N‑Sulfated Heparan Sulfate Promotes Reelin Signaling as a Co-receptor
Lin Pan, Xuehong Song, Guowei Su, Lauren A Gandy, Biqin Fang, Mason Buttaci, James Gibson, Ke Xia, Fuming Zhang, Jian Liu, Lianchun Wang, Sally Temple, Chunyu Wang

TL;DR
N-sulfated heparan sulfate helps Reelin signaling in the brain, acting as a co-receptor and influencing diseases like Alzheimer's.
Contribution
HS N-sulfation is identified as a key determinant for Reelin signaling, establishing HS as a co-receptor.
Findings
Full-length Reelin binds HS with high affinity (K_D = 17 ± 5 nM), enhanced by the COLBOS variant.
HS N-sulfation is critical for Reelin–HS binding, confirmed by SPR, glycan arrays, and Haddock modeling.
Heparinase treatment or NDST1 knockout reduces Reelin binding and receptor activation, showing HS's coreceptor role.
Abstract
Heparan sulfate (HS) plays a central role in signal transduction, while Reelin is an essential signaling protein in both the developing and adult brain. A Reelin COLBOS variant was recently discovered with enhanced HS binding and resilience against autosomal dominant Alzheimer’s disease (ADAD), underscoring the importance of Reelin–HS interactions. However, the glycan determinants of Reelin–HS interactions have not been well-characterized, which we systematically investigated here. Surface plasmon resonance (SPR) showed that full length Reelin binds HS with high affinity (K D = 17 ± 5 nM), which is enhanced by the COLBOS variant (K D = 10 ± 2 nM). Competition SPR and glycan array studies further revealed that HS N-sulfation is critical for Reelin–HS binding, consistent with Haddock modeling. In cell surface binding assays, heparinase treatment, which degrades HS, or the knockout of a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
