Integration of light quality signals regulates ABA abundance and stomatal movements during seedling establishment
Mathilda Gustavsson, Lionel Hill, Keara A. Franklin, Ashley J. Pridgeon

TL;DR
Seedlings adjust stomatal opening based on light quality, balancing growth and water use through ABA regulation.
Contribution
Discovery of how light quality signals converge to regulate ABA and stomatal movements in seedlings.
Findings
Low R:FR light inhibits stomatal opening via PIF4 and increased ABA.
UV-B promotes stomatal opening through UVR8 and phototropin photoreceptors.
Phytochrome and UVR8 signaling converge to control ABA and coordinate growth and water use.
Abstract
Obtaining sufficient light for photosynthesis and avoiding desiccation are two key challenges faced by seedlings during early establishment. Perception of light quality via specialised photoreceptors signals the availability of sunlight for photosynthesis. Canopy shade is depleted in red (R) and enriched in far‐red (FR) light, lowering R : FR ratio, while direct sunlight and sunflecks contain UV‐B. The balance between these wavelengths can determine the developmental strategy adopted by seedlings to either avoid shade, via stem elongation, or promote the expansion of photosynthetic organs. How seedlings regulate stomatal movements in different light environments is poorly understood.Using FR and UV‐B supplementation to mimic aspects of canopy shade and sunlight, respectively, we monitored stomatal apertures in Arabidopsis thaliana cotyledons and gas exchange in the cotyledons of Chinese…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
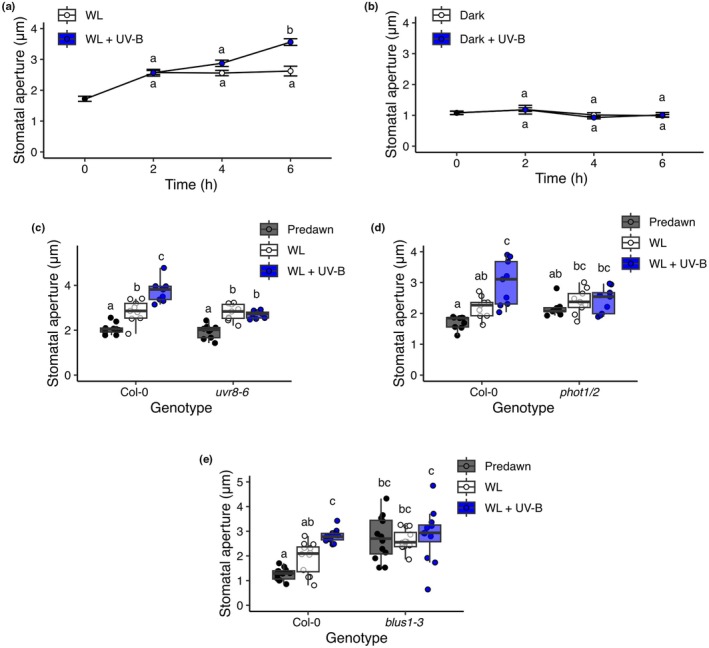 Fig. 1
Fig. 1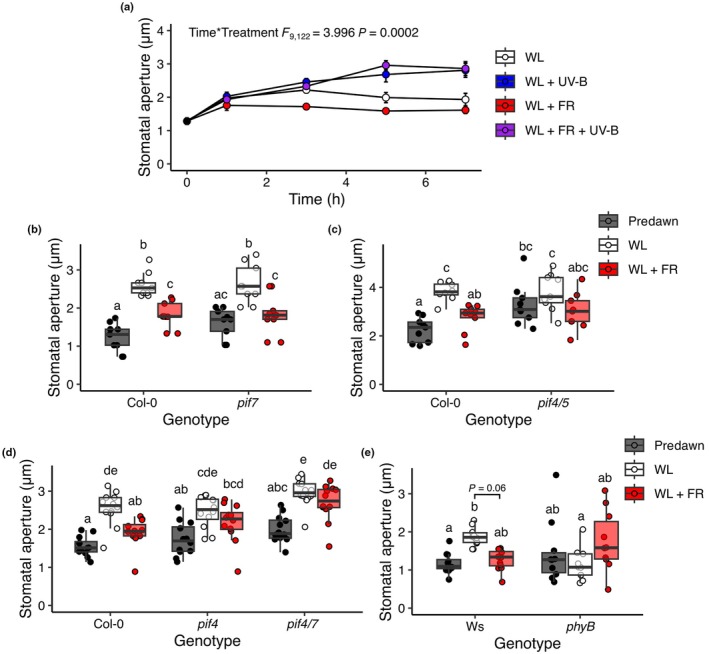 Fig. 2
Fig. 2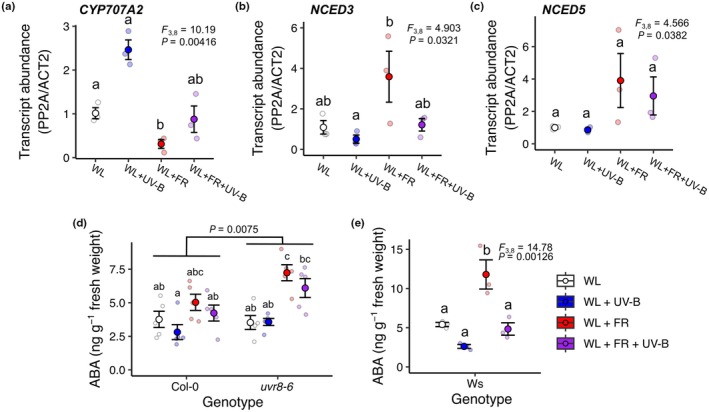 Fig. 3
Fig. 3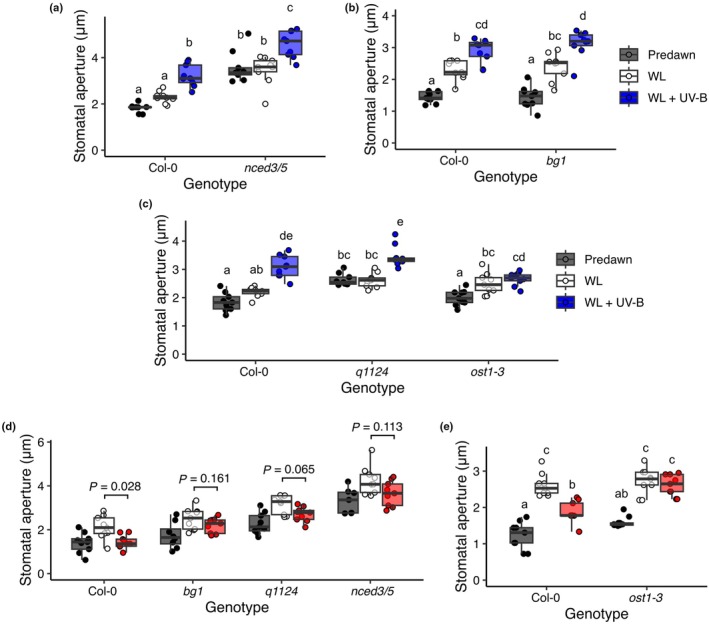 Fig. 4
Fig. 4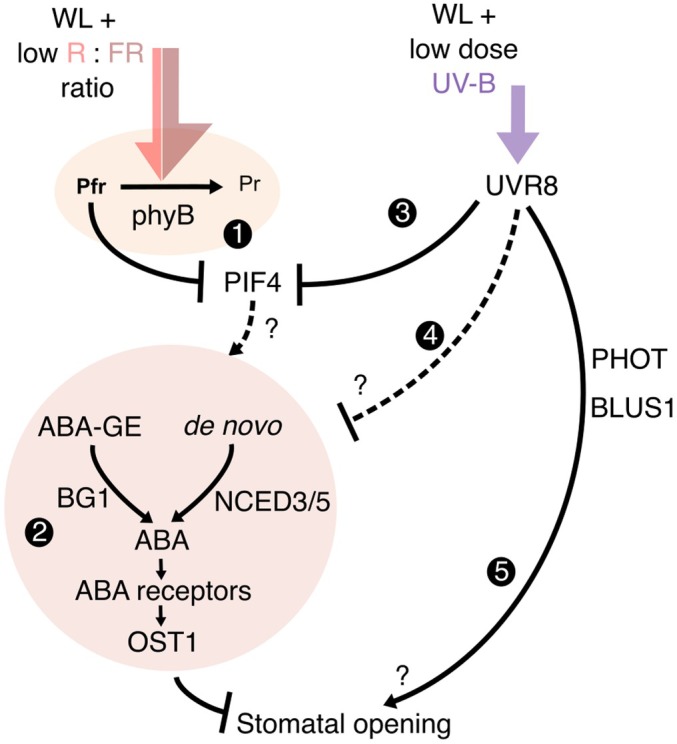 Fig. 5
Fig. 5- —Leverhulme Trust10.13039/501100000275
- —Biotechnology and Biological Sciences Research Council10.13039/501100000268
