A start codon-targeted genome editing strategy for generating hypomorphic mutants of lethal plant genes
Mika Yoshimura, Tsubasa Mamiya, Naoki Takahashi, Takashi Ishida

TL;DR
This paper introduces a genome editing strategy to study essential plant genes by creating partial-function mutants, avoiding lethal effects of complete gene loss.
Contribution
A novel CRISPR-based strategy using start codon removal to generate hypomorphic alleles of essential genes in plants.
Findings
Start codon removal in the NSE1 gene caused severe developmental defects and genome instability in Arabidopsis.
Homozygous mutants showed increased DNA fragmentation and cell death in root meristems.
Complementation assays confirmed partial gene function remains in these hypomorphic alleles.
Abstract
In plants, the functional characterization of essential genes is often hindered by the lethality associated with complete loss-of-function alleles. Here, we present a genome editing-based strategy to generate viable hypomorphic alleles through selective removal of the translation start codon. Using Arabidopsis thaliana NON-SMC ELEMENT 1, which encodes a conserved component of the Structural Maintenance of Chromosomes (SMC) 5/6 complex involved in DNA repair and genome stability, as a model, we generated CRISPR-Cas9-edited alleles lacking the start codon. These homozygous mutants exhibited severe developmental defects, including stunted growth and failure to form true leaves. Moreover, they displayed molecular hallmarks of genome instability, such as increased DNA fragmentation, upregulation of DNA repair and cell cycle checkpoint genes, and root meristem cell death. Complementation…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