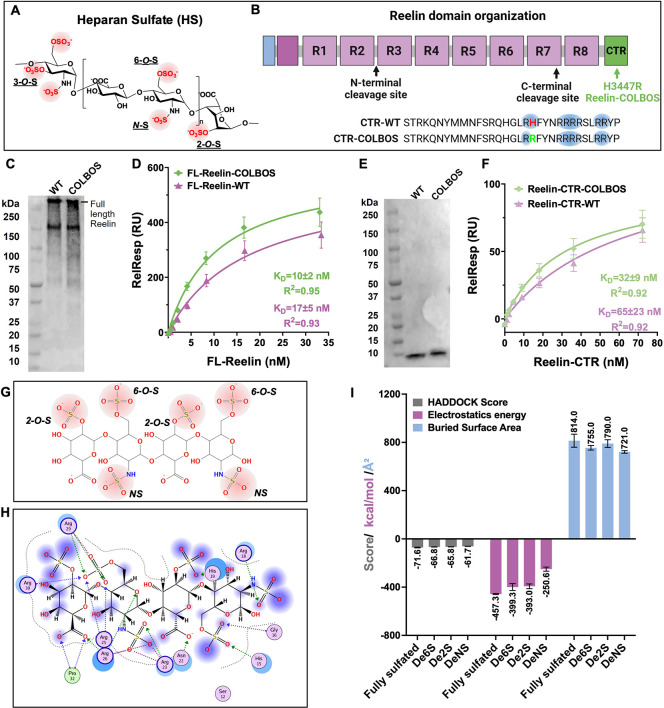 Figure 1
Figure 1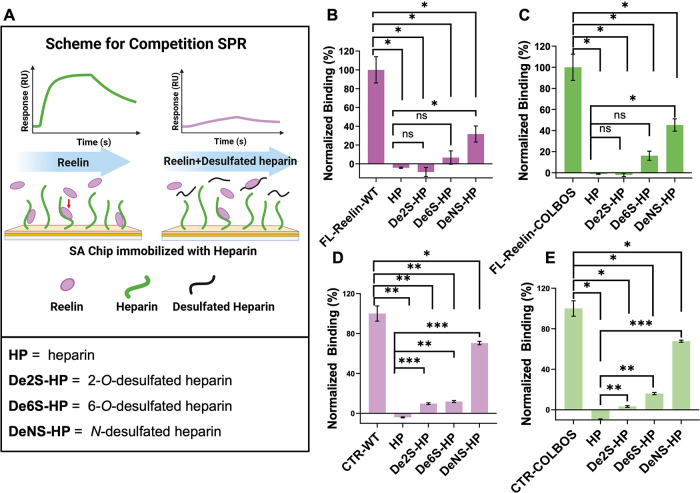 Figure 2
Figure 2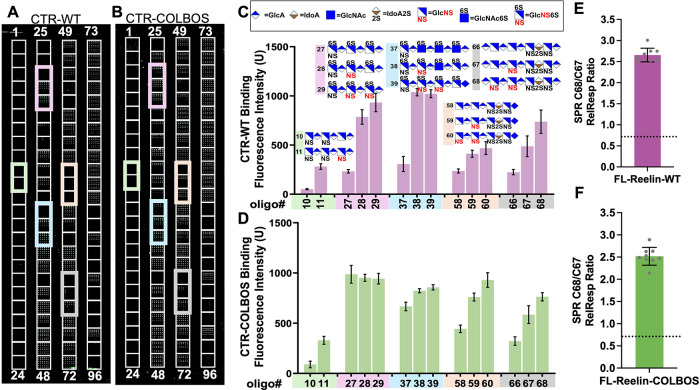 Figure 3
Figure 3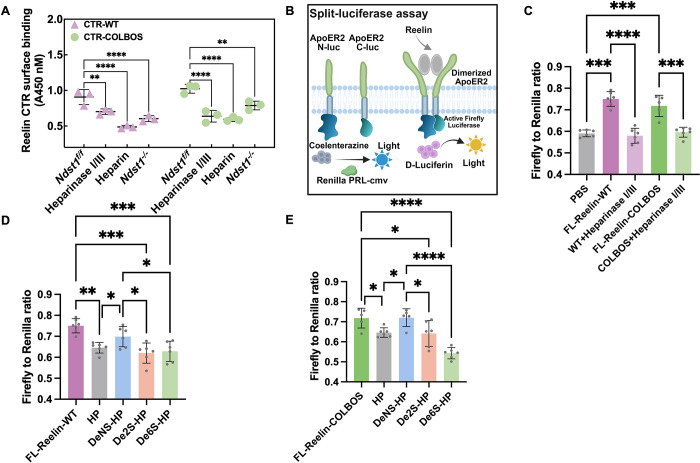 Figure 4
Figure 4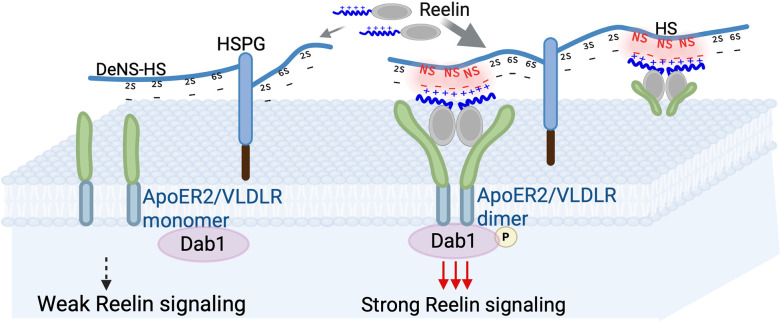 Figure 5
Figure 5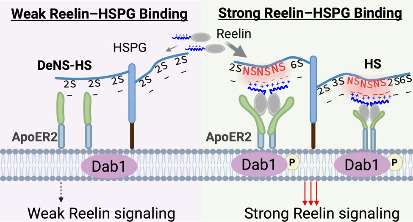 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurogenesis and neuroplasticity mechanisms · Nerve injury and regeneration · Alzheimer's disease research and treatments
Reelin is a large, secreted extracellular glycoprotein? (FigureB), which binds to receptors such as ApoER2 (LRP8) and VLDLR to activate intracellular signaling cascades.? Disruptions in Reelin signaling are linked to many neuropsychiatric disorders, including depression, schizophrenia, autism spectrum disorders, epilepsy, and recently AD. ?,? In 2023, Quiroz et al. reported the world’s second case of extreme resilience to autosomal dominant AD (ADAD): a patient with a RELN-COLBOS (H3447R) genetic variant was protected against dementia despite carrying an ADAD mutation, PSEN1 E280A.? Interestingly, H3447R does not affect Reelin binding to ApoER2 or VLDLR. Quiroz et al. demonstrated that the H3447R promotes Reelin binding to HS, likely by increasing electrostatic interactions between the positive charges on Reelin (e.g., arginine side chains) and negatively charged sulfo groups on HS. These intriguing data from the COLBOS genetic variant suggest that heparan sulfate may play an important role in the Reelin signaling pathway.
