Phytochemical composition and synergistic antimicrobial effects of Rosmarinus officinalis essential oils during flowering in an arid mediterranean region
Imane Abbad, Bouchra Soulaimani, Imane El Hakioui, Soraia El Baz, Elena Maria Varoni, Marcello Iriti

TL;DR
This study examines how the timing of harvesting rosemary affects the quality and antimicrobial power of its essential oils in a dry Mediterranean climate.
Contribution
First systematic month-by-month evaluation of rosemary essential oils during flowering in arid Mediterranean conditions.
Findings
EO yields peaked in autumn, with 1,8-cineole, α-pinene, and camphor as dominant compounds.
Autumn-harvested EOs showed strong antimicrobial activity against fungi, especially C. krusei and C. albicans.
EOs significantly enhanced streptomycin efficacy against E. coli, up to 32-fold in autumn.
Abstract
Rosmarinus officinalis has attracted significant attention due to its broad-spectrum antimicrobial activity, largely attributed to its bioactive essential oils (EOs). Several studies indicate that the flowering period is crucial for harvesting rosemary's aerial parts for optimal EO extraction. However, its prolonged flowering period complicates the determination of an optimal harvest time, potentially affecting yield, chemical composition, and efficacy. This study provides, for the first time, a systematic month-by-month evaluation of EO yield, chemical composition, and synergistic antimicrobial potential of rosemary cultivated under arid Mediterranean conditions during its flowering period (September to March). EO samples were analyzed by GC-MS and assessed for antimicrobial activity against clinically relevant pathogens, including S. aureus, E. coli, S. enterica, and four Candida…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
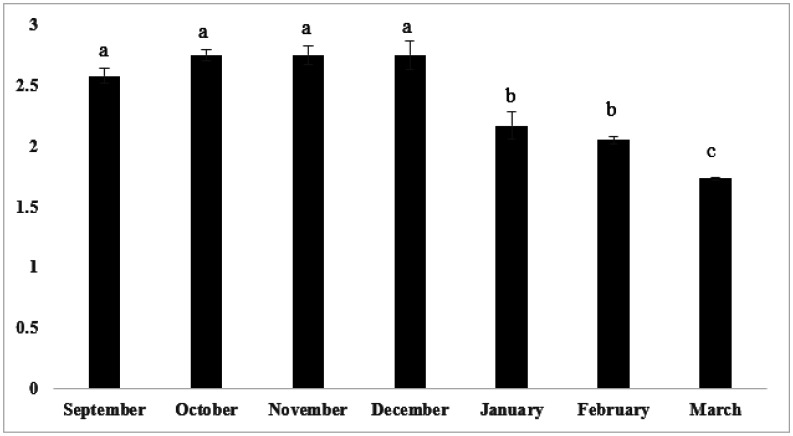 Figure 1
Figure 1| Evaporation (mm) | Rainfalls (mm) | Temperature (°C) | Humidity (%) | |
| SEP 2021 | 41.7 | 0.2 | 25.19 | 52.38 |
| OCT 2021 | 31.8 | 0 | 22.73 | 46.25 |
| NOV 2021 | 20.9 | 22 | 24.72 | 49.81 |
| DEC 2021 | 17.4 | 32 | 14.28 | 60.44 |
| JAN 2022 | 17.4 | 10.8 | 13.06 | 51.19 |
| FEB 2022 | 19 | 35 | 16.07 | 52.06 |
| MAR 2022 | 16.7 | 61 | 14.01 | 67 |
| RTb | RIc | RIlit | Compoundsa | SEP | OCT | NOV | DEC | JAN | FEB | MAR |
| 2.63 | 924 | 924 | α-Thujene | 0.22 ± 0.15 | 0.39 ± 0.00 | 0.48 ± 0.00 | 0.39 ± 0.01 | |||
| 2.71 | 936 | 932 | α-Pinene | 19.92 ± 0.96 | 18.96 ± 0.35 | 16.03 ± 0.18 | 15.85 ± 0.23 | 20.44 ± 0.08 | 15.36 ± 0.23 | 28.97 ± 0.10 |
| 2.85 | 945 | 946 | Camphene | 5.60 ± 0.12 | 4.81 ± 0.11 | 4.58 ± 0.09 | 5.05 ± 0.09 | 6.00 ± 0.07 | 5.60 ± 0.09 | 5.91 ± 0.05 |
| 2.96 | 977 | 974 | 1-Octen-3-ol | 0.15 ± 0.05 | 0.14 ± 0.03 | 0.12 ± 0.00 | ||||
| 3.08 | 978 | 974 | β-Pinene | 6.74 ± 0.00 | 6.21 ± 0.00 | 6.29 ± 0.05 | 10.64 ± 0.08 | 9.29 ± 0.02 | 10.24 ± 0.09 | 8.49 ± 0.04 |
| 3.30 | 998 | 1002 | α-Phellandrene | 0.24 ± 0.00 | 0.29 ± 0.00 | 0.24 ± 0.00 | 0.21 ± 0.00 | 0.25 ± 0.00 | 0.19 ± 0.00 | 0.27 ± 0.00 |
| 3.42 | 1012 | 1014 | α-Terpinene | 0.70 ± 0.01 | 0.78 ± 0.02 | 0.60 ± 0.01 | 0.60 ± 0.02 | 0.72 ± 0.03 | 0.56 ± 0.02 | 0.73 ± 0.03 |
| 3.50 | 1025 | 1020 | p-Cymene | 0.40 ± 0.54 | 1.55 ± 0.02 | 1.26 ± 0.01 | 1.93 ± 0.01 | |||
| 3.61 | 1034 | 1026 | 1,8-Cineole | 34.06 ± 0.50 | 33.96 ± 0.25 | 36.28 ± 0.26 | 34.94 ± 0.00 | 27.57 ± 0.76 | 35.30 ± 0.12 | 29.57 ± 0.09 |
