Microbiome characterization of the sea slugs Elysia viridis and Placida dendritica: insights into potential roles in kleptoplasty
Patrícia Martins, Paulo Cartaxana, Sónia Cruz

TL;DR
This study compares the microbiomes of two sea slugs that retain chloroplasts, finding significant differences that may relate to their ability to maintain functional chloroplasts.
Contribution
The study identifies a unique carotenoid-producing bacterial genus in Elysia viridis, potentially linked to chloroplast retention.
Findings
E. viridis has a smaller, more specialized microbiome dominated by Bacteroidota and Actinomycetota.
A ZOTU related to Fulvibacter was found exclusively in E. viridis, suggesting a role in chloroplast retention.
P. dendritica has a more diverse microbiome dominated by Pseudomonadota.
Abstract
Kleptoplasty is the process by which functional chloroplasts from algae food sources are sequestered and retained by a host organism. Some sacoglossan sea slugs display this ability, enabling them to survive extended periods of food shortage, as they can obtain organic compounds from photosynthesis. While research has focused on the mechanisms underlying chloroplast retention and functionality, the contribution of the microbiome to kleptoplasty in these photosynthetic sea slugs is largely unexplored. In this study, we assessed the bacterial communities of Elysia viridis and Placida dendritica, two photosynthetic sacoglossan species that share the same habitat and macroalga food source, but exhibit distinct abilities to retain chloroplasts. High-throughput 16S rRNA gene sequencing revealed highly significant differences in bacterial community composition between E. viridis and P.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
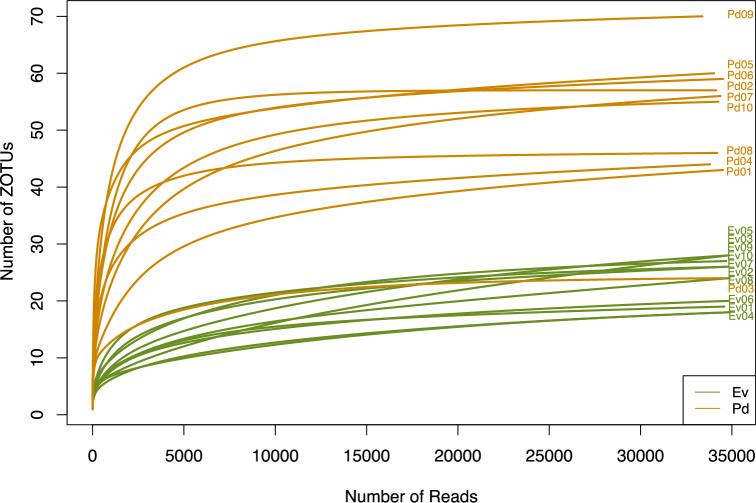 Figure 1
Figure 1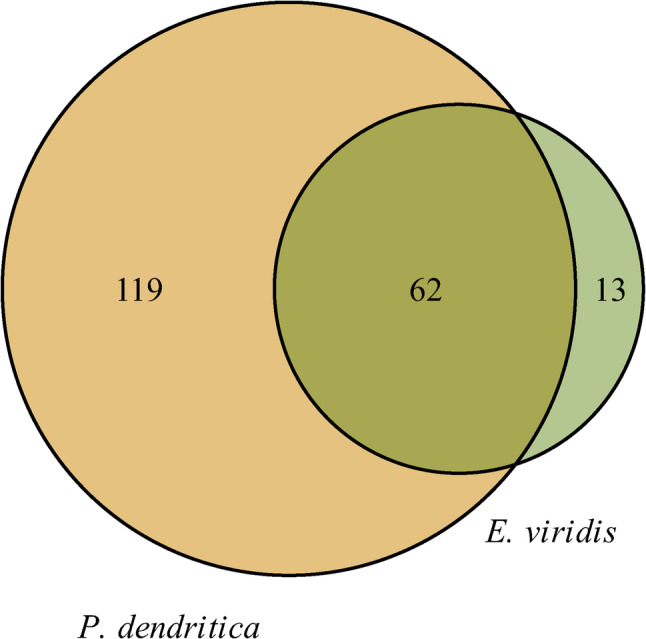 Figure 2
Figure 2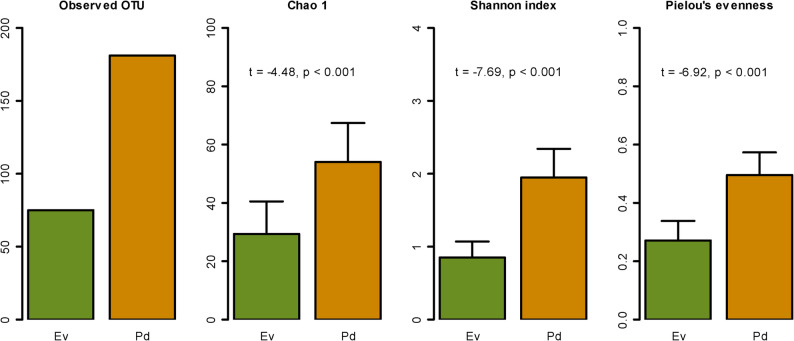 Figure 3
Figure 3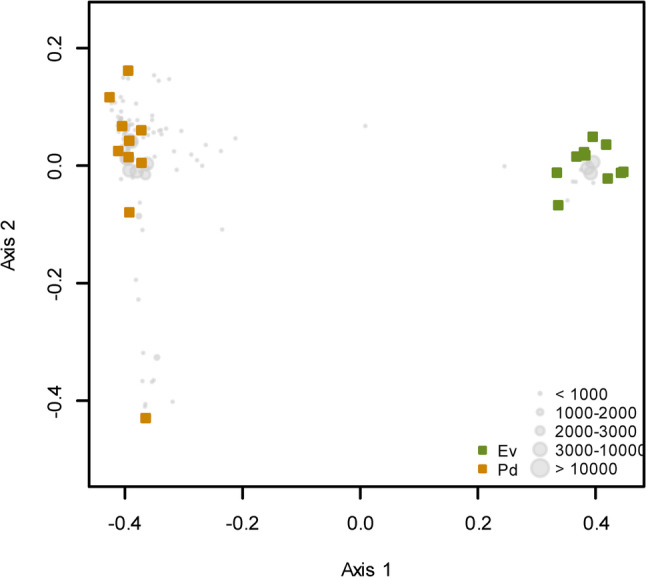 Figure 4
Figure 4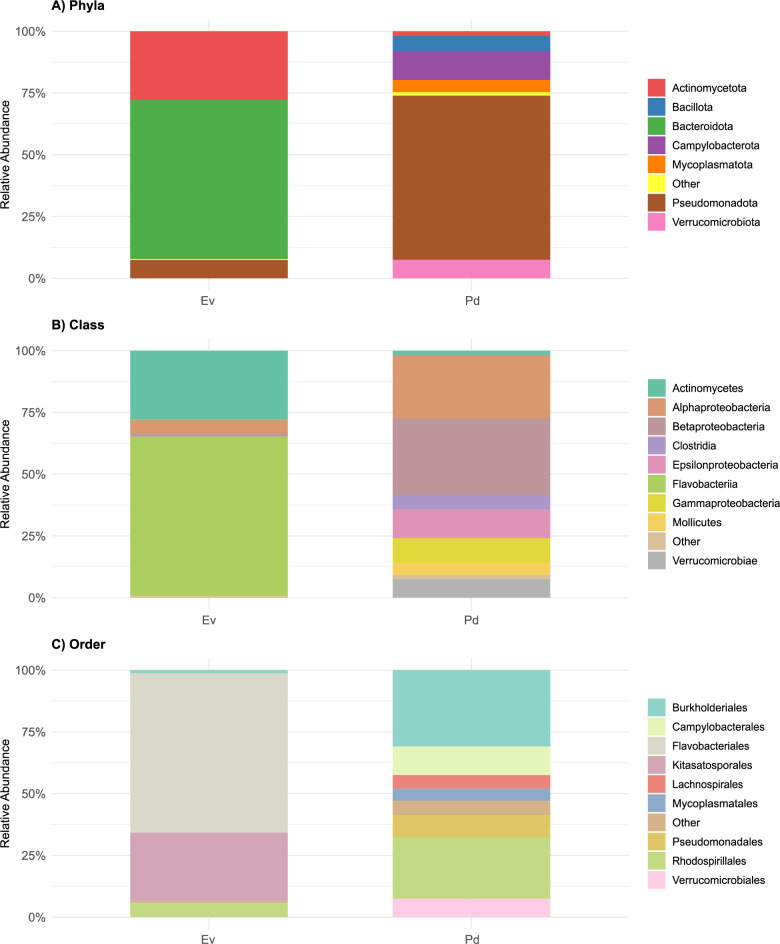 Figure 5
Figure 5- —https://doi.org/10.13039/501100000781European Research Council
- —https://doi.org/10.13039/501100001871Fundação para a Ciência e a Tecnologia
