Depression Aggravates Immune‐Mediated Hepatitis Through NLRP3 Overactivation Induced by Intestinal Microbiota
Simin Zhou, Liping Guo, Nian Chen, Haifeng Liu, Xin Liu, Jiwen Li, Shijing Dong, Jiangpeng Liu, Xiaoyi Wang, Ying Ran, Man Liu, Hongyu Chu, Yanni Li, Hui Yang, Jingwen Zhao, Lu Zhou

TL;DR
Depression worsens liver inflammation in autoimmune hepatitis by disrupting gut barriers and overactivating liver immune responses, with a specific gut bacteria playing a key role.
Contribution
This study identifies a novel mechanism linking depression to immune-mediated liver injury through gut microbiota and NLRP3 inflammasome activation.
Findings
Depression in AIH patients is linked to worsened liver outcomes and increased cirrhosis risk.
Gut microbiota from depressed individuals induces liver inflammation via NLRP3 overactivation in mice.
Lactococcus formosensis from the gut can translocate to the liver and cause liver injury.
Abstract
Depression is associated with adverse effects in patients with autoimmune hepatitis (AIH). However, the underlying mechanism remains unclear. This study explores the impact of depression and related intestinal microbiota on immune‐mediated hepatitis. We assessed depression in 260 AIH patients receiving 2‐year standardized treatment and 173 healthy controls. In mice, depressive‐like behaviors were induced by chronic unpredictable mild stress (CUMS), and immune‐mediated hepatitis was induced by intravenous injection of concanavalin A (ConA). Fecal microbiota transplantation (FMT) was performed using samples from patients with major depressive disorder (MDD) and controls. Depression was common in patients with AIH (106/260, 40.8%) and was associated with cirrhosis. Compared with nondepressed AIH patients, those with depression showed exacerbated intestinal barrier dysfunction and hepatic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
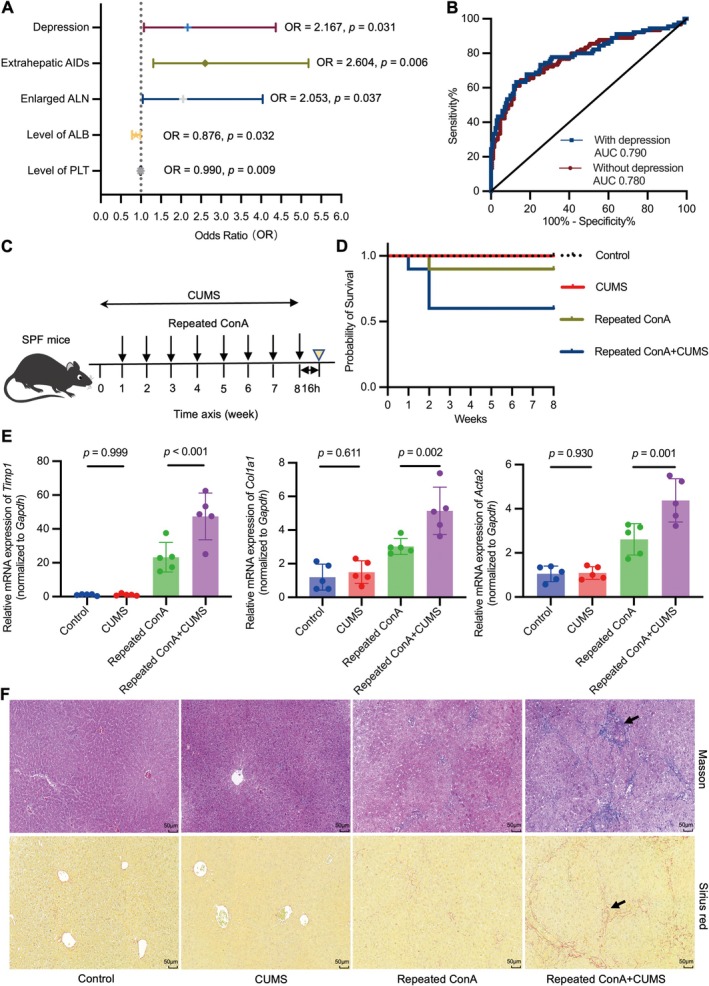 FIGURE 1
FIGURE 1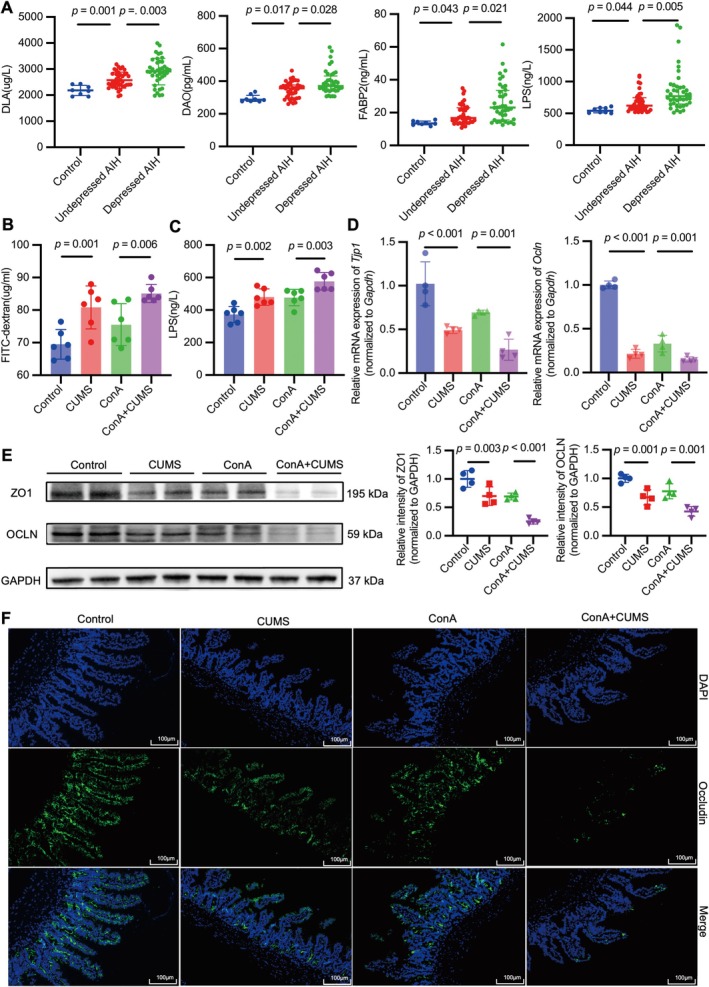 FIGURE 2
FIGURE 2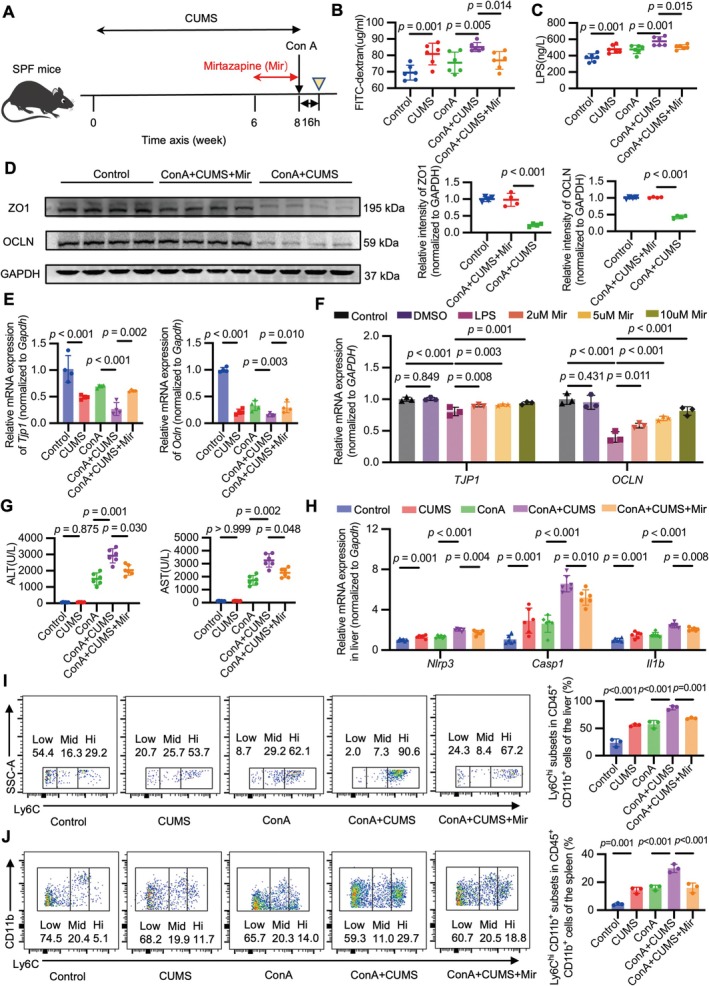 FIGURE 3
FIGURE 3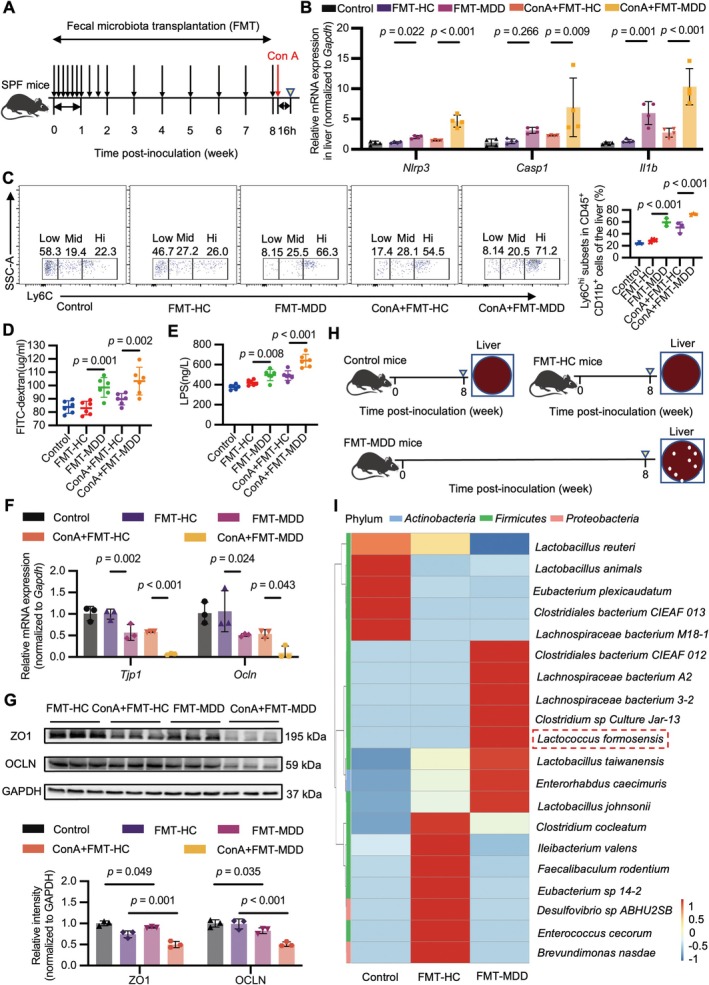 FIGURE 4
FIGURE 4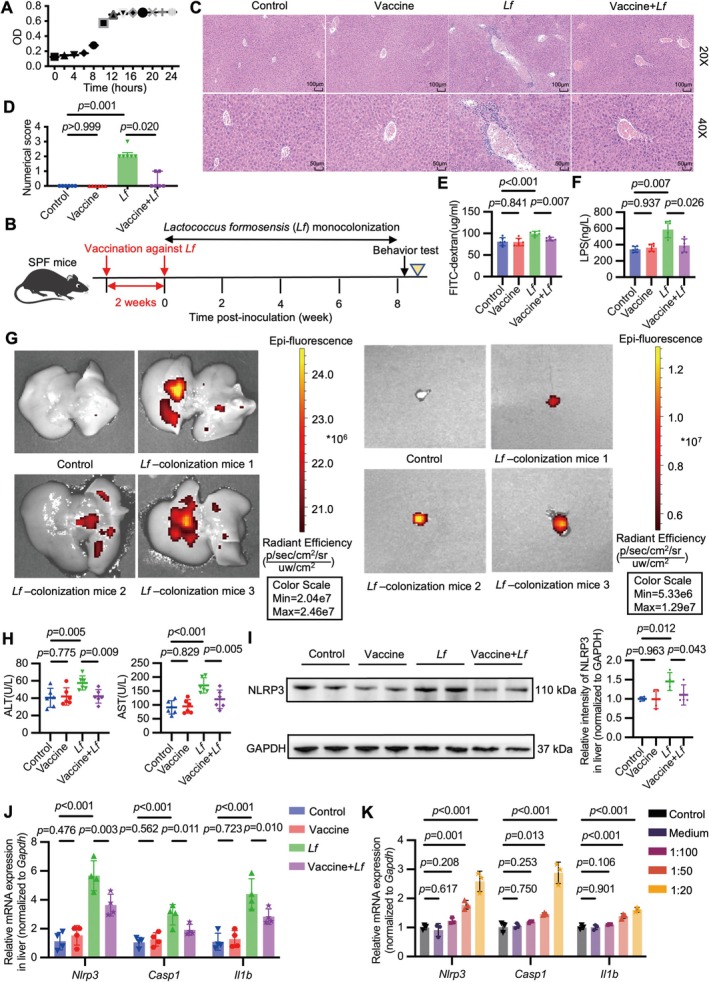 FIGURE 5
FIGURE 5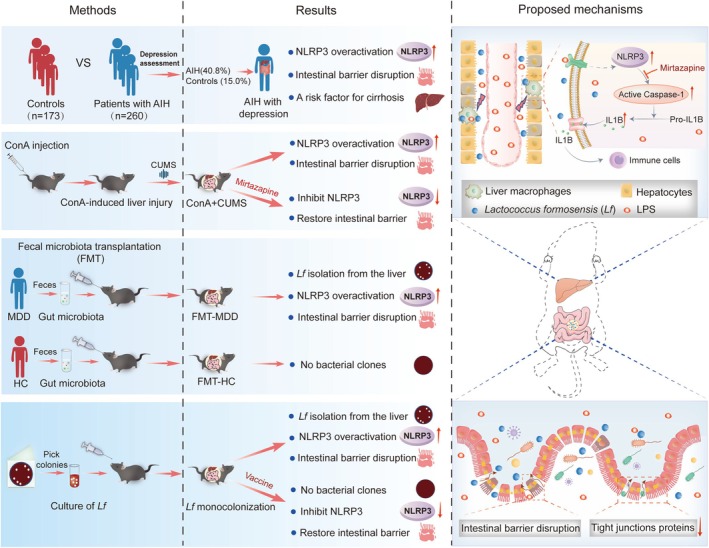 FIGURE 6
FIGURE 6| Without depression ( | With depression ( |
| |
|---|---|---|---|
| General characteristics | |||
| Age, median (IQR) | 58.5 (48.0–65.0) | 57.5 (49.0–63.0) | 0.509 |
| Sex: women, | 131 (85.1%) | 94 (88.7%) | 0.401 |
| Laboratory parameters (normal range) | |||
| ALT (5.0–40.0 U/L), median (IQR) | 31.0 (27.0–36.0) | 36.5 (27.0–51.0) | 0.002 |
| AST (8.0–40.0 U/L), median (IQR) | 33.0 (28.0–38.0) | 37.5 (27.0–50.0) | 0.005 |
| IgG (751.0–1560.0 mg/dL), median (IQR) | 1450.0 (1320.0–1530.0) | 1610.0 (1400.0–1752.5) | < 0.001 |
| C3 (79.0–152.0 mg/dL), median (IQR) | 100.0 (85.5–116.3) | 97.8 (79.0–109.3) | 0.044 |
| C4 (16.0–38.0 mg/dL), median (IQR) | 21.0 (17.4–25.9) | 19.5 (15.8–21.9) | 0.003 |
| Disease assessment | |||
| Cirrhosis, | 35 (22.7%) | 55 (51.9%) | < 0.001 |
| Extrahepatic AIDs, | 47 (30.5%) | 73 (68.9%) | < 0.001 |
| Enlarged ALN, | 68 (44.2%) | 67 (63.2%) | 0.003 |
- —National Natural Science Foundation of China10.13039/501100001809
- —Natural Science Foundation of Tianjin Municipality10.13039/501100006606
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGut microbiota and health · Liver Diseases and Immunity · Tryptophan and brain disorders
Introduction
1
