Polyphasic characterization of Biscogniauxia papillata sp. nov. (Graphostromataceae) and isolation of the cytotoxic cyclic pentapeptide cyclobiscognioxin A
Sarunyou Wongkanoun, Esteban Charria-Girón, Boonchuai Chainuwong, Prasert Srikitikulchai, Natapol Pornputtapong, Jennifer Luangsa-ard, Sherif S. Ebada, Marc Stadler

TL;DR
Scientists discovered a new fungus species in Thailand and isolated a toxic compound from it.
Contribution
The discovery of Biscogniauxia papillata and the isolation of cyclobiscognioxin A, a new cytotoxic cyclic pentapeptide.
Findings
Biscogniauxia papillata is a new species identified through morphology and phylogenetic analysis.
Cyclobiscognioxin A, a cytotoxic cyclic pentapeptide, was isolated and lacks antimicrobial activity.
The chemical diversity of Biscogniauxia suggests potential for biosynthetic exploration.
Abstract
The Graphostromataceae (Xylariales) mainly contains species known for their endophytic lifestyle, although some may shift to opportunistic pathogenicity under host stress and subsequently colonize woody tissues as saprotrophs. The largest genus, Biscogniauxia, remains largely unexplored and would greatly benefit from a polyphasic taxonomic revision. Likewise, the secondary metabolism of these fungi has been only partially studied compared to related families within the Xylariales, despite their ecological versatility and potential latent pathogenicity. Herein, we introduce Biscogniauxia papillata as a new species from Thailand, based on morphological features and robust multi-locus phylogenetic analyses. Secondary metabolite profiling in different media, followed by preparative chromatography, led to the isolation of a cyclic pentapeptide named cyclobiscognioxin A (1), along with a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3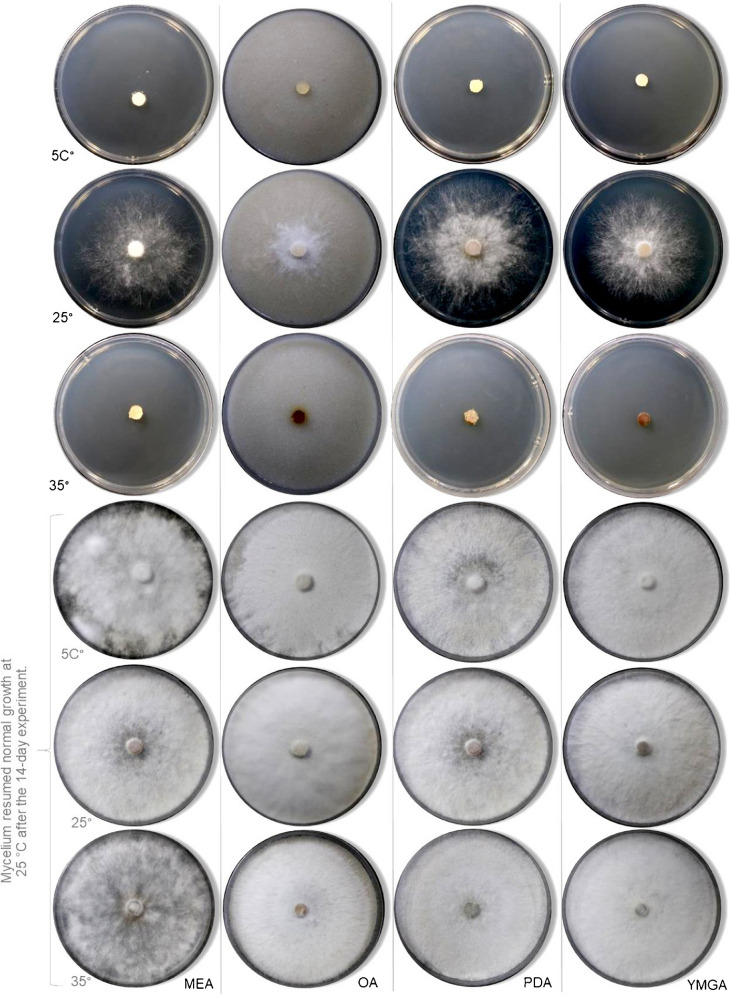 Figure 4
Figure 4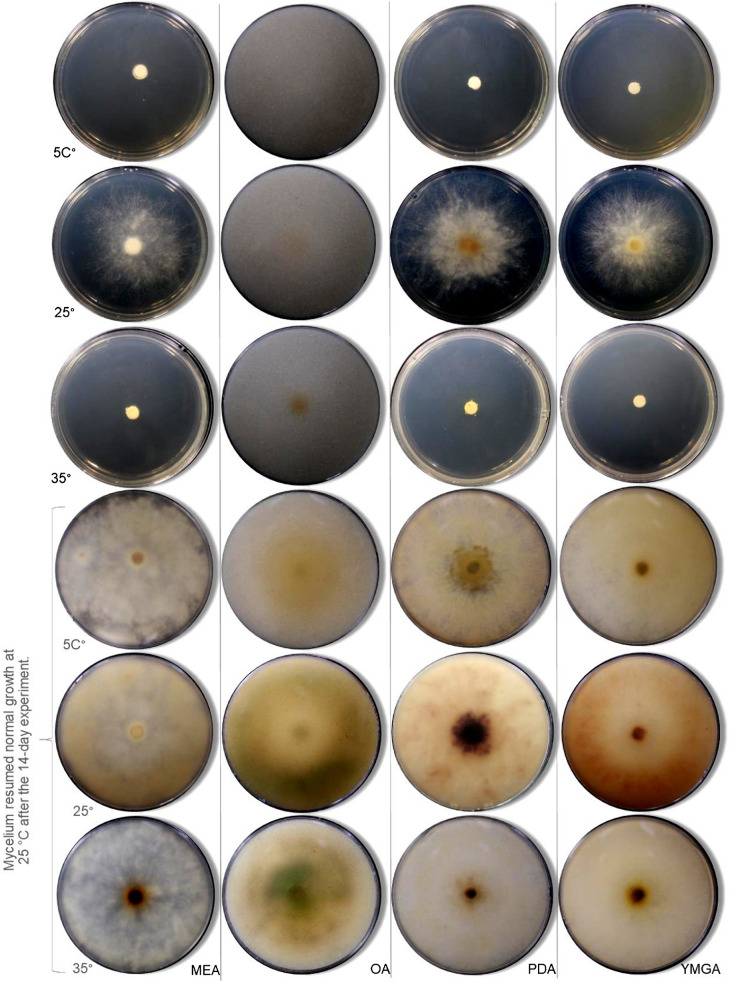 Figure 5
Figure 5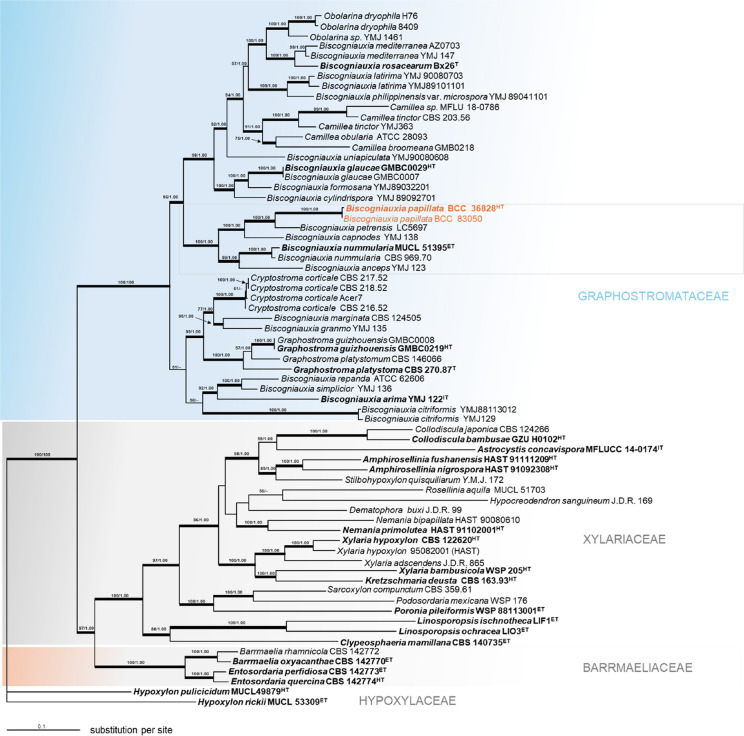 Figure 6
Figure 6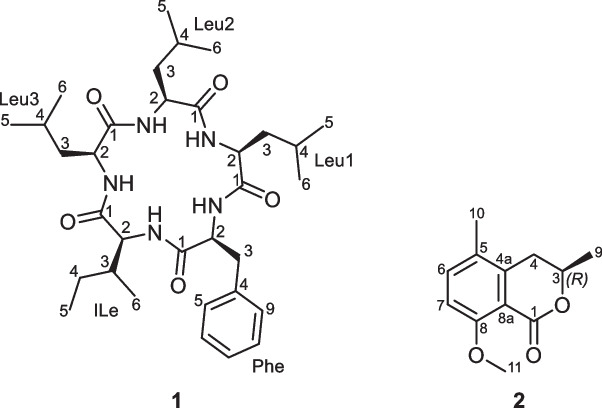 Figure 7
Figure 7- —http://dx.doi.org/10.13039/100010663H2020 European Research Council
- —http://dx.doi.org/10.13039/100021828German Academic Exchange Service
- —http://dx.doi.org/10.13039/100007224National Foundation for Science and Technology Development
- —http://dx.doi.org/10.13039/100005156Alexander von Humboldt-Stiftung
- —Helmholtz-Zentrum für Infektionsforschung GmbH (HZI) (4214)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Microbial Natural Products and Biosynthesis · Entomopathogenic Microorganisms in Pest Control
Introduction
The taxonomic revision of Biscogniauxia by Ju et al. (1998) recognized 49 taxa within the genus. Since then, the number of accepted species has significantly increased, with 118 taxa currently listed worldwide Robert et al. (2005) www.mycobank.org. Morphologically, Biscogniauxia is characterized by widely effuse stromata with separate ostioles on the surface. The perithecia are primarily arranged in a single layer, though sometimes polystichous. Asci are cylindrical, 8-spored, and may or may not possess an amyloid apical apparatus while ascospores are predominantly uniseriate, rarely biseriate, ellipsoid, and brown, with or without germ slits (Ju et al. 1998; Vasilyeva et al. 2007).