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine Biology and Ecology Research · Marine and coastal plant biology · Marine Invertebrate Physiology and Ecology
Background
Some Sacoglossa sea slugs (Gastropoda, Mollusca) are characterized by their remarkable ability to sequester chloroplasts from algae [1]. The stolen chloroplasts – kleptoplasts – are maintained functionally and capable of harnessing the energy of sunlight, converting it through photosynthesis into usable resources [2]. This capacity, named kleptoplasty, enables these sea slugs to survive extended periods of food shortage, as they can supplement their diet with energy derived from photosynthesis [3, 4]. Studies have revealed that photosynthetic sea slugs may exhibit a wide range of kleptoplast retention times [5, 6]. Some species, like Placida dendritica, show non-functional retention, kleptoplasts are rapidly degraded and no photosynthetic activity is detected even immediately after chloroplast acquisition [7]. Others, like Elysia viridis, exhibits long-term retention of functional chloroplasts, with photosynthetic activity lasting for some weeks to a few months [8]. Among metazoans, the capacity for long-term maintenance of photosynthetically active kleptoplasts is exclusive to some Sacoglossa, mostly within the genus Elysia [9].
Extensive research has been conducted on these photosynthetic sacoglossan sea slugs, particularly focusing on the mechanisms underlying chloroplast sequestration and maintenance. Different sacoglossan species have been found to preferentially feed on and retain functional chloroplasts from specific types of algae, while other species have a more generalist diet, retaining functional chloroplasts from a variety of algae species [6]. The long-term retention species E. viridis failed to retain functional chloroplasts when fed on the macroalga Cladophora rupestris [10], which indicates a level of selectivity in kleptoplast retention. Therefore, the retention of functional chloroplasts cannot be solely attributed to the alga or sacoglossan individually, depending on a combination of specific characteristics from both species involved [11]. An important and conspicuous factor that remains overlooked in this host-donor relationship is the role of the associated bacteria.