Autoimmune hepatitis (AIH), characterized by the disruption of immune tolerance, is an immune‐mediated hepatitis with an increasing incidence [1]. A major depressive syndrome is five times more frequent in patients with AIH than in the general population [2]. Moreover, patients with other autoimmune diseases (AIDs) exhibit a significantly higher prevalence of depression than healthy controls [3, 4]. Depression increases the risk of nonadherence to standard therapy and relapse in patients with AIH [5], and is indeed an important risk factor for liver fibrosis [6, 7, 8]. Notably, patients with major depressive disorder (MDD) exhibit elevated frequencies of antinuclear antibody (ANA), thyroid gland antibody, and parietal cell antibody [9]. Similarly, another study demonstrated the potential role of T helper 17 cells in autoimmune disorders in patients with MDD [10], further supporting the existing evidence of autoimmune dysregulation in depression. Therefore, the role of depression in immune disorders during AIH progression warrants clinical attention.
Accumulating evidence suggests that depression drives inflammation and promotes immune activation [9, 10]. Macrophages play vital roles in maintaining immune homeostasis in the liver [11]. However, the persistent activation of hepatic macrophages results in pathological inflammation and fibrosis in chronic liver disease [12]. Meanwhile, liver‐resident macrophages can produce significant amounts of NLR family pyrin domain containing 3 (NLRP3) and interleukin 1 beta (IL1B) [13]. The NLRP3 inflammasome complex, which consists of NLRP3, pro‐caspase‐1 (proCASP1), and apoptosis‐associated speck‐like protein, can regulate the cleavage and activation of serine protease CASP1 and drive the production of pro‐inflammatory cytokine IL1B [14]. Several studies have indicated that the NLRP3 inflammasome is involved in the progression of depression [15, 16, 17]. NLRP3 activation is elevated in peripheral blood mononuclear cells of patients with depression and can be reversed by antidepressant therapy [18]. In the chronic unpredictable mild stress (CUMS)‐treated rats, NLRP3 inflammasome overactivation is observed in the liver [19, 20, 21]. In addition, the concanavalin A (ConA)‐induced murine hepatitis model exhibits aberrant NLRP3 inflammasome activation [22, 23, 24, 25], whereas relevant research in patients with AIH remains limited. Collectively, these findings demonstrate that NLRP3 activation is involved in the progression of both depression and immune‐mediated hepatitis.
Many studies have demonstrated that intestinal dysbiosis [26, 27] and intestinal barrier disruption [28, 29] contribute to the pathogenesis of AIH. Additionally, altered gut microbiota composition [30, 31, 32] and increased intestinal permeability [33] also participate in the pathogenesis of depression. Fecal microbiota transplantation (FMT) from patients with depression resulted in depressive‐like behaviors in recipient germ‐free mice [34]. Importantly, transplantation of fecal microbiota from NLRP3‐deficient mice, whose production of pro‐inflammatory cytokines was limited, alleviated depressive‐like behaviors in recipient wild‐type mice [35]. In the Mdr2^−/−^ murine model of primary sclerosing cholangitis, gut microbiota dysbiosis prompted the translocation of endotoxin to the hepatic portal system, causing severe liver injury due to pronounced NLRP3 inflammasome activation [36]. Hence, it is important to elucidate the impact of depression‐associated gut dysbiosis on the activation of the NLRP3 inflammasome during the progression of immune‐mediated hepatitis.
