigger strong direct and indirect effects that cascade through ecological communities, shaping both structure and function (Paine, 1966; Thomsen et al., 2010; Thomsen & Wernberg, 2014; Stachowicz, 2001; Witman, 1987; Stachowicz & Hay, 1999; Lafferty & Suchanek, 2016). Within the framework proposed by Thomsen et al. (2010), such interactions may propagate via “keystone” or “cascading” facilitation pathways. These facilitators often enhance biodiversity and ecological resilience, with keystone species contributing through biogenic habitat provision (Paine, 1966; Dayton, 1975; Jones, Lawton & Shachak, 1997; Lafferty & Suchanek, 2016), and Stachowicz (2001) argued that they can promote diversity similarly to predation by suppressing dominant competitors. The primary mechanism involves habitat‐forming organisms functioning as ecosystem engineers (Jones et al., 1994); by modifying the physical environment – altering sedimentation, nutrient dynamics, water flow, or structural complexity – they provide refuge, enhance local productivity, reduce environmental stress, and mitigate predation risks, thus facilitating other species' survival, growth, or reproduction (Bertness & Leonard, 1997; Bulleri, 2009; Bertness & Callaway, 1994; Jones et al., 1994). However, most of this empirical research has focused on sessile or sedentary groups, such as mussel beds, oyster reefs, coral reefs, macroalgae canopies, seagrasses, salt marshes, and mangroves (Stachowicz, 2001; Bulleri, 2009), whereas mobile marine vertebrates remain comparatively understudied as facilitators.
While direct behavioural interactions are the focus of this review, it is important first to recognise that mobile vertebrates also act as powerful facilitators through indirect, ecosystem‐engineering pathways. For instance, the “whale pump” – nutrient‐rich faeces released by whales near the surface – enhances primary productivity, increasing phytoplankton and potentially supporting larger primary consumer populations (Roman & McCarthy, 2010). Similarly, seabird guano provides substantial nutrient inputs to coastal ecosystems, fertilising primary producers and influencing the abundance and reproductive success of organisms across trophic levels (Polis, Anderson & Holt, 1997). These bottom‐up, nutrient‐mediated pathways demonstrate how ecosystem‐level facilitation directly shapes the productivity and persistence of recipient populations.
Beyond these indirect engineering roles, mobile marine vertebrates are also central to community dynamics through the direct behavioural interactions that are the primary focus of this review. For instance, Goyert et al. (2014) linked common tern (Sterna hirundo) and roseate tern (Sterna dougallii) distribution and abundance to associations with subsurface predators (tunas and dolphins) in the North Atlantic, exploring the implications of commensalism and local enhancement at the community level. This research classified tunas as “keystone facilitators” due to their impact on forage fish (Ashmole & Ashmole, 1967; Au & Pitman, 1986), thus impacting tern assemblages. Such interactions demonstrate how prey abundance dictates whether species compete under low prey availability or engage in mutualistic/commensal relationships to facilitate predation (Goyert et al., 2014; Au & Pitman, 1986). Under scarcity, terns may experience increased competition, leading to agonistic interactions or spatial segregation. Conversely, abundance or concentration of prey by facilitators like tunas allows species to benefit from information transfer and local enhancement, where individuals use others' presence and foraging success to locate and exploit prey more efficiently. This dynamic interplay highlights the context‐dependent nature of species interactions along a competition–facilitation spectrum.
This context‐dependent dynamic extends beyond the nature of the interaction to the very composition of the interacting groups themselves. Gostischa, Massolo & Constantine (2021) extensively explored mixed‐species feeding associations (MSFAs) among large marine predators (seabirds, cetaceans, sharks, and seals) analysing association patterns based on species composition, temporality, and prey preferences. They identified three MSFA categories by feeding habits and functional group composition. Over a decade, MSFA composition and dynamics showed temporal variability, emphasising the sensitivity of these facilitative communities to long‐term environmental changes. Such findings underscore the need to understand that the structure of these communities is not static, but shifts over time, which in turn influences the costs and benefits for all participants.