The concept of the “holobiont” has gained increasing recognition, emphasizing the complex and dynamic interactions between a host and its microbiome [12, 13]. Growing awareness of the microbiome’s potential influence on host physiology, metabolism, and adaptation has underscored its fundamental role in shaping the biology of diverse organisms [14]. It has become increasingly evident that many compounds once attributed solely to animal physiology are, in fact, produced by bacterial symbionts [15–17]. Furthermore, symbiotic bacteria contribute to the overall health, metabolism, and ecological interactions of their hosts [18, 19]. Several studies have already demonstrated the importance of microbial communities, and especially bacteria, in maintaining the functionality of algal symbionts in corals [20, 21].
Advances in high-throughput sequencing technologies have revolutionized our ability to understand the complexities of bacterial communities, allowing us to explore their diversity, functional potential, and ecological interactions with unprecedented depth and accuracy [22–24]. However, only few studies have addressed the bacterial communities associated with photosynthetic sacoglossan sea slugs. Devine et al. [25] reported changes in the bacterial diversity associated with two populations of the long-term kleptoplastic sea slug Elysia chlorotica, which also differed from its algal prey Vaucheria litorea. Two additional studies assessed the bacterial communities of the sacoglossan sea slugs Elysia rufescens and Elysia crispata [26, 27].
In this study, we characterized the microbial communities of E. viridis and P. dendritica inhabiting intertidal and shallow coastal waters of the northeastern Atlantic, where they are commonly found on the green macroalga Codium tomentosum, their primary food source [28]. Despite sharing the same habitat and algal food source, E. viridis can retain functional chloroplasts for up to 12 weeks [29], whereas P. dendritica exhibits non-functional chloroplast retention [7]. With this study, we aim to characterize and compare the microbiomes of E. viridis and P. dendritica using high-throughput sequencing of the 16S rRNA gene. Identification of key bacterial taxa may elucidate their relevance and potential role in the process of kleptoplasty.
Materials and methods
Sea slug collection and maintenance
The sacoglossan species Elysia viridis (Montagu, 1804) and Placida dendritica (Alder & Hancock, 1843) (Supplementary fig. S1), along with their food source, the macroalga Codium tomentosum (Stackhouse, 1797), were collected during low tide from the intertidal rocky zone of Tamargueira Beach in Figueira da Foz, Portugal (40° 9’ 58.730” N; 8° 52’ 57.285” W) in mid-September 2024. The animals and algae were maintained under controlled laboratory conditions for three weeks to ensure environmental uniformity prior to sampling. The animals were housed in 150 L recirculating life support systems (LSS) filled with artificial seawater (ASW) at a salinity of 35 PSU and a temperature of 18 °C. The two species, E. viridis and P. dendritica, were kept in separate maternity nets within the system and regularly fed with C. tomentosum. The LSS was operated on a 12 h light:12 h dark photoperiod and equipped with T5 fluorescent lamps, providing a photon scalar irradiance of 60–80 µmol photons m⁻² s⁻¹. Light intensity was measured at the water surface by using a Spherical Micro Quantum Sensor connected to a ULM-500 Universal Light Meter (Walz, Germany). Water changes (approximately 10% of the system’s volume) were performed weekly using freshly prepared ASW, with the same salinity. Additionally, fish maternity nets were replaced once a week.
Sampling and DNA extraction
After three weeks, the reared animals were collected and washed with autoclaved and filtered ASW to remove transient and loosely attached bacteria. They were then transferred to a disinfected 200 μm pore-sized sieve and thoroughly rinsed with autoclaved and filtered ASW to eliminate debris and excess mucus. The sieve was subsequently placed on tissue paper to remove residual water. Samples were snap-frozen in liquid nitrogen and stored at −20 °C, as DNA extraction was performed within two days of collection.
For E. viridis, 10 replicates were prepared, with each replicate consisting of a pool of five individuals. In contrast, for P. dendritica, 10 replicates were also used, but each replicate contained a pool of seven individuals due to species-specific differences in body size. Individuals used ranged from 9 to 15 mm in length for E. viridis and 4 to 7 mm for P. dendritica. Total community DNA (TC) was extracted from whole animal samples of E. viridis (10 replicates) and P. dendritica samples (10 replicates) using the FastDNA^®^ SPIN Kit (MP biomedicals) following manufacturer’s instructions.
Next-generation sequencing analysis
The V4 hypervariable region of the 16S rRNA gene amplicons were amplified using PCR primers 515 F (5’-GTGCCAGCMGCCGCGGTAA-3’) and 806R (5’-GGACTACHVHHHTWTCTAAT-3’) [30] with barcode on the forward primer. A 30 cycle PCR assay using the HotStarTaq Plus Master Mix Kit (Qiagen, USA) was performed under the following conditions: 95 °C for 5 min, followed by 30 cycles of 95 °C for 30 s, 53 °C for 40 s and 72 °C for 1 min, and a final elongation step at 72 °C for 10 min. After amplification, PCR products were checked in 2% agarose gel to determine the success of amplification and the relative intensity of bands.