The phylogenetic relationships of Biscogniauxia and its allied genera have been firmly established within the family Graphostromataceae M.E. Barr, J.D. Rogers and Y.M. Ju, and refined by M. Stadler, L. Wendt and Sir based on multi-gene phylogenetic analyses and comprehensive macro- and microscopic features (Wendt et al. 2018; Samarakoon et al. 2022). This family includes Graphostroma Piroz., a genus characterized by its hyaline, elongated ascospores and bipartite stromata, as well as Biscogniauxia, Camillea Fr., Cryptostroma P.H. Greg. and S. Waller, Obolarina Pouzar, and Vivantia J.D. Rogers, Y.M. Ju and Cand. These morphological similarities further support their classification within Graphostromataceae (Wendt et al. 2018; Daranagama et al. 2018). These taxa display significant ecological versatility, acting as both saprotrophs and opportunistic pathogens depending on environmental conditions. This dual lifestyle underscores the ecological role of Biscogniauxia spp. in forest ecosystems, where it contributes to wood decomposition as well as plant-pathogen interactions (Ju et al. 1998).
While recent studies have started to delve into the translational potential of the ecological versatility of Graphostromataceae, particularly regarding their biotechnological relevance and secondary metabolism, the actual ecological functions of tthe molecules they produce remain poorly understood, especially in the context of plant diseases (Helaly et al. 2018; Purbaya et al. 2023). For instance, the phytotoxic properties of certain metabolites, such as biscopyran, suggest a possible role in plant–pathogen interactions and may contribute to the virulence mechanisms of Biscogniauxia species (Evidente et al. 2005). However, our understanding of the pathogenic traits in these fungi is hindered by the lack of mechanistic studies on their secondary metabolites, uncertainty about the production of these compounds across related taxa, and the absence of evidence for their involvement during infection.
As a part of our taxonomic studies on xylarialean fungi in Thailand, supported by extensive fieldwork, we identified a novel species of Biscogniauxia. This study is dedicated to its comprehensive taxonomic description, incorporating detailed macro- and microscopic features, accompanied by illustrations. Additionally, we evaluated its secondary metabolite production under different conditions, characterized its main components after preparative isolation and structure elucidation, and assessed their antimicrobial and cytotoxic properties.
Materials and methods
Sample collection and cultivation
The specimens in this study were collected from the community forests in northern Thailand. Pictures of the specimens in their natural habitat were captured using a Canon 60D digital camera (Canon, Tokyo, Japan). Pure isolates were obtained by culturing on potato dextrose agar (PDA) using the multiple spore isolation technique on the same day as field collection. After a few days of incubation, hyphal tips were excised and transferred to fresh agar plates. Pure cultures were preserved in the BIOTEC Culture Collection (BCC) and the National Biobank of Thailand (NBT), while the dried voucher specimens were deposited in the BIOTEC Bangkok Herbarium (BBH), Thailand, respectively.
Morphological characterization and temperature growth profile
Morphological features, such as stromatal size and shape, perithecia, asci, apical apparatus, and ascospores were examined in accordance with Ju et al. (1998) using a Nikon (Bangkok, Thailand) Eclipse Ni connected with a Nikon microscope camera DS-Ri2 and a stereo dissecting microscope Nikon SMZ18 (Bangkok, Thailand). The abbreviations used in the descriptions were as follows: x̄ = mean of the measurements per specimen, and n = number of measurements per specimen. Fungal cultures were grown on several media, i.e., oatmeal agar (Difco OA; Becton Dickinson, Carlsbad, CA, USA); potato dextrose agar (Difco PDA); yeast malt glucose agar (1% malt extract, 0.4% glucose, and 0.4% yeast extract; agar 1%; YMGA); and malt extract agar (2% malt extract, 1% glucose, and 0.1% peptone; agar 1%; MEA). The strains were inoculated in one point on 90 mm Petri dishes and incubated at 25 °C in darkness. Conidiogenous cells and conidiophore branching patterns of the anamorph were examined as proposed by Ju and Rogers (1998). Furthermore, the cultures colors were documented following the color chart of Rayner (1970). Cardinal temperatures were determined for Biscogniauxia papillate. The strains were grown on MEA, OA, PDA, and YMGA for 14 d at 5, 25, and 35 °C in darkness.