Heparan sulfate (HS) is an important class of polyanionic glycosaminoglycans (GAGs), with crucial signaling roles in the central nervous system? such as mediating neuronal migration in development.? Structurally, HS and heparin (HP, a highly sulfated HS analogue) are linear GAGs composed of repeating disaccharide units of N-acetylglucosamine (GlcNAc) and uronic acid (either d-glucuronic acid (GlcA) or l-iduronic acid (IdoA)) (FigureA). These units are modified by sulfotransferases in the Golgi, starting with N-sulfation (NS) of glucosamine, followed by 2-O-sulfation (2S) of uronic acids, and finally 6-O-sulfation (6S) and 3-O-sulfation (3S) of glucosamine, creating diverse sulfation patterns.? Changes in sulfation of HS chains can greatly alter the affinity of HS-binding proteins, with important consequences on cellular signaling and biological function. ?−? ?
We first evaluated the binding affinity between Reelin and heparin (a commonly used HS analog). We expressed and purified full-length Reelin (FL-Reelin; ∼400 kDa; FigureB) with either the wild-type (FL-Reelin-WT) or COLBOS (FL-Reelin-COLBOS) sequence (FigureC; Figure S1A and B) in mammalian cells. The COLBOS variant is in the Reelin C-terminal region (CTR), which plays an important role in heparin binding.? The CTR of Reelin WT (Reelin-CTR-WT) and the COLBOS (Reelin-CTR-COLBOS) variant were expressed in E. coli and purified (FigureE; Figure S1C–E). Then we performed the SPR analysis on a heparin biochip as described in our previous studies.? Binding affinity was determined using a steady-state affinity equation.? FL-Reelin-WT exhibited a K D of 17 ± 5 nM, while the FL-Reelin-COLBOS variant showed an enhanced binding affinity, with a K D of 10 ± 2 nM (FigureD). The CTR retains high-affinity binding to heparin with a K D of 65 ± 23 nM for the Reelin-CTR-WT. As expected, the Reelin-CTR-COLBOS variant displayed enhanced binding, with a K D of 32 ± 9 nM (FigureF). These data are consistent with previous bilayer interferometry (BLI) and isothermal titration calorimetry (ITC) studies of CTR–heparin interaction.? However, in Quiroz et al.? only studied Reelin-heparin binding with CTR. Here, we have extended the binding studies to FL-Reelin. Because FL-Reelin and the CTR have K D values on the same order of magnitude, CTR is likely the major heparin or HS binding site in Reelin.
To dissect the role of individual sulfate groups in HS–Reelin binding, we employed Haddock? to dock Reelin CTR to a canonical HS tetrasaccharide containing two N-, two 2-O-, and two 6-O-sulfate groups (FigureG). Targeted removal of each sulfate type yielded three selectively desulfated variants (DeNS, De2S, and De6S), which were docked to the Reelin-CTR to assess their relative binding contributions. Docking analysis showed that clustered sulfate groups form salt bridges and hydrogen bonds with multiple basic residues in the Reelin-CTR, consistent with multivalent electrostatic recognition (Figure.H). R18 and R23 in the Reelin-CTR contribute to recognition of N-sulfated heparan sulfate motifs, providing a plausible structural explanation for its preference toward specific HSPG species. A fully sulfated HS tetrasaccharide displayed the strongest complementarity and largest buried surface area, whereas removal of the N-sulfate group caused the greatest loss of electrostatic energy (>+200 kcal/mol), reduced binding interface (∼90 Å^2^), and markedly impaired docking score. In contrast, 2-O-desulfation had little effect, and 6-O-desulfation only moderately weakened the interaction in docking studies (Figure.I). These results suggest that N-sulfation is the key glycan determinant of charge-mediated Reelin-HS binding.