In this study, we investigated the effects of depression on AIH progression and elucidated the role of depression‐associated intestinal dysbiosis in this process. Specifically, we explored whether this exacerbation was mediated by the activation of the NLRP3 inflammasome in hepatic macrophages.
Materials and Methods
2
The Data S1 include an expanded “Supporting Informations and Methods.”
Design and Subjects
2.1
Consecutive patients diagnosed with AIH who visited our hepatology clinic between December 2021 and December 2023 were enrolled (n = 260). To eliminate the potential influence of illness duration on depressive symptoms, only patients who received standardized treatment for 2 years were recruited. AIH was diagnosed in accordance with the European Association for the Study of the Liver guidelines [37]. The inclusion criteria were as follows: (1) AIH diagnosis; (2) age > 18 years; (3) standardized treatment of prednisone and/or azathioprine for 2 years; (4) willingness to participate in the assessment of depressive symptoms performed by two trained clinicians using the 24‐item Hamilton Rating Scale for Depression (HAMD‐24); (5) absence of viral hepatitis or malignant tumors. Healthy individuals (n = 173) were enrolled from a health management center based on the following criteria: (1) matched for age and sex; (2) willingness to participate in the HAMD‐24 assessment; (3) no history of liver diseases; (4) no history of autoimmune diseases or malignant tumors. Informed consent was obtained from all participants. This study was approved by the Tianjin Medical University Ethics Committee, based on the ethical guidelines of the Declaration of Helsinki (revised 2013, Fortaleza, Brazil) (Ethical Approval No. IRB2021‐WZ‐191).
Data Collection
2.2
Two trained clinicians independently assessed depressive symptoms in patients with AIH who received standardized treatment for 2 years. During the assessment period, we systematically collected the most recent hepatic pathology reports, diagnostic imaging reports, and laboratory parameters (including white blood cells [WBC], hemoglobin [Hb], platelets [PLT], total protein [TP], albumin [ALB], globulin [GLO], total bilirubin [TB], direct bilirubin [DB], alanine aminotransferase [ALT], aspartate aminotransferase [AST], alkaline phosphatase [ALP], γ‐glutamyl transpeptidase [GGT], complement component 3 [C3], complement component 4 [C4], immunoglobulin G [IgG], IgM, and ANA). The diagnosis of cirrhosis was established based on hepatic pathology. Liver biopsy was unnecessary when clinical evidence of cirrhosis was present, including imaging report of a nodular and shrunken liver, coagulopathy, and ascites.
Construction of Logistic Regression Model
2.3
Comparisons of continuous variables between cirrhotic and noncirrhotic AIH patients were performed using the unpaired t‐test for normally distributed data and the Mann–Whitney U test for non‐normally distributed data. Differences in categorical variables were analyzed using the Chi‐square test. Variables showing statistically significant differences between cirrhotic and noncirrhotic AIH patients (including age, levels of ALB, GLO, ALT, AST, ALP, WBC, Hb, PLT, IgG, C3, C4, ANA titer, and comorbidities, such as depression, extrahepatic AIDs, and enlarged abdominal lymph nodes [ALN]) were further selected to build the logistic regression model. The model performance was evaluated by the calculation of area under the receiver operating characteristic curve (AUC‐ROC).