DNA extraction, polymerase chain reaction (PCR), and sequencing
Genomic DNA from fungal mycelia was isolated according to the methods of O’Donnell and Cigelnek (1997) and Sakayaroj et al. (2011). The internal transcribed spacer regions (ITS), large subunit of the rDNA (LSU), RNA polymerase II (RPB2), and beta tubulin (TUB2) were amplified using standard primers introduced by White et al. (1990; ITS1, ITS4, and ITS5); Vilgalys and Hester (1990; LR5 and LROR); Liu et al. (1999: RPB2–5F and 7Cr); and O’Donnell and Cigelnik (1997; T1 and T22). PCR was conducted in 25 µL reaction volume consisting of 1 × PCR buffer, 200 µM of each of the four dNTPs, 2.5 mM MgCl_2_, 1 U Taq DNA polymerase recombinant (Thermo Scientific, USA), 0.5 µM of each primer, and 50–100 ng of DNA template. Amplification was performed using a T100TM thermal cycler (BIO-RAD Laboratories, Inc., California) under the following conditions: 94 °C for 2 min, followed by 35 cycles of denaturation at 94 °C for 1 min, annealing at a suitable temperature for 1 min, extension at 72 °C for 2 min, and a final extension of 72 °C for 10 min. The annealing temperatures were 55 °C for ITS and LSU, 54 °C for RPB2, and 53 °C for TUB2. PCR products were sent to Macrogen Co. (Seoul, Korea) for purification and sequencing using the same primers as in the PCR amplification. DNA sequences were checked and assembled using BioEdit v. 7.2.5 (Hall 1999) and AliView v. 1.28 (Larsson, A. 2014). All newly generated sequences were submitted to GenBank (https://www.ncbi.nlm.nih.gov/) and listed in Table 1. Table 1. Taxa used in the phylogenetic analyses and their corresponding GenBank accession numbersSpeciesStrain numberStatusGenBank accession numbersReferencesITSRPB2TUB2LSUAmphirosellinia fushanensisHAST 91111209holotypeGU339496GQ848339GQ495950N/A Hsieh et al. (2010) Amphirosellinia nigrosporaHAST 91092308holotypeGU322457GQ848340GQ495951N/A Hsieh et al. (2010) Astrocystis concavisporaMFLUCC 14–0174isotypeKP297404KP340532KP406615KP340545 Daranagama et al. (2015) Barrmaelia oxyacanthaeCBS 142770MF488988MF488997MF489016N/A Voglmayr et al. (2018) Barrmaelia rhamnicolaCBS 142772epitypeMF488990MF488999MF489018N/A Voglmayr et al. (2018) Biscogniauxia ancepsYMJ 123authorityEF026132JX507777AY951671N/A Hsieh et al. (2010) Biscogniauxia arimaYMJ 122IsotypeEF026150GQ304736AY951672N/A Hsieh et al. (2010) Biscogniauxia capnodesYMJ 138authorityEF026131JX507779AY951675N/A Hsieh et al. (2010) Biscogniauxia citriformisYMJ129authorityJX507801JX507781AY951678N/A Mirabolfathy et al. (2013) Biscogniauxia citriformisYMJ88113012authorityJX507800JX507780AY951677N/A Mirabolfathy et al. (2013) Biscogniauxia cylindrisporaYMJ 89092701authorityEF026133JX507782AY951679N/A Hsieh et al. (2010) Biscogniauxia formosanaYMJ89032201authorityJX507802JX507783AY951680N/A Mirabolfathy et al. (2013) Biscogniauxia glaucaeGMBC0007HolotypeMT624046MT622652MT622654N/A Li et al. (2021)) Biscogniauxia glaucaeGMBC0029MT624047MT622653MT622655N/A Li et al. (2021)) Biscogniauxia granmoiYMJ 135authorityJX507803JX507784AY951681N/A Mirabolfathy et al. (2013) Biscogniauxia latirimaYMJ 90080703authorityEF026135JX507786AY951683N/A Mirabolfathy et al. (2013) Biscogniauxia latirimaYMJ89101101authorityJX507804JX507785AY951682N/A Mirabolfathy et al. (2013) Biscogniauxia marginataCBS 124505-KU684016KU684310KU684124N/A U’Ren et al. (2016) Biscogniauxia mediterraneaYMJ 147authorityEF026134GQ844765AY951684N/A Hsieh et al. (2010) Biscogniauxia mediterraneaAZ0703-HM123416KU684217KU684122N/A U’Ren et al. (2016) Biscogniauxia nummulariaCBS 969.70-MH860015KU684281KU684125N/A Vu et al. (2019) Biscogniauxia nummulariaMUCL 51395epitypeNR_153649KY624236KX271241KT281894 Wendt et al. (2018) Biscogniauxia papillataBCC 36828holotypePQ586363****PQ604605N/APQ586365****This studyBiscogniauxia papillata**BCC 83050authorityPQ586364**PQ604606N/APQ586366**This studyBiscogniauxia petrensisLC5697-KU746669KY883231KU746761KU746715 Zhang et al. (2017) Biscogniauxia philippinensis var. microsporaYMJ 89041101authorityEF026136JX507787AY951685N/A Hsieh et al. (2010) Biscogniauxia repandaATCC 62606-KY610383KY624237KX271242KY610428. Wendt et al. (2018) Biscogniauxia rosacearumBx26typeKT253493N/AKT253527N/A Raimondo et al. (2016) Biscogniauxia simpliciorYMJ 136authorityEF026130JX507788AY951686N/A Hsieh et al. (2010) Biscogniauxia uniapiculataYMJ90080608authorityJX507805JX507789AY951687N/A Mirabolfathy et al. (2013) Camillea broomeanaGMB0218MW854657GMB0218MW855491MW854663. Li et al. (2021)) Camillea obulariaATCC_28093-KY610384KY624238KX271243KY610429 Wendt et al. (2018) Camillea sp.MFLU 18–0786-MW240614MW342618N/AMN244210Samarakoon,M.C submitted directly.Camillea tinctorYMJ363authorityJX507806JX507790JX507795OQ871479Hsieh and Ju (direct submission)Camillea tinctorCBS 203.56-KU683753KU684282N/AN/A U’Ren et al. (2016) Clypeosphaeria mamillanaCBS 140735epitypeKT949897MF489001MH704637MH554225 Jaklitsch and Gardiennet (2016) Collodiscula bambusaeGZU H0102holotypeKP054279KP276675KP276674KP054280 Li et al. (2015) Collodiscula japonicaCBS 124266-JF440974KY624273KY624316MH874889 Jaklitsch and Voglmayr (2012; ITS, LSU), Wendt et al. (2018; RPB2,TUB2) Cryptostroma corticaleAcer7-HG934114HG934119HG934105N/A Koukol et al. (2015) Cryptostroma corticaleCBS 216.52-MH857008HG934116HG934102N/A Vu et al. (2019) Cryptostroma corticaleCBS 217.52-HG934111HG934117HG934103N/A Koukol et al. (2015) Cryptostroma corticaleCBS 218.52-HG934112HG934118HG934104N/A Koukol et al. (2015) Entosordaria perfidiosaCBS 142773epitypeMF488993MF489003MF489021N/A Voglmayr et al. (2018) Entosordaria quercinaCBS 142774holotypeMF488994MF489004MF489022N/A Voglmayr et al. (2018) Graphostroma guizhouenseGMBC0219holotypeMW854659MW855487MW855490MW854664 Li et al. (2021) Graphostroma guizhouenseGMBC0008-MW854658MW855486MW855489MW854662 Li et al. (2021) Graphostroma platystomumCBS 270.87typeJX658535DQ836893HG934108DQ836906 Stadler et al. (2014; ITS), Zhang et al. (2017; LSU), Koukol et al. (2015; TUB2), Wendt et al. (2018; RPB2) Graphostroma platystomumCBS:146066-MT223799MT223680MT223734N/A Hsieh et al. (2010) Hypocreodendron sanguineumJ.D.R. 169-GU322433GQ844819GQ487710N/A Hsieh et al. (2010) Hypoxylon pulicicidumMUCL49879holotypeJX183075KY624280JX183072KY610492. Bills et al. (2012; ITS, TUB2), Wendt et al.(2018; LSU, RPB2) Hypoxylon rickiiMUCL 53309epitypeKC968932KY624281KC977288N/A Kuhnert et al. (2014; ITS, TUB2), Wendt et al.(2018; LSU, RPB2) Kretzschmaria deustaCBS 163.93holotypeKC477237KY624227KX271251KY610458 Stadler et al. (2013, ITS), Wendt et al. (2018, LSU, RPB2, TUB2) Linosporopsis ischnothecaLIF1epitypeMN818952MN820708MN820715N/A Voglmayr and Beenken (2020) Linosporopsis ochraceaLIO3epitypeMN818958MN820714MN820721N/A Voglmayr and Beenken (2020) Nemania bipapillataHAST 90080610-GU292818GQ844771GQ470221N/A Hsieh et al. (2010) Nemania primoluteaHAST 91102001holotypeEF026121GQ844767EF025607N/A Hsieh et al. (2010) Obolarina dryophilaMUCL 49882-GQ428316KY624284GQ428321GQ428316 Pažoutová et al. (2010); Wendt et al. (2018; RBB2) Obolarina dryophilaH76-GQ428317N/AGQ428323N/APažoutová et al. (2010)Obolarina sp.YMJ 1461-JX507807JX507792JX507796N/AMirabolfathy M et al. (2013)Podosordaria mexicanaWSP 176-GU324762GQ853039GQ844840N/A Hsieh et al. (2010) Poronia pileiformisWSP 88113001epitypeGU324760GQ853037GQ502720N/A Hsieh et al. (2010) Rosellinia aquilaMUCL 51703-KY610392KY624285KX271253KY610460 Wendt et al. (2018) Dematophora buxiJ.D.R. 99-GU300070GQ844780GQ470228N/A Hsieh et al. (2010) Stilbohypoxylon quisquiliarumY.M.J. 172-EF026119GQ853020EF025605N/A Hsieh et al. (2010) Xylaria adscendensJ.D.R. 865-GU322432GQ844818GQ487709N/A Hsieh et al. (2010) Xylaria bambusicolaWSP 205holotypeEF026123GQ844802AY951762N/A Hsieh et al. (2010) Sarcoxylon compunctumCBS 359.61-KT281903KY624230KX271255KY610462 Senanayake et al. (2015; ITS), Wendt et al.(2018; LSU, RPB2, TUB2) Xylaria hypoxylonCBS 122620-KY610407KY624231KX271279KY610495 Sir et al. (2016; TUB2), Wendt et al. (2018; ITS, LSU, RPB2) Xylaria hypoxylon95082001 (HAST)-GU300095GQ844811GQ487703N/A Hsieh et al. (2010) New taxa proposed in this study are in bold.