- —Bristol Centre for Agricultural Innovation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLight effects on plants · Plant Gene Expression Analysis · Photosynthetic Processes and Mechanisms
Introduction
Plants have evolved a suite of photoreceptors to detect light quality and quantity, signalling the availability of sunlight to fuel photosynthesis. Seedlings emerging under canopy shade are exposed to increased amounts of FR light, reflected and transmitted from vegetative tissue. This lowers the ratio of red to far‐red light (R : FR), inactivating phytochrome photoreceptors and driving stem elongation to overtop competitors (Ballare et al., 1990). Direct sunlight and sunflecks contain UV‐B light, which antagonises shade avoidance responses (Hayes et al., 2014; Sharma et al., 2019; Tavridou et al., 2020a), promoting a compact stature (Moriconi et al., 2018). Continuous monitoring of light quality changes, therefore, enables plants to optimise developmental strategy in fluctuating environments (Fernández‐Milmanda & Ballaré, 2021).
Mortality rates are high during early seedling establishment (Leck et al., 2008), with water scarcity presenting a significant stress (Fenner & Thompson, 2005). Stomata are microscopic pores found predominantly on the epidermis of leaves. They perform a crucial role in the prevention of water loss and the uptake of carbon through the regulation of gas exchange. This can involve short‐term alterations in the size of the stomatal pore and longer term alterations in stomatal development, changing the number of stomatal pores on a plant's surface (Lawson & Matthews, 2020). Stomata are sensitive to a range of exogenous and endogenous signals (including light, temperature, CO_2_ concentration, water availability, pathogen and microbe‐associated signals). The signalling processes involved in blue light‐induced stomatal opening (Inoue & Kinoshita, 2017) and abscisic acid (ABA)‐induced stomatal closure are well characterised (Munemasa et al., 2015; Lawson & Matthews, 2020). Although guard cells can respond autonomously to environmental signals such as blue light and ABA, responses can be modulated