Bioproduction of N6-benzyladenine N9-β-D-glucopyranoside using suspension cells of bamboo (Phyllostachys nigra)
Taiji Nomura, Karin Okazaki, Mikihisa Umehara, Yasuo Kato

TL;DR
Bamboo cells can produce a specific cytokinin derivative, BA-9G, which may be useful as a plant growth regulator due to its moderate activity and lower toxicity.
Contribution
Discovery of a regio-specific N-glucosyltransferase in bamboo cells for producing BA-9G.
Findings
Bamboo cells specifically produce BA-9G when cultured with N6-benzyladenine.
BA-9G shows moderate cytokinin activity and lower cytotoxicity compared to BA.
Bamboo cells may offer a viable system for bioproducing the rare cytokinin derivative BA-9G.
Abstract
Suspension-cultured cells of a temperate bamboo species (Phyllostachys nigra) accumulate substantial amounts of hydroxycinnamic acid derivatives and lignin under culture conditions that promote xylogenesis. In our previous study, we found a metabolite specifically produced in bamboo cells cultured under lignification-inducing conditions in a medium containing N6-benzyladenine (BA), but the chemical structure was not elucidated. In this study, we purified and identified this compound as BA N9-β-D-glucopyranoside (BA-9G). Despite the presence of three nitrogen positions (N-3, N-7, and N-9) that may be glucosylated in the adenine moiety of BA, bamboo cells specifically produced BA-9G (i.e., without other glucoside types) when