| 3.90 | 1048 | 1054 | γ-Terpinene | 1.17 ± 0.04 | 1.18 ± 0.03 | 0.96 ± 0.02 | 1.15 ± 0.02 | 1.40 ± 0.02 | 1.13 ± 0.02 | 1.23 ± 0.01 |
| 4.03 | 1056 | 1065 | Cis-Sabinene hydrate | 0.11 ± 0.00 | 0.09 ± 0.00 | 0.09 ± 0.00 | 0.20 ± 0.00 | 0.09 ± 0.00 | 0.13 ± 0.00 | 0.05 ± 0.01 |
| 4.32 | 1082 | 1086 | Terpinolene | 0.55 ± 0.02 | 0.55 ± 0.00 | 0.44 ± 0.00 | 0.41 ± 0.00 | 0.63 ± 0.02 | 0.44 ± 0.00 | 0.64 ± 0.01 |
| 4.38 | 1098 | 1095 | Linalool | 0.49 ± 0.03 | 0.50 ± 0.03 | 0.43 ± 0.03 | 0.34 ± 0.02 | 0.44 ± 0.01 | 0.33 ± 0.01 | 0.65 ± 0.00 |
| 4.48 | 1105 | 1114 | Fenchol | 0.04 ± 0.00 | 0.06 ± 0.00 | 0.09 ± 0.00 | 0.05 ± 0.00 | 0.06 ± 0.00 | 0.06 ± 0.00 | 0.07 ± 0.00 |
| 4.75 | 1114 | 1124 | Chrysanthenone | 0.15 ± 0.00 | 0.15 ± 0.00 | 0.08 ± 0.00 | 0.09 ± 0.00 | 0.23 ± 0.01 | 0.11 ± 0.00 | 0.23 ± 0.00 |
| 5.30 | 1131 | 1141 | Camphor | 14.21 ± 0.05 | 14.51 ± 0.00 | 15.37 ± 0.12 | 13.48 ± 0.16 | 10.21 ± 0.06 | 14.19 ± 0.16 | 7.12 ± 0.00 |
| 5.65 | 1167 | 1165 | Borneol | 4.75 ± 0.15 | 4.43 ± 0.00 | 4.57 ± 0.01 | 4.63 ± 0.05 | 4.78 ± 0.00 | 5.68 ± 0.04 | 3.26 ± 0.05 |
| 5.84 | 1176 | 1174 | Terpinen-4-ol | 0.68 ± 0.02 | 0.80 ± 0.03 | 0.77 ± 0.04 | 0.69 ± 0.04 | 0.62 ± 0.02 | 0.73 ± 0.04 | 0.55 ± 0.02 |
| 6.08 | 1187 | 1186 | α-Terpineol | 2.12 ± 0.06 | 2.73 ± 0.09 | 2.61 ± 0.10 | 2.25 ± 0.13 | 1.91 ± 0.07 | 2.09 ± 0.08 | 1.52 ± 0.05 |
| 6.51 | 1204 | 1204 | l-Verbenone | 0.35 ± 0.00 | 0.39 ± 0.01 | 0.11 ± 0.00 | 0.07 ± 0.00 | 0.20 ± 0.00 | 0.05 ± 0.00 | 0.55 ± 0.01 |
| 8.11 | 1287 | 1284 | Bornyl acetate | 1.62 ± 0.01 | 1.52 ± 0.02 | 0.83 ± 0.01 | 1.07 ± 0.01 | 1.91 ± 0.01 | 1.28 ± 0.00 | 1.06 ± 0.00 |
| 8.44 | 1297 | 1298 | Carvacrol | 0.02 ± 0.01 | 0.05 ± 0.02 | 0.09 ± 0.01 | ||||
| 10.41 | 1376 | 1374 | α-Copaene | 0.03 ± 0.00 | 0.06 ± 0.00 | 0.01 ± 0.00 | 0.14 ± 0.00 | |||
| 11.60 | 1417 | 1417 | Caryophyllene | 4.36 ± 0.14 | 5.42 ± 0.07 | 6.31 ± 0.10 | 6.18 ± 0.01 | 8.45 ± 0.05 | 3.37 ± 0.00 | 4.28 ± 0.00 |
| 12.49 | 1453 | 1452 | Humulene | 0.60 ± 0.01 | 0.89 ± 0.00 | 0.76 ± 0.00 | 0.84 ± 0.01 | 1.22 ± 0.00 | 0.42 ± 0.00 | 0.94 ± 0.00 |
| 13.06 | 1477 | 1478 | γ-Muurolene | 0.03 ± 0.00 | 0.01 ± 0.00 | 0.05 ± 0.00 | 0.12 ± 0.00 | |||
| 13.63 | 1500 | 1500 | Bicylogermacrene | 0.11 ± 0.00 | ||||||
| 13.97 | 1508 | 1505 | β-Bisabolene | 0.06 ± 0.01 | 0.09 ± 0.00 | 0.06 ± 0.00 | 0.02 ± 0.02 | 0.08 ± 0.00 | 0.02 ± 0.00 | 0.03 ± 0.00 |
| 14.10 | 1515 | 1513 | γ-Cadinene | 0.04 ± 0.00 | 0.04 ± 0.00 | 0.02 ± 0.00 | 0.05 ± 0.00 | 0.02 ± 0.00 | 0.11 ± 0.02 | |
| 14.33 | 1524 | 1522 | δ-Cadinene | 0.07 ± 0.03 | 0.06 ± 0.00 | 0.01 ± 0.00 | 0.12 ± 0.00 | 0.01 ± 0.01 | 0.25 ± 0.00 | |
| 16.00 | 1561 | 1582 | Caryophyllene oxide | 0.09 ± 0.01 | 0.13 ± 0.01 | 0.16 ± 0.00 | 0.20 ± 0.00 | 0.46 ± 0.00 | 0.27 ± 0.00 | 0.27 ± 0.00 |
| Oxygen-containing monoterpenes | 57.00 ± 0.52 | 57.68 ± 0.08 | 60.41 ± 0.53 | 56.8 ± 0.42 | 46.10 ± 0.93 | 58.68 ± 0.46 | 43.56 ± 0.24 | |||
| Monoterpene hydrocarbons | 35.15 ± 0.43 | 33.18 ± 0.03 | 30.68 ± 0.39 | 34.29 ± 0.49 | 39.19 ± 0.20 | 35.17 ± 0.49 | 48.17 ± 0.24 | |||
| Oxygen-containing sesquiterpenes | 0.09 ± 0.01 | 0.13 ± 0.01 | 0.16 ± 0.00 | 0.20 ± 0.00 | 0.46 ± 0.00 | 0.27 ± 0.00 | 0.27 ± 0.00 | |||
| Sesquiterpene hydrocarbons | 5.15 ± 0.19 | 6.53 ± 0.07 | 7.13 ± 0.10 | 7.07 ± 0.04 | 10.03 ± 0.06 | 3.95 ± 0.00 | 5.88 ± 0.02 | |||