by mesophyll‐produced signals such as sugar and malate to coordinate stomatal aperture with leaf carbon assimilation (Lawson & Matthews, 2020; Flütsch & Santelia, 2021). Red light‐mediated stomatal opening is less well characterised than the corresponding blue light response. As the red‐light response saturates at similar light intensities to photosynthesis, it is thought to be linked to photosynthetic carbon assimilation; however, the exact nature of this link is unclear (Matthews et al., 2020; Taylor et al., 2024). The red‐light photoreceptor, phytochrome B (phyB) and downstream phytochrome signalling components have been shown to contribute to the coordination of stomatal apertures (Wang et al., 2010; Li et al., 2022; Rovira et al., 2024).
In this study, we analysed how light quality signals controlling light foraging affect stomatal aperture in developing seedlings. Stomatal development is established during embryogenesis, with stomata forming rapidly after germination (Smit et al., 2023). The stomata of young seedlings are responsive to light signals (Rovira et al., 2024), but the role of stomatal movement during seedling establishment is poorly understood. Stomatal apertures were quantified from cotyledons treated with white light (WL) supplemented with FR and/or low‐dose UV‐B to simulate aspects of canopy shade and direct sunlight, respectively. We show that low R : FR inhibits stomatal opening in cotyledons in a response involving the basic helix–loop–helix (bHLH) transcription factor, PIF4 and accumulation of ABA. By contrast, UV‐B supplementation promotes the sustained opening of stomata in a process requiring the UVR8 and phototropin photoreceptors as well as the signalling component BLUE LIGHT SIGNALLING 1 (BLUS1). When low R : FR and UV‐B signals are combined, UV‐B signalling overrides the effects of low R : FR on ABA abundance and stomatal aperture. Our data suggest that the integration of light quality signals by multiple photoreceptors coordinates seedling light‐foraging strategy with water use through the alteration of hormone levels.
