Insecticide resistance in Aedes aegypti and Aedes albopictus in southern Benin: quantification, investigation of kdr mutations, and detection of detoxification enzyme activity
Alphonse Keller Konkon, Rock Aikpon, Isidore Hoyochi, David Mahouton Zoungbédji, Arthur Sovi, Albert Sourou Salako, Camus Konkon, Brice Dangnon, Geoffroy Yahoue, Romuld Victoir Adjovi, Lokossou Antoine, Juvenal Ahouandjinou, Nsele Kisambu Grace, Bruno Adjottin

TL;DR
This study finds that Aedes aegypti mosquitoes in southern Benin are resistant to certain insecticides, while Aedes albopictus remains susceptible, highlighting the need for better resistance management.
Contribution
The study identifies specific kdr mutations and elevated detox enzyme activity in Aedes aegypti, contributing to pyrethroid resistance in southern Benin.
Findings
Aedes aegypti showed resistance to deltamethrin and permethrin but remained susceptible to bendiocarb.
Three kdr mutations (F1534C, V1016G, S989P) were found in Aedes aegypti with moderate allele frequencies.
Elevated oxidase, GST, and esterase activity was observed in Aedes aegypti populations.
Abstract
Insecticide resistance in arbovirus vectors threatens the effectiveness of vector control strategies in many endemic regions. Understanding resistance profiles and identifying underlying mechanisms are essential for preventing operational failures. This study assessed the susceptibility of Aedes aegypti and Aedes albopictus to commonly used insecticides in southern Benin and investigated the contribution of knockdown resistance mutations and metabolic detoxification pathways. Entomological surveillance was conducted in Lokossa, Bohicon, and Dassa, where ovitraps were used to collect eggs of Aedes species. Eggs were reared to the adult stage under controlled insectary conditions. Nonblood-fed females aged 2–5 days were tested using standard tube assays to determine susceptibility to deltamethrin at 0.05%, permethrin at 0.75%, and bendiocarb at 0.1%. Additional concentrations at fivefold…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1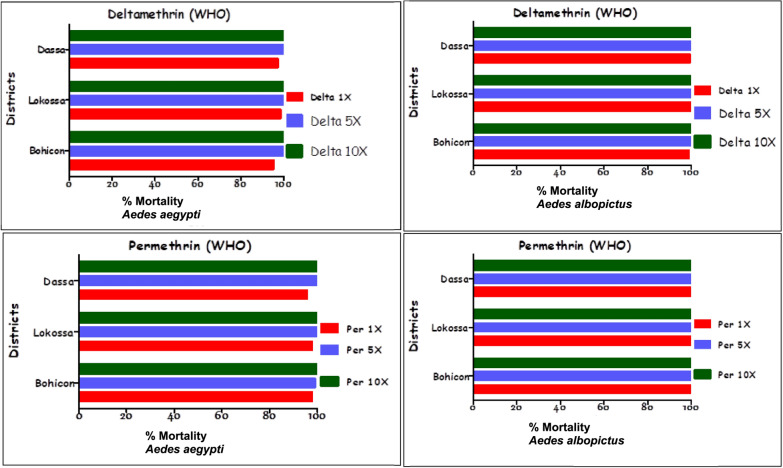 Figure 2
Figure 2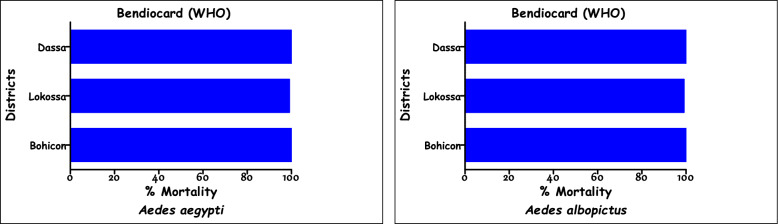 Figure 3
Figure 3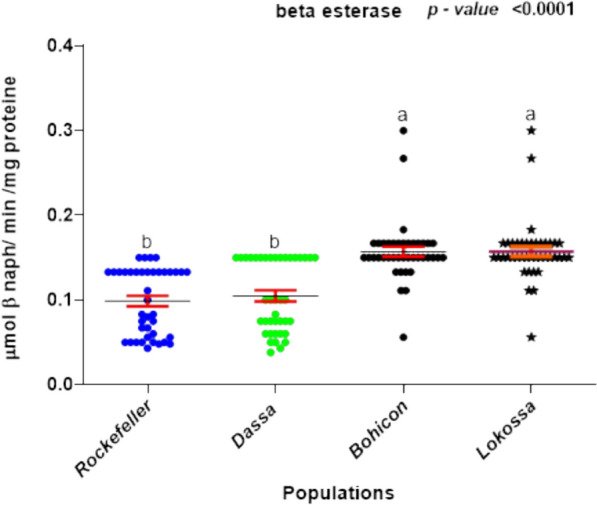 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Insect Resistance and Genetics · Insect Pest Control Strategies
Introduction
Insecticide resistance among arbovirus vectors poses a major challenge to vector control efforts aimed at reducing disease incidence and ultimately achieving elimination. Recent studies have reported a significant increase in resistance levels, particularly to pyrethroids [1, 2], and have linked this trend to genetic mutations in vector populations [2, 3]. In West Africa, the public health situation has become increasingly concerning. Countries such as Burkina Faso have experienced recurrent dengue outbreaks in recent years, including reports of dengue-related deaths [4–6]. According to the World Health Organization's 2024 report, over half of the