Animals and Treatments
2.4
Seven‐week‐old female C57BL/6 mice (18–20 g) were purchased from the Beijing Animal Study Center. The experimental animals were raised under specific‐pathogen free conditions at a constant temperature of 22°C and relative humidity of 55%. All mice received the same autoclaved water and chow under a 12‐h light/dark cycle. Female mice were selected because of the female predominance observed in patients with AIH (75%) [38] and depression (64.7%) [39]. In ConA‐induced hepatitis model, female mice exhibit more severe hepatic damage and elevated production of pro‐inflammatory cytokines than male mice [40]. An ovariectomy in female mice decreases levels of ALT and pro‐inflammatory cytokines and ameliorates ConA‐induced liver injury [40]. In contrast, orchiectomy in male mice exacerbates hepatic damage by promoting the production of pro‐inflammatory cytokines [40]. Consistent with the observations in the ConA‐induced liver injury model, female subjects are inclined to be more pro‐inflammatory and more sensitive to CUMS stimulation [41]. Furthermore, testosterone treatment correlates with improvement in depressive symptoms in men [42]. Based on these considerations, only female mice were included in the present study. Mice were randomized to different experimental groups using a random number table. After the behavioral evaluation, mice were sacrificed under anesthesia. The animal experiments complied with the ARRIVE guidelines. All experimental procedures were approved by the Ethics and Welfare Committee of Animals at Tianjin Medical University (Ethical No. IRB2023‐DWFL‐257).
Statistical Analysis
2.5
Statistical analysis was performed using IBM SPSS Statistics, version 26.0. The normality of the data distribution was assessed using the Kruskal–Wallis test (for a sample size ≥ 50) or Shapiro–Wilk test (for a sample size < 50). Continuous data are presented as mean ± standard deviation (SD) (normally distributed) or median (interquartile range [IQR]) (non‐normally distributed). Variables with normal distribution were analyzed using unpaired t‐tests or one‐way analysis of variance (ANOVA). Multiple comparisons were performed based on least significant difference (LSD) post hoc test when variances were equal and Tamhane's T2 post hoc test when variances were unequal. Nonparametric tests were conducted to compare non‐normally distributed variables. Differences in categorical variables were determined using the Chi‐square test. All reported p‐values were two‐sided. Statistical significance was set at p < 0.05. All figures were plotted using GraphPad Prism version 9.0.
Results
3
Depression Correlates With Disease Severity in Patients With AIH
3.1
Patients with AIH who received standardized treatment for 2 years (n = 260) were recruited. Age‐ and sex‐matched healthy participants (n = 173) were selected as controls (Table S1). When assessed using the HAMD questionnaire, 40.8% (106/260) of AIH patients presented with symptoms of depression, significantly higher than the rate in healthy controls (15.0%, 26/173; p < 0.001). Furthermore, severe symptoms of depression (HAMD score > 20) were more frequent in patients with AIH than in controls (1.2% vs. 0.0%, p < 0.001). AIH patients with depression presented with higher serum levels of ALT, AST, and IgG and lower serum levels of C3 and C4 than those without depression (p = 0.002, p = 0.005, p < 0.001, p = 0.044, p = 0.003; Table 1 and Table S2). Moreover, the comorbidities of cirrhosis, extrahepatic AIDs, and enlarged ALN were more frequent in AIH patients with depression than in those without depression (p < 0.001, p < 0.001, p = 0.003; Table 1 and Table S2).
Depression Promotes the Progression of Fibrosis in Immune‐Mediated Hepatitis
3.2
Among the 260 patients with AIH, 90 patients (34.6%) presented with liver cirrhosis. The rate of cirrhosis was higher in AIH patients with depression (55/106, 51.9%) than in those without depression (35/154, 22.7%) (p < 0.001). The following 16 parameters showed a statistical difference between AIH patients with and without cirrhosis: age, ALB, GLO, ALT, AST, ALP, WBC, Hb, PLT, IgG, C3, C4, ANA titer, and comorbidities such as depression, extrahepatic AIDs, and enlarged ALN (Table S3). In the binary logistic regression model incorporating these 16 indicators, depression (B = 0.773, odds ratio [OR] = 2.167, 95% confidence interval [CI], 1.075–4.366, p = 0.031) and the other four variables, including extrahepatic AIDs (B = 0.957, OR = 2.604, 95% CI: 1.308–5.185, p = 0.006), enlarged ALN (B = 0.719, OR = 2.053, 95% CI: 1.044–4.039, p = 0.037), level of ALB (B = −0.132, OR = 0.876, 95% CI: 0.777–0.988, p = 0.032), and level of PLT (B = −0.010, OR = 0.990, 95% CI: 0.983–0.998, p = 0.009), were identified as independent predictors of cirrhosis in patients with AIH (Figure 1A). The model including depression and the other four indicators exhibited a higher AUC value (0.790, 95% CI: 0.726–0.853; p < 0.001) compared with the model excluding the variable of depression (0.780, 95% CI: 0.716–0.844; p < 0.001) (Figure 1B).
Depression promotes the progression of fibrosis in immune‐mediated hepatitis. (A) Logistic regression analysis was employed to identify indicators associated with cirrhosis in patients with autoimmune hepatitis (AIH). (B) The performance of the logistic model was evaluated by the calculation of area under the receiver operating characteristic curve (AUC‐ROC). With depression: Depression, extrahepatic autoimmune diseases (AIDs), enlarged abdominal lymph node (ALN), level of albumin (ALB), and level of platelet (PLT); Without depression: The above factors, excluding depression. (C) Study design: Mice were challenged with chronic unpredictable mild stress (CUMS) daily and concanavalin A (ConA) once a week during the course of 8 weeks. The mice were sacrificed 16 h after the last injection of ConA (yellow triangle). (D) The survival curve of different groups. (E) Quantitative real‐time polymerase chain reaction (PCR) analysis for tissue inhibitor of metalloproteinase 1 (Timp1), collagen, type I, alpha 1 (Col1a1), and Actin alpha 2 (Acta2) relative to glyceraldehyde‐3‐phosphate dehydrogenase (Gapdh) in liver tissues from the indicated groups. F = 36.411, 19.865, 29.729, all Df = 3, all n = 5. (F) Photo micrographs of Masson trichrome staining (upper) and Sirius red staining (lower) of the liver sections from the indicated groups. Scale bars, 50 μm. Data are presented as mean ± standard deviation (SD). Statistical comparisons between groups were performed using one‐way analysis of variance (ANOVA) and the least significant difference (LSD) post hoc test.
To validate the impact of depression on liver fibrosis, mice with fibrosis induced by repeated ConA treatment were subjected to CUMS intervention (Figure 1C). CUMS intervention induced depressive‐like behaviors, manifested by weight loss, lower sucrose preference index, reduced total distance traveled, and decreased frequency of entering the central area (Figure S1A,D,G,J). In addition, CUMS intervention increased mortality in mice subjected to repeated ConA‐induced liver fibrosis (p = 0.006; Figure 1D). Importantly, the gene expression of fibrosis markers, including collagen, type I, alpha 1 (Col1a1), tissue inhibitor of metalloproteinase 1 (Timp1), and actin alpha 2 (Acta2), was significantly upregulated in the liver tissues of repeated ConA‐treated mice with CUMS intervention compared with those without (Figure 1E). Further, hepatic fibrosis was pathologically more severe in ConA‐treated mice with CUMS intervention than in those without, as measured by Masson trichrome staining and Sirius red staining (Figure 1F). Collectively, these observations provide experimental evidence that depression promotes fibrosis progression in immune‐mediated hepatitis.