These shifting costs and benefits are often determined by the spatial co‐occurrence and composition of the groups. Indeed, studies on dynamic MSGs (seabirds, cetaceans, fish, and pinnipeds) emphasise how the net benefit for an individual fluctuates with group composition (Ashmole & Ashmole, 1967; Anderwald et al., 2011; Au & Pitman, 1986; Hebshi et al., 2008). A clear example is the commensal relationship observed in Norwegian fjords, where humpback whales (Megaptera novaeangliae) benefit from the presence of killer whales (Orcinus orca) while foraging on herring (Clupea harengus) (Jourdain & Vongraven, 2017). Although some group members may experience temporary disadvantages, these associations persist, demonstrating the resilience and adaptability of marine communities. This variation in group composition and community dynamics is often driven by environmental factors, as illustrated by Correia et al.'s (2019) investigation of MSGs of terns and subsurface predators during the terns' non‐breeding period in West Africa's shallow marine waters. They observed various interspecific interactions among tern species, with black terns (Chlidonias niger) foraging with heterospecifics in 89% of observations, indicating high dependence. The study highlighted how MSG composition varied with water depth: smaller predators like jacks (Caranx hippos) in shallow waters and larger predators like tunas, cetaceans, and pinnipeds in deeper waters. These insights reveal the complex dynamics of interspecific interactions, emphasising how environmental factors like water depth shape group composition.
Shifting the focus beyond apex predators, recent research on fish has explored community‐ and ecosystem‐level facilitation using interaction webs and network theory, particularly in coral reef ecosystems. For example, Fernández‐Cisternas et al. (2021) examined empirical facilitative interactions – such as mutualistic cleaning and following behaviour – within reef fish assemblages around Rapa Nui and Robinson Crusoe Islands. Their analysis also incorporated trophic interactions enabling indirect facilitation, such as predator‐mediated effects and trophic cascades. The study revealed how both top‐down pressures (e.g. fisheries) and bottom‐up drivers (e.g. productivity) shape the structure and resilience of facilitative networks. Robinson Crusoe Island exhibited a robust network, with a balance of agonistic and facilitative links contributing to higher community stability, whereas Rapa Nui's more fragile network suggested greater vulnerability to perturbations.
While such empirical studies advance our understanding of facilitation in marine networks, many interaction web models (e.g. Kéfi et al., 2015, 2016) are largely based on inferred or assumed interactions. These conceptual models are useful for exploring potential ecological outcomes, but they require validation with field data to assess the strength and direction of actual species interactions. Moreover, indirect facilitation is often already embedded in classical food‐web structures, where trophic pathways link species in complex but ecologically meaningful ways (e.g. species A suppresses predator B of species C). Mutualistic networks have been extensively studied in terrestrial ecosystems (e.g. plant–pollinator systems) and, to a lesser extent, in marine contexts such as anemone–fish associations (but see marine cleaning mutualisms, e.g. Quimbayo et al., 2018). An alternative empirical approach involves individual‐based network analyses that document species co‐occurrence and spatial associations without presupposing inhibition or facilitation (Thomsen & South, 2019). These approaches are promising for uncovering patterns of facilitation in dynamic, multi‐species marine systems.
Providing such an empirical approach, Auster et al. (2019) analysed a piscivorous fish assemblage engaged in group hunting within the coral reef ecosystem of Isla del Coco, Costa Rica. Using a network approach to study collective behaviour, they identified dominant species and highlighted the importance of quantifying facilitative interaction attributes within trophic guilds. This research sheds light on cooperative behaviours underpinning group hunting dynamics and contributes to conservation strategies by emphasising facilitative roles essential to ecosystem integrity.
Taken together, these examples illustrate that FII influences marine ecosystems via direct and indirect pathways. By integrating concepts such as interaction networks, keystone facilitation, niche modification, and non‐trophic mutualisms, we emphasise that facilitation is not limited to pairwise benefits but can restructure entire communities. Viewing facilitation through these ecological lenses reveals it as a powerful yet underappreciated mechanism shaping biodiversity, resilience, and trophic dynamics in marine systems.