global dengue cases were reported in Burkina Faso alone [7]. Other West African nations, including Côte d’Ivoire, Mali, Senegal, and Nigeria, have also been substantially affected by the disease [8–13]. Owing to its geographical proximity to these countries, Benin is considered a high-risk area and warrants close attention from public health authorities In Benin, between July and August 2010, two dengue cases were diagnosed in travelers returning to Cotonou from France. In addition, in 2019, Benin reported 11 dengue cases in the municipalities of Cotonou, Porto-Novo, Sèmè-Kpodji, and Abomey-Calavi, including two fatalities among the confirmed cases (one in Abomey-Calavi and one in Porto-Novo) [14, 15]. Dengue cases were confirmed in the human population in Benin as early as 2019 [16]. Despite this, dengue continues to be classified as a neglected tropical disease, largely owing to insufficient surveillance and limited policy focus. In 2023, a nationwide study documented the widespread distribution and abundance of major arbovirus vectors in Benin [17]. Further investigation in 2024 revealed the circulation of three distinct dengue virus serotypes within local mosquito populations [18]. This cocirculation poses a heightened risk, as secondary infection with a different serotype has been associated with more severe manifestations of the disease, including dengue hemorrhagic fever and dengue shock syndrome [19, 20]. Given this context, strengthening entomological surveillance is critical for preventing or responding rapidly to potential outbreaks. This study was, therefore, designed to assess the current status of insecticide resistance in Ae. aegypti and Ae. albopictus populations in southern Benin and to evaluate the operational implications for vector control strategies. In addition to guiding local interventions, this work addresses a key knowledge gap regarding the resistance status of Ae. albopictus to insecticides in the West African region.
Methodology
Study sites
The study was conducted in three communities of southern Benin: Lokossa (6°37′60″ N, 1°43′0″ E), Bohicon (7°11′00″ N, 2°04′00″ E), and Dassa-Zoumè (7°45′00″ N, 2°11′00″ E), as shown in the map of Benin (Fig. 1).Fig. 1. Study sites
Insecticide resistance status of Ae. aegypti and Ae. albopictus to pyrethroids and carbamates
Egg collection and mosquito rearing
Ovitraps were deployed in peridomestic environments across the three selected communities for a period of 7 days. The traps were then retrieved and transported to the insectary of the Centre de Recherche Entomologique de Cotonou (CREC). The collected eggs were immersed in water to stimulate hatching under favorable conditions. Following hatching, the larvae were reared under optimal insectary conditions (28 °C and ~ 80% relative humidity) [21]. Pupae were collected, pooled at the collection site, and transferred to holding cages for adult emergence. Adult mosquitoes were morphologically identified via taxonomic keys developed by Edwards and Huang [22, 23].