Materials and Methods
Plant material and growth conditions
Arabidopsis thaliana seedlings and Chinese kale (Brassica oleracea var Alboglabra) were grown in a 3 : 1 Levingtons F2 compost: silver sand (Melcourt) mixture. Seeds were washed with 70% ethanol, sown on soil and stratified for 2–3 d in the dark at 4°C. Seedlings were grown in long‐day conditions (16 h : 8 h, day : night) in 75 μmol m^−2^ s^−1^ WL (see Supporting Information Fig. S1 for light spectra) at a constant humidity of 70% and a temperature of 20°C in growth cabinets (Microclima 1600E; Snijder Scientific, Tilburg, the Netherlands). All Arabidopsis genotypes are in a Col‐0 background apart from the hy5hyh, and phyB mutants, which are in the Wassilewskija (Ws) background. A list of genotypes (McNellis et al., 1994; Shirley et al., 1995; Kagawa et al., 2001; Holm et al., 2002; Yoshida et al., 2002; Franklin et al., 2003a; Lee et al., 2006; de Lucas et al., 2008; Leivar et al., 2008a,b; Favory et al., 2009; Park et al., 2009; Frey et al., 2012) and their descriptions can be found in Table S1.
Light treatments
Light measurements were performed using an Ocean Optics FLAME‐S‐UV–VIS spectrometer with a cosine corrector. For UV‐B treatments, a Philips TL100W/01 narrowband tube light wrapped in strips of heatproof tape was used to provide treatments of 1 μmol m^−2^ s^−1^ UV‐B light (between 280 and 315 nm). Other than in Fig. 1(b), UV‐B was applied in a background of 75 μmol m^−2^ s^−1^ WL. For FR supplementation, LEDs emitting at a max peak of 735 nm were used in conjunction with 75 μmol m^−2^ s^−1^ WL. R : FR ratios were determined by dividing R light photon irradiance (between 660–670 nm) by FR light photon irradiance (between 725–735 nm). Low R : FR conditions were set between 0.06 and 0.08, whereas high R : FR conditions were between 4 and 7. During gas exchange measurements, light treatments were applied to the measured leaves using the red, blue and FR LEDs within the Licor 6800 multiphase flash fluorometer, while the plant outside the measuring cuvette was treated with light as described above.
Low‐dose UV‐B enhances stomatal opening in a UVR8‐ and phototropin‐dependent manner. Stomatal apertures of 7‐d‐old Arabidopsis seedlings in response to different light treatments. Seedlings were treated with (a) white light (WL) ± UV‐B and (b) darkness ± UV‐B (Dark + UV‐B) over a 6‐h time course. Data are presented as the mean ± SE of each plant's average stomatal aperture. The stomatal responses of (c) uvr8‐6, (d) phot1/2, and (e) blus1‐3 mutants following 6 h of WL ± UV‐B treatment. Treatments were started at dawn, and apertures were measured before dawn (predawn). Data are presented as boxplots showing the median and interquartile range of each group. The upper and lower whiskers represent data within 1.5 × IQR. Each individual plant's mean stomatal aperture is represented as a point on the plot. For all genotype treatment combinations, n = 9–12 seedlings over three independent experiments. Each mean seedling stomatal aperture was calculated from 10 to 12 stomatal measurements. Data were analysed using a two‐way ANOVA, followed by Tukey multiple comparison test.