Depression Promotes Hepatic NLRP3 Activation in Immune‐Mediated Hepatitis
3.3
To explore the effects of depression on the activation of NLRP3 inflammasome in patients with AIH, we collected liver tissues from AIH patients with depression, AIH patients without depression, and age‐ and sex‐matched controls (Table S4). The mRNA expression levels of NLRP3, CASP1, and IL1B were higher in liver biopsies from AIH patients with depression than in those without depression (Figure S2A). Immunofluorescence analysis showed a more pronounced co‐localization of NLRP3 with the macrophage marker CD68 in the hepatic tissues of depressed AIH patients compared with nondepressed patients (Figure S2B).
Next, we investigated the impact of CUMS intervention on hepatic NLRP3 expression in mice (Figure S2C). CUMS intervention resulted in elevated hepatic NLRP3 expression relative to control mice (Figure S2D,E). Furthermore, co‐localization of NLRP3 and F4/80 was more frequent in hepatic sections of CUMS‐treated mice compared with control mice (Figure S3A). Previous studies have demonstrated that CD11b^+^Ly6C^hi^ macrophages, a subset of pro‐inflammatory cells, released higher amounts of NLRP3 and IL1B than CD11b^+^Ly6C^lo^ and CD11b^−^Ly6C^−^ cells [43]. This prompted us to investigate the infiltration of CD45^+^CD11b^+^Ly6C^hi^ macrophages in mice. Notably, CD45^+^CD11b^+^Ly6C^hi^ cells accumulated to a greater extent in the liver and spleen of CUMS‐treated mice than in control mice (Figures S2F and S3B).
MCC950, a highly specific NLRP3 inhibitor, was used to determine whether targeting NLRP3 could alleviate liver injury (Figure S2C). After ConA‐treated mice were subjected to CUMS intervention, serum ALT and AST levels and hepatic NLRP3 expression were significantly elevated, which could be alleviated by MCC950 treatment (Figures S2D,E and S3A,C). Significantly, histological analysis using hematoxylin and eosin (H&E) staining revealed severe portal inflammatory cell infiltration in the liver of ConA mice with CUMS intervention, which could be ameliorated by MCC950 (Figure S3D). Additionally, MCC950 treatment reduced the accumulation of CD45^+^CD11b^+^Ly6C^hi^ macrophages in the liver and spleen of ConA‐treated mice subjected to CUMS intervention (Figures S2F and S3B).
Depression Exacerbates the Disruption of the Intestinal Barrier Function in Immune‐Mediated Hepatitis
3.4
To investigate the role of depression in intestinal barrier dysfunction in patients with AIH, we measured serum biomarkers of intestinal permeability, including D‐lactic acid (DLA), diamine oxidase (DAO), fatty acid‐binding protein‐2 (FABP2), and lipopolysaccharide (LPS). Intestinal barrier dysfunction was more common in AIH patients with depression than in those without depression (Figure 2A).
Depression exacerbates the disruption of the intestinal barrier function in immune‐mediated hepatitis. (A) Serum levels of D‐lactic acid (DLA), diamine oxidase (DAO), fatty acid‐binding protein‐2 (FABP2), and lipopolysaccharide (LPS) were analyzed by enzyme‐linked immunosorbent assay (ELISA) in the control group (n = 8), AIH without depression group (n = 40), and AIH with depression group (n = 40). F = 13.430, KW = 20.266, 18.607, 22.166, all DF = 2. (B, C) The serum concentration of fluorescein isothiocyanate‐dextran (FITC‐D) in vivo permeability assay (B) and the serum concentration of LPS by ELISA (C). F = 9.639, 15.902, both DF = 3, both n = 6. (D) PCR analysis of tight junction protein 1 (Tjp1) or occludin (Ocln) relative to Gapdh in the intestine from the indicated groups. F = 20.463, 173.780, both DF = 3, both n = 4. (E) Western blot analysis of Zonula occludens protein 1 (ZO1) or OCLN relative to GAPDH in the intestine from the indicated groups. F = 28.575, 19.922, both DF = 3, both n = 4. (F) Representative immunofluorescence staining with occludin in the intestine from the indicated groups. Scale bars, 100 μm. The data for B–E are presented as mean ± SD. Statistical comparisons between groups were performed using one‐way ANOVA and the LSD post hoc test. The data for DLA are expressed as mean ± SD (analyzed by one‐way ANOVA with Tamhane's T2 post hoc test), whereas the data for DAO, FABP2, and LPS are presented as median (interquartile range [IQR]) (analyzed by nonparametric tests).
Compared with vehicle control mice, exposure to CUMS increased intestinal permeability, as reflected by increased levels of fluorescein isothiocyanate‐dextran (FITC‐D) and LPS in the serum and decreased levels of Zonula occludens protein 1 (ZO1) and occludin (OCLN) in the intestine (Figure 2B–E). Immunofluorescence staining of occludin further confirmed more severe intestinal barrier disruption in CUMS‐treated mice (Figure 2F). In particular, compared with mice treated with ConA alone, those with combined exposure to CUMS and ConA intervention exhibited more severe intestinal barrier disruption, as indicated by altered levels of FITC‐D, LPS, ZO1, and OCLN (Figure 2B–F). Overall, depression can exacerbate the disruption of the intestinal barrier function in immune‐mediated hepatitis.
Mirtazapine Exerts Protective Effects by Alleviating the Disruption of the Intestinal Barrier and the Overactivation of NLRP3 in Mice
3.5
The antidepressant mirtazapine was administered to investigate its effect on the destruction of the intestinal barrier integrity (Figure 3A). Mirtazapine alleviated intestinal permeability in ConA mice with CUMS intervention, as demonstrated by decreased levels of FITC‐D and LPS in the serum and increased levels of ZO1 and OCLN in the intestine (Figure 3B–E). Immunofluorescence staining of occludin further confirmed the protective role of mirtazapine in maintaining intestinal epithelial integrity (Figure S5A). As it has been demonstrated that LPS could cause intestinal barrier dysfunction in Caco‐2 epithelial cell monolayers [44, 45, 46], we next explored whether mirtazapine exerted a protective effect on LPS‐treated Caco‐2 cells in vitro. The results supported our hypothesis and demonstrated that mirtazapine increased the mRNA expression of TJP1 and OCLN in a concentration‐dependent manner, which further proved its protective effect on the intestinal barrier function (Figure 3F).
Mirtazapine exerts protective effects by alleviating the disruption of the intestinal barrier and the overactivation of NLR family pyrin domain containing 3 (NLRP3) inflammasome in mice. (A) Study design: CUMS‐induced depression mice were intraperitoneally injected with mirtazapine daily during the last 2 weeks. Two hours after the last injection of mirtazapine, mice were intravenously injected with ConA. The mice were sacrificed 16 h after ConA injection (yellow triangle). (B, C) Serum concentration of FITC‐D in vivo permeability assay (B) and the serum concentration of LPS by ELISA (C). (B) F = 7.216, DF = 4, n = 6. (C) F = 14.250, DF = 4, n = 6. (D) Western blots analysis of ZO1 or OCLN relative to GAPDH in the intestine from the indicated groups. F = 51.304, 328.339, both DF = 2, both n = 4. (E) PCR analysis of Tjp1 or Ocln relative to Gapdh in the intestine from the indicated groups. F = 19.112, 96.428, both DF = 4, both n = 4. (F) PCR analysis of TJP1 or OCLN relative to GAPDH in LPS‐treated Caco‐2 monolayers. F = 14.148, 28.119, both DF = 5, both n = 3. (G) The serum levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) in mice from the indicated groups. F = 110.511, 105.418, both DF = 4, both n = 6. (H) PCR analysis of Nlrp3, caspase‐1 (Casp1), and interleukin 1 beta (Il1b) relative to Gapdh in liver samples from the indicated groups. F = 39.049, 39.358, 36.442, all DF = 4, all n = 6. (I, J) The proportions of CD45+CD11b+Ly6Chi subsets among CD45+CD11b+ cells from the liver (I) and spleen (J) were quantified using flow cytometry analysis. (I) F = 67.774, DF = 4, n = 3. (J) F = 42.182, DF = 4, n = 3. All the values are shown as mean ± SD. Statistical significance was calculated using one‐way ANOVA with LSD post hoc test (B–F, H–J) and Tamhane's T2 post hoc test (G).