cells were cultured in the presence of BA. This finding suggests that bamboo cells possess a regio-specific N-glucosyltransferase for catalyzing cytokinin glucoside…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
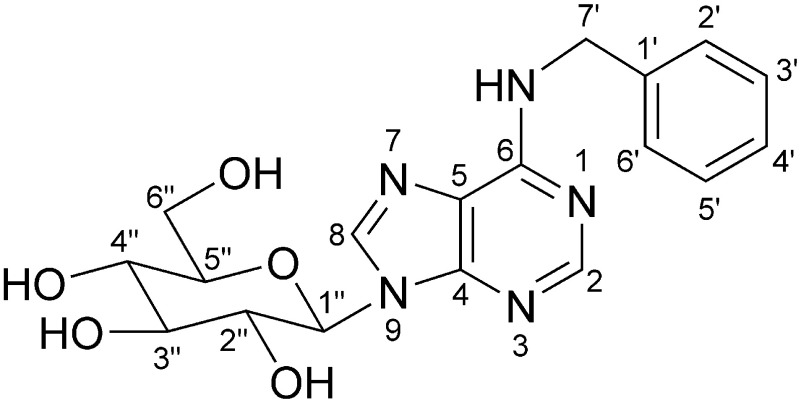 Figure 1
Figure 1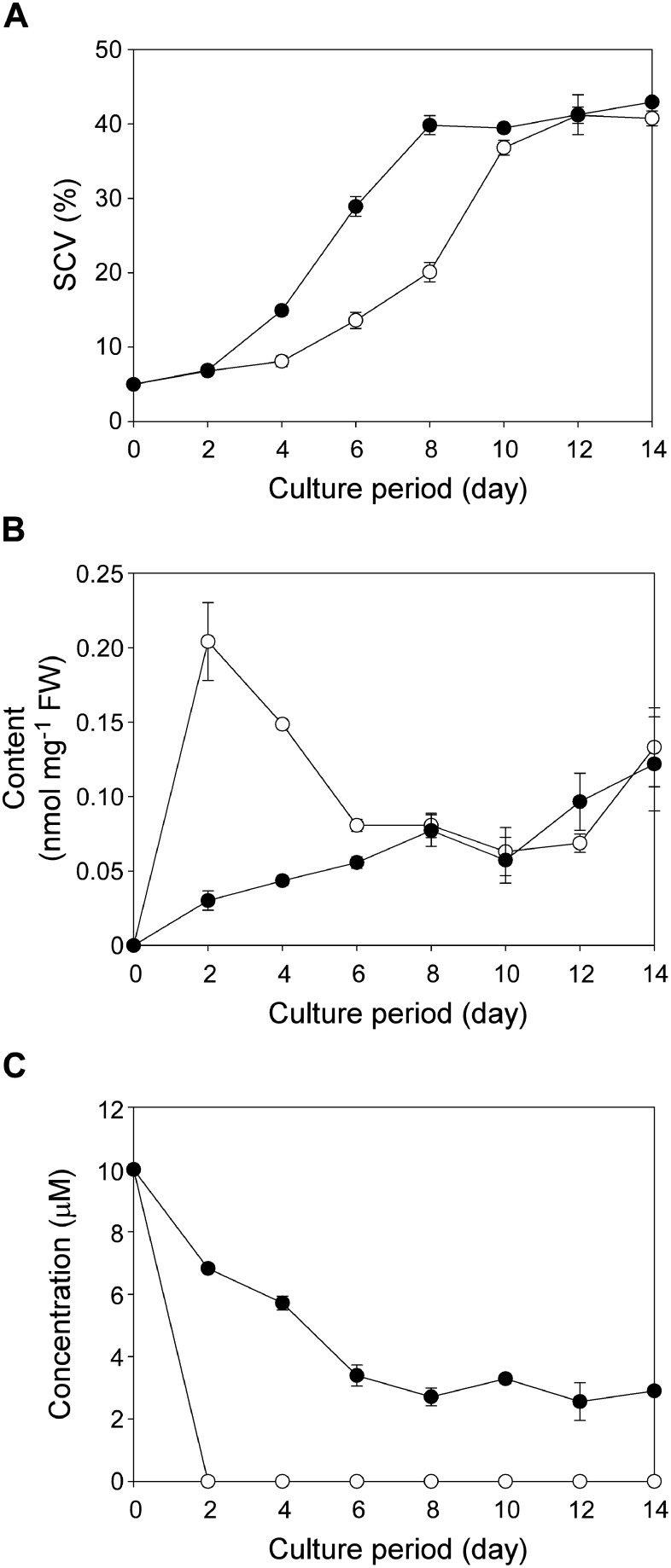 Figure 2
Figure 2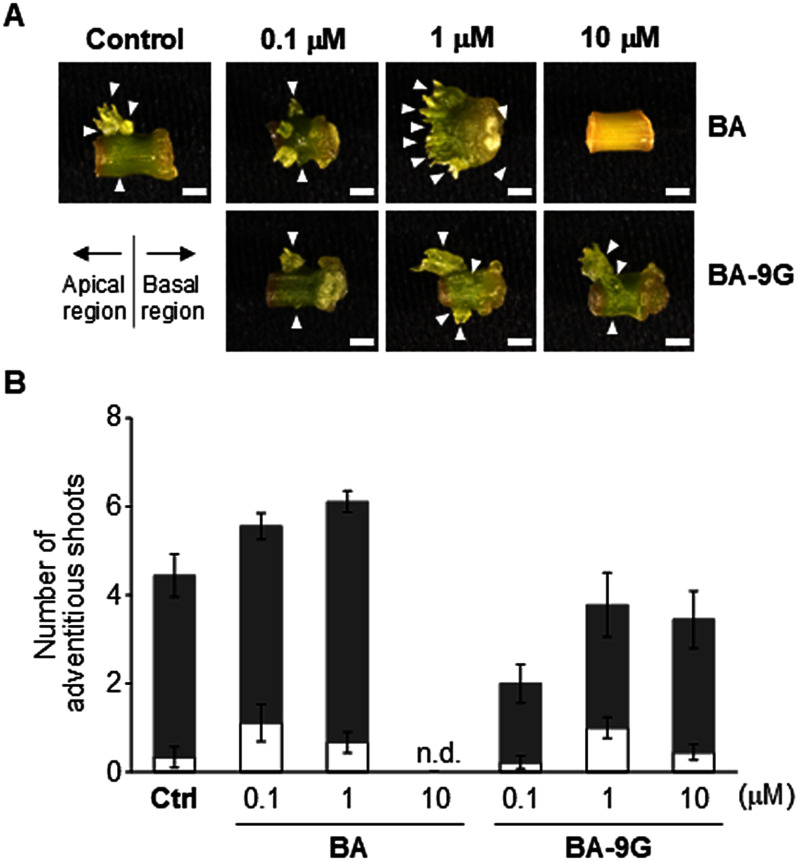 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBamboo properties and applications · Plant tissue culture and regeneration
Suspension-cultured cells of a temperate bamboo species (Phyllostachys nigra; Pn), which we established previously, are suitable for the bioproduction of phenylpropanoid-derived compounds via a rational metabolic-flow switching strategy (Kitaoka et al. 2020, 2021; Nomura et al. 2018; Ube et al. 2024). This strategy exploits Pn cell growth and metabolic features. Notably, the cell phase for rapid proliferation (inactive secondary metabolism) or active secondary metabolism (reduced proliferation) can easily be controlled by changing the culture medium (Nomura et al. 2013; Ogita et al. 2012). Pn cells cultured in half-strength (1/2) Murashige and Skoog (MS) medium (Murashige and Skoog 1962) supplemented with 10 µM N^6^-benzyladenine (BA), a synthetic cytokinin, accumulate substantial amounts of phenylpropanoid-derived metabolites, including hydroxycinnamoylputrescines and lignin (Nomura et al. 2013; Ogita et al. 2012).
