dock11 Knockdown in Zebrafish Disrupts Embryogenesis: Insights Into the Genetic Causes of Early Pregnancy Loss
Chang Liu, Meng Wang, Feng Chen, Mei Chen, Yonghua Yao, Wei Huang

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
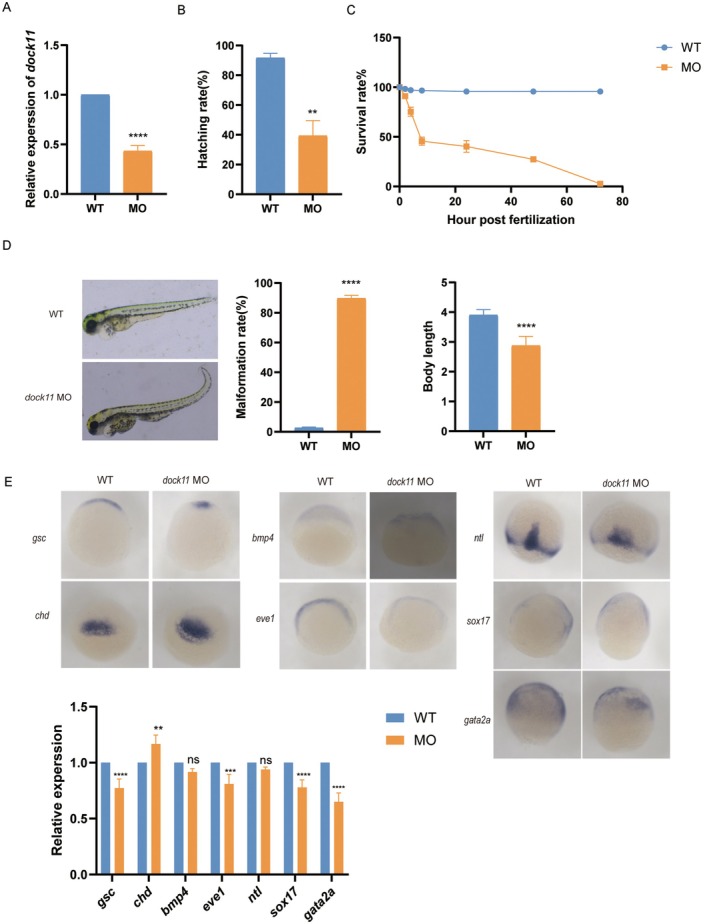 FIGURE 1
FIGURE 1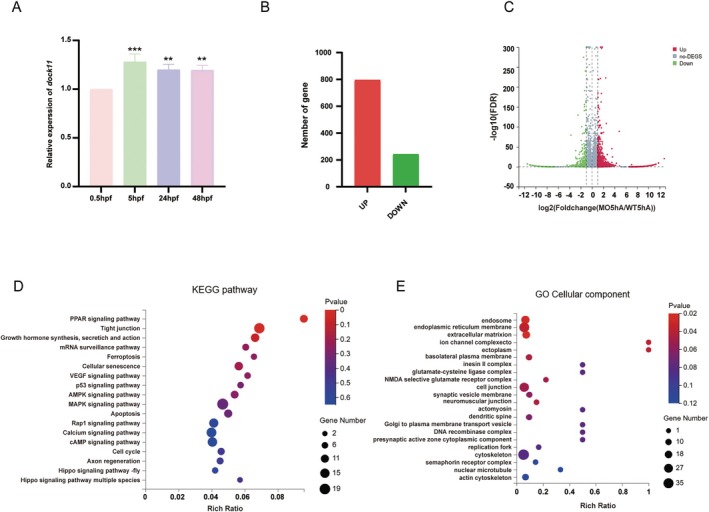 FIGURE 2
FIGURE 2- —National Key Research and Development Program of China10.13039/501100012166
- —General Program of Sichuan Provincial Natural Science Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsReproductive System and Pregnancy · Prenatal Screening and Diagnostics · Zebrafish Biomedical Research Applications
Dear Editor,
Recurrent pregnancy loss (RPL), affecting approximately 5% of couples worldwide, represents a major challenge in reproductive medicine and causes psychological distress [1]. While embryonic chromosomal errors account for 40%–65% of early pregnancy losses, a substantial proportion of cases remain unexplained despite extensive clinical evaluation [2]. This diagnostic gap is further highlighted by the observation that pregnancy losses still occur even after the transfer of euploid embryos following preimplantation genetic testing for aneuploidy (PGT‐A) in assisted reproduction [3]. This clinical dilemma underscores a critical gap in our understanding of the molecular pathogenesis of early pregnancy loss, particularly the role of embryonic‐intrinsic factors [2, 3, 4]. While existing research has largely centered on deficits in implantation and placental development, the critical window of early embryogenesis—a period governed by the embryo's autonomous developmental program and fundamental to embryonic survival—has received comparatively less attention [5, 6, 7].