Molecular phylogenetic inference
All sequences were aligned using Multiple Sequence Comparison by Log-Expectation (MUSCLE) (Edgar 2004) and manually refined. Multiple sequence alignments were analyzed together with closely matched sequences and other reference taxa obtained from GenBank as shown in Table 1. Phylogenetic analyses were performed using maximum likelihood (ML) and Bayesian algorithm (MB). The ML tree and bootstrap analyses were conducted through the CIPRES Science Gateway V. 3.3 (Miller et al. 2010) using RAxML 8.2.4 (Stamatakis 2014) with the BFGS method to optimize GTR rate parameters. Bayesian posterior probabilities of the branches were estimated using MrBayes 3.0B4 (Huelsenbeck and Ronquist 2001) with the best-fit model (GTR + I + G) selected by AIC in Mr Modeltest 2.2 (Nylander 2004) and tested using hierarchical likelihood ratio tests (hLRTs). Three million generations were run in four Markov chains, sampling every 100 generations, with a burn-in value set at 5000 sampled trees. Sequences of Hypoxylon pulicicidum (MUCL49879) and Hypoxylon rickii (MUCL 53309) were used as outgroups. Table 2. Comparison of morphological features among Biscogniauxia species closely related to B. papillata and other similar speciesTaxaAscosporePeritheciaOstiolesAnamorphB. capnodes (Berk.) Y.M. Ju and J.D. Rogers, Mycotaxon 66: 23. 1998.8.5–15 × 5–7.5 µmObovoid to tubular, 0.4–0.8 mm high., 0.2–0.4 mm diamPunctate and usually surrounded by slightly raised rim, sometimes overlain with white substance.Periconiella-likeB. papillata********(12–) 13–15 (–16) × (7–) 8–9Numerous individuals arranged in rosettes and discharging through a single ostiolar 0.6–1 mm high, 0.2–0.3 mm**Papillate, without surrounded by a slightly raised rimPericoniella-likeB. plana (Petch) Y.M. Ju and J.D. Rogers, Mycotaxon 66: 48. 1998.11–14 × 5.5–7.5Numerous individuals arranged in rosettes and discharging through a single ostiolar 0.3–0.5 mm high, 0.05 (–0.1) mm diamSlightly lower than to slightly higher than stromatal surface, with openings punctate or slightly papillatePericoniella-likeB.communapertura Y.M. Ju and J.D. Rogers, Mycotaxon 66: 31. 19988–11.5 × 4.5–6Numerous individuals arranged in rosettes and discharging through a single ostiolar canal, 0.3–0.5 high, 0.1–0.2 diamWidely spaced, higher than stromatal surface, with openings coarsely papillateN/aB. capnodes var. limonispora* Y.M. Ju and J.D. Rogers, Mycotaxon 66: 26. 1998.11.5–14 × 6–7.5Obovoid to tubular, 0.4–0.8 mm high, 0.2–0.4 mm diamPunctate and usually surrounded by slightly raised rim, sometimes overlain with white substancePericoniella-likeB. capnodes var. limonispora Y.M. Ju and J.D. Rogers Mycotaxon 66: 26. 1998. Specimens reported from Martinique (French West Indies) Fournier et al. (2017)(11.7–) 12.8–14.5 (–15) × (7.3–) 7.7–8.9 (–9.3)Individual ostioles, two adjacent perithecia at times sharing a common ostiole. 0.3–0.5 mm high × 0.1–0.25 diamPunctate, surrounded by a slightly raised rim, 25–70 µm diam, inconspicuous, evenly distributed, at times plugged with greyish to whitish substancePericoniella-likeB. nummularia (Bull.: Fr.) Kuntze, Revis. Gen. Pl. 2: 398. 1891.10–13 (–14) × (6.5–) 7.5–8.5Obovoid, 0.3–0.5 mm diam, 0.5–0.7 mm high.Higher than stromatal surface with openings slightly papillate, or lower than stromatal surface with openings punctate and usually surrounded by slightly raised rimPericoniella-likeB. anceps (Sacc.) J.D. Rogers, Y.M. Ju and Cand., Mycol. Res. 100: 669. 1996.(13–) 14–18 (–19) µm total length × 7–9 (–10) µm broad at the broadest part, the larger cell 10–13 µm long and the smaller cell 5–7 µm long × 4–7 µm broad.0.2–0.4 mm diamUmbilicate, located in grey depressed areasPericoniella-like*This study
General analytical procedures
Electrospray mass (ESI–MS) spectra were recorded with an UltiMate® 3000 Series UHPLC system (Thermo Fisher Scientific, Waltham, MA, USA) connected to an amazon speed® ESI-Ion Trap-MS (Bruker, Billerica, MA, USA) mass spectrometer. A C_18_ Acquity® UPLC BEH column (2.1 × 50 mm, 1.7 µm; Waters, Milford, MA, USA) was used as stationary phase. HPLC parameters were set as follows: solvent A: H_2_O + 0.1% formic acid, solvent B: acetonitrile (MeCN) + 0.1% formic acid; gradient 5% B for 0.5 min, increasing to 100% B over 19.5 min, followed by 5 min at 100% B; flow rate 0.6 mL/min, with diode array (DAD) detection in the range of 200–600 nm.
High-resolution electrospray ionization mass spectrometry (HR-ESI–MS) spectra were obtained using an Agilent 1200 Infinity Series HPLC (Agilent Technologies, Santa Clara, CA, USA) connected to a maXis® Electrospray Time-of-flight mass spectrometer (ESI-TOF–MS; Bruker). The HPLC conditions were the same as for ESI–MS measurements. NMR spectra were recorded with an Avance III 600 spectrometer (Bruker, ^1^H NMR: 600 MHz, ^13^C NMR: 150 MHz) in deuterated methanol. Optical rotation was measured with a MCP 100 circular polarimeter (Anton Paar, Graz, Austria), and UV/Vis spectra were acquired using a UV-2450 spectrophotometer (Shimadzu, Kyoto, Japan). Electronic circular dichroism (ECD) spectra were acquired using a J-815 spectropolarimeter (JASCO, Pfungstadt, Germany).
Fungal cultivation and isolation of secondary metabolites
For scaled-up cultivation of BCC 36828, this fungus was first grown in YM agar at 23 °C. The seed culture was done in 250-mL flasks each containing 50 mL semi-viscous SMYA medium (Serrano et al. 2017) (maltose 40 g/L, yeast extract 10 g/L, meat peptone 10 g/L, agar 4 g/L). Subsequently, inoculation was done by adding five pieces (ca. 25 mm 2 each) of a well-grown agar-plate to each flask. Seed cultures were incubated for 5 days on a shaker (23 °C, 230 rpm). Afterwards, 6 mL of the seed culture were transferred into the new 500-mL conical flasks, containing the solid rice culture medium (28 g brown rice and 100 mL of base liquid medium). The base liquid medium consisted of yeast extract (1 g/L), di-sodium tartrate dihydrate (0.5 g/L), and KH₂PO₄ (0.5 g/L). Finally, solid cultures were incubated for 15 days in the dark at 23 °C without shaking.
For secondary metabolite extraction, the mycelia on the rice were covered with acetone and sonicated for 30 min at 40 °C. The acetone extract was separated from the mycelia using cellulose filter paper (MN 615 1/4 Ø 185 mm, Macherey–Nagel GmbH & Co. KG, Düren, Germany). The extraction and filtration steps were repeated twice, and the obtained acetone phases were combined and evaporated under reduced pressure at 40 °C (evaporator: Heidolph Instruments GmbH & Co. KG, Germany; pump: Vacuubrand GmbH & Co. KG, Wertheim am Main, Germany) to obtain an aqueous residue. The remaining aqueous residue was dispersed in 1 L of H_2_O and extracted twice, using 1 L of ethyl acetate. The ethyl acetate phases were combined and evaporated to dryness under reduced pressure at 40 °C to yield 547.33 mg of the crude extract.
For the isolation of cyclobiscognioxin A (1), the obtained crude extract obtained was portioned to 2 × 270 mg and purified using a PLC 2250 preparative HPLC system (Gilson, Middleton, WI, USA) with a Gemini C_18_ (50 × 250 mm, 10 µm; Phenomenex®, Torrance, CA, USA as stationary phase. The HPLC conditions were as follows: solvent A: H_2_O + 0.1% formic acid, solvent B: MeCN + 0.1% formic acid; flow rate: 50 mL/min, gradient: isocratic conditions at 5% B for 5 min, followed by an increase to 55% B in 50 min, then increasing to 100% B in 5 min, and ending with isocratic conditions at 100% B for 10 min. This yielded 1 (1.73 mg, tR = 18 min) and 2 (0.9 mg, tR = 38 min).