WHO tube bioassays for pyrethroids and carbamates
The susceptibility of female Ae. aegypti and Ae. albopictus, aged 2–5 days, to insecticides was assessed via the WHO tube test protocol. For each species, batches of 20 to 25 nonblood-fed females were aspirated from rearing cages and exposed for 60 min to diagnostic (1 ×), 5 × , and 10 × concentrations of deltamethrin, and permethrin, respectively. The diagnostic concentration of bendiocarb was used. The diagnostic concentrations (1 ×) of insecticide-impregnated papers used in WHO tube bioassays were as follows: 0.05% deltamethrin, 0.75% permethrin, and 0.1% bendiocarb. The corresponding increased concentrations used to assess resistance intensity were 5 × (0.25% deltamethrin; 3.75% permethrin) and 10 × (0.5% deltamethrin; 7.5% permethrin). During exposure, the number of mosquitoes whose expression was knocked down was recorded at 15-min intervals. The control groups (20–25 females per group) were simultaneously exposed to untreated filter papers to validate the test results. After exposure, the mosquitoes were transferred to observation tubes, provided with a sugar solution, and monitored for 24 h to determine mortality rates [24].
Molecular and biochemical analyses
Detection of kdr mutations in Ae. aegypti
Genomic DNA was extracted from 30 individual Ae. aegypti samples per commune via the 2% CTAB protocol developed by the Centre de Recherche Entomologique de Cotonou (CREC). Sterile distilled water was added to the DNA pellet for resuspension. The reconstituted DNA was then used as a template for allele-specific PCR amplification to detect target-site mutations in the voltage-gated sodium channel (vgsc) gene. PCR was carried out using the following program: an initial denaturation and hot-start enzyme activation at 95 °C for 10 min, followed by 37 cycles of 94 °C for 30 s, 60 °C for 30 s, and 72 °C for 30 s, with a final extension at 72 °C for 7 min. [25]. Four kdr mutations were analyzed: S989P, V1016G, F1534C, and V1016I. The primers used for their detection are listed in Table 1.Table 1. List of primers used for the detection of the Kdr mutation in Ae. aegyptiMutationPrimersSequences (5′–3′)S989PM1-ForAATGATATTAACAAAATTGCGCM2-RevGCACGCCTCTAATATTGATGCM1-SGCGGCGAGTGGATCGAATM1-PGCGGCGAGTGGATCGAACV1016GM2-ForGCCACCGTAGTGATAGGAAATCM2-RevCGGGTTAAGTTTCGTTTAGTAGCM2-VGTTTCCCACTCGCACAGGTM2-GGTTTCCCACTCGCACAGGGF1534CM3-ForGGAGAACTACACGTGGGAGAACM3-RevCGCCACTGAAATTGAGAATAGCM3-FGCGTGAAGAACGACCCGAM3-CGCGTGAAGAACGACCCGCV1016IIso1016fGCG GGC ACA AAT TGT TTC CCA CCC GCA CTG AVal1016fGCG GGC AGG GCG GGG GCG GGG CCA CAA ATT GTT TCC CAC CCG CAC CGG
Biochemical assays
At each study site, thirty female Ae. aegypti mosquitoes, aged 2–5 days and not exposed to any insecticide, were selected for biochemical assays. The objective of this study was to quantify and compare the activity levels of major detoxification enzymes, including mixed-function oxidases (MFOs), non-specific esterases, and glutathione-S-transferases (GSTs), across field-collected populations relative to the insecticide-susceptible Rockefeller strain. The procedures followed the protocol established by Hemingway et al. [26]. Depending on the enzymatic activity measured, the following procedures were performed:
Glutathione-S-transferase (GST) activity
For the assay of GST activity, 10 µL of mosquito homogenate (in duplicate) was dispensed into each well of a Nunc microplate. To each well, 200 µL of a reaction mixture containing reduced glutathione (GSH) and 1-chloro-2,4-dinitrobenzene (CDNB) was added. After incubation at 25 °C, absorbance was recorded kinetically at 340 nm for 5 min using a spectrophotometer.
Oxidase activity
For oxidase determination, 20 µL of mosquito homogenate (in duplicate) was added to each well, followed by 80 µL of 0.0625 M potassium phosphate buffer (pH 7.2), 200 µL of 0.25 M tetramethylbenzidine (TMBZ, pH 5.0), and 25 µL of 3% hydrogen peroxide (H₂O₂) solution. The plate was incubated for 1 h, and absorbance was measured at 630 nm using a spectrophotometer.