INTEGRATING FACILITATIVE INTERACTIONS IN ECOSYSTEM‐BASED MANAGEMENT
VI.
As discussed above, FIIs play a critical role in marine ecosystems, influencing individual fitness, population dynamics, and community structure. These positive interactions, involving both apex predators and lower trophic‐level species, can shape food‐web resilience and ecosystem function. Yet, despite their ecological significance, FII remains largely absent from ecosystem models used in EBFM. EBFM frameworks seek to integrate ecological, social, and economic dimensions to manage fisheries sustainably (Link, 2010). To support this goal, a variety of ecosystem models (EMs) have been developed that differ in structure, assumptions, and complexity (Plagányi, 2007; Table 2). These models provide insight into the cumulative effects of fishing and environmental change by treating ecosystems as dynamic, interconnected systems. However, most EMs emphasise antagonistic interactions – such as predation and competition – while positive interspecific relationships are rarely represented. This limits their ability to capture the full complexity of marine ecosystem dynamics and may overlook important mechanisms that support biodiversity and ecosystem stability.
One of the most widely used tools for EBFM is Ecopath with Ecosim (EwE), a modelling suite that simulates energy flow and trophic dynamics in marine ecosystems. We discuss this model in more detail here to illustrate our argument regarding the inclusion of FII in models for EBFM. Initially developed by Polovina (1984) and expanded by Christensen & Pauly (1992), the Ecopath component constructs static, mass‐balance models of trophic interactions based on species biomass, diet composition, and fishing mortality. Its dynamic counterpart, Ecosim, incorporates temporal processes using differential equations to simulate population changes in response to fishing pressure, environmental variability, and habitat change (Walters, Christensen & Pauly, 1997; Ahrens, Walters & Christensen, 2012). Together, these tools enable scenario testing, long‐term trend analysis, and adaptive management planning (Christensen & Walters, 2004; Walters et al., 2005).
Despite EwE's broad application – evident in the 477 models archived in the EcoBase database (Colléter et al., 2015) – none currently incorporate FII involving apex predators or forage fish. This absence reflects the model's default structure, designed to track energy transfer through antagonistic links, illustrating a critical gap and a broader historical bias in ecological modelling towards competition and predation. This bias risks neglecting the stabilising or diversity‐enhancing effects of FII. However, EwE offers potential avenues for incorporating facilitation. The ‘mediation’ function can modify trophic interactions based on the biomass of a third species, offering a practical route for modelling facilitation. For example, a facilitating species could increase prey vulnerability or enhance foraging efficiency for another predator – mechanisms common in MSFAs.
Additionally, the vulnerability parameter in Ecosim, which controls the strength of top‐down versus bottom‐up control in predator–prey interactions, can be adapted to reflect facilitative mechanisms. By increasing prey vulnerability to a specific predator when facilitation occurs (e.g. through cooperative hunting), users can simulate enhanced foraging efficiency or prey accessibility in scenarios involving positive interspecific interactions. Adapting EwE and similar models to include facilitation would be a major step forward for EBFM. FIIs can modulate species coexistence, buffer ecosystems against perturbations, and influence community assembly – dynamics essential for anticipating ecological responses to fishing and climate change (see Section VII). Integrating these interactions would enhance the ecological realism of EMs, support better‐informed management strategies, and help safeguard marine biodiversity in an era of increasing human impact.
CONSERVATION RELEVANCE
VII.