During an earlier analysis of Pn cell extracts for secondary metabolite(s), we detected a compound that was specific to the extracts of Pn cells cultured in the presence of BA (designated as “Rt 7.0” on the basis of the high-performance liquid chromatography (HPLC) retention time; Nomura et al. 2013). We predicted that this compound (hereafter designated as compound 1) is a metabolite of BA, but the chemical structure was not determined. In this study, we first optimized culture conditions to enhance the production of compound 1 for its expedient isolation from cells, after which compound 1 was isolated and structurally characterized as a glucoside of BA with a limited availability. On the basis of the study results (e.g., comparative biological activity assay with BA), the metabolic and practical importance of this compound, which can be readily produced using Pn cells, is discussed.
BA was purchased from Fujifilm Wako Pure Chemical Corporation (Osaka, Japan). Authentic analytical standards of BA-glucosides, BA N^3^-β-D-glucopyranoside (BA-3G), BA N^7^-β-D-glucopyranoside (BA-7G), and BA N^9^-β-D-glucopyranoside (BA-9G), were purchased from Fujifilm Wako Pure Chemical Corporation, Olchemim (Olomouc, Czech Republic), and Toronto Research Chemicals (Toronto, ON, Canada), respectively.
Pn suspension cells, which are available from the RIKEN BioResource Research Center (no. rpc00047; https://web.brc.riken.jp/ (Accessed Mar 22, 2025)), were maintained and subcultured every 2 weeks as previously described (Nomura et al. 2013). Cell densities were determined according to the sedimented cell volume (SCV) as previously described (Ogita et al. 2011). To optimize culture conditions conducive to the production of compound 1, subcultured Pn suspension cells were transferred to the following fresh medium with an initial cell density of 5, 10, or 20% SCV: 1/2 MS medium containing 3% (w/v) sucrose and 1, 10, 100, or 250 µM BA. Cells were cultured, with samples collected every 2 days for 14 days as previously described (Nomura et al. 2013).
To analyze Pn cell extracts for compound 1, cells (approximately 100 mg fresh weight; FW) were mixed with 10 volumes of MeOH containing 2% (v/v) AcOH for a 10-min ultrasonication at room temperature. The resulting extract was centrifuged (21,500×g, 10 min, 4°C), and the supernatant was analyzed using a reversed-phase HPLC system as previously described (Nomura et al. 2013). For the HPLC analysis of the culture medium, the filtrate obtained after suspension-cultured cells were collected on filter paper was passed through a Millex-HV filter (0.45 µm; Merck, Darmstadt, Germany). The filtrate was analyzed by HPLC.
We first optimized Pn cell culture conditions for the high-level production of compound 1. Pn suspension cells were cultured in the presence of different concentrations of BA with an initial cell density of 5% SCV. Cell growth levels were similar between 1 µM and 10 µM BA (original concentration; Nomura et al. 2013), but decreased substantially at BA concentrations of 100 µM and 250 µM (Supplementary Figure S1A). Although we also cultured Pn suspension cells in the presence of 500 µM BA, the cells became bleached and were non-viable, indicating that the BA concentration was excessive for the cells. The production of compound 1 increased as the concentration of BA added to the culture medium increased (Supplementary Figure S1B), indicating that Pn cells can produce compound 1 at BA concentrations up to 250 µM, despite the suppressed cell growth at high BA concentrations.