| Other | 1.76 ± 0.06 | 1.66 ± 0.01 | 0.95 ± 0.01 | 1.07 ± 0.01 | 1.91 ± 0.01 | 1.28 ± 0.00 | 1.06 ± 0.00 | |||
| Total | 99.16 ± 0.17 | 99.18 ± 0.02 | 99.33 ± 0.03 | 99.43 ± 0.03 | 97.69 ± 0.69 | 99.36 ± 0.02 | 98.94 ± 0.02 |
| Essential oils | Antibiotics | ||||||||
| Microorganisms | SEP | OCT | NOV | DEC | JAN | FEV | MAR | Amphotericin B | Streptomycin |
| 9.88 ± 0.06a | 10.91 ± 1.51 | 10.41 ± 1.69 | 8.74 ± 0.34 | 9.2 ± 0.52 | 6.52 ± 0.40 | 8.41 ± 0.84 | 11.5 ± 0.93 | ||
| C. | 13.31 ± 1.58 | 15.68 ± 1.24 | 16.49 ± 1.78 | 11.25 ± 1.34 | 10.72 ± 1.33 | 8.06 ± 0.32 | 13.25 ± 0.68 | 13.4 ± 0.62 | |
| 11.47 ± 0.34 | 15.72 ± 0.73 | 14.3 ± 0.04 | 10.38 ± 0.2 | 8.94 ± 0.9 | 7.5 ± 0.55 | 10.4 ± 0.52 | 8.98 ± 1.15 | ||
| 13.8 ± 1.22 | 15.07 ± 2.52 | 15.83 ± 0.39 | 12.2 ± 0.73 | 9.44 ± 1.26 | 7.73 ± 1.19 | 9.16 ± 0.41 | 10.56 ± 0.4 | ||
|
| 9.3 ± 0.05 | 9.6 ± 0.05 | 10.6 ± 0.05 | 9 ± 0.1 | 8.6 ± 0.2 | 9.6 ± 0.37 | 9 ± 0.1 | 14.8 ± 0.34 | |
|
| 10 ± 0.35 | 11.3 ± 0.35 | 11.3 ± 0.05 | 10 ± 0.17 | 9.3 ± 0.05 | 8.6 ± 0.2 | 9.3 ± 0.05 | 17.7 ± 1.6 | |
|
| 8.1 ± 0.25 | 10.1 ± 0.05 | 9.3 ± 0.12 | 9.6 ± 0.03 | 8.2 ± 0.2 | 8.1 ± 0.1 | 7.1 ± 0.21 | 13.08 ± 0.01 | |
| Microorganisms tested | ||||||||||||||
|
|
|
| ||||||||||||
| EOs/antibiotics | MIC | MMC | MIC | MMC | MIC | MMC | MIC | MMC | MIC | MMC | MIC | MMC | MIC | MMC |
| SEP | 5 | 5 | 2.5 | 2.5 | 10 | 10 | 5 | 5 | 11.12 | 11.12 | 5.56 | 11.12 | 22.25 | 22.25 |
| OCT | 2.5 | 2.5 | 2.5 | 2.5 | 5 | 5 | 2.5 | 2.5 | 11.12 | 11.12 | 2.78 | 5.56 | 11.12 | 11.12 |
| NOV | 2.5 | 2.5 | 2.5 | 2.5 | 5 | 5 | 2.5 | 2.5 | 11.12 | 11.12 | 2.78 | 2.78 | 11.12 | 11.12 |
| DEC | 5 | 5 | 2.5 | 2.5 | 5 | 5 | 2.5 | 2.5 | 22.25 | 22.25 | 2.78 | 2.78 | 22.25 | 22.25 |
| JAN | 10 | 10 | 5 | 5 | 10 | 10 | 5 | 5 | 22.25 | 22.25 | 5.56 | 22.25 | 22.25 | 22.25 |
| FEB | 10 | 10 | 5 | 5 | 10 | 10 | 20 | 20 | 22.25 | 22.25 | 5.56 | 11.2 | 22.25 | 22.25 |
| MAR | 5 | 5 | 5 | 5 | 10 | 10 | 5 | 5 | 22.25 | 22.25 | 22.25 | 22.25 | 44.5 | 44.5 |
| Amphotericin B | 1.953 | 1.953 | 0.976 | 0.976 | 1.953 | 1.953 | 0.976 | 0.976 | ||||||
| Streptomycin | 0.072 | 0.072 | 0.009 | 0.018 | 0.072 | 0.072 | ||||||||
| Bacterial strains | MICc | Gain | FIC | FICI | Interaction | |||
| EO | ATB | EO | ATB | |||||
|
| SEP | 2.78 | 0.00225 | 32 | 0.25 | 0.03 | 0.28 | Synergism |
| OCT | 2.78 | 0.00225 | 32 | 0.25 | 0.03 | 0.28 | Synergism | |
| NOV | 2.78 | 0.00225 | 32 | 0.25 | 0.03 | 0.28 | Synergism | |
| DEC | 5.56 | 0.0045 | 16 | 0.25 | 0.06 | 0.31 | Synergism | |
| JAN | 5.56 | 0.0045 | 16 | 0.25 | 0.06 | 0.31 | Synergism | |
| FEB | 5.56 | 0.0045 | 16 | 0.25 | 0.06 | 0.31 | Synergism | |
| MAR | 5.56 | 0.0045 | 16 | 0.25 | 0.06 | 0.31 | Synergism | |
|
| SEP | 1.39 | 0.0011 | 8 | 0.25 | 0.12 | 0.37 | Synergism |
| OCT | 0.695 | 0.0011 | 8 | 0.25 | 0.12 | 0.37 | Synergism | |
| NOV | 0.695 | 0.0011 | 8 | 0.25 | 0.12 | 0.37 | Synergism | |
| DEC | 0.695 | 0.0011 | 8 | 0.25 | 0.12 | 0.37 | Synergism | |
| JAN | 1.39 | 0.00225 | 4 | 0.25 | 0.12 | 0.37 | Synergism | |
| FEB | 1.39 | 0.00225 | 4 | 0.25 | 0.25 | 0.50 | Synergism | |
| MAR | 5.56 | 0.00056 | 16 | 0.25 | 0.06 | 0.31 | Synergism | |
|
| SEP | 5.56 | 0.018 | 4 | 0.25 | 0.25 | 0.50 | Synergism |
| OCT | 2.78 | 0.009 | 8 | 0.25 | 0.12 | 0.37 | Synergism | |
| NOV | 2.78 | 0.009 | 8 | 0.25 | 0.12 | 0.37 | Synergism | |