Facilitative dynamics may profoundly impact species ecology (Bruno et al., 2003; Bronstein, 1994; Stachowicz, 2001), and here we examine their implications for conservation success and biodiversity. Conservation efforts prioritise protecting endangered, threatened, or vulnerable species and their habitats to prevent extinction and preserve biodiversity (Pullin, 2002; Lindenmayer & Burgman, 2005). Frameworks like ‘umbrella’ and ‘keystone’ species enhance conservation planning by highlighting species' ecological roles and significance within ecosystems (Pullin, 2002; Lindenmayer & Burgman, 2005; Caro, 2010; Barua, 2011). Umbrella species are typically large, wide‐ranging organisms whose conservation benefits numerous cohabiting species, while keystone species are vital to maintaining ecosystem structure and function (Pullin, 2002; Lindenmayer & Burgman, 2005; Caro, 2010; Barua, 2011). Recognising and protecting these species maximises conservation and biodiversity outcomes. Studies on FII have introduced the idea of ‘keystone facilitators’ (Au & Pitman, 1986; Ashmole & Ashmole, 1967; Stachowicz, 2001; Goyert, Manne & Veit, 2014; Goyert et al., 2018), which attract other species to MSGs through their behaviours (Goodale et al., 2017). Identifying such facilitators may further improve conservation strategies (Gostischa et al., 2021; Goyert et al., 2014; Veit & Harrison, 2017).
Moreover, keystone and umbrella species often serve as ‘flagship species’ due to their charisma, aesthetic appeal, or cultural significance (Caro, 2010; Barua, 2011). Understanding these species, particularly in the context of MSGs, strengthens conservation by utilising their emblematic status for broader actions. Many marine apex predators discussed herein (whales, dolphins, pinnipeds, albatrosses, and sharks) are flagship species (IUCN, 2024). These groups face significant threats from fisheries (Wade et al., 2021; Žydelis, Small & French, 2013; Pott & Wiedenfeld, 2017), with bycatch impacting various taxa (Dulvy et al., 2021) and overfishing affecting sharks and tunas (Hunter et al., 1986; Kompas, Grafton & Che, 2010). While some apex predators like sharks influence ecosystems through trophic cascades (e.g. mesopredator suppression), they are not necessarily facilitative as defined here, as they do not provide non‐trophic benefits to other species. Clarifying this distinction avoids conflating top‐down control with facilitation. Thus, conservation should also prioritise less‐charismatic but ecologically vital facilitative species that directly contribute to community structure and resilience.
The ecosystem‐based conservation approach is widely regarded as effective for preserving ecosystem structure and functionality (Garcia & Cochrane, 2005). Conservation biology emphasises identifying keystone, umbrella, and flagship species, which are central to this ecosystemic paradigm, as their protection is expected to create cascading benefits across species networks, including FIIs. Furthermore, this approach integrates socio‐economic considerations (Long, Charles & Stephenson, 2015; Curtin & Prellezo, 2010; Gruber, 2010; Garcia & Cochrane, 2005), promoting sustainable fisheries practices and stakeholder engagement via bioeconomic models (e.g. Azevedo et al., 2024). Employing ecosystem models that explicitly characterise facilitative interactions alongside antagonistic ones therefore will provide a more comprehensive strategy for species conservation.
Beyond their intrinsic ecological value, FIIs are increasingly shaped by anthropogenic pressures. The five major drivers of biodiversity loss – climate change, habitat destruction, resource extraction (e.g. overfishing), biological invasions, and pollution/eutrophication – can significantly disrupt FIIs across marine ecosystems. Among these, climate change has received the most attention. For example, shifts in ocean temperature and prey distribution have been shown to destabilise MSFAs involving krill predators such as seabirds and seals (Monier, Veit & Manne, 2020), while environmental variability can alter MSFA composition (Gostischa et al., 2021). By contrast, the effects of habitat degradation, pollution, overfishing, and invasive species on FIIs remain poorly understood. However, it is plausible that these stressors have significant impacts: habitat degradation could reduce refuge‐based facilitation; pollution may impair the sensory mechanisms essential for interspecific coordination; overfishing can remove key facilitator species; and invasive species may dismantle native facilitative networks. Despite these potential consequences (Fig. 3), FIIs are still largely overlooked in conservation planning. Explicitly incorporating FII into management frameworks could enhance ecosystem resilience and adaptive capacity in the face of global change.