Non-specific esterase activity
For non-specific esterases, 10 µL of mosquito homogenate (in duplicate) was placed into each well, followed by 90 µL of 1% Triton phosphate-buffered saline (PBS, pH 6.5). After mixing, the plate was left for 10 min at 25 °C (room temperature). Then, 100 µL of a substrate solution consisting of 0.3 M α-naphthyl acetate (or β-naphthyl acetate), 2.5 mL of Triton PBS (pH 6.5), and 7 mL of distilled water was added. The plate was incubated again for 30 min, after which 100 µL of Fast Garnet Salt (FGBC) solution was added. The mixture was incubated for an additional 10 min at 25 °C (covered), and absorbance was read at 550 nm using a spectrophotometer.
Data analysis
Mortality was recorded 24 h after insecticide exposure, and the results were interpreted according to World Health Organization (WHO) guidelines [27].
- A mortality rate between 98% and 100% indicates that the tested mosquito population is fully susceptible to the insecticide.
- A mortality rate less than 98% suggests suspected resistance, which requires further investigation.
- A mortality rate less than 90% confirms the presence of resistance genes within the tested population.
For tests conducted with increased diagnostic concentrations (5 × and 10 ×), the following interpretations were applied:
- A mortality rate > 98% at a 5 × concentration indicates low resistance**.**
- A mortality rate > 98% at a 10 × concentration indicates moderate resistance.
- A mortality rate < 98% at a 10 × concentration indicates high resistance**.**
Allelic frequency estimation of kdr mutations
The frequency of kdr alleles was calculated via the basic principles of Mendelian genetics, which is based on the following formula:
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$F = \left( {2n{\text{RR}} + n{\text{RS}}} \right)/2\left( {n{\text{RR}} + n{\text{RS}} + n{\text{SS}}} \right)$$\end{document}where n represents the number of mosquitoes with each specific genotype (RR: homozygous resistant, RS: heterozygous, SS: homozygous susceptible).
This formula has been widely applied and popularized in entomological research by Martinez-Torres et al. [28].
Exact binomial tests were used to compute 95% confidence intervals for both mortality rates and kdr allelic frequencies.
To evaluate differences in enzyme activity across mosquito populations, a linear regression model coupled with ANOVA was applied. Comparisons between enzyme activity levels in field-collected populations and the reference Rockefeller strain were performed via the nonparametric Mann–Whitney U test.
Results
Diversity of culicids collected using ovitraps
The results obtained from ovitrap collections revealed a clear predominance of the genus Aedes across the three districts surveyed. Aedes aegypti was the most abundant species in Lokossa (58.55%), Bohicon (57.50%), and Dassa (62.33%), followed by Aedes albopictus, whose proportions ranged from 34.23% in Dassa to 41.36% in Bohicon. Species of the genus Culex were less represented, with Culex quinquefasciatus accounting for 1.14–3.44% and Culex nebulosus observed only in Lokossa (0.70%) (Table 2). The dominance of Aedes, as evidenced by eggs collected in ovitraps, indicates a high level of arbovirus vector activity in these urban and peri-urban areas and underscores the need to strengthen entomological surveillance and targeted vector control interventions.Table 2. Species diversity of culicids collected using ovitrapsDistrictsMosquitoes speciesLokossa % (n)Bohicon % (n)Dassa % (n)Aedes aegypti58,55 (3600)57,50 (4383)62,33 (2027)Aedes albopictus38,33 (2357)41,36 (3153)34,23(1113)Culex quinquefasciatus2,42 (149)1,14 (87)3,44 (112)Culex nebulosus0,70 (43)0,00 (0)0,00 (0)Total614976233252
Susceptibility of Ae. aegypti and Ae. albopictus to pyrethroids and carbamates