Stomatal aperture measurements
Arabidopsis seedlings were grown for 7 d in long‐day (16 h : 8 h, light : dark) conditions and transferred at dawn on the 8^th^ day to the appropriate light treatment, unless otherwise stated. Chinese kale seedlings were grown for 8 d and transferred at dawn on the 9^th^ day to appropriate light treatments. The length of light treatment is indicated in each figure. The aerial portion of the seedling was rapidly placed on a microscope slide, and images were taken of the abaxial surface of the cotyledons using a ×40 lens on a Zeiss Axiovert 200M inverted microscope fitted with a Hamamatsu ORCA‐ER digital camera. Images were randomised and stomatal apertures (pore widths) were then measured using Fiji (ImageJ) (Schindelin et al., 2012). For each experiment, stomatal apertures were measured predawn in addition to control and light treatments. Each experiment was repeated independently at least three times, with 3–4 plants measured per genotype per treatment. For Arabidopsis, a total of 10–12 stomatal apertures were measured per plant, and the average of these stomatal aperture values was considered n = 1. For Chinese kale, a total of 12–28 stomatal apertures were measured per plant, with the average of these values considered n = 1. Overall, each experiment has n = 9–12 for each genotype and treatment combination unless otherwise stated.
For Fig. S2(d,e), leaf discs and epidermal peels were harvested from the 5^th^ and 6^th^ rosette leaves of 4–5‐wk‐old plants grown in the same conditions as previously described for seedlings. Leaf discs were harvested using a 4‐mm biopsy punch. After tissue harvesting, leaf discs and epidermal peels were transferred to 50‐mm petri dishes containing 10 ml 10/50 buffer (10 mM MES, 50 mM KCl, pH adjusted to 6.15 using KOH) prewarmed to 20°C. Dishes were incubated in darkness for 2 h before predawn samples were removed for measurement. Remaining dishes were transferred to 75 μmol m^−2^ s^−1^ WL or WL supplemented with 1 μmol m^−2^ s^−1^ UV‐B. Leaf discs were measured 2, 4, and 6 h after treatment. Epidermal peels were measured 6 h after treatment. Each experiment was repeated independently at least three times, with three plants measured per genotype per treatment. A total of 10 stomatal apertures were measured per plant, and the average of these stomatal aperture values was considered n = 1. Overall, each experiment has n = 9 for each genotype and treatment combination.
Gas exchange measurements
Chinese kale seedlings were grown for 14–18 d in long‐day conditions. On the day of measurement, seedlings were kept in darkness until the light treatment. Gas exchange measurements were recorded from seedling cotyledons using a Licor 6800 fitted with a multiphase flash fluorometer with conditions set to a constant flow: 400 μmol m^−2^ s^−1^, T air: 20°C, sample CO_2_: 400 ppm, sample H_2_O: 15 000 ppm, and mixing fan set to 7000 rpm. Cotyledon gas exchange was monitored in darkness for 30 min before treatment with 75 μmol m^−2^ s^−1^ red and blue light (67 and 8 μmol m^−2^ s^−1^, respectively) ± 10 μmol m^−2^ s^−1^ far‐red light for 2 h (see Fig. S1b for light spectra).
Stomatal density measurements
Arabidopsis seedlings were grown for 7 d in long‐day (16 h : 8 h, light : dark) conditions. On the 8^th^ day, cotyledons were harvested and cleared (Sharma, 2017). Cleared tissue was imaged using a ×20 DIC lens on a Leica DMIRE2 microscope with a Leica DFC350 FX camera. A stack of images containing both the abaxial and adaxial epidermises was taken. Stomata were counted using the cell counter plugin of Fiji (ImageJ) within a 0.208‐mm^2^ region.
ABA quantification
Arabidopsis seedlings were grown and treated similarly to stomatal aperture experiments. After light treatment, aerial seedling tissues were rapidly harvested, weighed and flash frozen in liquid nitrogen. Seedling tissue was ground into a fine powder and resuspended in 1.9 ml extraction buffer (1% v/v acetic acid in 100% isopropanol), spiked with 10 μl d6‐ABA (2.5 μg ml^−1^ in 100% MeOH). Samples were mixed overnight at 4°C and spun at 13.4 krcf for 5 min at 4°C. Supernatant was evaporated 950 μl at a time into a new tube using an Eppendorf Concentrator Plus set to V‐AL mode at 45°C for 1 h. The original sample tubes were resuspended in 950 μl of extraction buffer (without d6‐ABA) and centrifuged further for an hour at 4°C. The sample was centrifuged again at 13.4 krcf at 4°C, and the supernatant was transferred to the tube containing the evaporated sample, which was evaporated a final time using the same settings as discussed before. Dried samples were stored at −70°C until mass spectrometry analysis.