Considering these results, we next examined the effects of the initial cell density on the production of compound 1 in culture medium in which the initial BA concentration was 250 µM. As shown in Supplementary Figure S2A, cell growth recovered when the initial cell density was 20% SCV, suggesting that Pn cells metabolized BA to a less cytotoxic form (i.e., compound 1). In fact, increases in the initial cell density accelerated the decrease in BA levels in the culture medium (Supplementary Figure S2B). The compound 1 content per cell FW started to increase immediately after starting the culture (Supplementary Figure S2C). By multiplying the compound 1 content by the cell FW at each collection time point, the highest production titer per liter culture was obtained on day 4 for the culture with an initial cell density of 20% SCV (Supplementary Figure S2D), which coincided with the complete consumption of BA in the culture medium (Supplementary Figure S2B). On the basis of these results, we decided to culture Pn cells for 4 days with an initial cell density of 20% SCV in the presence of 250 µM BA for the expedient isolation of compound 1 (Supplementary Figure S3).
We purified 122 mg compound 1 from Pn suspension cells (187 g FW) obtained from a 2.3-l culture under optimized conditions according to the procedures described in Supplementary Methods. The NMR spectra of compound 1 were recorded using an AVANCE 400 spectrometer (Bruker, Karlsruhe, Germany), with a mixture comprising 0.5 ml (CD_3_)2_SO and 0.1 ml D_2_O (for ^1^H-NMR) or (CD_3)2_SO (for ^13^C-NMR) serving as the solvent. The UV spectrum was measured using a U-2000 spectrophotometer (Hitachi, Tokyo, Japan), with H_2_O serving as the solvent. The high-resolution electrospray ionization time-of-flight mass spectrometry (HR-ESI-TOF-MS) spectrum was obtained using a micrOTOF focus spectrometer (Bruker) and a published direct infusion method (Nomura and Kato 2020). The spectral properties of purified compound 1 were as follows: HR-ESI-TOF-MS (Supplementary Figure S4) (positive) m/z (relative intensity) 388.1634 [M+H]^+^ (100) (calcd. for C_18_H_22_N_5_O_5^+^, 388.1615), m/z 410.1453 [M+Na]^+^ (47) (calcd. for C_18_H_21_N_5_O_5_Na^+^, 410.1435); UV (H_2_O) λ_max_ (logε) 268 nm (4.28); ^1^H-NMR (400 MHz, (CD_3_)2_SO 0.5 ml + D_2_O 0.1 ml, Supplementary Figure S5A): δ (ppm) 3.32–4.03 (6H, m, H-2″, 3″, 4″, 5″, 6″), 4.74 (2H, brs, H-7′), 5.44 (1H, d, J=9.4 Hz, H-1″), 7.21–7.37 (5H, m, H-2′, 3′, 4′, 5′, 6′), 8.22 (1H, s, H-8), 8.33 (1H, s, H-2); ^13^C-NMR (100 MHz, (CD_3)_2_SO, Supplementary Figure S5B): δ (ppm) 43.3 (C-7′), 61.4 (C-6″), 70.3 (C-4″), 71.7 (C-2″), 77.7 (C-5″), 80.5 (C-3″), 83.3 (C-1″), 119.5 (C-5), 127.0 (C-4′), 127.5 (C-2′, 6′), 128.7 (C-3′, 5′), 140.2 (C-1′), 140.6 (C-8), 149.7 (C-4), 153.0 (C-2), 154.9 (C-6).
The molecular formula (C_18_H_21_N_5_O_5_) and molecular weight (387) determined on the basis of the HR-ESI-TOF-MS data (Supplementary Figure S4) indicated that compound 1 is a glucoside of BA. Glucosylation of cytokinins, N^6^-substituted adenine derivatives, occurs at the hydroxy group of the N^6^-substituent (i.e., hydroxylated isoprenoid type) and at the N^3^, N^7^, or N^9^ position of the purine moiety (Chen et al. 2021). Because BA does not have a glucosylation site on the N^6^-benzyl moiety, compound 1 was predicted to be one of the three N-glucosides. Thus, compound 1 was analyzed by HPLC for a comparison of its retention time with that of the authentic N-glucosides of BA (BA-3G, BA-7G, and BA-9G), which indicated that compound 1 is BA-9G (Supplementary Figure S6). Moreover, in the ^1^H-NMR spectrum of compound 1, the doublet signal of an anomeric proton (H-1″) at 5.44 ppm (J=9.4 Hz) reflected the β-configuration of C-1″. Considered together, these findings suggest that compound 1 is N^6^-benzyladenine N^9^-β-D-glucopyranoside (BA-9G, Figure 1). The spectral data of compound 1 were in accordance with published data (Cowley et al. 1978; Duke and MacLeod 1978; Hashizume and Yoshida 1976).