Our previous multi‐omics analysis of chorionic villi from euploid pregnancy‐loss patients revealed epigenetic silencing of DOCK11 (dedicator of cytokinesis 11) and its consequent transcriptional downregulation in extra‐embryonic tissues, implicating DOCK11 as a potential contributor to pregnancy failure (our unpublished data). This finding prompted us to investigate the potential intrinsic role of DOCK11 within the embryo proper.
To functionally validate the role of DOCK11 in early embryogenesis, we turned to the zebrafish model. This model is uniquely suited for such an investigation, as its external development and optical transparency enable direct visualization of embryogenesis while being free from the confounding influences of the maternal uterine environment and placental function. Morpholino (MO)‐mediated knockdown of dock11 was confirmed via a significant reduction in its mRNA levels (Figure 1A). Dock11‐knockdown embryos exhibited markedly compromised viability, with significantly reduced hatching rates and elevated embryonic mortality compared to wild‐type (WT) controls (Figure 1B,C). Detailed morphological assessment revealed a spectrum of severe developmental defects, including pronounced axial curvature, a high incidence of malformations, and reduced overall body length (Figure 1D). To determine the impact on early patterning, we further performed whole‐mount in situ hybridization. Although the spatial domains of key lineage markers—including gsc and chd (dorsal mesoderm, assessed at 5 hpf), bmp4 and eve1 (ventral mesoderm, 5 hpf), ntl (axial mesoderm, assessed 8 hpf), sox17 (endoderm, 8 hpf), and gata2a (ectoderm, 8 hpf) —remained largely unaltered in dock11 MO embryos compared to WT embryos, their expression levels were markedly altered: the expression of chd was significantly increased, while that of gsc, eve1, sox17, and gata2a was decreased. Changes in bmp4 and ntl were not significant (Figure 1E). Collectively, these findings establish that dock11 is essential for normal embryogenesis and germ layer formation in zebrafish.
*dock11 affects development of zebrafish embryos. (A) The expression of dock11 was determined by RT‐qPCR in wild‐type (WT) and morpholino (MO). mRNA levels were normalised to gapdh and are expressed relative to the WT control (n = 3). (B) Column chart shows the hatching rate of zebrafish. (C) Line chart represents the survival curve of zebrafish, with circular dotted line representing WT and square solid line indicating MO (n = 50). (D) Column graphs show the malformation rate (p < 0.0001) and body length (p < 0.0001) of zebrafish, respectively (n = 50). (E) Whole‐mount in situ hybridization of germ layer markers in WT and dock11 MO zebrafish embryos. Column charts display the relative expression levels quantified from the signal intensity. Genes assessed at 5 hpf include gsc, chd, bmp4, and eve1; those assessed at 8 hpf include nt1, sox17, and gata2a. Data are presented as mean ± SD. *p < 0.05; **p < 0.01; ***p < 0.001, ***p < 0.0001.
Having established that the loss of dock11 disrupts early embryonic development in zebrafish, we next sought to delineate the underlying molecular mechanisms. We first characterised the spatiotemporal expression profile of dock11, finding its mRNA to be dynamically expressed, with a pronounced peak at 5 h post‐fertilisation (hpf, Figure 2A)—a stage immediately preceding the onset of gastrulation that is critical for establishing the molecular patterns underlying subsequent cell fate determination. We therefore conducted transcriptomic profiling at this critical stage. RNA‐seq analysis revealed extensive dysregulation in dock11 morphants, with 796 upregulated and 243 downregulated (Figure 2B,C, Table S1). Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis of these differentially expressed genes revealed enrichment in pathways related to embryonic development, including VEGF signalling, tight junction assembly, Hippo and MAPK signalling pathways (Figure 2D, Table S2). Furthermore, pathways governing cell fate, such as apoptosis, p53 signalling, ferroptosis, and cellular senescence, were prominently enriched, as were metabolic regulators like PPAR and AMPK signalling, and mRNA surveillance mechanisms (Figure 2D, Table S2). Gene ontology (GO) analysis of cellular component highlighted structures essential for tissue integrity and morphogenesis, including the extracellular matrix (ECM), endoplasmic reticulum membrane, ion‐channel complexes, the basolateral plasma membrane, and cell–cell junctions. Enrichment was also observed for cytoskeletal elements and vesicle‐related complexes (Figure 2E, Table S3). These results strongly suggest that dock11 regulates a broad transcriptional network essential for cell adhesion, intra‐cellular signalling, and the maintenance of lineage integrity during early embryogenesis.
*dock11 regulates early transcriptional programs in zebrafish embryogenesis. (A) Column graphs show the mRNA expression levels of dock11 in zebrafish embryos at 0.5 hpf, 5 hpf, 24 hpf, and 48 hpf (n = 3). (B) Column graphs show the number of DEGs related to dock11‐knockdown at 5 hpf zebrafish embryos. (C) Volcano plot shows the DEGs associated with dock11 mRNA downregulation in the MO group compared with the WT group. (D) The bubble chart shows the representative KEGG pathways of DEGs associated with dock11 mRNA downregulation. The p‐value was determined using a hypergeometric test. (E) Bubble chart shows the representative biological process GO terms enriched by DEGs related to the dock11 mRNA downregulation. The p‐value was determined using a hypergeometric test. *p < 0.05; **p < 0.01; **p < 0.001.