Schematic overview of how anthropogenic stressors impact facilitative interspecific interactions (FIIs) in marine ecosystems. The diagram presents four illustrative examples of FII (left column) and links them to major human‐driven stressors (centre column), showing their respective ecological consequences (right column). Arrows indicate the causal pathways through which stressors alter the structure, function, or persistence of these interactions. This conceptual synthesis highlights how different stressors can disrupt species coordination, communication, sheltering, and mutualistic behaviours essential to ecosystem functioning. MSFA, multi‐species feeding association.
IMPLICATIONS FOR HUMAN–WILDLIFE INTERACTIONS
VIII.
FIIs are also evident in human–wildlife contexts (van der Wal et al., 2022). A notable example occurs in southern Brazil, where artisanal fishers collaborate with wild bottlenose dolphins (T. truncatus gephyreus) to catch migratory mullet (Mugil spp.) (Simões‐Lopes, 1998; Cantor et al., 2023). In this traditional fishery, dolphins herd mullet towards shallow waters and produce cues that signal when and where fishers should cast their nets. Disoriented by the nets, mullet become easier prey, allowing dolphins to forage more effectively using echolocation (Cantor et al., 2023). This interaction is one of the few remaining examples of human–wildlife cooperation and represents a flagship ecological interaction, a rare but culturally and ecologically significant relationship that fosters conservation awareness and strengthens human–nature connections (see Table 1). When foraging in synchrony, dolphins are nearly three times more likely to succeed in hunting, while fishers are 17 times more likely to catch mullet (Cantor et al., 2023). This cooperation can scale up to influence population dynamics, community structure, and ecosystem‐level processes (Bezamat et al., 2018; Dalpaz et al., 2024; Wooster et al., 2023). However, this mutualism may shift towards commensalism or even kleptoparasitism depending on ecological conditions, particularly when synchrony is low or prey is scarce.
Beyond enhanced foraging, human–wildlife FIIs also generate non‐material benefits. For southern Brazilian fishers, these include cultural services such as a sense of place, belonging, and cooperation among fishers (Machado et al., 2019; Santos‐Silva et al., 2022). For dolphins, cooperation is associated with increased survival (Bezamat et al., 2018, 2021) and stronger social bonds (Daura‐Jorge et al., 2012; Daura‐Jorge, Ingram & Simões‐Lopes, 2013; Machado et al., 2019). Still, the interaction faces multiple threats, including changing and declining fisher participation (Smith et al., 2009; Cantor et al., 2024), overfishing (Santos, Lemos & Viera, 2018; Simões‐Lopes, 1998), and bycatch from illegal fishing (Bezamat et al., 2021; Peterson, Hanazaki & Simões‐Lopes, 2008). Recognising the importance of such FII in conservation planning is essential for ecosystem health, the maintenance of ecosystem services, and the well‐being of coastal communities.
FILLING THE GAPS AND FUTURE DIRECTIONS
IX.
Individual level
(1)
While FIIs have moved beyond being a niche topic in ecology (Carlson et al., 2023), significant knowledge gaps remain. Key questions focus on the behavioural differences between conspecifics engaged in FIIs and those typically involved in single‐species interactions. Establishing a framework to measure foraging efficiencies explicitly – such as bite rates, foraging duration, prey quality, and other metrics – between mixed‐ and single‐species groups could provide valuable insights into the comparative advantages and trade‐offs of these interaction types. Further research should also examine the level of behavioural coordination among parties in FIIs, aggregations, MSGs, or MSFAs. Additionally, investigating the necessity of communication for achieving common goals, as well as potential differences in communication strategies between conspecifics and heterospecifics, deserves further attention. Understanding how individuals balance trade‐offs between positive and negative outcomes is critical, as variations within populations ultimately shape the dynamics of interspecific interactions.