Next, we investigated whether mirtazapine participated in immune regulation. In ConA mice with CUMS intervention, mirtazapine reduced the levels of ALT and AST in the serum (Figure 3G) and the expression of NLRP3 in the liver (Figure S2E, Figure 3H, and Figure S5B). Additionally, flow cytometry analysis showed that CD45^+^CD11b^+^Ly6C^hi^ cells accumulated to a lower extent in the liver and spleen after mirtazapine administration (Figure 3I,J). Moreover, in the isolated murine peritoneal macrophages, LPS stimulation upregulated the expression of Nlrp3, Casp1, and Il1b, which was counteracted by mirtazapine pretreatment in a concentration‐dependent manner (Figure S5C). These research findings demonstrate that mirtazapine exerts protective effects by alleviating the disruption of the intestinal barrier function and the overactivation of the NLRP3 inflammasome.
Colonization With Intestinal Microbiota of Patients With MDD Disrupts Intestinal Epithelial Integrity and Overactivates Hepatic NLRP3 Inflammasome in Mice
3.6
Colonization with MDD microbiota resulted in depressive‐like behaviors in mice, manifested by weight loss, lower sucrose preference index, reduced total distance traveled, and decreased frequency of entering the central area, in contrast to those with control microbiota (Figure 4A and Figure S1B,E,H,K). Furthermore, when subjected to ConA injection, mice receiving gut microbiota transplantation from patients with MDD exhibited more severe liver damage than those harboring control microbiota (Figure S4A), as evidenced by elevated serum levels of ALT and AST (Figure S4B) and hepatic expression of NLRP3 (Figure 4B and Figure S4C). The co‐localization of F4/80 and NLRP3 further indicated the involvement of hepatic macrophages in liver injury (Figure S4D). Additionally, CD45^+^CD11b^+^Ly6C^hi^ cells accumulated to a greater extent in the liver and spleen of FMT‐MDD and ConA + FMT‐MDD mice compared with FMT‐HC and ConA + FMT‐HC mice (Figure 4C and Figure S4E).
Colonization with intestinal microbiota of patients with major depressive disorder (MDD) disrupts intestinal epithelial integrity and overactivates hepatic NLRP3 inflammasome in mice. (A) Study design: Seventy‐two hours after antibiotic discontinuation, mice received fecal microbiota transplantation (FMT) from patients with MDD or healthy controls (HC). The mice were sacrificed 16 h after ConA injection (yellow triangle). (B) PCR analysis of Nlrp3, Casp1, and Il1b relative to Gapdh in liver biopsies from the indicated groups. F = 37.843, 4.630, 23.158, all DF = 4, all n = 4. (C) The proportions of CD45+CD11b+Ly6Chi cells among CD45+CD11b+ cells in the liver were quantified by flow cytometry analysis. F = 46.149, DF = 4, n = 3. (D, E) The serum concentration of FITC‐D in vivo permeability assay (D) and the serum concentration of LPS by ELISA (E). (D) F = 10.499, DF = 4, n = 6. (E) F = 27.756, DF = 4, n = 6. (F) PCR analysis of Tjp1 or Ocln relative to Gapdh in the intestine from the indicated groups. F = 29.744, 7.447, both DF = 4, both n = 3. (G) Western blot analysis of ZO1 or OCLN relative to GAPDH in the intestine from the indicated groups. F = 38.268, 21.866, both DF = 3, both n = 3. (H) Gut‐derived Lactococcus formosensis was successfully isolated from the liver of FMT‐MDD mice. (I) 16S rRNA gene sequencing of intestinal microbiota from the indicated groups. All the values are presented as mean ± SD. Statistical difference was calculated by one‐way ANOVA with LSD post hoc test.
Next, we demonstrated that transplantation of MDD microbiota notably increased intestinal permeability and further aggravated endotoxemia (Figure 4D,E). Mice colonized with MDD microbiota exhibited decreased levels of ZO1 and OCLN in the intestine compared with those colonized with control microbiota (Figure 4F,G). To identify the presence of bacteria in the extraintestinal organs, the liver, spleen, and mesenteric lymph nodes (MLN) from mice colonized with microbiota from patients with MDD (n = 4) were aseptically collected and cultured. Of these, bacterial clones were isolated from the livers of 3 out of 4 mice when cultured on Columbia blood agar plates under aerobic or anaerobic conditions. However, no bacterial clones could be grown from the spleen or MLN of mice receiving FMT from MDD patients. Similarly, no bacterial growth was observed in any organs from the control mice or mice receiving FMT from healthy controls. The bacteria were subsequently identified as Lactococcus formosensis using 16S rRNA gene sequencing (Figure 4H and Figure S6D). Simultaneously, we confirmed that the abundance of L. formosensis was higher in stool samples of mice receiving FMT from patients with MDD than in those receiving FMT from healthy controls (Figure 4I). Hence, these data collectively demonstrate that colonization with MDD microbiota can disrupt intestinal epithelial integrity and promote the activation of hepatic NLRP3 inflammasome in mice.
Translocation of Gut‐Derived
L. formosensis Overactivates the Hepatic NLRP3 Inflammasome and Induces Liver Injury in Mice
3.7
The growth and genetic characteristics of L. formosensis were summarized in Figure 5A and Figure S6A–D. We found that mice monocolonized with L. formosensis exhibited depressive‐like behaviors (e.g., weight loss, lower sucrose preference index, reduced total distance traveled, and decreased frequency of entering the central area) (Figure S1C,F,I,L). Histological analysis using H&E staining revealed severe portal inflammatory cell infiltration in the liver of mice monocolonized with L. formosensis , which was ameliorated by a specific vaccine targeting L. formosensis (Figure 5B–D). In addition, mice monocolonized with L. formosensis exhibited disrupted intestinal epithelial integrity, manifested by elevated serum levels of FITC‐D and LPS. Notably, the elevated levels of FITC‐D and LPS were counteracted by vaccination (Figure 5E,F). Consistently, mice monocolonized with L. formosensis showed bacterial translocation from the intestine to the liver and MLN (Figure S6D), as confirmed by green fluorescent protein (GFP)‐labeled bacteria (Figure 5G). Vaccination prevented bacterial translocation, as no growth of L. formosensis was observed in any organ of L. formosensis monocolonized mice pretreated with the vaccine.