We previously demonstrated that adding 10 µM BA to the culture medium promotes the xylogenesis of Pn cells via the accumulation of lignin (Nomura et al. 2013; Ogita et al. 2012). To examine whether the same phenomenon is observed when BA-9G is added, Pn cells were cultured in the presence of 10 µM BA-9G with an initial cell density of 5% SCV. As shown in Figure 2A, Pn cells cultured in medium containing 10 µM BA-9G started to proliferate earlier than those cultured in medium supplemented with 10 µM BA. This is likely because BA-9G is less cytotoxic or biologically active than BA. When BA was added to the medium, the BA-9G content in cells peaked on day 2 (Figure 2B), which was consistent with the complete consumption of BA in the culture medium in 2 days (Figure 2C). The addition of 10 µM BA-9G to the culture medium resulted in a gradual increase in the BA-9G content in cells (Figure 2B). Moreover, the BA-9G content in the culture medium gradually decreased, but was not completely consumed during the 14-day culture period (Figure 2C). BA-9G was incorporated into cells less efficiently than BA likely because BA-9G is more hydrophilic than BA. Staining cells with a phloroglucinol–HCl reagent indicated that BA-9G can also promote Pn cell lignification (Supplementary Figure S7). Although lignification appeared to be slightly delayed under the BA-9G condition, the extent of lignification (i.e., intensity of the red staining) was similar between the BA and BA-9G conditions.
We next examined the biological activity of BA-9G as a cytokinin using the adventitious shoot formation system involving cultured internodal segments of ipecac (Carapichea ipecacuanha) (Okazaki et al. 2025). These segments form adventitious shoots under phytohormone-free culture conditions, but shoot formation is biased toward the apical region because of differences in the endogenous cytokinin/auxin proportions between the apical and basal regions, with cytokinin/auxin proportions higher and lower in the apical and basal regions, respectively (Koike et al. 2017). Thus, this internode culture system is considered to be appropriate for evaluating the biological activity of exogenously applied cytokinins. The assay was performed as previously described (Okazaki et al. 2025). Briefly, 2.5-mm internodal segments were cut from sterile ipecac plants cultured in vitro. The segments were placed on B5 medium containing 0.1–10 µM BA-9G or BA and solidified with 0.8% (w/v) agar in a Petri dish (90 mm × 20 mm), which was then incubated at 24°C under a 14-h light/10-h dark photoperiod (10–20 µmol photons m^−2^ s^−1^). After culturing the internodal segments for 5 weeks, the number of adventitious shoots (longer than 0.3 mm) was determined for the apical and basal halves of each segment. As shown in Figure 3, adventitious shoots formed mainly in the apical region of internodal segments. When these segments were cultured in the presence of BA, the number of adventitious shoots increased as the BA concentration increased up to 1 µM, but adventitious shoots did not form at 10 µM. By contrast, a treatment with 0.1 µM BA-9G suppressed adventitious shoot formation (relative to the control). However, the number of adventitious shoots increased following the treatment with 1 µM BA-9G. Notably, 10 µM BA-9G induced adventitious shoot formation, which was in contrast to the fatal effect of BA at the same concentration, although the number of shoots was similar to that observed at 1 µM. These findings along with the observed effects on Pn cells suggest that BA-9G acts as a cytokinin with moderate biological activity and reduced cytotoxicity at high concentrations.
N-Glucosides of cytokinins have been identified in a wide variety of plant species (Hoyerová and Hošek 2020). Although the purine moiety can be glucosylated at the N^3^, N^7^, or N^9^ position, N^7^- and N^9^-glucosides are the predominant forms, whereas N^3^-glucosides are relatively rare (Chen et al. 2021; Hoyerová and Hošek 2020). The cytokinin activity of N^3^-glucosides markedly exceeds that of N^7^- and N^9^-glucosides, likely because N^3^-glucosides are more susceptible to the hydrolysis catalyzed by β-glucosidase to form the free base than N^7^- and N^9^-glucosides (Letham et al. 1975). Although N^7^- and N^9^-glucosides have historically been considered to be inactive or have low to negligible biological activity, a contrasting view has also been reported (Chen et al. 2021). It is still controversial whether N-glucosides are biologically active or if they need to be hydrolyzed to their free base (Chen et al. 2021; Hošek et al. 2020; Hoyerová and Hošek 2020). Despite this uncertainty, our results suggest that BA-9G may be useful as a plant growth regulator with moderate biological activity and reduced cytotoxicity (relative to free BA).