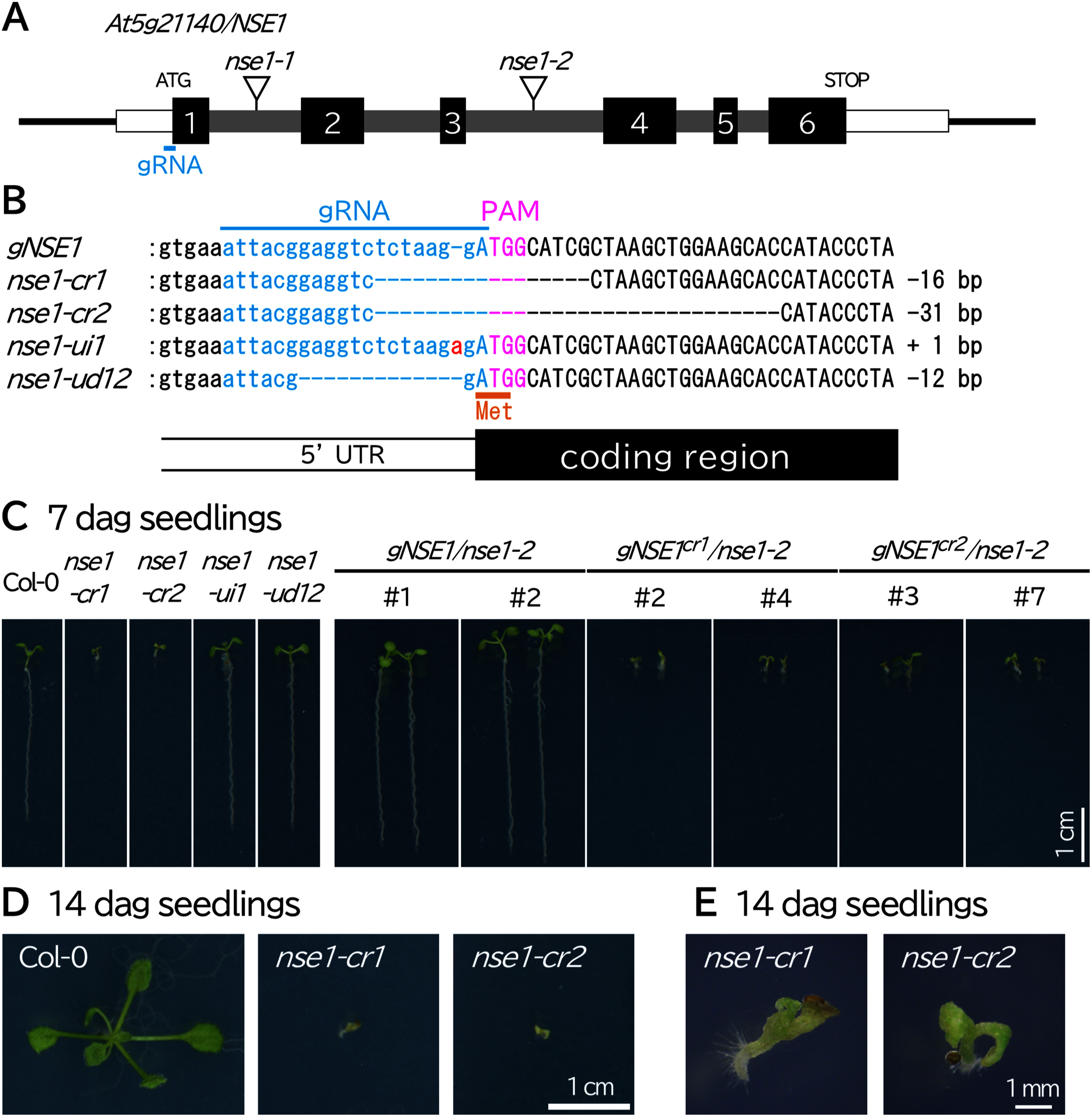 Figure 1
Figure 1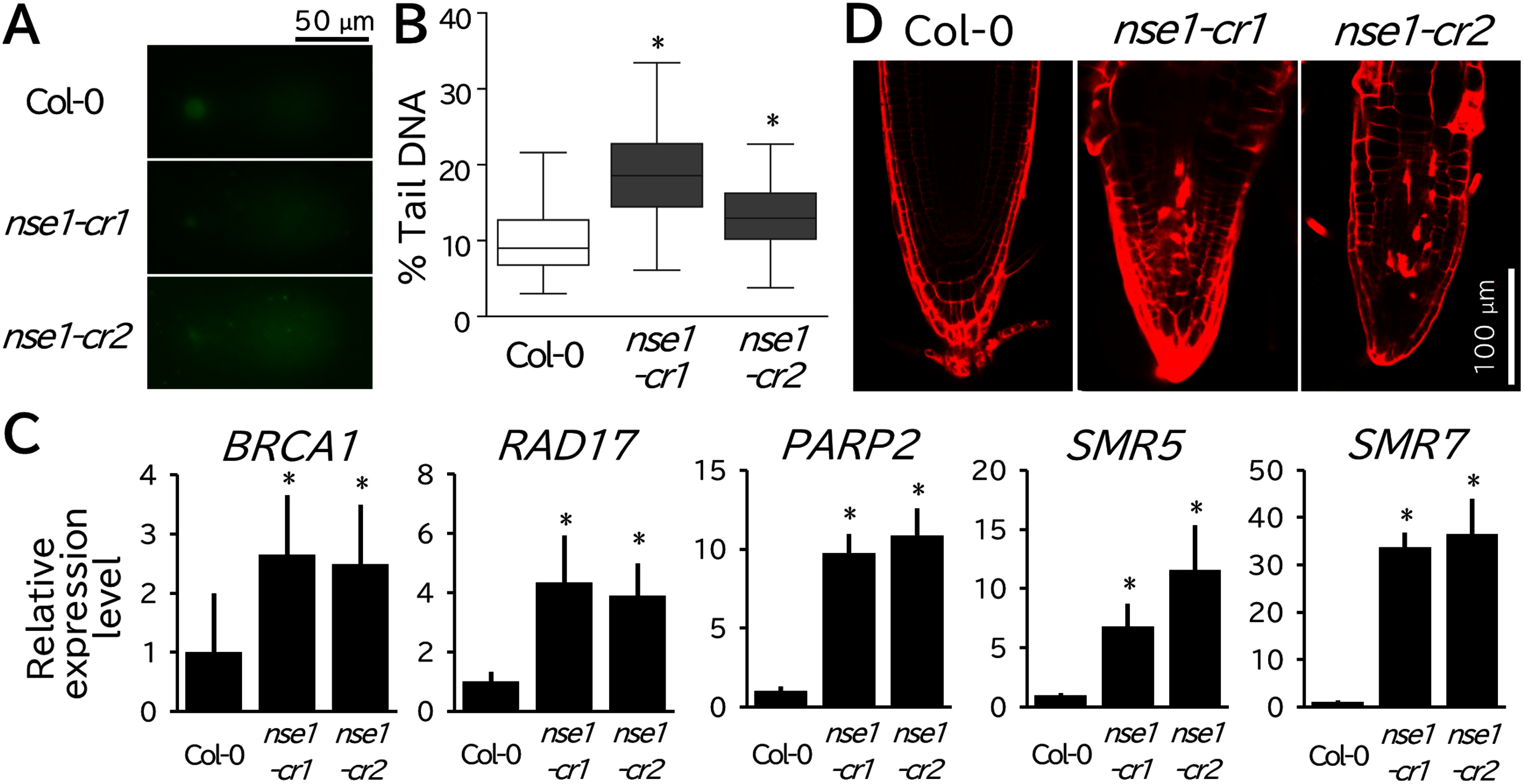 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCRISPR and Genetic Engineering · RNA and protein synthesis mechanisms · Photosynthetic Processes and Mechanisms
Functional analyses of specific genes in plant science require meticulous genetic analysis. Bioresources, such as a collection of mutants, have been used to accelerate our understanding of gene function, and recent advances in genome editing have revolutionized multiple fields in the life sciences. However, many mutants largely eliminate the function of the gene of interest, which makes them unsuitable for studying genes where complete loss-of-function is lethal. To overcome this difficulty, gene knockdown technologies such as RNAi are often used to elucidate gene function. Recently, we developed a method for creating viable mutant alleles by using CRISPR-Cas9 to introduce mutations into a splicing acceptor site (Yoshimura and Ishida 2025). However, since the applicability of this strategy may vary depending on gene structure, we now propose a new methodology that makes hypomorphic allele generation more versatile through genome editing, thereby stimulating alternative approaches for producing diverse mutants.