| DEC | 5.56 | 0.018 | 4 | 0.25 | 0.25 | 0.50 | Synergism | |
| JAN | 5.56 | 0.009 | 8 | 0.25 | 0.12 | 0.37 | Synergism | |
| FEB | 5.56 | 0.018 | 4 | 0.25 | 0.25 | 0.50 | Synergism | |
| MAR | 11.12 | 0.009 | 8 | 0.25 | 0.12 | 0.37 | Synergism |
| Candida | MIC EO | MIC ATB | Gain | FIC EO | FIC ATB | FICI | Interaction | |
|
| SEP | 1.25 | 0.488 | 4 | 0.25 | 0.25 | 0.50 | Synergism |
| OCT | 0.625 | 0.976 | 2 | 0.25 | 0.50 | 0.75 | Additive | |
| NOV | 0.625 | 0.488 | 4 | 0.25 | 0.50 | 0.75 | Additive | |
| DEC | 1.25 | 0.976 | 2 | 0.25 | 0.50 | 0.75 | Additive | |
| JAN | 2.5 | 0.976 | 2 | 0.25 | 0.50 | 0.75 | Additive | |
| FEB | 2.5 | 0.976 | 2 | 0.25 | 0.50 | 0.75 | Additive | |
| MAR | 1.25 | 0.976 | 2 | 0.25 | 0.50 | 0.75 | Additive | |
|
| SEP | 0.625 | 0.488 | 2 | 0.25 | 0.50 | 0.75 | Additive |
| OCT | 0.625 | 0.488 | 2 | 0.25 | 0.50 | 0.75 | Additive | |
| NOV | 0.625 | 0.488 | 2 | 0.25 | 0.50 | 0.75 | Additive | |
| DEC | 0.625 | 0.488 | 2 | 0.25 | 0.50 | 0.75 | Additive | |
| JAN | 1.25 | 0.488 | 2 | 0.25 | 0.50 | 0.75 | Additive | |
| FEB | 1.25 | 0.488 | 2 | 0.25 | 0.50 | 0.75 | Additive | |
| MAR | 1.25 | 0.488 | 2 | 0.25 | 0.50 | 0.75 | Additive | |
|
| SEP | 2.5 | 1.953 | 1 | 0.25 | 1 | 1.25 | Indifference |
| OCT | 1.25 | 1.953 | 1 | 0.25 | 1 | 1.25 | Indifference | |
| NOV | 1.25 | 1.953 | 1 | 0.25 | 1 | 1.25 | Indifference | |
| DEC | 1.25 | 1.953 | 1 | 0.25 | 1 | 1.25 | Indifference | |
| JAN | 2.5 | 1.953 | 1 | 0.25 | 1 | 1.25 | Indifference | |
| FEB | 2.5 | 1.953 | 1 | 0.25 | 1 | 1.25 | Indifference | |
| MAR | 2.5 | 1.953 | 1 | 0.25 | 1 | 1.25 | Indifference | |
|
| SEP | 2.5 | 0.976 | 2 | 0.25 | 0.50 | 0.75 | Additive |
| OCT | 0.625 | 0.488 | 2 | 0.25 | 0.50 | 0.75 | Additive | |
| NOV | 0.625 | 0.976 | 1 | 0.25 | 1 | 1.25 | Indifference | |
| DEC | 0.625 | 0.976 | 1 | 0.25 | 1 | 1.25 | Indifference | |
| JAN | 2.5 | 0.976 | 1 | 0.25 | 1 | 1.25 | Indifference | |
| FEB | 5 | 0.976 | 1 | 0.25 | 1 | 1.25 | Indifference | |
| MAR | 2.5 | 0.976 | 1 | 0.25 | 1 | 1.25 | Indifference | |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEssential Oils and Antimicrobial Activity · Plant biochemistry and biosynthesis · Flowering Plant Growth and Cultivation
Introduction
In recent years, the use of herbal medicine has seen significant growth in combating pathogenic bacteria, driven by the increasing emergence of bacterial resistance to existing antibiotics. This rise in multidrug-resistant infections has been linked to an alarming escalation in mortality rates annually. It is projected that if no measures are taken, antimicrobial resistance, currently causing around 700,000 deaths globally each year, could result in as many as 10 million deaths annually by 2050 [1],[2]. This urgent challenge has intensified the search for innovative treatments capable of combating multi-drug-resistant bacteria. Plant-derived essential oils (EOs) have emerged as a promising alternative due to their potent antimicrobial properties against resistant strains, their availability, and favorable perceptions among consumers [3]–[5]. Research has shown that various EOs from plants exhibit strong antimicrobial effects on both Gram-positive and Gram-negative bacteria, which is largely attributed to their complex chemical structures and the synergistic actions of their components [5]–[7].