However, because of its limited availability, BA-9G is relatively expensive, which is not conducive to its widespread use. In this study, the maximum production titer of BA-9G per liter culture (set to 100% in Supplementary Figure S2D) was calculated to be approximately 50 mg; this was produced after only 4 days. BA-9G bioproduction using Pn cells may be better than chemical synthesis, which requires complex multistep reactions for regio-specific N-glucosylation (Cowley et al. 1978), in terms of simplicity and low environmental loads. This bioproduction system may increase the availability of BA-9G, a highly expensive and rare cytokinin derivative. Future studies should examine whether this system is also applicable to the bioproduction of N^9^-glucosides of natural cytokinins (e.g., trans/cis-zeatin, isopentenyl adenine, and dihydrozeatin) with isoprenoid-type N^6^-substituents.
According to earlier research, N^7^- and N^9^-glucosides of cytokinins are often found together (Chen et al. 2021; Hošek et al. 2020; Hoyerová and Hošek 2020). To the best of our knowledge, the accumulation of N^9^-glucoside unaccompanied by N^7^-glucoside, as observed in this study, is quite rare. Indeed, the regio-specific N-glucosyltransferase of cytokinins has not been reported (Hou et al. 2004). Considering that neither BA-7G nor BA-3G were found in the extract of BA-treated Pn cells, Pn cells likely contain an N^9^-specific glucosyltransferase. This enzyme is currently being analyzed in our laboratory.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chen L, Zhao J, Song J, Jameson PE (2021) Cytokinin glucosyl transferases, key regulators of cytokinin homeostasis, have potential value for wheat improvement. Plant Biotechnol J 19: 878–89633811433 10.1111/pbi.13595 PMC 8131048 · doi ↗ · pubmed ↗
- 2Cowley DE, Duke CC, Liepa AJ, Mac Leod JK, Letham DS (1978) The structure and synthesis of cytokinin metabolites. I The 7- and 9-β-D-glucofuranosides and pyranosides of zeatin and 6-benzylaminopurine. Aust J Chem 31: 1095–1111
- 3Duke CC, Mac Leod JK (1978) The structure and synthesis of cytokinin metabolites. III 13C N.M.R. of cytokinin glycosides. Aust J Chem 31: 2219–2223
- 4Hashizume T, Yoshida K (1976) Glycosylation of N 6-benzyladenine by fusion procedure using bis-(p-nitrophenyl)hydrogen phosphate. Agric Biol Chem 40: 2001–2004
- 5Hošek P, Hoyerová K, Kiran NS, Dobrev PI, Zahajská L, Filepová R, Motyka V, Müller K, Kamínek M (2020) Distinct metabolism of N-glucosides of isopentenyladenine and trans-zeatin determines cytokinin metabolic spectrum in Arabidopsis. New Phytol 225: 2423–243831682013 10.1111/nph.16310 · doi ↗ · pubmed ↗
- 6Hou B, Lim E-K, Higgins GS, Bowles DJ (2004) N-Glucosylation of cytokinins by glucosyltransferases of Arabidopsis thaliana. J Biol Chem 279: 47822–4783215342621 10.1074/jbc.M 409569200 · doi ↗ · pubmed ↗
- 7Hoyerová K, Hošek P (2020) New insight into the metabolism and role of cytokinin N-glucosides in plants. Front Plant Sci 11: 74132582261 10.3389/fpls.2020.00741 PMC 7292203 · doi ↗ · pubmed ↗
- 8Kitaoka N, Nomura T, Ogita S, Kato Y (2020) Bioproduction of glucose conjugates of 4-hydroxybenzoic and vanillic acids using bamboo cells transformed to express bacterial 4-hydroxycinnamoyl-Co A hydratase/lyase. J Biosci Bioeng 130: 89–9732192841 10.1016/j.jbiosc.2020.02.010 · doi ↗ · pubmed ↗