In any given gene, if the primary start codon functions incorrectly during translation, initiation from an alternative start codon may produce a protein that is incomplete and retains partial functionality (Makino et al. 2016). Based on this observation, we hypothesized that the removal of a start codon via genome editing can generate hypomorphic alleles. To test this possibility, we targeted the Arabidopsis thaliana NON-SMC ELEMENT 1 (NSE1) gene. The protein encoded by NSE1, which was originally identified via its yeast homolog, is a component of the conserved eukaryotic SMC5/6 complex (Fujioka et al. 2002). This complex is known to play a crucial role in DNA repair and maintaining genome stability (Aragón 2018; Diaz and Pecinka 2018). In plants, the critical importance of the Structural Maintenance of Chromosomes (SMC) 5/6 complex is well documented. For example, loss-of-function mutations in its components generally result in embryonic lethality or, if embryos are viable, severe developmental defects (Diaz and Pecinka 2018; Ishida et al. 2009; Yang and Pecinka 2022). Moreover, two T-DNA insertion alleles in the intron of NSE1 have been reported, and both have been described as embryonic lethal mutants (Figure 1A) (Li et al. 2017, 2019). Consequently, analyses of NSE1 have largely been confined to studies of embryogenesis, with post-germination functions assessed using transgenic plants in which ectopic expression from non-native promoters is used to partially rescue the mutant phenotype. Given this context, we posited that generating hypomorphic alleles of NSE1 could substantially aid in its functional dissection.