Populations of Ae. aegypti and Ae. albopictus from Dassa, Lokossa, and Bohicon were exposed to two pyrethroids (deltamethrin and permethrin) and one carbamate (bendiocarb) at three concentrations (1 × , 5 × , and 10 ×) to assess their level of resistance to these insecticides. For Ae. aegypti, all populations exhibited resistance (mortality < 90%) when exposed to the diagnostic concentration (1 ×) of deltamethrin and permethrin (Fig. 2). However, high levels of susceptibility to bendiocarb at the same concentration were observed in Dassa (98.2%), Lokossa (99%), and Bohicon (100%) (Fig. 3). When exposed to relatively high concentrations (5 × and 10 ×), mortality rates increased, ranging from 99% to 100% with permethrin, and reached 100% with both deltamethrin and bendiocarb in all the tested populations. In contrast, Ae. albopictus populations from all three locations were fully susceptible (mortality > 98%) to the diagnostic concentrations (1 ×) of all three insecticides. Specifically, the mortality rates for deltamethrin were 99.5% in Dassa, 99.7% in Lokossa, and 98.8% in Bohicon. With permethrin, mortality reached 100% in both Dassa and Lokossa and 99.8% in Bohicon (Fig. 2). Bendiocarb induced mortality rates of 98.8%, 99.9%, and 100% in Dassa, Lokossa, and Bohicon, respectively (Fig. 3). Exposure to increased concentrations (5 × and 10 ×) of deltamethrin, permethrin, and bendiocarb resulted in 100% mortality across all Ae. albopictus populations (Figs. 2 and 3).Fig. 2. Mortality rates of Ae. aegypti and Ae. albopictus exposed to permethrin and deltamethrinFig. 3Mortality rates of Ae. aegypti and Ae. albopictus exposed to bendiocarb
Frequency of kdr mutations in Ae. aegypti
Three kdr mutations were detected across all study sites, with allelic frequencies varying between localities (Table 3). For the F1534C mutation, the frequency ranged from 23.33% [8.20–38.47] in Dassa to 43.33% in Bohicon, with an overall average of 33.89% [24.11–43.67] across all locations. These results may reflect a transitional phase in which local Ae. aegypti populations shift from susceptible to resistant. With respect to the V1016G mutation, allelic frequencies varied from 28.33% [12.21–44.46] in Dassa to 56.67% [38.93–74.40] in Lokossa. For the S989P mutation, the frequency ranged from 25.00% [9.50–40.50] in Dassa to 41.67% [24.02–59.31] in Bohicon, with an overall mean of 33.89% [24.11–43.67]. Overall, the mean allelic frequencies across all sites were 33.89% [24.11–43.67] for F1534C, 46.67% [36.36–56.97] for V1016G, and 33.89% [24.11–43.67] for S989P. No individuals carrying the V1016I mutation were detected at any of the study sites.Table 3. Allelic frequencies of kdr mutations (S989P, V1016G, and F1534C) in Ae. aegypti populations from Dassa, Bohicon, and LokossaN testedRRRSSSKdr frequency95% CIF15434C Dassa301121723,338,2038,47 Lokossa301191035,0017,9352,07 Bohicon30222643,3325,6061,07 All area904533333,8924,1143,67V1016G Dassa301151428,3312,2144,46 Lokossa30426056,6738,9374,40 Bohicon30719455,0037,2072,80 All area9012601846,6736,3656,97S989P Dassa30552025,009,5040,50 Lokossa306131141,6724,0259,31 Bohicon304131335,0017,9352,07 All area9015314433,8924,1143,67
Assessment of oxidase, esterase, and GST activity in Ae aegypti
Populations of Ae. aegypti collected from Dassa, Bohicon, and Lokossa presented significantly greater mixed-function oxidase activity than did the insecticide-susceptible Rockefeller reference strain. Glutathione-S-transferase (GST) activity was detected in all field populations, with particularly elevated levels observed in Bohicon and Lokossa. In addition, increased esterase activity was recorded in the Bohicon and Lokossa populations, where both α- and β-esterases showed significantly greater activity than in the Rockefeller strain. In contrast, no statistically significant difference in α- or β-esterase expression was detected between the Dassa population and the susceptible reference strain (Fig. 4a–d).Fig. 4a Measurement of the activity of detoxification enzymes (oxidases) in Ae. aegypti. b Measurement of the activity of detoxification enzymes (GSTs) in Ae. aegypti. c Measurement of the activity of detoxification enzymes (alpha esterases) in Ae. Aegypti. d Measurement of the activity of detoxification enzymes (beta-esterases) in Ae. aegypt
Discussion
This study provides an overview of insecticide resistance in Aedes mosquitoes, the primary arbovirus vectors in Benin. It quantifies resistance levels to commonly used insecticides and explores the possible involvement of mutations in voltage-gated sodium channel genes. The results revealed that Ae. aegypti populations were resistant to the diagnostic doses (1 ×) of deltamethrin and permethrin, whereas Ae. albopictus remained fully susceptible. Both species were completely susceptible to the diagnostic dose of bendiocarb across all the sites. Furthermore, Ae. aegypti remained susceptible to increased doses (5 × and 10 ×) of deltamethrin and permethrin at all locations.