Rosmarinus officinalis (rosemary), belonging to the Lamiaceae family, is one of the medicinal and aromatic plants that has attracted significant research interest. In fact, the species is known to be a rich source of EOs, which find extensive use across agri-food and pharmaceutical industries [8]. Its EOs are highly valued for their diverse biological and pharmacological properties, including antioxidant, antibacterial, and antifungal activities [9]. These biological properties have been attributed mainly to their richness in several bioactive molecules, like 1,8-cineole, borneol, β- and α-pinene, limonene, camphene, camphor, and myrcene [10]–[12]. Importantly, beyond their intrinsic antimicrobial activity, rosemary EOs have also been shown to enhance the efficacy of conventional antibiotics through synergistic interactions. Several studies reported that rosemary EO can potentiate the activity of many antimicrobial agents, including some aminoglycosides, tetracyclines, amphenicols, fluoroquinolone, and cephalosporins against Gram-negative pathogens, mainly through disruption of bacterial membranes and interference with efflux pump activity [13],[14]. However, like many medicinal and aromatic plants, the yield and chemical composition of rosemary EOs are strongly affected by various factors. These include the phenological stages of the plant, season of harvesting, bioclimatic conditions, geographical origin, organs, plant age among others [15]–[18]. Each of these elements can significantly influence the concentrations and types of bioactive compounds present in the oil, thus impacting its overall properties and efficacy. To optimize oil yields, specific chemical compositions, and enhance the biological activity of rosemary EO, research efforts have been focused on determining the ideal harvesting season under defined climatic conditions. Several studies indicate that the flowering period is crucial for harvesting rosemary's aerial parts for optimal EO extraction [19]–[23]. During this stage, rosemary significantly increases its production of secondary metabolites, a reaction possibly linked to enhanced pollinator activity [18]. This accumulation maximizes the richness of EOs in bioactive compounds, enhancing their therapeutic potential. In addition, cultivation under controlled aridity has been shown to increase the concentration and quality of rosemary EOs, making it a potentially interesting crop for marginal arid lands, particularly in Mediterranean regions [24]. However, it is well recognized that rosemary exhibits a prolonged flowering period, which complicates the determination of the optimal harvesting time for its EO, corresponding to the highest concentration of antimicrobial active compounds [25]. Given this extended flowering period of rosemary, this study aimed to (i) assess the monthly variation in EO yield and chemical profile throughout the flowering phase, (ii) evaluate monthly antimicrobial activity and synergistic interaction of rosemary EO with two conventional antibiotics (streptomycin and amphotericin B), and (iii) identify key compositional constituents influencing the observed bioactivity.
Materials and methods
Crop experimental design
2.1.
This research was conducted on an experimental parcel situated in Oulad Dlim, Marrakech-Morocco (32°01′23″N/8°13′36″W, 388 m above sea level). The site is in a region with an arid bioclimate, with a mean maximum temperature of 38.3 °C in July and a mean minimum temperature of 4.5 °C in February, along with an average annual rainfall of 242 mm. The soil of experimental plot was a red clay loam type having a pH of 8.04, electrical conductivity of 0.49 mS/cm, organic matter content of 4.1%, total nitrogen at 0.66%, P_2_O_5_ at 18.20 mg/100 g, K_2_O at 49.9 mg/100 g, and CaCO_3_ at 34.1%. In March of 2018, during the vegetative development stage, 600 stem cuttings of rosemary were randomly selected from a natural population located in the Er-Rich region, Central of Morocco (32°08′25″N, 5°16′48″W, 1460 m above sea level). Clonal propagation was used to preserve the genetic identity of the natural population (chemotype). The cuttings were immediately transplanted into the Polyethylene bags (15/10) containing sand (1/3) and peat (2/3). After 4 weeks, rooted cuttings (15 to 20 cm high) were transplanted randomly to the experimental area with a planting distance of 0.5 m × 0.5 m. For the first month (establishment phase), a watering level equivalent to 60% of soil field capacity was provided early in the morning twice a week. For the next months, the young shoots were drip-irrigated weekly during the non-rainy season (dry season), with water applied at a rate of 5 liters per hour for three hours per session. No phytosanitary treatments were applied. In the period from September 2020 to March 2021, which corresponds to the flowering season, 20 flowering branches were randomly collected each month from a rosemary cultivated population. Monthly rainfalls and average monthly temperature, humidity and evaporation during the period of sample collection are reported in Table 1. Voucher specimens of cultivated (ROC-14) and wild (ROW-34) rosemary are deposited at the Laboratory of Microbial Biotechnology, Agrosciences and Environment, Faculty of Sciences, Cadi Ayyad University. Before EO extraction, the sampled plant materials (flowering branches) were air-dried in the shade at a temperature of 25 °C until reaching a constant weigh.
EO extraction and gas chromatography/mass spectrometry (GC/MS) analyses
2.2.
EOs were extracted using a hydrodistillation process with a Clevenger-type apparatus (Borosil 3451029 Clevenger Apparatus) over a duration of 3 hours to ensure complete recovery. The process was conducted in triplicate (200 g × 3). The yields were determined by calculating the ratio of the volume of oil obtained to the weight of the dried plants used (v/w). The extracted EOs were then separated, dried using anhydrous sodium sulfate, and stored in opaque flasks at 4 °C in a dark environment until they were analyzed.
The qualitative and quantitative identification of the EO constituents was conducted using gas chromatography coupled to mass spectrometry (GC-MS) as previously described by Soulaimani et al. [26]. To accurately determine the compounds in each EO chromatographic profile during a single analysis, mass spectra were compared to known reference compounds when available, and consulted against the NBS75K and WILEY275 libraries, as well as a documented terpene library (Adams, 2007). Retention indices (RIs) were calculated based on the retention times of a range of C7–C30 n-alkanes using linear interpolation. These indices were then compared with those of known compounds or corroborated against published data to ensure accuracy.
Assessment of antimicrobial activity
2.3.
The antimicrobial efficacy of the EOs was evaluated against four clinically isolated pathogenic Candida strains: C. albicans (CCMM L4), C. glabrata (CCMM L7), C. krusei (CCMM L10), and C. parapsilosis (CCMM L18). Additionally, three well-known pathogenic bacteria were tested: Staphylococcus aureus (CCMM B3), Escherichia coli (ATCC 8739), and Salmonella enterica (ATCC 14028). The antimicrobial activity of EOs was assessed using the agar disc diffusion and microwell dilution methods, in accordance with the Clinical and Laboratory Standards Institute (CLSI) guidelines [27]. For the disc diffusion assay, sterile discs (6 mm in diameter) impregnated with 10 µL of EOs were placed on the surface of either Sabouraud Dextrose Agar (SDA) or Mueller Hinton Agar (MHA) plates that had been previously inoculated with 0.1 mL of yeast or bacterial suspensions at concentrations of 10^5^ and 10^8^ CFU/mL, respectively. The plates were stored at 4 °C for four hours to facilitate EO diffusion, then incubated at 37 °C for 24 hours for bacterial cultures and at 28 °C for 48 hours for yeast cultures. Antimicrobial activity was determined by measuring the inhibition zones (IZ) around the discs. Streptomycin (15 µg/disc) and amphotericin B (5 µg/disc) served as positive controls, and all tests were conducted in triplicate.