Farine, Garroway & Sheldon (2012) advocate for a ‘bottom‐up’ approach, focusing on individuals as the primary unit of analysis to deepen our understanding of FII dynamics. Social network analysis, grounded in individual‐centric rather than species‐centric paradigms, is pivotal to advancing this bottom‐up approach. Initially applied in avian studies, no theoretical barriers (other than logistical challenges in aquatic sampling) prevent extending individual‐based social network analysis to marine fish and apex predators, as explored by Krause, Croft & James (2007) and further analysed by Krause et al. (2015) and Croft, James & Krause (2008). Recent developments now enable statistical inference at the individual level. For example, Thomsen & South (2019) introduced frameworks allowing error estimation on individual nodes within ecological networks, offering more robust assessments of species interactions and their variability. However, understanding how the environment influences individual decisions and behaviours during mixed‐species encounters and sequential facilitation is a major challenge. Many FIIs occur in specific spatiotemporal contexts, underscoring the need to explore the interaction between environmental factors and individual behavioural responses. Such studies promise to illuminate the dynamics and ecological implications of FIIs while assessing the broader ecosystem effects of individual choices and behaviours.
Population level
(2)
As noted by Hart (2023), unlike research on competition and predation, studies on facilitation – aside from mutualism – lack a solid foundation in population dynamic theory. While mutualisms have been more extensively studied, other forms of facilitation remain less explored. Consequently, there has been less focus on estimating state variables and population‐level parameters that predict the outcomes of facilitative interactions. Despite these challenges, research has increasingly explored how facilitation influences population abundance, distribution, and demography (Bruno et al., 2003; Bulleri, 2009).
Several challenges persist in understanding the connections between FII and population ecology. As Agrawal et al. (2007) and Chu et al. (2008) have emphasised, population density and environmental factors determine whether interactions are mainly positive (facilitative) or negative (competitive). Evaluating how fish and marine apex predator population densities involved in FII influence population dynamics could yield valuable insights, particularly given environmental change. Cameron, Coulson & Marshall (2019), Lin et al. (2012, 2016), Malanson & Resler (2016), and Zepeda & Martorell (2019) have noted that both population density and individual size structure significantly impact the frequency and intensity of facilitation versus competition, affecting the life‐history traits of species involved.
Fish and marine apex predators could serve as focal groups to test the abiotic stress gradient hypothesis proposed by Bertness & Callaway (1994), which suggests that facilitation is more prevalent under high abiotic stress, while competition dominates under less‐stressful conditions. However, recent findings suggest strong facilitation can occur under low‐stress conditions, particularly when interactions are driven by predator avoidance rather than environmental harshness (Kawai & Tokeshi, 2007). For example, small fish shelter among jellyfish tentacles in benign pelagic environments, gaining protection from predators (Griffin et al., 2019). This suggests a unimodal pattern between stress and facilitation, peaking at moderate stress levels and declining at both extremes (Michalet et al., 2006, 2014; Xiao et al., 2009; Holmgren & Scheffer, 2010; Verwijmeren et al., 2013; Le Bagousse‐Pinguet et al., 2014). Testing this hypothesis in diverse ecological contexts, particularly for mobile fauna, could clarify whether FIIs involving fish and marine apex predators also intensify under high environmental stress, as the theory was initially developed for foundation species that facilitate through physical habitat modification, like mussel beds, coral reefs, and mangroves.
Additionally, examining the demographic aspects of fish and apex predator populations involved in FII could deepen our understanding of population dynamics, as suggested by Caswell (2006). Demographic models provide a framework for integrating empirically estimated rates, such as survival, growth, and reproduction, across life stages to project future population growth under current ecological conditions. Analysing species distribution ranges could also shed light on how facilitation influences population dynamics, as highlighted in studies by Bulleri et al. (2016) and Butterfield (2009).