It is noteworthy that while previous studies using dock11 knockout models in mice and zebrafish reported no embryonic lethality but rather focused on immune and hematopoietic phenotypes [8, 9], our study focuses on the role of dock11 during early embryogenesis.
In conclusion, our integrated findings suggest the importance of embryonic‐intrinsic factors in pregnancy loss. We demonstrate that dock11 is essential for body patterning and germ layer specification in the early vertebrate embryo. This work thus expands our understanding of the genetic aetiology of early pregnancy loss and establishes DOCK11 dysfunction as a previously unappreciated mechanistic contributor to the aetiology of early pregnancy loss.
Author Contributions
Wei Huang: Conceptualised, designed, and supervised the research, revised the manuscript, and handled the submission. Chang Liu and Meng Wang: Performed the experiments, collected and analysed the data, and drafted the original manuscript. Feng Chen, Mei Chen, and Yonghua Yao: Assisted with experiments.
Funding
This work was supported by National Key Research and Development Program of China, 2023YFC2705502. Natural Science Foundation of Sichuan province, 2025ZNSFSC0742. We acknowledge support from the National Key R&D Program of China (2023YFC2705502) and the General Program of Sichuan Provincial Natural Science Foundation (2025ZNSFSC0742).
Ethics Statement
The study was approved by the Ethics Committee of West China Second University Hospital of Sichuan University (Approval No. 2025–51) and was approved by the Ethics Committee of Animal Experiments of Sichuan University (Approval No. 2025214). All participants provided written informed consent prior to their inclusion in the study.
Consent
All authors reviewed the final manuscript and consented to its publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Table S1: List of Differentially Expressed Genes from RNA‐seq Analysis of dock11 Morphants.
Table S2: List of KEGG Pathway Enrichment Analysis for Differentially Expressed Genes from RNA‐seq of dock11 Morphants.
Table S3: List of GO Enrichment Analysis for Differentially Expressed Genes from RNA‐seq of dock11 Morphants.
Table S4: Sequences of primers used for quantitative real‐time PCR.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Obstetrics Subgroup, Chinese Society of Obstetrics and Gynecology; Chinese Medical Association; Chinese Expert Consensus Group on Diagnosis and Management of Recurrent Spontaneous Abortion , “Chinese Expert Consensus on Diagnosis and Management of Recurrent Spontaneous Abortion,” Zhonghua Fu Chan Ke Za Zhi 57, no. 9 (2022): 653–667, 10.3760/cma.j.cn 112141-20220421-00259.36177576 · doi ↗ · pubmed ↗
- 2K. Sonehara , Y. Yano , T. Naito , et al., “Common and Rare Genetic Variants Predisposing Females to Unexplained Recurrent Pregnancy Loss,” Nature Communications 15, no. 1 (2024): 5744, 10.1038/s 41467-024-49993-5.PMC 1125529639019884 · doi ↗ · pubmed ↗
- 3X. Dong , J. Shi , X. Liu , et al., “Risk Factors Associated With Pregnancy Loss After Single Euploid Blastocysts Transfer,” Frontiers in Endocrinology 15 (2025): 1461088, 10.3389/fendo.2024.1461088.39944231 PMC 11813741 · doi ↗ · pubmed ↗
- 4A. L. Wilkinson , I. Zorzan , and P. J. Rugg‐Gunn , “Epigenetic Regulation of Early Human Embryo Development,” Cell Stem Cell 30, no. 12 (2023): 1569–1584, 10.1016/j.stem.2023.09.010.37858333 · doi ↗ · pubmed ↗
- 5Q. Zhu , Y. Liu , X. Hao , et al., “Dissecting Pre‐ To Post‐Implantation Transition of DNA Methylome‐Transcriptome Dynamics in Early Mammalian Development,” Cell Reports 44 (2025): 115790, 10.1016/j.celrep.2025.115790.40471785 · doi ↗ · pubmed ↗
- 6W. Peng , Y. Chen , X. Luo , et al., “DNA Methylation‐Associated Repression of MEST/PEG 1 Expression Contributes to the Invasion of Extravillous Trophoblast Cells,” Placenta 46 (2016): 92–101, 10.1016/j.placenta.2016.08.093.27697227 · doi ↗ · pubmed ↗
- 7A. L. Mattei , N. Bailly , and A. Meissner , “DNA Methylation: A Historical Perspective,” Trends in Genetics 38 (2022): 676–707, 10.1016/j.tig.2022.03.010.35504755 · doi ↗ · pubmed ↗
- 8T. Matsuda , S. Yanase , A. Takaoka , and M. Maruyama , “The Immunosenescence‐Related Gene Zizimin 2 Is Associated With Early Bone Marrow B Cell Development and Marginal Zone B Cell Formation,” Immunity & Ageing 12 (2015): 1, 10.1186/s 12979-015-0028-x.25729399 PMC 4343071 · doi ↗ · pubmed ↗