To validate the computational prediction, competition SPR (FigureA) was performed by using selectively desulfated heparin. Unmodified heparin (HP) fully blocked both FL-Reelin and Reelin-CTR binding to chip-immobilized heparin, whereas N-desulfated heparin (DeNS) showed much less inhibition, indicating the loss of binding of DeNS to Reelin in solution (FigureB-E). In contrast, 2-O- and 6-O-desulfated heparin inhibited binding almost as effectively as unmodified heparin in FL-Reelin (FigureB, C). A similar trend was observed for the Reelin-CTR results (FigureD, E). These results highlight the critical role of N-sulfation in mediating both FL-Reelin and Reelin-CTR interaction with HS, consistent with the computational prediction. In addition, the COLBOS mutation enhanced overall binding affinity but did not alter sulfation-site preferences for sulfation. Using competition SPR, we also examined the effect of heparin chain length on Reelin binding (Figure S4). The results indicated that the longer heparin chains exhibited stronger binding to Reelin.
We then used a glycan microarray of 96 chemically defined HS oligosaccharides? (Figure S2) to validate the above results from desulfated heparin. Biotin-labeled Reelin-CTR bound to the HS-microarray was detected with fluorescence-labeled streptavidin. The results showed that Reelin binding intensity correlated strongly with chain length and sulfation level (Figure S3), consistent with competition SPR results (Figure S4). Among sulfation patterns, N-sulfation had the most pronounced positive effect: within oligosaccharides of the same chain length and disaccharide composition, the addition of each N-sulfate group markedly increased Reelin-CTR-WT binding (e.g., compound 11 vs compound 10), a trend reproduced across multiple oligosaccharide sets (FigureA, C). Reelin-CTR COLBOS displayed higher overall binding than WT but retained the same dependence on Nsulfation (FigureB, D). In contrast, 6S or 2S modifications showed no consistent effect on binding (Figure S3). To further confirm the role of N-sulfation in FL-Reelin binding, we compared two representative HS oligosaccharides, C67 and C68 (FigureC), which differ only by a single N-sulfate group. SPR analysis showed that C68 bound significantly more strongly to FL-Reelin, with FL-Reelin-WT exhibiting an ∼2.5-fold higher response to C68 than C67 (FigureE). Similar preferences were observed for COLBOS variants (FigureF), demonstrating that a single N-sulfation markedly enhances Reelin–HS interactions, highlighting the key role of N-sulfation.
To evaluate the role of N-sulfation in Reelin-HS binding at the cellular level, we conducted cell surface binding assays. N-deacetylase/sulfotransferase 1 (Ndst1) is responsible for N-sulfation in heparan sulfate biosynthesis. We utilized wild-type (Ndst1f/f) mouse lung endothelial cells (MLECs) which have abundant HS on their cell surface, and Ndst1 knockout (Ndst1–/−) MLECs which show reduced levels of N-sulfation (by ∼40%), 6S (by ∼10%), and 2S (by ∼15%).? The additional treatment with Heparinase I/III or heparin was included to examine the involvement of HS in binding of Reelin to the cell surface. Biotin-labeled Reelin-CTR (WT and COLBOS) bound to cell surfaces was detected using streptavidin-HRP. Heparinase I/III treatment or soluble heparin competition markedly reduced binding in Ndst1f/f cells (FigureA), confirming the critical role of HS in mediating Reelin’s interaction with the cell surface. The cell surface binding was further diminished in Ndst1–/– cells to ∼40% for WT and ∼30% for COLBOS, demonstrating that N-sulfation is essential for Reelin–HS interactions, consistent with SPR and glycan array results (FigureA).
Reelin-mediated dimerization or higher-order multimers of ApoER2 are involved in the activation of the Reelin signaling pathway. ? ? To assess the role of HS in Reelin receptor activation, we employed a split-luciferase assay.? HEK293T cells were transfected with plasmids encoding ApoER2-N-Luc, ApoER2-C-Luc, and Renilla Luciferase (as a control for transformation efficiency), where N-Luc and C-luc represent N- and C-terminal halves of firefly luciferase, respectively. Upon Reelin binding, ApoER2 will dimerize and reconstitute firefly luciferase activity (FigureB). In the luciferase assay, both FL-Reelin-WT and COLBOS significantly increased the Firefly/Renilla ratio relative to PBS, confirming receptor activation (FigureC). Heparinase I/III treatment degraded cell-surface HS and markedly reduced Reelin-induced ApoER2 dimerization in both WT and COLBOS, without affecting the Firefly/Renilla ratio (FiguresC, S5). Exogenous heparin strongly inhibited ApoER2 dimerization, while desulfated derivatives (DeNS-HP, De2S-HP, and De6S-HP) showed differential effects. DeNS-HP had the weakest inhibition, highlighting the essential role of N-sulfation, whereas 2-O- and 6-O-sulfation were less critical, as De2S-HP and De6S-HP inhibited ApoER2 dimerization similarly to heparin (FiguresD–E, S6). These results demonstrated that N-sulfation plays a crucial role in Reelin-HS binding at the cellular level.