Community and ecosystem levels
(3)
McIntire & Fajardo (2014) highlight facilitation as a key driver of biodiversity, as species modify local biotic or abiotic conditions through resource exploitation, thus reshaping multidimensional niches. This alteration can shift species composition, often enhancing both beta and, occasionally, alpha diversity. The importance of facilitation is amplified in complex communities where species not only exploit but also create niches via interactions with the environment. While McIntire & Fajardo's (2014) work focused primarily on sessile organisms, all organisms, regardless of mobility, can influence their environment and shape ecological outcomes. Foundational studies on facilitation by sessile or low‐mobility organisms (e.g. Bertness & Callaway, 1994; Dayton, 1972; Jones et al., 1994, 1997; Ellison et al., 2005; Tews et al., 2004; Townsend, 2021) have underscored the importance of foundation species, keystone structures, and allogenic ecosystem engineers in promoting biodiversity and stabilising communities.
However, recent research shows that facilitation also plays a critical role among mobile organisms. Although historically overlooked, mobile species can function as biodiversity reservoirs, expanding the conceptual scope of facilitation (Altieri & Witman, 2014). Despite logistical challenges such as spatial and temporal segregation (e.g. Karban, Grof‐Tisza & Holyoak, 2012; Li et al., 2018), facilitative interactions among mobile species are increasingly recognised as ecologically relevant, contributing to coexistence, community resilience, and ecosystem functioning (e.g. Canepuccia et al., 2020; van der Wal et al., 2022). Such interactions often involve one species enhancing another's access to or quality of resources, influencing species distributions and community dynamics, potentially driving biodiversity (e.g. Bronstein, 2009; Hale, Valdovinos & Martinez, 2020).
Canepuccia et al. (2024) demonstrate how indirect and reciprocal facilitative effects intricately influence species' behaviour, abundance, and resource use within interaction networks. These modifications apply selection pressures across communities, expanding niches and enhancing biodiversity. Empirical evidence across botanical contexts consistently underscores the importance of facilitation for ecosystem structure, function, and dynamics (Callaway, 2007). Understanding facilitation mechanisms and their broader ecosystem impacts is critical, especially for mobile species like fish and apex predators. Michalet & Pugnaire (2016), for instance, suggest that understanding the role of facilitation could improve ecosystem management and restoration by leveraging positive feedback loops to avert catastrophic shifts. Fully comprehending species‐level facilitation and its ecosystem‐level implications requires investigating complete interaction networks among species and their abiotic environments, merging ecosystem and community ecology approaches (Kéfi et al., 2016).
Ecosystem modelling has explored facilitation in diverse contexts, although gaps remain in addressing interactions among mobile species. Ecopath models have considered facilitation, such as those of Harvey (2014), who explored eelgrass (Zostera marina) as a refuge habitat for juvenile Pacific salmon (Oncorhynchus spp.). However, scaling mediation functions to ecosystem levels poses challenges in Ecopath, as detailed by Espinosa‐Romero et al. (2011) and Plummer et al. (2013). Harvey (2014) emphasises the need for exploratory approaches to refine model assumptions, while Espinosa‐Romero et al. (2011) exemplify the application of Ecopath's mediation function, showing how kelps enhance feeding areas and food availability for other species, significantly affecting biomass and energy flow of fish, benthic invertebrates, otters, and detritus. Although these findings diverge from this review's focal groups, they underscore the potential value of integrating facilitation within an ecosystem framework. In addition to the mediation function, the Ecosim's ‘vulnerability’ parameter also provides a promising tool to represent facilitation, allowing predator–prey interactions to shift based on the presence or behaviour of a facilitator, consistent with the principles of the foraging arena theory (Walters et al., 1997).
CONCLUSIONS
X.
- (1)This review synthesises the multifaceted roles of facilitative interspecific interactions (FIIs) across multiple biological scales in the marine environment, connecting individual‐level behaviours to their broader consequences for populations, communities, and ecosystems.
- (2)Significant progress has been made in descriptive studies over recent decades; however, important gaps remain in understanding the impacts of FIIs on population dynamics, community structure, and ecosystem processes within marine environments.