Dried samples were resuspended in 100 μl 100% MeOH and run on an Acquity UPLC equipped with a XevoTQS tandem mass spec (Waters, Milford, MA, USA). Separation was performed on a 50 × 2.1 mm 2.6 μ Kinetex EVO C18 column (Phenomenex, Torrance, CA, USA) using the following gradient of acetonitrile vs 0.1% formic acid in water, run at 0.7 ml min^−1^ and 30°C (0 min – 5%, 3 min – 95%, 3.5 min – 95%, 3.6 min – 5%, 5.1 min – 5%). Samples were maintained at 10°C, and the instrument injected 5 μl. Hormones were detected by negative mode electrospray, with the following mass transitions: ABA, 263 > 153; d6‐ABA, 269 > 159 (collision energy 10 V). Spray chamber conditions were 900 l h^−1^ drying gas at 500°C, 150 l h^−1^ cone gas, 7.0 bar nebuliser pressure and a spray voltage of 1.5 kV.
Thermal imaging
Thermal images were recorded using a FLIR A665sc thermal imaging camera. ResearchIR software (v.4.40.9.30) was used to generate TIF images, which were then analysed using Fiji (ImageJ). Cotyledon temperatures were analysed within a 24‐pixel box, and the mean of the 24 pixels was used to represent each cotyledon's temperature.
Chlorophyll fluorescence
Chlorophyll fluorescence parameters of 7‐d‐old seedlings were measured using an IMAGING‐PAM M series system with ImagingWin software (v.2.56p, Walz). Seedlings were treated with WL ± UV‐B and ± FR for 6 h. Following this, seedlings were dark‐adapted for 30 min before applying a saturating pulse of light. Maximal photosystem II efficiency (F V/F M) was calculated using the formula F V/F M = (F M – F 0)/F M, where F 0 and F M represent fluorescence measurements before and after the saturating light pulse, respectively. Images were exported from the ImagingWin software as .tif files and analysed using Fiji (ImageJ). Manual thresholding was used to generate masks of the seedling aerial tissue before calculating the mean F V/F M for that region of interest.
RNAseq analysis
The raw count RNAseq data from GSE146125 (Tavridou et al., 2020b) and GSE192469 (Sharma et al., 2023) were downloaded from the NCBI GEO database. These studies treated 7‐d‐old Arabidopsis seedlings with supplemental far‐red and/or UV‐B for 3 and 4 h, respectively. EdgeR (v.4.6.2) was used to filter out low‐expressed genes, normalise sample library sizes and perform differential expression analysis using the quasi‐likelihood pipeline (quasi‐likelihood negative binomial model). Genes with FDRs (calculated using the Benjamini‐Hochberg method) less than 0.05 were classed as differentially regulated. Heat maps were generated for genes tagged with ABA biosynthesis, catabolism and signalling GO terms. Normalised log_2_(CPM) Z‐scores were used for heat map visualisation.
qPCR analysis
Arabidopsis aerial tissue was harvested similarly to ABA quantification. RNA was extracted from samples using the Spectrum Total RNA kit (Sigma, St Louis, MO, USA). cDNA was synthesised using the high‐capacity cDNA reverse transcription kit with RNase inhibitor (Applied Biosystems, Foster City, CA, USA). qPCR was performed using the Brilliant III Ultra‐Fast SYBR Green qPCR Master Mix (Agilent, Santa Clara, CA, USA) and an Mx3000P (Agilent). Transcript abundances were calculated using the ∆∆C t method, with PP2A and ACT2 used as reference genes unless otherwise stated. Statistics were performed using the −∆∆C t values. Primers are described in Table S2.
Data presentation and analysis
All data were statistically analysed in R (v.4.3.1) (R Core Team, 2021) and plotted using the ggplot2 package (Wickham, 2016).
Statistical analysis
Data were analysed using R (v.4.3.1). Datasets were analysed using one or two‐way ANOVAs with post hoc Tukey multiple comparison tests. Due to the increased number of mutant genotypes and the resulting large number of uninformative multiple comparisons that would arise from post hoc Tukey test, Holm‐corrected t‐tests were used to assess the FR responsiveness of Col‐0 and the bg1, q1124 and nced3/5 mutants in Fig. 4(d) (see later).