Cyclobiscognioxin A (1): White powder; UV–Vis (MeOH): λ_max_ 230 nm; HR-ESI–MS: m/z 600.4118 [M + H]^+^ (calcd. 600.4125 for C_33_H_54_N_5_O_5_^+^), 622.3948 [M + Na]^+^ (calcd. 622.3944 for C_33_H_53_N_5_NaO_5_^+^); ^13^C/^1^H NMR data (methanol-d4, 150 and 600 MHz): see Table S1 and comparable to those reported in literature (Li et al. 2004); C_33_H_53_N_5_O_5_ (599.82 g mol^−1^).
3,5-Dimethyl-8-methoxy-3,4-dihydroisocoumarin (2): Colorless amorphous solid; UV–Vis (MeOH): λ_max_ 210, 247, 316 nm; HR-ESI–MS: m/z 207.1017 [M + H]^+^ (calcd. 207.1021 for C_12_H_15_O_3_^+^), 229.0837 [M + Na]^+^ (calcd. 229.0841 for C_12_H_14_NaO_3_^+^); ^1^H NMR data (methanol-d4, 600 MHz): comparable to those reported in literature (Kokubun et al. 2003); ^13^C NMR data (methanol-d4, 150 MHz): δ_C_ 161.5 (C-1), 158.6 (C-8), 140.3 (C-5), 135.3 (C-6), 125.9 (C-4a), 113.2 (C-8a), 110.6 (C-7), 72.8 (C-3), 55.5 (C-11), 32.1 (C-4), 20.2 (C-9), 17.8 (C-10); C_12_H_14_O_3_ (206.24 g mol^−1^).
Biological activities
The evaluation of the antimicrobial and cytotoxic properties from the isolated compounds was performed following the methods reported by Charria-Girón et al. (2025).
Results and discussion
Taxonomy
Biscogniauxia papillata Srikit., Wongkan., Charria-Girón and Luangsa-ard, sp. nov.
Figures 1, 2, and 3.Fig. 1. Stromata of Biscogniauxia papillata (BBH 25799) on the bark of an unidentified host tree. Stromata effused-pulvinate to irregular in outline, black, carbonaceous, solitary to coalescent, and partially embedded in the bark. Margins indistinct or lobed. Scale bar = 20 mmFig. 2Biscogniauxia papillata (BBH 25799). a, d Stroma in longitudinal section showing the thick carbonaceous subsurface, the perithecia and the indistinct subperithecial tissue. b Shiny black papillate ostioles with truncate top. c Ostioles without the external surface. e Cross-section of the stroma showing rosette arrangement of perithecia and the presence of sterile tissue between them (white arrow). f Ascus. g–j Apical apparatus in Melzer’s reagent (white arrows). k Apical apparatus in 10% KOH reagent (white arrow). l Ascospore in 10% KOH reagent, showing conspicuous germ slits (white arrow). m–o Ascospores in distilled water, showing germ slits (white arrows). p–u Ascospores in distilled water. Scale bars: a–b,** d**–e 0.5 mm, c 100 µm, f 20 µm, g–u 10 µmFig. 3Biscogniauxia papillata (BCC 36828) a–b Conidiophores showing compact fertile head (arrow indicating melanized main axis) morph showing conidiophores with virgariella-like to nodulisporium-like branching patterns. c Fertile head with densely packed branchlets bearing conidiogenous cells (arrow). d Conidiogenous cells, cylindrical, hyaline, finely roughened, bearing inconspicuous collarettes (arrow). e Conidia, hyaline, smooth-walled, ellipsoid, often with flattened base (arrow). Scale bars: a–b = 40 µm; c = 20 µm; d–e = 10 µm
Mycobank MB 857465.
Etymology: “papillata,” referring to the ostioles of the new species having papillae or nipple-like or rounded projections on the stromatal surface*.*
Diagnosis: Differs from related Biscogniauxia species by having perithecia in clusters of 2–4 sharing a common ostiolar canal, papillate ostioles lacking a white rim, larger flask-shaped perithecia, and broadly ellipsoid, dark brown ascospores with a straight, full-length germ slit.
Holotype: Thailand: Chiang Rai Province, Phu Kaeng Waterfall, 19.443° N, 99.670° E, on decaying wood, 18 January 2012, P. Srikitikulchai, (BBH 25799), (ex-BCC 36828); DNA sequences of ex-holotype strain ITS– PQ586363, LSU– PQ586365, RPB2–PQ586365, TUB2–N/A.
Teleomorph: Stromata effused-pulvinate or irregular in shape, solitary to coalescent and distinctly bipartite, solitary to coalescent,10–120 mm long, 10–30 mm broad, 1.63–1.77 mm thick (Fig. 1); surface brown to blackish brown or black 0.035–0.071 mm thick (x̅ = 0.048 mm; n = 10), carbonaceous forming a thin crust above perithecia layer 0.26–0.47 mm thick (x̅ = 0.36 mm; n = 10) (Fig. 2a, d); the tissue between perithecia brownish, soft-textured; tissue beneath perithecia gray soft-textured and a blackish line separates the stromatal tissue from the host surface 0.14–0.35 mm, and the stromatal tissue is almost inseparable from the host substrate. Perithecia narrowly cylindrical to clavate, obpyriform to flask-shaped (lageniform) (0.76–)0.90–0.97(–1.01) × (0.18–)0.22–0.30(–0.44) mm (x̅ = 0.94 × 0.28 mm; n = 25). Ostioles papillae or nipple-like or rounded projections on the stromatal surface (Fig. 2b–c). Asci cylindrical, the spore-bearing parts 82–89 × (6–)7–9 µm, the total length (86–)105–123 µm, with apical apparatus 4–5 × 1–2 µm (x̅ = 4.48 × 2.02 µm; n = 10), bluing in Melzer’s reagent. Ascospores broadly ellipsoid, inequilateral, (12–)13–15(–16) × (7–)8–9 µm (x̅ = 13.85 × 8.26 µm; n = 25), dark brown, smooth-walled, with a straight, full-length germ slit on the convex side.
Culture characteristics: Colonies on PDA, reaching the edge of the Petri dish in 1 week, azonate, at first whitish becoming Umber (9); reverse Umber (9) and Chestnut (40). Colonies on OA, reaching the edge of the Petri dish in 1 week, aerial mycelium at first whitish becoming Pale Olivaceous Grey (120) and Olivaceous Grey (121); reverse Olivaceous Grey (121). Colonies on YMGA reaching the edge of the Petri dish in 1 week, azonate, at first whitish becoming Umber (9); reverse Umber (9) and Chestnut (40). Colonies on MEA reaching the edge of the Petri dish in 1 week, azonate, at first whitish becoming Umber (9); reverse olivaceous (46) and Umber (9).
Anamorph: On OA conidiophores structure with periconiella-like to nodulisporium-like branching patterns, as described by Ju and Rogers (1996)
Conidiophores (Fig. 3a–b) erect, straight to slightly flexuous, unbranched along most of their length, 200–300 µm long, 5–6 µm wide, septate, with thickened walls; the main axis is melanized, brown to blackish brown, with a surface ranging from smooth to roughened. The fertile region is apical, composed of a dense cluster of short, closely packed branchlets bearing conidiogenous cells.
Conidiogenous cells (Fig. 3c–d) phialidic, cylindrical, hyaline, finely roughened, (7–)9.8–19.7 × 2.6–5.2 µm (x̅ = 14.37 × 3.62 µm; n = 25), arranged in whorls or irregular clusters at the tips of branchlets; collarettes inconspicuous to short, flaring slightly. Conidia are produced holoblastically in a sympodial sequence from the apical region of each conidiogenous cell.
Conidia (Fig. 3e) hyaline, smooth-walled, aseptate, ellipsoid, (5–)5.8–6.9(–7.7) × 2.3–5.2 µm (x̅ = 6.15 × 3.12 µm; n = 25), often with a flattened base marking the point of attachment to the conidiogenous cell; accumulating in slimy masses at the apex of the fertile head.
Cardinal temperatures: Biscogniauxia papillata grows at 10 °C but not at 5 °C, with optimum growth at 25 °C. Growth is restricted at 35 °C (Figs. 4, 5). After incubating cultures at each test temperature for 14 days, those subsequently returned to the optimum temperature resumed normal growth in all cases, indicating that the species can survive suboptimal thermal conditions and rapidly recover when favorable temperatures are restored. Such thermal resilience suggests that B. papillata may be well adapted to withstand short-term temperature extremes predicted under climate change scenarios, enabling persistence in host tissues during unfavorable periods and rapid colonization when conditions improve, a strategy also reported for other members of the genus (Granata and Sidoti 2004).Fig. 4. Temperature growth profile of the newly described species after 7 d of incubation on MEA, OA, PDA and YMGA at 5 °C, 25 °C, and 35 °C. Normal mycelial growth resumed at 25 °C after incubation under different temperatures 14 daysFig. 5Temperature growth profile of the newly described species after 6 d of incubation on MEA, OA, PDA and YMGA (reverse) at 5 °C, 25 °C, and 35 °C. Normal mycelial growth resumed at 25 °C after incubation under different temperatures 14 days
Secondary metabolites: Cyclobiscognioxin A (1), and 3,5-dimethyl-8-methoxy-3,4-dihydroisocoumarin (2) (Fig. 7).