In the microwell dilution method, EOs were diluted in a 2-fold series in 4% dimethyl sulfoxide (DMSO), with 100 µL of each dilution added to microwells that contained 100 µL of yeast or bacterial suspensions at densities of 1–2 × 10^3^ and 10^8^ CFU/mL, respectively. The microplates were incubated at 28 °C for 18–24 hours for Candida strains and at 37 °C for bacteria. The minimum inhibitory concentration (MIC) was defined as the lowest concentration of EOs that prevented the visible growth of the tested strains. To determine the minimum microbicidal concentration (MMC), 0.1 mL from each clear well, indicating no growth in MIC tests, was sub-cultured on appropriate agar (MHA or SDA) and incubated under the same conditions previously described. The MMC was identified as the lowest EO concentration that effectively killed 99,9% of the bacteria or yeasts. Streptomycin and amphotericin B were employed as standard antibacterial and antifungal agents, respectively.
Synergistic effect of EOs with standard antimicrobials
2.4.
The interaction between rosemary EOs and two standard antimicrobial agents, amphotericin B for Candida spp. and streptomycin for bacteria, were evaluated using a checkerboard assay based on the broth microdilution method. Briefly, mixtures were prepared by combining 50 µL of the EO dilution with 50 µL of the antimicrobial agent dilution. Each well was then inoculated with 100 µL of microbial suspension. The microplates were incubated at 37 °C for 24 hours for bacterial strains and at 28 °C for 48 hours for Candida spp. The antimicrobial efficacy of each combination was assessed by calculating the MIC gain, which reflects the fold reduction in the MIC of the antimicrobial when used in combination. It was calculated using the formula: MIC gain = MIC of the antimicrobial alone/MIC of the antimicrobial in combination. To determine the nature of the interaction, the fractional inhibitory concentration index (FICI) was also calculated as follows: FICI = FIC (EO) + FIC (antimicrobial), where FIC (EO) = MIC of EO in combination/MIC of EO alone, and FIC (antimicrobial) = MIC of antimicrobial in combination/MIC of antimicrobial alone. All tests were conducted in triplicate. According to EUCAST standards, interactions were classified as synergistic when FICI ≤ 0.5, additive when 0.5 < FICI ≤ 1, indifferent when 1 < FICI ≤ 2, and antagonistic when FICI > 2 [28].
Statistical analysis
2.5.
Data were expressed as the mean ± standard deviation, group means were compared by one way ANOVA and Tukey test to identify significance (p < 0.05) among groups using SPSS 21.0 statistical software.
Results and discussion
Yields and chemical compositions of rosemary EOs
3.1.
The yields of EOs from rosemary, quantified as the volume of oil per weight of dried plant material, ranged between 1.73% and 2.75%. These values are comparatively higher than those reported for some rosemary populations cultivated under Mediterranean climate [25],[29]. Significant variations in EO yields were noted, with the highest yields occurring during the months of October, November, and December. In contrast, March, coinciding with the end of the flowering season, consistently showed the lowest EO productions (Figure 1). This trend is consistent with previous findings in both wild and cultivated rosemary, where higher EO yields were recorded during the autumn months, likely associated with climatic water stress conditions that favor the accumulation of secondary metabolites [19],[23],[25],[29],[30]. In fact, at our experimental site, this flowering season is characterized by reduced rainfall, elevated temperatures, and increased evapotranspiration rates, as shown in Table 1. The chemical composition of rosemary EOs collected throughout the flowering season (September to May) is presented in Table 2. The constituents, listed in order of elution, were identified using GC/MS analysis combined with retention indices. A total of 31 compounds were identified, representing between 97.69 ± 0.69% and 99.43 ± 0.03% of the total oil content. Monoterpenes constituted the predominant class across all EO samples. Sesquiterpenes, both hydrocarbon and oxygenated forms, were consistently present in low concentrations, confirming their scarcity in Moroccan Rosemary EOs, as reported in numerous previous studies [31],[32]. Regarding the main constituents of EOs, 1,8-cineole consistently exhibited the highest concentrations throughout the entire sampling period, ranging from 27.57 ± 0.76% to 35.30 ± 0.12% (Table 2). The other main compounds of EOs were α-pinene (15.36 ± 0.23%–28.97 ± 0.10%), camphor (7.12 ± 0.00%–15.37 ± 0.12%), β-pinene (6.21 ± 0.00%–10.24 ± 0.09%), borneol (3.26 ± 0.05%–5.68 ± 0.04%), and caryophyllene (4.36 ± 0.14–8.45 ± 0.05%). This result confirms the predominance of the 1,8-cineole/α-pinene/camphor chemotype in Moroccan rosemary, in line with findings from several previous studies [31]–[33]. The variation in the main EO components throughout the sampling period revealed that 1,8-cineole maintained the highest concentrations across all months, except in January, where it exhibited a marked decrease. Camphor was more prominent during autumn, followed by a gradual decline through winter and spring. In contrast, α-pinene exhibited significant monthly variation, reaching its highest concentration in March, which coincided with the end of the flowering period. This monthly variation is consistent with previous findings on rosemary EOs, which highlight the sensitivity of their main chemical constituents, particularly monoterpenes like 1,8-cineole, camphor, and α-pinene, to climatic factors [19],[31],[32],[34]. It is evident that these environmental influences can alter the biosynthesis and relative abundance of EO compounds, ultimately affecting the oil's biological activity.
Monthly variation of rosemary EO yields during the flowering period.
Antimicrobial assessment of the studied rosemary EOs
3.2.
This part of our study aimed to assess whether monthly variations during the flowering period influence the antimicrobial activity of rosemary EOs. The antimicrobial properties were tested against seven clinically relevant pathogens: Four Candida species (C. albicans, C. glabrata, C. parapsilosis, C. krusei), two Gram-negative bacteria (S. enterica and E. coli), and one Gram-positive strain (S. aureus). Antimicrobial efficacy was determined through the evaluation of IZ, MIC, and MMC, with detailed results presented in Tables 3 and 4.