Translocation of gut‐derived L. formosensis overactivates the hepatic NLRP3 inflammasome and induces liver injury in mice. (A) The growth curve of L. formosensis was determined by optical density (OD) measurements at 600 nm. (B) Study design: Seventy‐two hours after antibiotic discontinuation, mice were administered L. formosensis once a week during the course of 8 weeks by oral gavage. Before the first gavage, the vaccine targeting L. formosensis ‐specific DNA was intramuscularly injected once a week during the course of 2 weeks. The evaluation of depressive‐like behaviors was conducted 24 h after the last administration. The yellow triangle represents the point of analysis. (C) Representative hematoxylin and eosin (H&E) staining images of liver tissues were shown (Scale bars: 100 μm, 50 μm). (D) The severity of inflammatory cell infiltration was graded using the 4‐point scoring system. KW = 19.677, DF = 3, n = 6. (E, F) The concentration of FITC‐D in vivo permeability assay (E) and the serum concentration of LPS by ELISA (F). (E) F = 9.190, DF = 3, n = 6. (F) F = 15.299, DF = 3, n = 6. (G) The translocation of green fluorescent protein (GFP)‐labeled L. formosensis to liver (left) or mesenteric lymph nodes (MLN) (right). Fluorescence in the liver or MLN indicated bacterial translocation. (H) The serum levels of ALT and AST in mice from the indicated groups. F = 4.516, 10.612, both DF = 3, both n = 6. (I) Western blot analysis of NLRP3 relative to GAPDH in the liver from the indicated groups. F = 3.984, DF = 3, n = 4. (J) PCR analysis of Nlrp3, Casp1, and Il1b relative to Gapdh in the liver from the indicated groups. F = 28.603, 12.039, 18.164, all DF = 3, all n = 4. (K) PCR analysis of Nlrp3, Casp1, and Il1b relative to Gapdh in the isolated murine peritoneal macrophages treated with the supernatant of L. formosensis in a concentration‐dependent manner (1:100, 1:50, 1:25). F = 34.235, 52.209, 42.390, all DF = 4, all n = 3. The data for E–F, H–K are presented as mean ± SD. Statistical significance was calculated by one‐way ANOVA with LSD post hoc test (E, H–K) and Tamhane's T2 post hoc test (F). The data for D are presented as median (IQR) and were analyzed using nonparametric tests.
Monocolonization with L. formosensis elevated serum levels of ALT and AST (Figure 5H) and hepatic expression of NLRP3 (Figure 5I,J, Figure S6E), which could also be alleviated by the specific vaccine against L. formosensis . In the isolated murine peritoneal macrophages, the supernatant of L. formosensis upregulated the expression of Nlrp3, Casp1, and Il1b in a concentration‐dependent manner (Figure 5K). In conclusion, monocolonization with L. formosensis is sufficient to disrupt intestinal epithelial integrity and lead to subsequent bacterial translocation to the liver, which can amplify NLRP3‐mediated liver injury.
Discussion
4
This study demonstrated that depression exacerbated immune‐mediated hepatitis through the gut‐liver axis. Clinically, our findings highlighted a high prevalence of depression in patients with AIH and further demonstrated its significant association with an increased risk of cirrhosis and intestinal barrier disruption. Mechanistically, we revealed that the detrimental effects of depression were mediated by intestinal barrier dysfunction and NLRP3 overactivation. The translocation of gut‐derived L. formosensis to the liver played pivotal roles in the overactivation of the hepatic NLRP3 inflammasome and consequent liver injury.
Previous studies have demonstrated a high prevalence of depression in patients with AIH [2, 47]. Depression, a key factor responsible for nonadherence to standard treatment, increases the risk of relapse in patients with AIH [5]. Moreover, depression is related to adverse clinical outcomes in patients with chronic liver disease [48] and is recognized as an important risk factor for liver fibrosis [6, 7, 8]. Consistent with previous studies, we observed that depression was common in patients with AIH (40.8%) and correlated with disease severity. Additionally, depression was an independent risk factor for cirrhosis in patients with AIH. We validated the above clinical results in a mouse model. Repeated ConA injections increased susceptibility to liver fibrosis in mice [49]. Based on this liver fibrosis model, we demonstrated that CUMS intervention promoted fibrosis progression in mice repeatedly injected with ConA. These results highlight the clinical significance of identifying and managing depression in AIH.
Patients with depression commonly exhibit systemic inflammation with elevated levels of pro‐inflammatory cytokines in plasma [50]. The NLRP3 inflammasome, involved in CASP1 activation, exerts a crucial effect on depression [15, 16, 17]. In the livers of ConA‐induced hepatitis mice, the expression levels of NLRP3, CASP1, and IL1B are significantly upregulated [25]. The NLRP3 inflammasome activation also participates in the liver inflammation of other forms of immune‐mediated hepatitis [51, 52]. Consistent with previous findings, we demonstrated upregulated expression of Nlrp3, Casp1, and Il1b in the livers of CUMS‐treated mice. Furthermore, CUMS intervention upregulated the expression of hepatic Nlrp3, Casp1, and Il1b in ConA‐treated mice, which was attenuated by the highly specific NLRP3 inhibitor MCC950. In patients with AIH, elevated expression levels of NLRP3, CASP1, and IL1B were more commonly detected in the liver biopsies from depressed AIH patients compared with nondepressed patients. Taken together, these data imply that depression may promote the activation of NLRP3 and subsequent disease progression in immune‐mediated hepatitis.
Gut‐derived systemic inflammation acts as a driver of depression in chronic liver disease [53]. Gut dysbiosis has been identified as a shared pathogenic mechanism in AIH [26, 27] and depression [30, 31, 32]. A recent study suggested that gut‐derived Enterococcus gallinarum could translocate to the liver through the disrupted intestinal barrier, inducing autoimmunity in patients and mouse models of AIH [54]. Another study demonstrated that gut‐derived Klebsiella pneumonia promoted intestinal barrier dysfunction through epithelial pore formation, leading to bacterial translocation and subsequent hepatic inflammation in primary sclerosing cholangitis [55]. In depression, gut dysbiosis also causes impairment of intestinal mucosal permeability and culminates in increased bacterial translocation from the intestine to extraintestinal organs [33, 56]. In this study, we isolated L. formosensis from the livers of mice colonized with the microbiota of patients with MDD. Furthermore, disruption of the intestinal barrier integrity and overactivation of the hepatic NLRP3 were observed in mice receiving FMT from patients with MDD and in mice monocolonized with L. formosensis . Therefore, our data provide evidence for a mechanism through which depression aggravates immune‐mediated hepatitis, which involves hepatic NLRP3 inflammasome overactivation induced by depression‐associated intestinal dysbiosis.
In our study, mirtazapine, a competitive inhibitor of serotonin type 2 and type 3 receptors, was used to investigate its effect on immune‐mediated hepatitis. Patients with depression are more likely to develop ulcerative colitis. Interestingly, mirtazapine protects against ulcerative colitis by alleviating behavioral abnormalities and modulating neuronal excitability in the brain [57]. In primary biliary cholangitis, mirtazapine protects against a poor prognosis by decreasing the incidence of cirrhosis and mortality [58]. In mouse models, mirtazapine attenuates ConA‐induced hepatic injury in a concentration‐dependent manner [59]. In this study, we verified that mirtazapine conferred protective effects by alleviating the impairment of intestinal barrier integrity and abnormal activation of the NLRP3 inflammasome in ConA‐treated mice with CUMS intervention. Therefore, investigating the therapeutic potential of mirtazapine in patients with AIH, particularly those with comorbid depression, is of vital clinical importance.
This study presented several key innovations. Clinically, it highlighted the necessity of considering depressive symptoms in patients with AIH, given their association with an increased risk of cirrhosis and disease progression. Mechanistically, we verified that depression exacerbated immune‐mediated hepatitis by impairing the intestinal barrier and overactivating the hepatic NLRP3 inflammasome. Critically, gut dysbiosis played a pivotal role in depression‐related liver injury. Our study is the first to successfully isolate gut‐derived L. formosensis from the livers of mice receiving FMT from patients with MDD, and to elucidate the pivotal role of L. formosensis in immune regulation. Furthermore, a vaccine targeting L. formosensis was shown to prevent bacterial translocation and alleviate liver injury in mice, demonstrating therapeutic potential. These findings pave the way for considering mirtazapine as a promising therapeutic option for AIH.