Accordingly, we used the CRISPRdirect online tool to design guide RNAs capable of inducing cleavage immediately upstream of the translation initiation codon. Subsequently, we constructed genome editing vectors employing the pDe-Cas9 system using a previously described method (Fauser et al. 2014; Naito et al. 2015; Yamaguchi et al. 2017; Yamamoto et al. 2019). Next, in the T_2_ generation, we isolated mutants harboring either a 16-bp or a 31-bp deletion in one allele; these were designated as nse1-cr1 and nse1-cr2, respectively (Figure 1B, Supplementary Figure S1). Both alleles likely lack the first coding region that contains the start codon. Intriguingly, we recovered homozygous seedlings from heterozygote parents of nse1-cr1 and nse1-cr2, indicating that these mutant alleles remain viable, at least during embryogenesis (Figure 1C). Thus, in contrast to the previously reported T-DNA insertion alleles, which are embryonically lethal, these mutants were classified as hypomorphic alleles.
We further characterized the morphology of the nse1-cr1 and nse1-cr2 mutants. To this end, mutant seedlings were grown for seven days on half-strength MS agar medium; they were found to be markedly dwarfed relative to wild-type seedlings (Figure 1C). Moreover, at 14 days after germination, most mutants failed to develop true leaves, and their roots were so reduced that they were barely visible under a dissecting microscope (Figure 1D, E). Finally, none survived to the reproductive stage. Next, we took advantage of the availability of viable homozygous mutant seedlings and investigated the biological relevance of NSE1. Using comet assays to quantify DNA integrity loss, we found that nse1-cr1 and nse1-cr2 mutants exhibited pronounced DNA fragmentation (Figure 2A, B). Furthermore, the expression levels of the DNA repair genes BRCA1, RAD17, and PARP2, which are induced by DNA damage, were strongly elevated in both mutants (Figure 2C) (Ogita et al. 2018). Similarly, SMR5 and SMR7, which are two important regulators of the cell cycle checkpoint and are expressed in response to DNA damage, were strongly upregulated in mutant seedlings (Figure 2C) (Yi et al. 2014). As part of the DNA damage response, we also observed cell death in the root apical meristem (Figure 2D). Overall, these findings are consistent with those of previous studies employing an ectopic NSE1 expression line and further highlighting the usefulness of our strategy for enabling the recovery of viable hypomorphic mutants for functional analysis.
In parallel, we isolated additional genome-edited alleles. One, nse1-ui1, carried a 1-bp insertion, and another, nse1-ud12, carried a 12-bp deletion; both mutations were located in their respective 5′ UTR regions (Figure 1B, Supplementary Figure S1). Interestingly, homozygous plants harboring these alleles exhibited phenotypes that were nearly indistinguishable from the wild-type, thereby reinforcing the notion that loss of the start codon is critical for generating hypomorphic alleles (Figure 1C).
Notably, the NSE1 sequences of nse1-cr1 and nse1-cr2 exhibited residual functional capacity, indicating that they are not nonfunctional. To further examine these functions, we conducted a complementation assay using a genomic fragment of NSE1. Specifically, we cloned a 4,646-bp fragment and introduced it into a nse1-2 mutant. This fully restored normal seedling development confirms that the fragment was sufficient to restore NSE1 function (Figure 1C). We then engineered modified genomic sequences with deletions analogous to those found in the nse1-cr1 and nse1-cr2 mutants by site-directed mutagenesis. When these constructs were transformed into the nse1-2 mutant, they reproduced a dwarf phenotype reminiscent of that observed in the original hypomorphic alleles (Figure 1C). Taken together, these findings provide genetic evidence that the nse1-cr1 and nse1-cr2 alleles compromise but do not eliminate NSE1 function, consistent with their classification as hypomorphic.