Multiple samples were pooled together in equal proportions based on their molecular weight and DNA concentrations. Pooled samples were purified using calibrated Ampure XP beads, with pooled samples and purified PCR product being used to prepare an Illumina DNA library. Sequencing was performed at MR DNA (www.mrdnalab.com, Shallowater, TX, USA) using Illumina MiSeq platform with paired-end 2 × 250 bp reads, following the manufacturer’s instructions. The average sequencing depth was approximately 20,000 reads per sample. Sequence data were processed using MR DNA analysis pipeline (MR DNA, Shallowater, TX, USA). Briefly, sequences were joined, depleted of barcodes and then sequences < 150 bp or with ambiguous base calls were removed. Sequences were quality filtered using a maximum expected error threshold of 1.0 and dereplicated. After denoising and chimera filtering, zero-radius operational taxonomic units (ZOTUs) were generated. Taxonomic classification was then performed using BLASTn against an internally curated 16S rRNA database (MR DNA version 2024.4) derived from NCBI sequences (https://ftp.ncbi.nlm.nih.gov/blast/db/), as implemented by MR DNA. The database undergoes standard curation and quality-control procedures, including the removal of duplicate, partial, ambiguously annotated, and improperly annotated sequences, and is updated quarterly (versioned by year–quarter). Although the database is proprietary and not publicly available, which limits full reproducibility, MR DNA’s 16S rRNA sequencing and classification pipeline has been widely applied in peer-reviewed microbiome studies, supporting the reliability and consistency of the results obtained.
Data analyses
An OTU table containing the raw abundance of all ZOTUs of Elysia viridis samples (Ev) and Placida dendritica samples (Pd) was imported to R version 4.2.3 (http://www.r-project.org/). To assess sequencing effort, rarefaction curves were generated using the rarecurve() function from the vegan package (version 2.6–4.6) in R.
To determine the distribution of ZOTUs in E. viridis and P. dendritica samples, a Venn diagram was assembled using the venn() function in the gplots package (version 3.1.3).
Alpha bacterial diversity was assessed by calculating the ZOTUs recorded, Chao1 index, Shannon diversity index and Pielou’s evenness index. All indices were calculated with the diversity() function from vegan package. Differences in bacterial alpha-diversity were assessed using a t-test after confirming normality using Shapiro’s test and homogeneity of variance using the Levene’s test.
All subsequent community composition analyses were performed using relative abundance data, obtained through proportional normalization of read counts within each sample. Variation in ZOTU composition was visualized using Principal Coordinates Analysis (PCoA) based on Bray-Curtis dissimilarity, calculated from relative abundance data with vegdist() function in the vegan package. Ordination plots were generated using the cmdscale() function in R. Differences in the bacterial composition of the two sacoglossan sea slugs were tested using PERMANOVA with the adonis2() function of the vegan package.
For visualization of the representative taxa (Phylum, Class, and Order) in E. viridis and P. dendritica, stacked bar plots were constructed using the ggplot2 package (version 3.4.4). Prior to plotting, read counts were aggregated at each taxonomic level, normalized to relative abundance (%) within each sample, and taxa contributing less than 1% to the total community were grouped as “Other”.
ZOTUs with more than 3,000 sequence reads were selected to identify the most dominant bacterial communities, and their closest relatives were determined through BLAST search.
The core microbiome of both sacoglossan species was assessed by selecting the ZOTUs that were present simultaneously in all individuals. All DNA sequences generated in this study were submitted to the NCBI SRA: Accession number PRJNA1256232. The R script and input data used for microbiome analysis and figure generation are available via GitHub at: https://github.com/Patriciatmartins/Elysia-viridis-and-Placida-dendritica-Microbiome_.
Pigment analysis
Pigment extraction and analysis were performed as described by Cruz et al. [31]. Briefly, freeze-dried sea slug and macroalgal samples were extracted for 30 min in 95% cold buffered methanol (2% ammonium acetate), after sonication for 1 min. Extracts were filtered through 0.2 μm PTFE membrane filters before injection into a Prominence-i LC 2030 C HPLC system (Shimadzu, Kyoto, Japan) equipped with a photodiode array detector. Chromatographic separation was carried out using a Supelcosil C18 reverse-phase column (250 × 4.6 mm, 5 μm particle size; Sigma-Aldrich, St. Louis, MO, USA) under a 35 min elution gradient. Pigments were identified based on absorbance spectra and retention times, in comparison with pure pigment standards from DHI (Hørsholm, Denmark).
Results and discussion
Overall assessment of bacterial composition
After quality filtering, 689,554 sequence reads were detected and assigned to 194 bacterial ZOTUs. Elysia viridis samples displayed 347,778 sequence reads, while Placida dendritica presented 341,776 sequence reads (Supplementary table S1).