In the context of indoor residual spraying (IRS), high levels of resistance can compromise vector control efficacy, as the insecticide concentrations achieved on sprayed surfaces under field conditions are unlikely to reach those used in laboratory bioassays. Consequently, the persistence and bioavailability of pyrethroids on treated walls may be insufficient to induce lethal effects in resistant mosquito populations, potentially reducing the overall impact of IRS interventions. Furthermore, these findings underscore the need for regular monitoring of resistance intensity and the adoption of alternative or combination insecticides with different modes of action to sustain the long-term effectiveness of IRS programs [29]. Previous studies have linked such patterns to the presence of mutations in the voltage-gated sodium channel gene (commonly referred to as kdr) or to increased enzymatic detoxification mechanisms in mosquitoes [30–34]. The resistance of Ae. aegypti to pyrethroids has already been documented in Benin and other West African countries [1, 2, 25], where this species has developed increasing tolerance to pyrethroids, the most widely used class of insecticides in public health. In contrast, Ae. albopictus, more recently reported in Benin, still shows full susceptibility to pyrethroids [1, 35]. Both vectors were fully susceptible to bendiocarb, a carbamate insecticide. This difference could be explained by the distinct modes of action of the two insecticide classes: organophosphates and carbamates inhibit acetylcholinesterase, whereas pyrethroids act by disrupting the function of voltage-gated sodium channels. [36, 37]. In addition, the selective pressure for resistance to carbamates remains low, given their limited use in public health interventions (e.g., indoor residual spraying, ITNs) and relatively modest application in agriculture, where they represent only 16.6% of insecticide use in Benin [31, 38–40]. This study also reported the presence of three kdr mutations, F1534C, V1016G, and S989P, in Ae. aegypti populations. These mutations were detected at varying frequencies across sites, ranging overall from 33.89% to 46.67%. This heterogeneity suggests ongoing genetic shifts within Ae. aegypti populations, supporting the phenotypic resistance profiles observed in this study. The allelic frequencies are consistent with an early phase of resistance emergence or a transitional stage from susceptibility to resistance [2, 32]. The partial restoration of susceptibility to permethrin and deltamethrin at relatively high concentrations might be due to the relatively low kdr allele frequencies, indicating moderate resistance levels [31, 41]. These findings highlight the importance of closely monitoring the frequencies of the F1534C, V1016G, and S989P alleles to track the potential evolution of resistance before allele fixation occurs. The high levels of mixed-function oxidase (MFO) activity detected in Ae. aegypti from Dassa, Bohicon, and Lokossa further support the involvement of oxidative metabolic pathways in pyrethroid resistance, a well-documented mechanism linked to vector control failure [42, 43]. Elevated GST activity, particularly in Bohicon and Lokossa, can contribute to pyrethroid detoxification, a well-established mechanism. This observation aligns with previous reports from West Africa suggesting a growing role of GSTs in resistance to conventional insecticides [44]. Moreover, the elevated activity of α- and β-esterases observed in Ae. aegypti populations from Bohicon and Lokossa indicates enhanced metabolic activity, although it does not appear sufficient to confer phenotypic resistance to bendiocarb, to which these populations remained fully susceptible. This suggests that these esterases may play a broader detoxification role, potentially involved in cross-resistance to organophosphates rather than directly mediating carbamate resistance [39, 45]. Although increased esterase activity was observed in certain Aedes aegypti populations, this biochemical response does not necessarily confer phenotypic resistance to carbamates, such as bendiocarb. The observed susceptibility may indicate that the level or isoform of esterases expressed is inadequate to catalyze the hydrolysis of carbamate molecules efficiently, or that these enzymes are preferentially involved in the detoxification of other insecticide classes, particularly organophosphates. Alternatively, the