However, several limitations of this study must be acknowledged. First, it was a single‐center investigation, which necessitates further validation in multicenter cohorts. Second, the lack of data on the clinical efficacy of mirtazapine limits the clinical applicability of our findings. Additionally, the clinical relevance of L. formosensis translocation, including its prevalence in human patients and its contribution to disease progression, remains to be fully established. Finally, it was difficult to determine causality between depression and the observed results. From the perspective of neuroimmune circuits, the frequent comorbidity of depression in patients with AIH may be ascribed to the interaction between the brain and the immune system. Therefore, future studies should explore the interaction between depression and AIH from different perspectives.
Conclusions
5
In conclusion, depression, a common mental disorder in patients with AIH, contributed to the progression of fibrosis. As graphically summarized (Figure 6), liver inflammation driven by depression was associated with intestinal barrier dysfunction and NLRP3 overactivation in liver macrophages. L. formosensis , isolated from the livers of mice colonized with microbiota from patients with depression, played pivotal roles in the disruption of intestinal epithelial integrity and overactivation of NLRP3 in the liver. Antidepressants and vaccines against L. formosensis may be strategies for preventing and treating patients with AIH, particularly those with depression.
Illustration depicting the role of depression and depression‐associated gut microbiota in immune‐mediated hepatitis. Depression is common in patients with AIH and is a risk factor for cirrhosis. AIH patients with depression, mice colonized with microbiota from patients with MDD, and ConA‐treated mice with CUMS intervention exhibit intestinal barrier dysfunction and hepatic NLRP3 inflammasome overactivation. Gut‐derived L. formosensis , isolated from the livers of mice colonized with microbiota from patients with MDD, plays pivotal roles in the disruption of intestinal epithelial integrity and overactivation of hepatic NLRP3 inflammasome. Antidepressants mirtazapine and vaccines against L. formosensis can alleviate liver injury in mice.
Author Contributions
Simin Zhou: conceptualization, data curation, formal analysis, writing – original draft. Liping Guo: data curation, investigation, visualization. Nian Chen: formal analysis, software, visualization. Haifeng Liu: data curation, writing – review and editing. Xin Liu: conceptualization, writing – original draft. Jiwen Li: conceptualization. Shijing Dong: methodology. Jiangpeng Liu: validation. Xiaoyi Wang: methodology. Ying Ran: methodology. Man Liu: conceptualization. Hongyu Chu: data curation. Yanni Li: methodology. Hui Yang: validation. Jingwen Zhao: conceptualization, project administration, supervision, visualization. Lu Zhou: funding acquisition, resources, writing – original draft.
Funding
This work was supported by National Natural Science Foundation of China (81860109) and Natural Science Foundation of Tianjin Municipality (21JCZDJC00880).
Disclosure
The authors have nothing to report.
Ethics Statement
This study involving human participants was approved by the Tianjin Medical University Ethics Committee, based on the ethical guidelines of the Declaration of Helsinki (revised 2013, Fortaleza, Brazil) (Ethical Approval No. IRB2021‐WZ‐191). The animal experiment was approved by the Ethics and Welfare Committee of Animals at Tianjin Medical University (Ethical No. IRB2023‐DWFL‐257).
Consent
Informed consent was obtained from all participants.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Data S1: The Data S1 includes Supporting Informations and Methods, Supplementary Figures, and Supplementary Tables. Figure S1: Assessment of depressive‐like behaviors in mice. Figure S2: Depression promotes the activation of NLRP3 in immune‐mediated hepatitis. Figure S3: A highly specific NLRP3 inhibitor MCC950 exerts protective effects in mice. Figure S4: Fecal microbiota transplantation from patients with MDD induces adverse effects in recipient mice. Figure S5: Mirtazapine exerts protective effects by alleviating the disruption of intestinal barrier and the overactivation of NLRP3 in mice. Figure S6: Translocation of gut‐derived L. formosensis drives hepatic NLRP3 overactivation and liver injury in mice. Table S1: Comparison of general characteristics and laboratory parameters between patients with AIH and healthy controls. Table S2: Comparison of general characteristics and laboratory parameters between AIH patients with and without depression. Table S3: Comparison of general characteristics and laboratory parameters between AIH patients with and without cirrhosis. Table S4: Clinical characteristics of participants included in the PCR‐based analysis of hepatic NLRP3 pathway expression.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1C. D. Slooter , F. F. van den Brand , A. Lleo , et al., “Lack of Complete Biochemical Response in Autoimmune Hepatitis Leads to Adverse Outcome: First Report of the IAIHG Retrospective Registry,” Hepatology 79, no. 3 (2024): 538–550.37676683 10.1097/HEP.0000000000000589 · doi ↗ · pubmed ↗
- 2C. Schramm , I. Wahl , C. Weiler‐Normann , et al., “Health‐Related Quality of Life, Depression, and Anxiety in Patients With Autoimmune Hepatitis,” Journal of Hepatology 60, no. 3 (2014): 618–624.24240053 10.1016/j.jhep.2013.10.035 · doi ↗ · pubmed ↗
- 3L. Yang , B. Gu , X. Wang , Q. Ren , M. Shen , and D. Su , “Association of Disease Activity With Depression and Anxiety in Systemic Lupus Erythematosus: A Comparison of SLEDAI‐2K and SLE‐DAS,” Rheumatology (Oxford) 64, no. 2 (2025): 632–638.38305645 10.1093/rheumatology/keae 070 · doi ↗ · pubmed ↗
- 4Y. Qian , Y. Chen , L. Liu , T. Wu , X. Chen , and G. Ma , “Depression and Anxiety in Inflammatory Bowel Disease: Mechanisms and Emerging Therapeutics Targeting the Microbiota‐Gut‐Brain Axis,” Frontiers in Immunology 16 (2025): 1676160.41280932 10.3389/fimmu.2025.1676160 PMC 12634526 · doi ↗ · pubmed ↗
- 5S. Sockalingam , D. Blank , N. Abdelhamid , S. E. Abbey , and G. M. Hirschfield , “Identifying Opportunities to Improve Management of Autoimmune Hepatitis: Evaluation of Drug Adherence and Psychosocial Factors,” Journal of Hepatology 57, no. 6 (2012): 1299–1304.22871503 10.1016/j.jhep.2012.07.032 · doi ↗ · pubmed ↗
- 6D. Kim , B. B. Dennis , G. Cholankeril , and A. Ahmed , “Association Between Depression and Metabolic Dysfunction‐Associated Fatty Liver Disease/Significant Fibrosis,” Journal of Affective Disorders 329 (2023): 184–191.36841305 10.1016/j.jad.2023.02.101 · doi ↗ · pubmed ↗
- 7A. B. Nasir , S. Zouridis , P. Aspichueta , P. Manka , and W. K. Syn , “Relationship Between Depression and Chronic Liver Disease: Potential Role of Antidepressants in Modulating Liver Fibrosis,” American Journal of the Medical Sciences (2025): S 0002‐9629(25)01214‐5, 10.1016/j.amjms.2025.07.018.41043608 · doi ↗ · pubmed ↗
- 8S. M. Zhou , J. W. Li , J. P. Liu , et al., “Depressive Symptom as a Risk Factor for Cirrhosis in Patients With Primary Biliary Cholangitis: Analysis Based on Lasso‐Logistic Regression and Decision Tree Models,” Brain and Behavior 14, no. 8 (2024): e 3639.39099389 10.1002/brb 3.3639 PMC 11298689 · doi ↗ · pubmed ↗