Rarefaction curves illustrate the relationship between sequencing depth (number of sequence reads) and the observed ZOTU richness for each sample (Fig. 1). The asymptotic nature of the curves suggests that sequencing effort was sufficient to capture the bacterial diversity within each sample. Sequence reads were evenly distributed across samples, minimizing biases and supporting robust comparisons between groups. These findings indicate that the dataset provides adequate coverage for assessing bacterial community composition and richness in the studied populations of E. viridis and P. dendritica.Fig. 1. Rarefaction curves illustrating the bacterial diversity and species richness in Elysia viridis (Ev) and Placida dendritica (Pd) samples
The distribution of shared and unique ZOTUs between the samples of E. viridis and P. dendritica is represented in the Venn diagram (Fig. 2). The sea slug P. dendritica contained the highest number of unique ZOTUs, with 119 exclusive to this species, while E. viridis samples exhibited only 13 unique ZOTUs. Both species shared a total of 62 ZOTUs, highlighting a core set of microbial taxa present in both organisms. However, if we consider only ZOTUs present in at least 5 replicates, E. viridis exhibited only 6 unique ZOTUs, while P. dendritica exhibited 13 unique ZOTUs. This reduction in the number of unique ZOTUs could be attributed to transient or low-abundance microbial taxa that were not consistently detected across replicates. These results suggest that, in the Portuguese populations examined, E. viridis harbors a smaller yet more specialized bacterial community, whereas P. dendritica supports a larger, more diverse, and generalist microbiome.Fig. 2. Venn diagram showing the number of shared and unique bacterial ZOTUs in Elysia viridis and Placida dendritica
In terms of bacterial community diversity, results indicate higher alpha diversity in P. dendritica samples, as assessed by the Chao1 index, Shannon diversity index, and Pielou’s evenness (Fig. 3). There were statistically significant differences between the bacterial communities associated with P. dendritica and E. viridis, with P. dendritica exhibiting higher values across all diversity indices (Chao1: t = −4.48, p < 0.001; Shannon: t = −7.69, p < 0.001; and Pielou’s evenness: t = −6.92, p < 0.001). These findings suggest that P. dendritica microbiome is not only more diverse but also characterized by a well-balanced distribution of bacterial species.Fig. 3. Alpha diversity metrics of bacterial communities associated with Elysia viridis (Ev) and Placida dendritica (Pd). The graphs display the number of observed ZOTUs, Chao1 richness estimator, Shannon diversity index, and Pielou’s evenness index. Statistical comparisons were performed to assess differences between groups
The PCoA ordination analysis illustrates a clear separation between two main groups, corresponding to E. viridis samples and P. dendritica samples (Fig. 4). The primary axis of variation, which explains 66.7% of the variance, highlights the distinct differences in the bacterial communities associated with each species. Statistical analysis revealed highly significant differences in bacterial community composition between E. viridis and P. dendritica samples (F = 81.058; p = 0.001).Fig. 4. Ordination showing the first two axes of the principal coordinates analysis (PCoA) of the bacterial composition present in Elysia viridis (Ev) and Placida dendritica (Pd). Grey circles represent ZOTUs, with their size proportional to abundance (number of sequence reads)
Taxonomic composition of bacterial communities
The overall taxonomic analysis grouped bacterial sequences into fifteen phyla, twenty-nine classes, and fifty-six orders. Fig. 5 highlights the relative abundance of the most representative bacterial groups found in E. viridis and P. dendritica. Bacteroidota and Actinomycetota were the most dominant phyla in E. viridis, together accounting for approximately 92% of the total sequence reads. In contrast, P. dendritica harbored thirteen phyla, with Pseudomonadota (former Proteobacteria) being the most prevalent, representing 66% of the total sequence reads (Fig. 5A).Fig. 5. Stacked bar plots showing the relative abundance (%) of representative bacterial groups in Elysia viridis (Ev) and Placida dendritica (Pd), for phyla (A), class (B), and order (C). Taxa contributing less than 1% to the overall abundance are grouped as “Other”. Bars represent the mean relative abundance (%) based on proportional normalization of read counts within each sample
Pseudomonadota, Bacteroidota and Actinomycetota are known to dominate the microbial communities in various marine habitats, including in marine invertebrates, such as sea snails, coral and sponges where they contribute to digestion, nutrient cycling, and symbiotic interactions [23, 25, 32].
In a previous study assessing the bacterial communities of the sacoglossan sea slug E. crispata, Spirochaetota and Bacteroidota were reported as the dominant phyla, although Mycoplasmatota and Pseudomonadota were also detected among other taxa [27]. Similarly, a study investigating the bacterial communities of two wild E. chlorotica populations (from near Nova Scotia and Martha’s Vineyard, USA) found a clear dominance of the phylum Pseudomonadota [25]. The predominance of the phylum Pseudomonadota observed in P. dendritica, was also reported in the microbiome of its main food source, the macroalga C. tomentosum, in a study examining its microbial community under natural conditions and in response to copper exposure [33].
More than 64% of the bacterial sequences identified in E. viridis samples were identified as belonging to the class Flavobacteriia, while in P. dendritica more than 50% of the bacterial sequences were identified as belonging to the classes Betaproteobacteria (31%) and Alphaproteobacteria (25%) (Fig. 5B). Previous studies addressing the host-associated microbiome of the sea slug E. chlorotica, identified Betaproteobacteria and Alphaproteobacteria as dominant groups [25]. However, notable differences in the relative abundance of Betaproteobacteria were observed between wild populations from Nova Scotia and Martha’s Vineyard, with higher proportions in individuals inhabiting environments that are subject to large salinity fluctuations [25]. These bacterial classes were also among the three most dominant detected in the C. tomentosum microbiome, with Gammaproteobacteria being the most abundant (57%), followed by Alphaproteobacteria (31%) and Betaproteobacteria, which accounted for 4.8% [33].
At the order level, 64% of E. viridis sequences were grouped under Flavobacteriales, with all Flavobacteriales-derived sequences being assigned to the Flavobacteriaceae family (Fig. 5C). Notably, these dominant families found in E. viridis were absent in P. dendritica. Members of the Flavobacteriales order are recognized for their involvement in the degradation of complex organic matter [34] and play a crucial role in the functioning of marine biofilms [35]. Although Flavobacteriales have not been identified as dominant orders in other studies of sea slug microbiomes, they have been observed in both photosynthetic [25] and non-photosynthetic sea slug species [36]. In P. dendritica, Burkholderiales (30%) and Rhodospirillales (24%) were the most abundant bacterial orders, accounting for 54% of the total sequences (Fig. 5C). A previous study by Devine et al. [25] investigating the bacterial communities associated with two populations of E. chlorotica also reported that Betaproteobacteria in sea slug samples were predominantly represented by the order Burkholderiales. These findings suggest that the order-level trends observed in the Portuguese populations of E. viridis and P. dendritica are consistent with those already reported for other sacoglossan sea slugs, with certain bacterial groups playing a central role in their microbial communities.