Other material examined: Thailand: Phitsanulok Province, Ban Phaothai Community Forest, 16.733° N, 100.658° E, hill evergreen forest on decaying wood, 22 November 2016, P.S. and S.W. (BCC83050).
Notes
Morphologically, Biscogniauxia papillata resembles B. communapertura Y.M. Ju and J.D. Rogers in producing numerous perithecia arranged in rosettes (Fig. 2a and e) and discharging through a single ostiolar canal, with both species also exhibiting papillate ostioles (Fig. 2b–c). In B. papillata, two perithecia frequently share a common ostiole (Fig. 2a–b). However, the new species differs from B. communapertura in ascospore morphology, being brown to dark brown and significantly larger [(12–)13–15(–16) × (7–)8–9 µm vs. 11.5 × 4.5–6 µm in B. communapertura] (Table 2).
Our fungus also resembles B. plana (Petch) Y.M. Ju and J.D. Rogers in having numerous perithecia arranged in rosettes and discharging through a single ostiolar canal, but differs by its larger, flask-shaped perithecia [(0.76–)0.90–0.97(–1.01) × (0.18–)0.22–0.30(–0.44) mm vs. 0.50 × 0.90–0.97 mm in B. plana] and larger ascospores [(12–)13–15(–16) × (7–)8–9 µm vs. 11–14 × 5.5–7.5 µm].
Biscogniauxia papillata shares several features with B. capnodes var. limonispora Y.M. Ju and J.D. Rogers from Thailand (Ju et al. 1998), including the general stromatal habit. It differs, however, in having papillate ostioles on the stromatal surface, larger ascospores [dark brown, ellipsoid to slightly inequilateral, (12–)13–15(–16) × (7–)8–9 µm vs. 11.5–14 × 6–7.5 µm], and multiple perithecia discharging through a common ostiolar canal. The anamorph also differs: B. papillata exhibits a periconiella-like to nodulisporium-like branching pattern in its conidiogenous structures, with the nodulisporium-like form occurring much more frequently, whereas B. capnodes var. limonispora has a strictly periconiella-like anamorph. Specimens of B. capnodes var. limonispora from Martinique (Fournier et al. 2017) resemble our collections in gross morphology but differ in ostiolar structure, with B. papillata lacking the white ring surrounding the ostioles.
Biscogniauxia petrensis Z.F. Zhang, F. Liu and L. Cai produces conidia of similar length [4.5–7.5 × 2.5–4.5 µm vs. (4–)5–6(–7) × (2–)3–4 µm in B. papillata], but multi-locus phylogenetic analyses clearly separate the two species with high support (100/1.00), despite their genetic proximity (Fig. 6). Multi-locus phylogenetic analyses also clearly distinguish B. papillata, B. capnodes (Berk.) Y.M. Ju & J.D. Rogers, B. nummularia (Bull.) Kuntze, and B. anceps (Sacc.) J.D. Rogers, Y.M. Ju and Cand. into distinct, well-supported lineages.Fig. 6. Phylogenetic relationships inferred from RAxML on multi-locus alignment of Biscogniauxia papillata and other selected Xylariales based on concatenated ribosomal (ITS and LSU) and proteinogenic (TUB2 and RPB2) DNA sequence data. Support values from ML and MB analyses higher than 50% ML and 70% MB are shown above the respective branches. Branches of significant support bootstrap (BS) 70% and posterior probability (PP) 95% are thickened. The sequences of B. papillata are highlighted in orange font. ET (ex-epitype), IT (ex-isotype), HT (ex-holotype), PT (ex-paratype), and T (ex-type) strains are highlighted in bold letters
In terms of morphology, B. papillata differs from B. capnodes in having larger, flask-shaped perithecia [(0.76–)0.90–0.97(–1.01) × (0.18–)0.22–0.30(–0.44) mm vs. 0.4–0.8 × 0.2–0.4 mm], papillate ostioles lacking the white rim sometimes present in B. capnodes, and larger asci [(86–)105–123 µm vs. 70–115 µm] with a more robust apical apparatus [4–5 × 1–2 µm vs. 1.5–4 × 3.5–4 µm]. Its ascospores are broadly ellipsoid, often inequilateral, and larger [(12–)13–15(–16) × (7–)8–9 µm vs. 8.5–15 × 5–7.5 µm], and the anamorph is predominantly nodulisporium-like, with the periconiella-like branching patterns encountered less frequently. The frequent occurrence of two perithecia sharing a common ostiolar canal, absent in B. capnodes, is a consistent diagnostic feature.
Biscogniauxia papillata is also readily distinguished from B. nummularia by stromatal morphology and ascospore form. Biscogniauxia nummularia produces applanate stromata with woody, greyish brown to dark brown tissue between the perithecia, whereas B. papillata has distinctly bipartite, effused-pulvinate stromata with softer-textured grey tissue beneath the perithecia and a blackish line demarcating the stromatal base from the substrate. The ascospores of B. nummularia are smaller, nearly equilateral, and more rounded [10–13(–14) × (6.5–)7.5–8.5 µm], contrasting with the larger, broadly ellipsoid and often inequilateral ascospores of B. papillata. Biogeographically, B. nummularia is largely restricted to Europe, B. capnodes is rarely encountered there (Ju et al. 1998), and B. papillata is presently known only from Thailand.
Biscogniauxia anceps is clearly distinct from B. papillata both molecularly and morphologically. It forms applanate, discoid to widespreading stromata with distinct margins and a cracked surface reminiscent of Diatrype stigma (Ju et al. 1998), smaller perithecia (0.2–0.4 mm diam), and umbilicate ostioles within grey depressed areas. Its ascospores are unequally two-celled, with the larger cell sometimes dark brown but most commonly hyaline, and the smaller cell always hyaline without a germ slit; in contrast, B. papillata produces uniseptate, dark brown, broadly ellipsoid ascospores with a straight, full-length germ slit, larger flask-shaped perithecia, and an anamorph that is most often nodulisporium-like, with the periconiella-like branching patterns occurring less frequently.
Molecular phylogenetic inference
After providing full taxonomic description of the newly introduced fungus from Thailand, we confirmed its phylogenetic placement using multi-locus DNA analyses as shown in Fig. 6. The six newly generated ITS, LSU, and RPB2 sequences were compared with data from GenBank NCBI nucleotide database (PCR amplifications yielded approximately 500 bp, 1000 bp, 800 bp, and 1000 bp of ITS rDNA, LSU rDNA, RPB2, and TUB2 sequences, respectively). The phylogenetic relationships were estimated using the ML and MB analyses. The dataset of the multi-locus DNA sequences included 69 taxa from Amphirosellinia (2), Astrocystis (1), Barrmaelia (2), Biscogniauxia (27), Camillea (5), Dematophora (1), Clypeosphaeria (1), Collodiscula (2), Cryptostroma (4), Entosordaria (2), Graphostroma (4), Hypocreodendron (1), Hypoxylon (2), Kretzschmaria (1), Linosporopsis (2), Nemania (2), Obolarina (3), Podosordaria (2), Rosellinia (1), Sacroxylon (1), Stilbohypoxylon (1), and Xylaria (4). The best phylogenetic tree inferred from RAxML had a likelihood of − 71969.270958. The alignment had 2969 distinct alignment patterns, with 39.98% undetermined characters or gaps. Estimated base frequencies were as follows: 0.237200, C = 0.273463, G = 0.246760, T = 0.242577; substitution rates were AC = 1.265867, AG = 3.551300, AT = 1.172200, CG = 0.892001, CT = 5.053569, GT = 1.000; gamma distribution shape parameter was α 0.340307. The likelihood of the Bayesian tree was − 82154.19. As shown in Fig. 6, the sequences of our new species are well separated from the previously reported species in Biscogniauxia that have been recently updated by Li et al. (2021). The topology of the RAxML tree is practically identical to the one presented by Wendt et al. (2018) and Samarakoon et al. (2022).