All rosemary EO samples collected monthly exhibited antimicrobial activity, with inhibition zone diameters revealing a more pronounced effect against fungal strains compared to bacterial ones (Table 3). For bacterial strains, the EO showed moderate activity. The strongest inhibition was observed against S. aureus in November (11.3 ± 0.05 mm), while the activity against E. coli and S. enterica was generally lower, with inhibition zones mostly below 10 mm. The highest recorded zone for S. enterica was in October (10.1 ± 0.05 mm). Among the yeasts, C. krusei showed the highest sensitivity, with inhibition zones reaching up to 16.49 ± 1.78 mm in November. C. albicans also exhibited strong sensitivity in November (15.83 ± 0.39 mm), whereas C. glabrata showed relatively lower inhibition values, with a minimum of 6.52 ± 0.40 mm in February. Regarding C. parapsilosis, the activity was highest in October and November (15.72 ± 0.73 mm and 14.3 ± 0.04 mm, respectively) and decreased during the later months. This higher susceptibility of fungi may be attributed to the greater permeability of fungal membranes to lipophilic constituents found in rosemary EOs, such as 1,8-cineole, camphor, and α-pinene, which are known to disrupt membrane integrity and impair cellular respiration [19],[36],[37]. The MIC values (Table 4) further supported the inhibitory potential of rosemary EOs, showing significant variation depending on the month of harvesting. The strongest antifungal activity was observed in oils collected in October and November, with MICs as low as 2.5 mg/mL across all Candida species. In contrast, weaker antifungal effects were recorded in January and February, with MICs rising to 10 or even 20 mg/mL, particularly against C. albicans. Regarding bacterial strains, S. aureus exhibited greater sensitivity to rosemary EOs, notably those from October and November (MIC: 2.78 mg/mL), whereas the Gram-negative strains E. coli and S. enterica were less susceptible, with MICs ranging from 11.12 to 44.5 mg/mL. This lower sensitivity is consistent with the structural features of Gram-negative bacteria, whose outer membrane acts as a barrier to hydrophobic compounds [38].
A clear seasonal trend in the antimicrobial efficacy of rosemary EOs emerges from these results. The observed month-to-month variation in bioactivity can be largely attributed to seasonal shift in the composition of major EO constituents, which are strongly influenced by climatic conditions. Notably, in October and November, the EOs showed enhanced antimicrobial and antifungal activities, likely associated with elevated concentrations of 1,8-cineole, camphor, and α-pinene, monoterpenes with well-documented antimicrobial properties [36],[39],[40]. These months were characterized by relatively low rainfall and high evaporation rates (Table 1), indicative of moderate climatic water stress (Table 1). Such stress conditions are known to induce adaptive metabolic responses in aromatic plants, including the upregulation of monoterpene biosynthesis pathways, particularly those leading to the accumulation of oxygenated compounds like 1,8-cineole and camphor[30]. In contrast, the reduced concentrations of these bioactive monoterpenes in the winter months (cooler climatic conditions) may account for the weaker inhibitory effects, particularly against C. albicans and Gram-negative bacteria. Nevertheless, as highlighted by Zaouali et al. (2010), the antimicrobial activity of rosemary EO is not solely attributable to its major compounds, but rather to the synergistic interactions between both major and minor constituents [20].
Synergistic antimicrobial effects of rosemary EO with antibiotics
3.3.
A month-by-month analysis of the checkerboard assay results reveals clear seasonal trends in the synergistic activity of rosemary EOs when combined with streptomycin across all tested bacterial strains (Table 5). In E. coli, the strongest synergistic effect was observed during the early autumn months (September to November), with a consistent 32-fold reduction in the MIC of streptomycin and a FICI of 0.28, reflecting robust potentiation of antibiotic efficacy. This pronounced synergy likely correlates with the peak seasonal accumulation of oxygenated monoterpenes, in particular 1,8-cineole and camphor in the EOs, which are known to disrupt bacterial membranes and inhibit efflux pump activity [39],[40]. This membrane disruption likely enhances streptomycin penetration, facilitating its bactericidal effect and increasing bacterial mortality [41]. This mechanism is supported by Seukep et al. who demonstrated that plant-derived products can potentiate aminoglycoside activity against multidrug-resistant bacteria by interfering with efflux pump function [42]. From December to March, a notable decline in synergy was observed in E. coli, as the MIC gain dropped to 16-fold and the FICI slightly increased to 0.31, likely due to seasonal changes in the EO profile marked by a reduction in oxygenated monoterpenes. In March, despite an increase in α-pinene content (a hydrocarbon monoterpene), the synergistic effect remained unchanged (16-fold gain; FICI = 0.31), reinforcing the hypothesis that oxygenated monoterpenes play a more critical role in synergy than hydrocarbons alone [43]. In S. aureus, a Gram-positive strain, synergistic effects were also observed throughout all months, but with slightly lower MIC gains compared to E. coli. This difference may be attributed to structural differences in their cell envelopes, as the outer membrane of Gram-negative E. coli presents a stronger permeability barrier that is more markedly disrupted by EOs, thereby enhancing antibiotic uptake and synergy [44]. From September to December, the MIC of streptomycin was consistently reduced 8-fold, with FICI values of 0.37. Notably, the lowest EO MIC (0.695 mg/mL) was recorded in October and November, reflecting the increased potency of the EO during the autumn harvest. In January and February, the gain dropped to 4-fold, likely due to a moderate increase in ATB MIC and a potential decrease in EO membrane-disrupting compounds. Interestingly, a marked increase in EO MIC was observed in March (5.56 mg/mL), yet the MIC of streptomycin was strongly reduced (to 0.00056 mg/mL), yielding a 16-fold gain and a FICI of 0.31. This suggests that while EO efficacy may decline in terms of direct inhibition (higher MIC), it may still synergize effectively when paired with antibiotics, possibly through mechanisms beyond simple membrane disruption, such as efflux pump inhibition. S. enterica showed a more variable synergy profile. In October and November, the MIC gain peaked at 8-fold (ATB reduced from 0.018 to 0.009 mg/mL), with a FICI of 0.37, consistent with the autumnal peak in EO activity. During the colder months (December and February), the synergy weakened (4-fold MIC gain; FICI = 0.50), while a moderate rebound was observed in March, with MIC gains returning to 8-fold and a FICI of 0.37. These fluctuations likely reflect both compositional changes in EO and possible strain-specific sensitivity to seasonal chemical variations.