Composition analysis of dominant ZOTUs
The most dominant ZOTUs (≥ 3,000 sequence reads) identified in this study are summarized in Table 1. The dominance analysis revealed four dominant ZOTUs found in E. viridis: ZOTU 1 (Fulvibacter), ZOTU 2 (unknown Actinomycetota), ZOTU 3 (Ralstonia syzygii) and ZOTU 8 (Aestuariispira) (Table 1). ZOTU 1 was the most abundant ZOTU detected, accounting for 224,282 sequence reads, and found exclusively in E. viridis samples (Table 1). According to Blast (http://www.ncbi.nlm.nih.gov/) this ZOTU was related Fulvibacter tottoriensis [GenBank accession number (acc.) NR041572] isolated from a marine sediment in Japan [37]. An important characteristic of Fulvibacter genus, as observed in many members of the Flavobacteriaceae family, is the production of carotenoid-type pigments [37]. Carotenoids are well known for their antioxidant properties, mitigating oxidative damage caused by reactive oxygen species (ROS) [38]. In marine environments, these pigments also play a crucial role in photoprotection from harmful ultraviolet (UV) radiation and intense light exposure [39, 40]. Additionally, carotenoids have been investigated for their role in enhancing stress tolerance in symbiotic and free-living marine organisms [41]. For example, Motone et al. [21] reported the discovery of a zeaxanthin-producing bacterium (belonging to Flavobacteriaceae family) with the ability to protect Symbiodiniaceae ‒ the algal symbionts of corals ‒ from thermal and light stress, potentially contributing to coral resilience under climate change conditions.Table 1. Taxonomic affiliation of the most abundant ZOTUs in Elysia viridis (Ev) and Placida dendritica (Pd) (≥ 3,000 sequences) including ZOTU-numbers (ZOTU); number of sequence reads; taxonomic assignment (Phylum, Class and Order); their closest relatives (Blast result); with respective accession number (Accession); sequence identity (Sq ident); and their source. Values represent total (absolute) sequence read counts. All comparative and statistical analyses were performed using relative abundance data normalized by sample read depthZOTUSequence readsSumPhylumClassOrderBlast resultAccessionSq. ident (%)SourceEvPdZOTU1224,2820224,282BacteroidotaFlavobacteriiaFlavobacterialesFulvibacter tottoriensisNR 114,16993.94marine sedimentZOTU296,145096,145ActinomycetotaActinomycetesKitasatosporalesStreptomyces sp.DQ84682182.49European soilsZOTU34,34897,115101,463PseudomonadotaBetaproteobacteriaBurkholderialesRalstonia insidiosaMW960229100.00Ralstonia insidiosa strainZOTU4084,96184,961PseudomonadotaAlphaproteobacteriaunknownunknown AlphaproteobacteriaHQ67553281.82South Atlantic OceanZOTU5039,72939,729CampylobacterotaEpsilonproteobacteriaCampylobacteralesMalaciobacter marinusCP03210193.28Pacific OceanZOTU6030,70530,705PseudomonadotaGammaproteobacteriaPseudomonadalesPseudomonas sp.MN22720991.70Pseudomonas sp. strainZOTU7025,44725,447VerrucomicrobiotaVerrucomicrobiaeVerrucomicrobialesRubritalea marinaNR_04370194.86marine spongeZOTU819,868019,868PseudomonadotaAlphaproteobacteriaRhodospirillalesAestuariispira insulaePP05312789.37Aestuariispira insulae strainZOTU9016,57316,573MycoplasmatotaMollicutesMycoplasmatalesCandidatus Mycoplasma mahoniiOP99547495.63brittle starZOTU10013,24513,245Pseudomonadotaunknownunknownunknown ProteobacteriaLN68128493.68marine algaZOTU1105,2195,219Pseudomonadotaunknownunknownunknown ProteobacteriaLN68128493.28marine algaZOTU121293,3123,441PseudomonadotaBetaproteobacteriaBurkholderialesRalstonia pickettiiOQ062259100.00rice root
Previous studies revealed the presence of an unidentified carotenoid in the sea slug E. viridis that was not present in their food source, the macroalgae C. tomentosum and Chaetomorpha sp [31, 42]. The authors hypothesized that the compound was synthesized by the animal cell using an algal precursor. However, the high abundance and exclusive presence of ZOTU 1 in E. viridis suggest a possible bacterial origin for the unidentified carotenoid. To obtain additional insights, we compared the pigment composition of the two sea slug species and the macroalgal food source (Supplementary fig. S2). The pigment profile of P. dendritica matched that of C. tomentosum, with the presence of chlorophylls a and b, and the carotenoids siphonaxanthin, all-trans-neoxanthin, 9’-cis-neoxanthin, violaxanthin, siphonaxanthin dodecenoate, and β,ε-carotene (Supplementary figs. S2B and C). Results confirm that E. viridis possesses an additional unidentified carotenoid absorbing in the 400–550 nm range and with an absorption maximum of 460 nm (Supplementary figs. S2A and D). Further investigations are required to elucidate its origin, but it is plausible that a carotenoid produced by the associated bacteria is accumulated by this long-term retention sea slug, helping the animal to cope with oxidative stress generated by the acquired chloroplasts. By mitigating ROS accumulation, this carotenoid could enhance the stability and functionality of kleptoplasts, ultimately benefiting the host organism. While our results highlight a correlation between the dominance of ZOTU 1, a putative carotenoid-producing bacterium, and the long-term retention of functional chloroplasts in E. viridis, this association remains speculative and does not establish a direct functional link. Because our analysis is based on 16S rRNA amplicon sequencing, which has limited taxonomic and functional resolution, future studies integrating metagenomic, transcriptomic, and metabolomic approaches, together with experimental manipulation of bacterial communities, will be essential to clarify the specific contributions of the microbiome to chloroplast stability and photoprotection in photosynthetic sea slugs.