Results
Low‐dose UV‐B enhances stomatal opening in a UVR8‐ and phototropin‐dependent manner in seedling tissue
To assess the effect of low‐dose UV‐B (1 μmol m^−2^ s^−1^) on stomatal aperture, 7‐d‐old Arabidopsis seedlings were transferred to WL ± UV‐B at dawn (light spectra are presented in Fig. S1). Cotyledon stomatal apertures were recorded over a 6‐h period and are presented in Fig. 1(a). In contrast to reports using older leaves and/or higher doses of UV‐B or epidermal strips (He et al., 2013; Tossi et al., 2014; Li et al., 2017; Ge et al., 2020), seedlings supplemented with UV‐B showed enhanced stomatal opening when compared with WL controls during the latter part of the time course. Exposing seedlings to monochromatic UV‐B (Fig. 1b) showed that the UV‐B‐induced enhancement of stomatal opening requires a background of light within the 400–700‐nm wavelength range. Stomatal responses to UV‐B were also analysed in mature plants. Here, apertures were measured from rosette leaves, and in leaf disc and epidermal peel tissue treated while floating on a buffer. In contrast to cotyledons, no UV‐B‐enhanced stomatal opening was observed in rosette leaves (Fig. S2a–c). Some UV‐B‐mediated increase in stomatal aperture was, however, observed in leaf disc and epidermal tissue (Fig. S2d,e).
Blue light‐induced stomatal opening involves the redundant actions of phototropin photoreceptors, phot1 and phot2 (Kinoshita et al., 2001). Analyses of the photoreceptor mutants uvr8‐6 and phot1/2 showed that both are required for UV‐B‐enhanced stomatal opening (Fig. 1c,d), suggesting a role for phototropin and UVR8 signalling. Additionally, mutants deficient in the downstream phototropin signalling component BLUS1 (blus1‐3) (Takemiya et al., 2013) displayed a similar phenotype to phot1/2 (Fig. 1e), with no additional opening in response to UV‐B supplementation observed. These data provide further support for a role for phototropin signalling in UV‐B–enhanced stomatal opening.
Following UV‐B absorption, UVR8 dimers monomerise and interact with the E3 ubiquitin ligase CONSTITUTIVELY PHOTOMORPHOGENIC (COP1) (Favory et al., 2009; Cloix et al., 2012). This stabilises the bZIP transcription factors ELONGATED HYPOCOTYL 5 (HY5) and HY5 HOMOLOGUE (HYH), which control the expression of a number of UV‐B‐regulated genes (Brown et al., 2005; Brown & Jenkins, 2008). We further investigated the involvement of these components in the enhanced stomatal opening response observed under UV‐B‐supplemented conditions. The hy5/hyh double mutant displayed wild‐type stomatal responses to UV‐B light, suggesting that these transcription factors are not required for UV‐B‐mediated stomatal opening (Fig. S3a). This mutant is in the Ws background, which behaved similarly to Col‐0, confirming the consistency of this response across different Arabidopsis accessions. Consistent with previous reports (Mao et al., 2005, p. 2; Khanna et al., 2014; An et al., 2022), the cop1 mutant showed consistently open stomata under all conditions (Fig. S3b). No further opening was observed following UV‐B supplementation, but interpretation of this result is confounded by the mutant's existing phenotype. Additionally, we observed that the tt4 mutant (a flavonoid biosynthesis mutant hypersensitive to UV‐B and more prone to UV‐B‐induced damage (Li et al., 1993) showed no UV‐B‐mediated enhancement of stomatal opening, suggesting damaging amounts of UV‐B may prevent the enhanced opening observed at 6 h (Fig. S3c).
Low R : FR ratio treatment inhibits stomatal opening and is antagonised by UV‐B
Reduced R : FR is a major component of vegetative shade that drives inactivation of phytochrome photoreceptors, auxin production and the elongation of stems for light foraging (Ballare et al., 1990; Franklin, 2008; Fernández‐Milmanda & Ballaré, 2021). The effect of reduced R : FR (achieved by supplementing WL with FR) on seedling stomatal apertures was investigated (Figs 2a, S2). Low R : FR decreased stomatal aperture in a response observed after 1–3 h of treatment. When WL was supplemented with both FR and UV‐B light, stomatal apertures were no longer reduced, suggesting that the presence of UV‐B overrides the effect of low R : FR. Decreased stomatal apertures were also observed in rosette leaves, although at later time points than in cotyledons (Fig. S2a–c).