Secondary metabolite profiling and isolation
After analyzing the secondary metabolite production of B. papillata, we decided to scale up its cultivation in BRFT solid medium. Among the major components, a metabolite of medium polarity with high molecular weight was initially assumed to be related to sansalvamide A, a compound first reported from a marine Fusarium sp., with a similar derivative previously identified in B. mediterranea (Belofsky et al. 1999; Wu et al. 2016). To confirm the identity of this compound, we targeted its isolation. Consequently, compound 1 was purified as an amorphous solid, whose HR-ESI–MS spectrum established its molecular formula as C_33_H_53_N_5_O_5_ by revealing a protonated molecular ion peak at m/z 600.4118 [M + H]^+^ (calculated 600.4125 for C_33_H_54_N_5_O_5_^+^) indicating ten degrees of unsaturation. The ^13^C NMR spectral data of 1 (Table S1, Figure S4) revealed the presence of five carbonyl carbon atoms at δ_C_ 174.64, 174.58, 174.1, 174.0, and 173.9 ppm that suggested, together with its ^1^H NMR spectral data (Table S1, Figure S3), being a cyclopentapeptide derivative. The ^1^H NMR spectral data of 1 (Table S1, Figure S3) revealed five α proton signals at δ_H_ 3.29 ~ 4.67 ppm; four β-diastereotopic methylene groups at δ_H_ 1.42/1.49, 1.57/1.64, 1.56/1.70, and 2.88/3.02 ppm and one β-methine proton at δ_H_ 2.38 ppm. In addition, the ^1^H NMR spectral data of 1 also revealed five aromatic protons (δ_H_ 7.20 ~ 7.27 ppm) denoting a monosubstituted aromatic ring together with three pairs of doublet methyl groups at δ_H_ 0.99/0.95, 0.97/0.89, and 0.90/0.85; one triplet methyl at δ_H_ 0.87; and a doublet methyl group at δ_H_ 0.72 ppm. Based on the obtained results, compound 1 was suggested to be a cyclic pentapeptide consisting of one phenylalanine, three leucine and one isoleucine residues. A literature search of 1 revealed its close similarity to a previously reported cyclopentapeptide that was isolated from the methanol extract of liquid culture medium mycelia of an unidentified endophytic fungal strain no. 2524 derived from a seed of Avicennia marina, a mangrove plant in Hongkong (Li et al. 2004). A careful comparison of ^13^C/^1^H NMR data of both 1 and cyclo-(L-Phe-L-Leu^1^-L-Leu^2^-L-Leu^3^-L-Ile) (Li et al. 2004) (Table S1) revealed that they are virtually identical. The amino acid sequence of 1 was established via acquiring its HMBC spectrum that revealed key correlations from αH of Phe at δ_H_ 4.67 to the carbonyl group of Leu^1^ at δ_C_ 174.0 and αH signals of Leu^1^-Leu^3^ at δ_H_ 4.44, 4.27, and 4.21 to carbonyl groups of Leu^2^, Leu^3^, and Ile at 174.58, 174.64, and 174.1 ppm, respectively. The αH of Ile at δ_H_ 3.29 revealed key HMBC correlation to the carbonyl group of Phe at δ_C_ 173.9 thus confirming its presence as a cyclic pentapeptide of Phe-Leu^1^-Leu^2^-Leu^3^-Ile. The absolute configuration of amino acid residues was determined via Marfey’s method as previously described (Wennrich et al. 2024; Holzenkamp et al. 2024). The results of Marfey’s method (data not shown) determined that all amino acids in 1 are present in L configurations. Thus, compound 1 was unambiguously characterized as cyclo-(L-Phe-L-Leu^1^-L-Leu^2^-L-Leu^3^-L-Ile) and it was given a trivial name cyclobiscognioxin A.
Compound 2 was obtained as a colorless amorphous solid. The HR-ESI–MS spectrum of 2 revealed a protonated molecular ion and a sodium adduct at m/z 207.1017 (calculated 207.1021) and 229.0837 (calculated 229.0841), respectively. Thus, the molecular formula of 2 was determined as C_12_H_14_O_3_ indicating six degrees of unsaturation. The ^1^H NMR spectral data of 2 (Table S2, Figure S11) revealed two ortho-coupled aromatic proton signals at δ_H_ 7.40 and 6.96 ppm with a coupling constant (J value) of 8.6 Hz; an aliphatic methine at δ_H_ 4.46 (dqd, J = 11.4, 6.4, 2.8 Hz); a diastereotopic methylene group at δ_H_ 2.63/2.95 and three methyl groups including one doublet at δ_H_ 1.38 (d, J = 6.4 Hz), two singlets at δ_H_ 2.19 and 3.78 ppm. A literature search of 2 revealed its close resemblance to 3,5-dimethyl-8-methoxy-3,4-dihydroisocoumarin (aka 5-methylmellein) that was previously reported from a culture filtrate of Cytospora eucalypticola (Kokubun et al. 2003). To further confirm the depicted structure of 2 (Fig.7), its 2D NMR spectra as ^1^H-^1^H COSY, HMBC, and HSQC (Figures S12–S15) were recorded. The ^1^H-^1^H COSY spectrum (Figures S12 and S15) revealed two spin systems: one between two ortho-coupled aromatic protons (H-6/H-7) and another spin system extends a diastereotopic methylene group (H_2_−4) to a methine proton (H-3) and ends at a double methyl moiety (H_3_−9). The HMBC spectrum of 2 (Figures S13 and S15) revealed key correlations from H-6 to C-5 (δ_C_ 140.3) and C-8 (δ_C_ 158.6); from an olefinic methyl group H_3_−10 to C-4a (δ_C_ 125.9), C-5 and C-6 (δ_C_ 135.3) confirming its position at C-5. In addition, a singlet methoxy group (H_3_−11) and H-6 to C-8 (δ_C_ 158.6) and from a doublet methyl group (H_3_−9) to a methylene carbon at δ_C_ 32.1 (C-4) confirming their positions at C-8 and C-3 (δ_C_ 72.8), respectively. The absolute configuration at C-3 was determined based on analyzing the coupling constant (J value) of H-3 that indicated its existence in axial orientation while the doublet methyl group H_3_-9 to be on equatorial orientation. Hence, the absolute configuration at C-3 was assigned as (3R) configuration.Fig. 7. Chemical structures of secondary metabolites isolated from the solid cultures of Bicogniauxia papillata, cyclobiscognioxin A (1), and (3R)−3,5-dimethyl-8-methoxy-3,4-dihydroisocoumarin (2)
Overall, cyclobiscognioxin A (1) was isolated together with the previously reported mellein derivative, (3R)−3,5-dimethyl-8-methoxy-3,4-dihydroisocoumarin (5-methylmellein, 2) (Figure 7). Given the fact that several cyclic peptides are known for their toxicity and diverse ecological roles, we decided to evaluate the antimicrobial and cytotoxic properties of 1. Our assays identified cyclobiscognioxin A (1) as a potent cytotoxic agent but no antimicrobial activity against the tested microorganisms was observed (Table S3). The obtained results revealed that 1 featured the strongest cytotoxic effects against A549 (lung), KB3.1 (cervix), and MCF-7 (breast) cancer cell lines at IC_50_ values of 4.7, 5.2, and 5.5 µM, respectively.
Discussion
The genus Biscogniauxia exhibits diverse ecological roles, including saprobic, endophytic, and pathogenic behaviors on weakened or stressed hosts, as illustrated by reports of disease in oak, almond, and strawberry by some of these taxa (Yangui et al. 2021). Species in this genus are frequently associated with hardwoods, colonizing bark, wood, and decayed plant material (Ju et al. 1998). For example, B. anceps occurs on Quercus and Corylus in temperate regions, whereas tropical congeners, including the species described here, thrive in the tropical forest ecosystems (Ju et al. 1998). Related taxa can also become pathogenic under suitable conditions such as Cryptostroma corticale, a close relative within the Graphostromataceae, which causes the sooty bark disease on sycamore (Acer pseudoplatanus) (Brenken et al. 2024). Motivated by these considerations, we evaluated whether B. papillata can grow outside its optimal temperature range, recognizing that climate warming and derived drought are likely to alter distributions, fitness, and host interactions by stressing trees and ultimately increasing susceptibility to opportunistic infections (Nugent et al. 2005; Desprez-Loustau et al. 2006). During our experiments, mycelial growth was optimal at 25 °C with intact hyphal structures, but growth declined sharply at ≤ 10 °C, consistent with persistence despite reduced competitive capacity (Lan et al. 2025). Growth was also slowed above 30 °C, which coincided with the development of the pigmentation on agar media (Fig. 4), suggesting a stress-mitigation response (Venkatachalam et al. 2019). Similar adaptability has been reported in B. mediterranea, which can grow from 5 to 45 °C, with an optimum near 35 °C, and recovers rapidly when conditions improve (Henriques et al. 2015; Bakhshi-Ganje et al. 2024). This pattern indicates a broader capacity across Biscogniauxia species to exploit variable thermal regimes. Altogether, these observations support a bet-hedging strategy in which prolonged, low-activity endophytism shifted to opportunistic pathogenicity during occasional heatwaves and drought.