ZOTU 2 was classified as an unknown Actinomycetota and was detected exclusively in the E. viridis. Blast (http://www.ncbi.nlm.nih.gov/) results related this ZOTU to the genus Streptomyces, however with a relatively low sequence similarity (< 85%) with organisms from the NCBI database (Table 1). The phylum Actinomycetota is one of the most important bacterial groups for the discovery of novel natural products [43]. ZOTU 8 was also dominant in E. viridis and was taxonomically assigned to the family Kiloniellaceae. BLAST analysis revealed that this ZOTU is closely related to Aestuariispira insulae, although with a sequence homology of less than 90% compared to known references (Table 1). Members of the Kiloniellaceae family have previously been identified in high abundances in various marine invertebrates, including sponges [44], jellyfish [45] and corals [46], as well as in algae [47].
The most abundant ZOTUs found in P. dendritica were ZOTU 3 and 12 (Ralstonia), ZOTU 4 (unknown Alphaproteobacteria), ZOTU 5 (Malaciobacter), ZOTU 6 (Pseudomonas), ZOTU 7 (Rubritalea), ZOTU 9 (Mycoplasma), ZOTU 10 and 11 (unknown Proteobacteria) (Table 1). ZOTUs 3 and 12, unlike the other dominant ZOTUs, were detected in both E. viridis and P. dendritica, however with a significant higher number of sequence reads in P. dendritica samples. Blast results related these ZOTU to Ralstonia insidiosa and Ralstonia pickettii (former Pseudomonas pickettii), respectively (Table 1). While both species are primarily recognized as opportunistic pathogens associated with plant and human diseases [48, 49], they are also known for their capacity to form robust biofilms [50]. Previous studies have demonstrated that members of the genus Ralstonia produce signaling molecules involved in quorum sensing and biofilm formation [51], although the extent and mechanisms may vary among species. Additionally, they exhibit exceptional metabolic versatility, enabling them to use a wide range of carbon sources, including aromatic compounds [52]. This metabolic flexibility allows the bacterium to thrive in a variety of environmental conditions, further contributing to its persistence in diverse habitats.
According to BLAST analysis, ZOTU 5 was identified as Malaciobacter marinus (formerly Arcobacter marinus) (Table 1). Although the ecological role of Malaciobacter in the marine environment remains unclear, it has been reported as a prevalent and abundant bacterial species in the microbiomes of oysters (Tiostrea chilensis) and corals (Acropora species) [53, 54]. Additionally, members of this genus have been detected in the skin and gill mucus of various fish species [55].
Blast results related ZOTU 6 to the genus Pseudomonas, a group of bacteria that are ubiquitous in marine environments. However, the sequence similarity was relatively low (91%) when compared to organisms in the NCBI database (Table 1). Pseudomonadaceae family plays a crucial role in the nitrogen and carbon cycles by fixing nitrogen, decomposing organic carbon, and converting nitrogenous compounds, thereby contributing to ecosystem balance [56, 57]. Additionally, several members of this family are known for producing antimicrobial compounds, including antibiotics and bacteriocins. Some Pseudomonas species also establish symbiotic relationships with marine organisms, such as fish and corals, supporting host health by promoting nutrient cycling and acting as biocontrol agents against harmful pathogens [58–60]. This family has previously been identified in the microbiome of sea slugs [35] including photosynthetic species [25].
BLAST analysis identified ZOTU 7 as Rubritalea marina, which was exclusively detected in P. dendritica, accounting for 25,447 sequence reads (Table 1). This species was originally isolated from the Mediterranean sponge Axinella polypoides [61]. The prevalence of Rubritalea species in marine sponges suggests their potential role in symbiotic interactions with marine organisms. Members of this genus are known for their ability to produce carotenoids and degrade organic matter, contributing to both carbon and nitrogen cycling [62].
ZOTU 9, detected exclusively in P. dendritica, was assigned to Candidatus Mycoplasma mahonii based on a BLAST search. Although generally regarded as parasitic, Mycoplasma species are frequently found in symbiotic associations with marine organisms, including fish and corals [63, 64]. They are integral members of the normal microbiota of these hosts, colonizing mucosal surfaces and the digestive tract [64, 65]. Their versatility makes them well-suited for host-associated niches such as fish gills, intestines, and coral mucus layers. As facultative anaerobes or microaerophiles, Mycoplasma species can thrive in oxygen-limited conditions, further supporting their adaptation to these environments [66]. Members of the genus Mycoplasma have been already detected in photosynthetic sea slugs, including Elysia crispata [67] and Elysia rufescens [26]. Although the microbiomes of P. dendritica, E. crispata, and E. rufescens share the presence of specific genera such as Pseudomonas and Mycoplasma, their overall bacterial communities exhibit significant differences at the genus level. This is not unexpected, considering the differences in their environments, feeding strategies, and dietary preferences.