In conclusion, this study demonstrates that targeted removal of a start codon via genome editing is an effective and broadly applicable strategy for generating hypomorphic alleles in Arabidopsis. By directly disrupting translation initiation, this approach enables the generation of viable mutant alleles even in genes for which complete loss-of-function is embryonically lethal—an outcome often unattainable with conventional knockout or RNAi-based methods. On the other hand, a key limitation of this strategy lies in the uncertainty regarding the exact transcripts and protein products generated by the mutant alleles. Nonetheless, the method offers a significant advantage in its high degree of design flexibility. Similar outcomes can likely be achieved using other gene targeting technologies (e.g., TALENs) or DNA rewriting strategies, such as base editing or prime editing. Applying this approach to NSE1, a component of the SMC5/6 complex, we successfully recovered viable hypomorphic mutants that displayed characteristic defects in development and genomic integrity. These findings lay the groundwork for future investigations into the genetic interplay between NSE1—as a component of the SMC5/6 complex—and canonical DNA damage regulators. In particular, studies focusing on ATM and ATR, which are key kinases that sense DNA damage and initiate signaling cascades, and SOG1, a master regulator of the DNA damage response, will be important for understanding the broader regulatory network. Furthermore, our results may guide investigations into the molecular mechanisms underlying cell cycle checkpoint control mediated by SMR and related factors. Overall, our work expands the toolkit for functional gene analysis and offers a promising avenue for basic research and biotechnological applications in the future.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aragón L (2018) The Smc 5/6 complex: New and old functions of the enigmatic long-distance relative. Annu Rev Genet 52: 89–10730476445 10.1146/annurev-genet-120417-031353 · doi ↗ · pubmed ↗
- 2Diaz M, Pecinka A (2018) Scaffolding for repair: Understanding molecular functions of the SMC 5/6 complex. Genes 9: 3629329249 10.3390/genes 9010036 PMC 5793187 · doi ↗ · pubmed ↗
- 3Fauser F, Schiml S, Puchta H (2014) Both CRISPR/Cas-based nucleases and nickases can be used efficiently for genome engineering in Arabidopsis thaliana. Plant J 79: 348–35924836556 10.1111/tpj.12554 · doi ↗ · pubmed ↗
- 4Fujioka Y, Kimata Y, Nomaguchi K, Watanabe K, Kohno K (2002) Identification of a novel non-structural maintenance of chromosomes (SMC) component of the SMC 5-SMC 6 complex involved in DNA repair. J Biol Chem 277: 21585–2159111927594 10.1074/jbc.M 201523200 · doi ↗ · pubmed ↗
- 5Ishida T, Fujiwara S, Miura K, Stacey N, Yoshimura M, Schneider K, Adachi S, Minamisawa K, Umeda M, Sugimoto K (2009) SUMO E 3 ligase HIGH PLOIDY 2 regulates endocycle onset and meristem maintenance in Arabidopsis. Plant Cell 21: 2284–229719666737 10.1105/tpc.109.068072 PMC 2751947 · doi ↗ · pubmed ↗
- 6Li G, Zou W, Jian L, Qian J, Deng Y, Zhao J (2017) Non-SMC elements 1 and 3 are required for early embryo and seedling development in Arabidopsis. J Exp Bot 68: 1039–105428207059 10.1093/jxb/erx 016PMC 5441860 · doi ↗ · pubmed ↗
- 7Li G, Zou W, Jian L, Qian J, Zhao J (2019) At NSE 1 and At NSE 3 are required for embryo pattern formation and maintenance of cell viability during Arabidopsis embryogenesis. J Exp Bot 70: 6229–624431408172 10.1093/jxb/erz 373PMC 6859727 · doi ↗ · pubmed ↗
- 8Makino S, Fukumura R, Gondo Y (2016) Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas 9. Sci Rep 6: 3960828000783 10.1038/srep 39608 PMC 5175197 · doi ↗ · pubmed ↗