Here, using SPR competition, glycan arrays, knockout cell lines, and luciferase assays, we show that HS acts as a glycan coreceptor in Reelin signaling activation, with its sulfation pattern serving as a key modulator of the Reelin–HS interaction (Figure). Both Reelin-WT and Reelin-COLBOS exhibit high-affinity binding to heparan sulfate and depend critically on N-sulfation for this interaction and downstream ApoER2 activation. The two proteins share similar overall binding profiles in surface plasmon resonance, glycan array, and cell-based assays, confirming that the COLBOS mutation does not alter the fundamental mechanism of Reelin–HS recognition. Nevertheless, the COLBOS variant shows a modest (∼1.5–2-fold) increase in overall affinity and a subtle shift in glycan preference, displaying stronger binding to HS structures containing fewer N-sulfation in both glycan array and cell-surface binding assays. H3447R substitution enhances the local positive charge density in the C-terminal domain, improving electrostatic complementarity with negatively charged HS motifs. Thus, our data suggest that the COLBOS mutation fine-tunes, rather than dramatically increases, Reelin–HS interactions, providing a plausible molecular basis for its reported neuroprotective effect. This work provides a potential basis for developing therapeutic strategies targeting Reelin signaling.
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1D’Arcangelo G.Nakajima K.Miyata T.Ogawa M.Mikoshiba K.Curran T.Reelin Is a Secreted Glycoprotein Recognized by the CR-50 Monoclonal Antibody J. Neurosci.19971712310.1523/JNEUROSCI.17-01-00023.19978987733 PMC 6793694 · doi ↗ · pubmed ↗
- 2Hiesberger T.Trommsdorff M.Howell B. W.Goffinet A.Mumby M. C.Cooper J. A.Herz J.Direct binding of Reelin to VLDL receptor and Apo E receptor 2 induces tyrosine phosphorylation of disabled-1 and modulates tau phosphorylation Neuron 199924248148910.1016/S 0896-6273(00)80861-210571241 · doi ↗ · pubmed ↗
- 3Bracher-Smith M.Leonenko G.Baker E.Crawford K.Graham A. C.Salih D. A.Howell B. W.Hardy J.Escott-Price V.Whole genome analysis in APOE 4 homozygotes identifies the DAB 1-RELN pathway in Alzheimer’s disease pathogenesis Neurobiol Aging 2022119677610.1016/j.neurobiolaging.2022.07.00935977442 PMC 9548409 · doi ↗ · pubmed ↗
- 4Joly-Amado A.Kulkarni N.Nash K. R.Reelin signaling in neurodevelopmental disorders and neurodegenerative diseases Brain Sciences 20231310147910.3390/brainsci 1310147937891846 PMC 10605156 · doi ↗ · pubmed ↗
- 5Lopera F.Marino C.Chandrahas A. S.O’Hare M.Villalba-Moreno N. D.Aguillon D.Baena A.Sanchez J. S.Vila-Castelar C.Ramirez Gomez L.Resilience to autosomal dominant Alzheimer’s disease in a Reelin-COLBOS heterozygous man Nature Medicine 20232951243125210.1038/s 41591-023-02318-3PMC 1020281237188781 · doi ↗ · pubmed ↗
- 6Bandtlow C. E.Zimmermann D. R.Proteoglycans in the developing brain: new conceptual insights for old proteins Physiol. Rev.20008041267129010.1152/physrev.2000.80.4.126711015614 · doi ↗ · pubmed ↗
- 7Ozsan Mc Millan I.Li J. P.Wang L.Heparan sulfate proteoglycan in Alzheimer’s disease: aberrant expression and functions in molecular pathways related to amyloid-beta metabolism Am. J. Physiol Cell Physiol 20233244 C 893C 90910.1152/ajpcell.00247.202236878848 PMC 10069967 · doi ↗ · pubmed ↗
- 8Li J. P.Kusche-Gullberg M.Heparan sulfate: biosynthesis, structure, and function International review of cell and molecular biology 201632521527310.1016/bs.ircmb.2016.02.00927241222 · doi ↗ · pubmed ↗