It is important to highlight that, among all the most abundant ZOTUs detected in the studied E. viridis and P. dendritica populations, only two (ZOTU 3 and 12) showed 100% sequence homology in the BLAST search. Six ZOTUs exhibited homology between 93% and 95% (ZOTU 1, 5, 7, 9, 10 and 11), two ZOTUs demonstrated homology ranging from 88% to 92% (ZOTU 6 and 8), while the remaining two displayed homology below 85% (ZOTU 2 and 4) in comparison to published sequences (Table 1). These findings suggest that the taxonomic assignment of several abundant ZOTUs remains uncertain at the species level, highlighting potential novel or highly divergent bacterial lineages associated with E. viridis and P. dendritica. The marine environment represents a vast and largely unexplored reservoir of biodiversity, with significant potential for the discovery of novel species. In recent years, numerous previously unidentified taxa have been described, including bacterial species associated with sea slugs, highlighting the complexity and ecological significance of host-associated microbiomes [15, 68]. The relatively low sequence homology observed for several dominant ZOTUs in this study suggests the potential presence of yet-undiscovered bacterial species, underscoring the uniqueness of these underexplored microbial communities. Future research should focus on the detailed characterization of these specific bacterial taxa to better understand their functional roles and evolutionary significance for photosynthetic sea slugs, including their putative involvement in kleptoplast establishment and maintenance. At the same time, it should be acknowledged that the present study, conducted without temporal replication and limited to a single sampling location, represents a snapshot from the Portuguese coast that may partly reflect local environmental influences. Expanding spatial and temporal sampling will be necessary to evaluate the consistency of these microbial associations across different habitats and populations.
Core microbiome characterization
The core microbiome is defined as the set of ZOTUs that are consistently present across all replicates. In E. viridis, the core microbiome comprised eight ZOTUs out of the 75 detected. In contrast, P. dendritica exhibited a core microbiome comprising nine ZOTUs out of the 181 identified (Supplementary table S2). The core microbiome of E. viridis was composed of five phyla, seven classes, seven orders, and seven families, while that of P. dendritica comprised six phyla, eight classes, eight orders, and eight families. Bacteroidota was the dominant phylum in the core microbiome of E. viridis, accounting for approximately 64% of total sequence reads. In contrast, Pseudomonadota dominated the core microbiome of P. dendritica, representing about 68%. At the family level, Flavobacteriaceae (64%) was the most abundant in E. viridis, whereas P. dendritica was dominated by Burkholderiaceae (26%) and Rhodospirillaceae (31%).
The taxonomic composition of the core microbiome of E. viridis and P. dendritica mirrors that of the most dominant ZOTUs identified in each species, because all the dominant ZOTUs identified in E. viridis and P. dendritica are part of the species core microbiome. All nine ZOTUs constituting the core microbiome of P. dendritica were identified as the dominant ones. In contrast, among the eight core ZOTUs of E. viridis, four were identified as dominant (ZOTU 1, 2, 3, and 8), whereas ZOTU 12, although dominant in P. dendritica, was not dominant in E. viridis. Additionally, three other ZOTUs (ZOTU 21, 22, and 24) were not dominant in either of the sea slug species. Those three ZOTUs exhibited a lower number of reads compared to the dominant taxa (< 3,000 sequence reads), with a total of only 581, 618, and 585 sequence reads, respectively. According to BLAST analysis, they were identified as Candidatus Mycoplasma mahonii (94.05% sequence homology), Reinekea sp. (87.5% sequence homology), and Gimesia algae (76.54% sequence homology) (Supplementary table S2).
Conclusions
This study provides novel insights into the bacterial communities associated with two sacoglossan sea slugs, E. viridis and P. dendritica, using a culture-independent approach based on 16S rRNA gene sequencing. Our findings reveal that E. viridis harbors a distinct and less diverse microbiome, characterized by the dominance of a few highly abundant ZOTUs, such as ZOTU 1 (Fulvibacter). Members of the genus Fulvibacter and the Flavobacteriaceae family are known for their carotenoid production, which may help mitigate oxidative stress associated with photosynthesis, potentially supporting the functionality of kleptoplasts in E. viridis. In contrast, P. dendritica supports a broader and more diverse bacterial community, likely providing functional redundancy and metabolic adaptability. The microbial assemblage in P. dendritica may support its heterotrophic lifestyle by facilitating nutrient acquisition and contributing to essential metabolic functions and defense. It is important to note that this study is based on evidence derived solely from 16S rRNA sequencing, which limits functional inference. Experimental and multi-omics approaches will be required to verify whether the bacterial taxa identified, particularly Fulvibacter, directly contribute to carotenoid biosynthesis and oxidative stress regulation in E. viridis.
Further research should characterize the identified bacterial taxa and elucidate their specific functional roles within sacoglossan sea slugs. Additionally, the low sequence homology observed between several ZOTUs and known database sequences suggests the presence of previously unidentified species, highlighting the uniqueness and ecological significance of these underexplored microbial communities.
Finally, it should be acknowledged that this study focused on populations of E. viridis and P. dendritica from the Portuguese coast, and therefore, the results should be interpreted within this geographic context. The core microbiome of these species may vary in other coastal regions due to environmental differences and local adaptations, which could shape host–microbe interactions.
Supplementary Information
Supplementary Material 1.
Supplementary Material 2.
Supplementary Material 3.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zan J, Li Z, Diarey Tianero M, Davis J, Hill RT, Donia MS. A microbial factory for defensive kahalalides in a tripartite marine symbiosis. Science. 2019;364:eaaw 6732. 10.1126/science.aaw 6732.10.1126/science.aaw 6732 PMC 1216815431196985 · doi ↗ · pubmed ↗
- 2Davis J. Characterization of the bacterial communities associated with the tropical sacoglossan mollusks Elysia rufescens and Elysia crispata. Ph D Thesis, University of Maryland; 2015. 10.13016/M 2QW 6D.