To place these findings in a taxonomic context, we constructed a multi-locus phylogeny, which was consistent with Ju et al. (1998) morphological studies and with recent multi-locus analyses that delimited the species in the family Graphostromataceae and its constituent genera, Biscogniauxia, Camillea, Cryptostroma, Graphostroma, Obolarina, and Vivantia (Wendt et al. 2018; Daranagama et al. 2018). In the combined-sequence analyses, our new collections form a well-supported clade within the lineage that includes B. capnodes and closely related taxa, reinforcing their placement in Biscogniauxia sensu lato. Our phylogenetic analyses further confirmed the distinction between B. capnodes and B. nummularia and were consistent with Ju et al.’s (1998) morphological studies that differentiated these species based on stromatal characteristics and ascospore morphology. Biscogniauxia capnodes is characterized by carbonaceous tissue between the perithecia and ellipsoid ascospores, whereas B. nummularia possesses grey to brown woody tissue between the perithecia and more rounded ascospores Ju et al. (1998). Geographically, B. nummularia is primarily found in Europe, while B. capnodes is rarely recorded in the region (Ju et al. 1998). Similarly, B. anceps is phylogenetically distinct from our new species, consistent with their morphological concept. Both differ in their ascospore structure and their ecology. While B. papillata has coalescent stromata, flattened perithecia with shared ostioles, and larger, lemon-shaped ascospores, B. anceps has smaller, two-celled ascospores, with the larger cell measuring 10–13 µm and the smaller cell 5–7 µm, typically ellipsoid to obovate in shape. In addition, B. anceps is primarily distributed in temperate regions, including Italy and France (Ju et al. 1998). Biscogniauxia papillata is closely related to B. capnodes, which agrees with their well-defined morphological features. However, our new species is distinguished by its smaller, ellipsoid-lobed, coalescent stromata with a dehiscing layer, carbonaceous tissue beneath the perithecia, and flattened perithecia with some sharing a common ostiole, characteristics rarely found in B. capnodes. Additionally, B. papillata exhibits larger, lemon-shaped ascospores with slightly pinched ends and a spore-length germ slit, giving this fungus a unique identity among its relatives. According to the phylogenetic analyses, B. papillata is clearly separated from B. petrensis. The latter was first reported in its anamorphic state and shown to form a sister group with B. capnodes, as the species was isolated from rock in a karst cave in China (Zhang et al. 2017). While the conidial features of our new species resemble those of B. petrensis, samples isolated from the roots of Dendrobium harveyanum in Thailand (Ma et al. 2020) were morphologically similar to those from karst caves in China. However, their distinction is supported by our molecular phylogeny, which provides strong support from both RAxML and MrBayes analyses.
Molecular phylogenetic analyses clearly separate our new species from Camillea, which, as defined by Læssøe et al. (1989) and further studied by Fournier et al. (2023), is characterized by highly carbonaceous stromata erumpent through bark with a fleeting ectostromatic layer of mixed bark and fungal tissues, asci bearing a massive rhomboid amyloid apical apparatus, subhyaline to yellowish ornamented ascospores visible under SEM, and a xylocladium-like anamorph. In contrast, B. papillata develops bipartite, effused-pulvinate stromata without an ectostromatic layer, asci with a smaller discoid apical apparatus, dark brown smooth-walled ascospores, and an anamorph that is predominantly nodulisporium-like with a less common periconiella-like form, and is consistently resolved within Biscogniauxia with strong statistical support.
The isolation of compound 1 as the major secondary metabolite of B. papillata is particularly relevant for the study of related species, especially given its observed in vitro toxicity in our assays. Cyclobiscognioxin A (1) showed potent, dose-dependent cytotoxicity with no detectable antimicrobial activity against the test pane l. This bioactivity profile aligns with hydrophobic cyclic pentapeptides that act in eukaryotic cells, often transiting membranes as “molecular chameleons” and defying small-molecule rule-sets (Buckton et al. 2021). Given the latent pathogenicity observed in some Biscogniauxia species and their parasitic status, the production of toxic metabolites might represent adaptive mechanisms associated to host interactions, competition, or environmental stress responses (Patejuk et al. 2022; Nugent et al. 2005). Cyclic peptides often exhibit bioactive properties, including cytotoxic, antifungal, and antibacterial effects (Ribeiro et al. 2022), suggesting roles in shaping fungal community dynamics and host–pathogen interactions. Destruxins, produced by Metarhizium species, contribute to fungal virulence against insect hosts by suppressing immune responses (Kobmoo et al. 2024), while beauvericins from Fusarium exhibit cytotoxic and antimicrobial activities that might influence fungal competition and pathogenicity (Logrieco et al. 1998). We recently showed in Cordyceps spp. that diverse cyclic peptides, including beauveriolides and beauvericins, accumulate in insect cadavers, supporting a role during infection (Charria-Girón et al.2023). Future chemotaxonomic studies within Graphostromataceae are essential to uncover the cryptic chemistry of this fungal family. A detailed evaluation of the biological properties of cyclobiscognioxin A (1) and related compounds is required to clarify their ecological roles and potential as a pathogenicity factor.
Recent phylogenomic studies have provided deeper insights into the evolutionary dynamics of Biscogniauxia, revealing extensive gene duplications and horizontal gene transfers (HGTs), particularly in relation to secondary metabolite gene clusters (SMGCs) (Franco et al. 2022). These genetic adaptations may have played a crucial role in the ecological versatility of Biscogniauxia, allowing species within the genus to act as both saprotrophs and opportunistic pathogens, depending on environmental conditions. Such evolutionary mechanisms highlight the significance of Biscogniauxia in forests, where it participates in both wood decay processes and plant-pathogen interactions (Ju et al. 1998; Henriques et al. 2014; 2016). This complexity is reflected in the ecological versatility of the genus, with species capable of shifting between saprotrophic and opportunistic pathogenic lifestyles depending on environmental conditions. Such adaptive strategies are particularly relevant in the context of fungal systematics, as they offer insight into the ecological roles and evolutionary history of Biscogniauxia. Concurrently, we are expanding sampling across Thailand and Southeast Asia to build a population-level framework for precise species delimitation, combining genotyping and multi-locus phylogeny. These data will be integrated with state-of-the-art metabolomics to delineate chemotaxonomic traits and map them onto the evolutionary history of the Xylariales. We also plan controlled host–interaction assays to examine the endophyte-to-pathogen transition and identify candidate virulence factors. Finally, we will establish an open reference library for Thai Biscogniauxia, including vouchers, living cultures, and curated metabolite profiles, to enable reproducible identification and support responsible bioprospecting by the broader mycological community.
Supplementary Information
Below is the link to the electronic supplementary material.ESM 1(PDF 2.27 MB)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bakhshi-Ganje M, Mahmoodi S, Ahmadi K, Mirabolfathy M (2024) Potential distribution of Biscogniauxia mediterranea and Obolarina persica, causal agents of oak charcoal disease in Iran’s Zagros forests. Sci Rep 14:7784. 10.1038/s 41598-024-57298-210.1038/s 41598-024-57298-2PMC 1098758238565553 · doi ↗ · pubmed ↗
- 2Charria-Girón E, Marin-Felix Y, Beutling U, Franke R, Brönstrup M, Vasco-Palacios AM, Caicedo NH, Surup F (2023) Metabolomics insights into the polyketide-lactones produced by Diaporthe caliensis sp. nov. an endophyte of the medicinal plant Otoba gracilipes. Microbiol Spectr 11(6):e 0274323. 10.1128/spectrum.02743-2310.1128/spectrum.02743-23PMC 1071520937921483 · doi ↗ · pubmed ↗
- 3Ju YM, Rogers JD (1996) A revision of the genus Hypoxylon. Mycologia Memoir No.° 20. American Phytopathological Society (APS Press)
- 4Ju YM, Rogers JD, San Martín F, Granmo A (1998) The genus Biscognauxia. Mycotaxon 66:1–98
- 5Raimondo ML, Lops F, Carlucci A (2016) Charcoal canker of pear, plum, and quince trees caused by Biscogniauxia rosacearum sp nov in southern Italy. Plant Dis 100:1813–1822. 10.1094/PDIS-09-15-1037-RE 10.1094/PDIS-09-15-1037-RE 30682981 · doi ↗ · pubmed ↗
- 6Robert V, Stegehuis G, Stalpers J (2005) The Myco Bank engine and related databases. https://www.mycobank.org