upregulation of esterase genes might reflect a generalized metabolic adjustment, serving a supportive rather than a direct role in resistance mechanisms. These results underscore that elevated esterase activity should not be regarded as definitive evidence of carbamate resistance, but rather as part of the broader biochemical plasticity that enables mosquito populations to cope with diverse insecticidal pressures. The detection of pyrethroid resistance in Aedes aegypti, despite the moderate occurrence of kdr mutations and the variable expression of detoxification enzymes among sites, indicates that several mechanisms may interact either synergistically or independently to produce the observed resistance phenotype. In certain areas, such as Dassa, the metabolic profile suggests that oxidases or esterases play a more prominent role, whereas in other locations, resistance appears to be primarily driven by kdr-associated target-site insensitivity. This spatial variability underscores the complex and multifactorial nature of pyrethroid resistance, where both metabolic detoxification and target-site modifications may act in a compensatory manner, influenced by local selection pressures, mosquito genetic background, and the intensity of insecticide exposure. These findings underscore the urgent need for continued resistance surveillance to anticipate potential failures in vector control programs. Given the confirmed pyrethroid resistance in Ae. aegypti, alternative insecticides like bendiocarb could be considered for emergency space spraying during outbreaks, while long-term strategies should invest in non-chemical methods. This study has several limitations. First, it was conducted in only three municipalities, limiting the generalizability of its findings at the national level. Second, it was not a longitudinal study and thus did not capture resistance dynamics over time. Nevertheless, it provides a critical snapshot of the current resistance status and emphasizes the need for ongoing monitoring to prevent the spread of resistance across Aedes populations in Benin.
Conclusion
This study confirms the resistance of Ae. aegypti to the diagnostic doses of permethrin and deltamethrin in the municipalities of Lokossa, Bohicon, and Dassa-Zounmè, while Ae. albopictus remains fully susceptible. Both species were susceptible to bendiocarb. The detection of kdr mutations F1534C, V1016G, and S989P in all sites, at frequencies indicative of emerging resistance, highlights a concerning trend. In addition, detoxification enzymes such as oxidases, GSTs, and esterases (α and β) were identified in Ae. aegypti, underscoring the metabolic component of insecticide resistance in these populations. These findings provide an essential baseline for establishing continuous resistance monitoring frameworks and for guiding adaptive vector control strategies aimed at preventing the future spread of insecticide resistance in Ae. albopictus populations.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1PNLP. Epidemie de dengue, Abidjan; 2023. https://www.pnlpcotedivoire.org/article-detail/8/158/epidemie-de-dengue-abidjan
- 2WHO. Dengue. World Health Organization; 2025. https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue
- 3WHO. Dengue-Global situation; 2024. https://www.who.int/emergencies/disease-outbreak-news/item/04-August-2017-dengue-cote-d-ivoire-en
- 4Gerome P. Evolution des épidémies de dengue en Afrique depuis le début novembre 2024. Mes Vaccins. 2024. https://www.mesvaccins.net/web/news/22570-evolution-des-epidemies-de-dengue-en-afrique-depuis-le-debut-novembre-2024
- 5WHO. Dengue and severe dengue; 2024. https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue
- 6Ledoux S, Gutierrez CT, Lobo NF, Murillo EM, Pérez S, Guerra R, et al. The need to harmonize insecticide resistance testing: methodology, intensity concentrations and molecular mechanisms evaluated in Aedes aegypti populations in Central America and Hispaniola. bio Rxiv. 2020; p. 2020.02.25.964270. 10.1101/2020.02.25.964270
- 7OMS. Manuel pour le suivi de la résistance aux insecticides chez les moustiques vecteurs et la sélection d’interventions appropriées; 2022. https://www.who.int/fr/publications-detail/9789240051089
- 8Direction de la Statistique Agricole (DSA). Les chiffres définitifs de la campagne agricole 2023–2024 au Bénin. MAEP. Cotonou, Bénin; 2024. https://dsa.agriculture.gouv.bj