- (3)Facilitation has been widely studied in sessile species, often supported by experimental evidence and long‐term demographic data demonstrating clear positive effects on population dynamics. By contrast, facilitation among pelagic animals remains less experimentally tested, with most studies relying on correlations. While these correlations suggest potential positive interactions, the absence of long‐term demographic data makes it difficult to confirm facilitation in the strictest sense. Future studies incorporating experimental manipulations or long‐term monitoring could help establish causal relationships and clarify the demographic consequences of these interactions.
- (4)Our synthesis also highlights a key research gap: despite growing recognition of the importance of FIIs, few studies have quantified their outcomes in terms of reproductive success, population growth, or community stability. Addressing this gap will be essential for improving ecological models, informing conservation strategies, and distinguishing facilitation more clearly from other ecological processes such as trophic interactions.
- (5)We strongly recommend incorporating FII into ecosystem models to improve Ecosystem‐Based Fishery Management (EBFM). This is particularly critical for conserving vulnerable taxa, including fish, sharks, seabirds, cetaceans, and pinnipeds, whose populations may depend on these essential interactions.
- (6)Finally, ensuring the resilience of human communities that rely on ocean ecosystems is an urgent priority. In this context, FIIs involving humans and apex predators pose unique management challenges, yet they also offer important opportunities for expanded data collection, model development, and understanding broader ecological and socio‐economic impacts.
AUTHOR CONTRIBUTIONS
E. D. V.: Conceptualisation; Data collection; Data curation; Investigation; Methodology; Visualisation; Writing – original draft. F. G. D‐J.: Conceptualisation; Data collection; Formal analysis; Methodology; Funding acquisition; Project administration; Writing – original draft; review & editing; Supervision; Validation; Visualisation; J. L. R‐F.: Investigation; Resources; Supervision; Writing – review & editing; R. A.: Investigation; Resources; Supervision; Writing – review & editing; M. C.: Investigation; Resources; Writing – review & editing; S. S.: Conceptualisation; Data collection; Formal analysis; Methodology; Writing – original draft; review & editing; Supervision;
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Acevedo‐Gutiérrez, A. (1999). Aerial behavior is not a social facilitator in bottlenose dolphins hunting in small groups. Journal of Mammalogy 80(3), 768–776.
- 2Agrawal, A. A. , Ackerly, D. D. , Adler, F. , Arnold, A. E. , Cáceres, C. , Doak, D. F. , Post, E. , Hudson, P. J. , Maron, J. L. , Mooney, K. A. , Power, M. , Schemske, D. , Stachowicz, J. , Strauss, S. , Turner, M. G. & Werner, E. (2007). Filling key gaps in population and community ecology. Frontiers in Ecology and the Environment 5(3), 145–152.
- 3Ahrens, R. N. , Walters, C. J. & Christensen, V. (2012). Ecosim: modelling ecosystem dynamics over time. Fisheries Centre Research Reports 20(2), 1–99.
- 4Ainsworth, C. H. , Samhouri, J. F. , Busch, D. S. , Cheung, W. W. , Dunne, J. & Okey, T. A. (2011). Potential impacts of climate change on Northeast Pacific marine foodwebs and fisheries. ICES Journal of Marine Science 68(6), 1217–1229.
- 5Altieri, A. H. & Witman, J. D. (2014). Modular mobile foundation species as reservoirs of biodiversity. Ecosphere 5(10), 1–11.
- 6Anderwald, P. (2011). Ecology and behaviour of minke whales in the Northeast Atlantic. Doctoral Dissertation. University of Aberdeen.
- 7Anderwald, P. , Evans, P. G. , Gygax, L. & Hoelzel, A. R. (2011). Role of feeding strategies in seabird‐minke whale associations. Marine Ecology Progress Series 424, 219–227.
- 8Ashmole, N. P. & Ashmole, M. J. (1967). Comparative feeding ecology of sea birds of a tropical oceanic Island. Bulletin of the Peabody Museum of Natural History 40(1), 1–131.