Regarding Candida spp., the synergistic effect of rosemary EOs with amphotericin B was overall limited and clearly strain and season dependent (Table 6). Among all tested isolates, only C. glabrata exhibited a true synergistic interaction in September (FICI = 0.50), accompanied by a 4-fold gain in the antibiotic's efficacy. This suggests that the EO profile during early autumn likely richer in oxygenated monoterpenes (1,8-cineole and camphor) may potentiate the antifungal activity of amphotericin B. In contrast, from October to March, the interaction in C. glabrata consistently dropped to an additive level (FICI = 0.75), with only a 2-fold gain in MIC, indicating a seasonal loss of bioactive synergy. Similarly, in C. albicans, a temporary additive effect was observed in September and October, followed by a shift to indifference (FICI = 1.25) in the subsequent months. This decline in synergistic potential may reflect seasonal changes in EO composition, especially the reduction of compounds that facilitate membrane disruption or ergosterol accessibility. For C. krusei, the interaction remained additive throughout the entire flowering period (FICI = 0.75), with stable MIC values and no significant seasonal fluctuation, suggesting a moderate but consistent EO–drug interaction that was insufficient to reach synergy. The most resistant profile was observed in C. parapsilosis, where the interaction remained indifferent across all months (FICI = 1.25), with no gain in amphotericin B activity. The limited synergistic activity observed with amphotericin B, particularly in C. parapsilosis, and C. albicans interactions, may be attributed to the polyene mode of action of the antibiotic. Amphotericin B binds to ergosterol in fungal cell membranes to form pores, thereby increasing membrane permeability and causing cell death [45]. It appears that certain components of rosemary EO may intercalate into the fungal lipid bilayer, altering its structural integrity and membrane environment. This modification could inadvertently reduce amphotericin B's access to ergosterol or disrupt membrane fluidity in a manner that impairs the formation of transmembrane pores. Consequently, rather than potentiating the antifungal activity of amphotericin B, co-treatment with rosemary EO may partially antagonize its binding efficiency, thereby diminishing the expected synergistic effect as reported in previous studies [45]. Consequently, rather than potentiating the antifungal activity of amphotericin B, co-treatment with rosemary EO may partially antagonize its binding efficiency, thereby diminishing the expected synergistic effect.
Conclusions
This study provides compelling evidence that the antimicrobial potential of R. officinalis EOs is profoundly influenced by seasonal variation during the flowering period in arid Mediterranean conditions. The findings clearly demonstrate that EOs collected during autumn, particularly in October and November with relatively high climatic water stress, exhibit the highest yields and superior antimicrobial efficacy, especially against Candida species and the Gram-positive S. aureus. This enhanced bioactivity was closely associated with elevated levels of oxygenated monoterpenes, in particular 1,8-cineole and camphor. Importantly, the synergistic assays revealed that rosemary EOs markedly potentiated the antibacterial action of streptomycin, achieving up to a 32-fold MIC reduction against E. coli in autumn, suggesting a promising role in overcoming antibiotic resistance in this Gram-negative bacterium. However, this synergy was more modest in S. aureus and more variable in S. enterica, reflecting strain-specific differences EO susceptibility. In contrast, the co-administration of rosemary EOs with amphotericin B yielded limited effects against Candida spp. These findings suggests that rosemary EOs harvested in early autumn, enriched in oxygenated monoterpenes, may act as effective natural adjuvants to enhance the efficacy of antibiotics and potentially enabling dose reductions that minimize toxicity. Standardizing harvest time to early autumn could thus optimize the therapeutic value of rosemary EO formulations and support their integration into antimicrobial strategies targeting some resistant multidrug resistant bacteria, including the Gram-negative E. coli. To facilitate their clinical integration, further in vivo pharmacological evaluations, toxicological assessments, and well-designed clinical trials are necessary to validate their safety, bioavailability, and synergistic efficacy when co-administered with conventional antibiotics. Nevertheless, the partial antagonism observed with amphotericin B warrants caution and highlights the need for strain-specific and drug-specific compatibility assessments before clinical use. These interactions must be considered when designing combinatory therapies to avoid compromising drug efficacy.
Use of AI tools declaration
The authors declare they have not used Artificial Intelligence (AI) tools in the creation of this article.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mc Connell MJ Where are we with monoclonal antibodies for multidrug-resistant infections?Drug Discov Today 2019241132113810.1016/j.drudis.2019.03.00230853568 · doi ↗ · pubmed ↗
- 2Fong IW Antimicrobial resistance: A crisis in the making New Antimicrobials: For the Present and the Future Cham Springer International Publishing 202312110.1007/978-3-031-26078-0_1 · doi ↗
- 3Vasireddy L Bingle LEH Davies MS Antimicrobial activity of essential oils against multidrug-resistant clinical isolates of the Burkholderia cepacia complex PLOS ONE 201813 e 020183510.1371/journal.pone.020183530071118 PMC 6072103 · doi ↗ · pubmed ↗
- 4Karnwal A Malik T Exploring the untapped potential of naturally occurring antimicrobial compounds: Novel advancements in food preservation for enhanced safety and sustainability Front Sustain Food Syst 20248130721010.3389/fsufs.2024.1307210 · doi ↗
- 5Ju J Xie Y Yu H Synergistic interactions of plant essential oils with antimicrobial agents: A new antimicrobial therapy Crit Rev Food Sci Nutr 2022621740175110.1080/10408398.2020.184649433207954 · doi ↗ · pubmed ↗
- 6Burt S Essential oils: Their antibacterial properties and potential applications in foods—a review Int J Food Microbiol 20049422325310.1016/j.ijfoodmicro.2004.03.02215246235 · doi ↗ · pubmed ↗
- 7Seow YX Yeo CR Chung HL Plant essential oils as active antimicrobial agents Crit Rev Food Sci Nutr 20145462564410.1002/ptr.662224261536 · doi ↗ · pubmed ↗
- 8Sharifi-Rad J Ezzat SM El Bishbishy MH Rosmarinus plants: Key farm concepts towards food applications Phytother Res 2020341474151810.1002/ptr.662232058653 · doi ↗ · pubmed ↗