Low R : FR inhibits stomatal opening in a phytochrome interacting factor‐dependent manner and is antagonised by UV‐B. (a) Arabidopsis Col‐0 seedling stomatal apertures were monitored over a 7‐h time course where plants were treated with white light (WL) ± far red (FR) and/or low dose UV‐B. (b–e) Col‐0, Ws, phyB and pif mutant seedlings were treated with WL ± FR for 2 h before measurement of stomatal apertures. These were also measured before dawn (predawn). In (a) data are presented as the mean ± SE of each plant's average stomatal aperture. In (b, c), data are presented as boxplots showing the median and interquartile range (IQR) of each group. The upper and lower whiskers represent data within 1.5 × IQR. Each individual plant's mean stomatal aperture is represented as a point on the plot. For all genotype treatment combinations, n = 9–12 seedlings over three independent experiments. Each mean seedling stomatal aperture was calculated from 10 to 12 stomatal measurements. Data were analysed using a 2‐way ANOVA, followed by Tukey's multiple comparison test.
To investigate whether these responses are conserved beyond Arabidopsis, the movement of the Brassica crop Chinese kale (Brassica oleracea var. alboglabra) cotyledon stomata was monitored in response to FR and UV‐B supplementation (Fig. S4a). The increased cotyledon size of this species additionally facilitated gas exchange analyses. In support of our Arabidopsis observations, stomatal apertures were reduced in plants treated with WL supplemented with FR for 6 h, when compared with WL controls. The addition of low‐dose UV‐B treatment also overcame the FR supplementation effect (Fig. S4a). Unlike Arabidopsis, Chinese kale did not show significant differences between WL‐ and FR‐supplemented plants at 2 h; however, both stomatal apertures and stomatal conductance trended towards decreased values (Fig. S4b,c). No significant differences in photosynthetic CO_2_ assimilation were recorded between WL‐ and FR‐supplemented plants over this time period (Fig. S4d,e).
The role of phytochrome interacting factor (PIF) transcription factors in Arabidopsis stomatal responses to low R : FR was next explored, as PIFs have been shown to regulate both shade avoidance (Lorrain et al., 2008; Leivar & Quail, 2011; Li et al., 2012) and stomatal aperture (Li et al., 2022; Rovira et al., 2024). PIF4, PIF5 and PIF7 are the major regulators of plant architectural responses to low R : FR, so we therefore focused on these genes (Lorrain et al., 2008; Hornitschek et al., 2009; Li et al., 2012; Burko et al., 2022). pif7 mutant seedlings displayed a wild‐type response to reduced R : FR (Fig. 2b), whereas pif4 and pif4/5 mutants showed more variable stomatal apertures, with no significant differences between WL and low R : FR (Fig. 2c,d). Furthermore, the pif4/7 double mutant displayed an insensitivity to low R : FR, with significantly more open stomata than Col‐0 (Fig. 2d). This suggests a dominant role for PIF4 in inhibiting stomatal opening in low R : FR conditions, with a potential minor redundant role for PIF7. Consistent with previous reports (Hayes et al., 2017), UV‐B supplementation reduced PIF4 transcript accumulation in a UVR8‐dependent manner (Fig. S5a). UVR8 has also been shown to target PIF4 protein for degradation in UV‐B (Hayes et al., 2017; Tavridou et al., 2020a), presenting the possibility that PIF4 integrates low R : FR and UV‐B signalling in the regulation of stomatal aperture. The effect of UV‐B supplementation on stomatal aperture was investigated in pif4/7 mutants at 6 h (Fig. S5b), when maximum responsiveness to UV‐B treatment was observed in wild‐type seedlings (Fig. 2a). UV‐B‐mediated increases in stomatal aperture were mediated by UVR8 at this time point (Fig. S5c). In contrast to low R : FR, responsiveness to UV‐B was retained in pif4/7 mutants (Fig. S5b), supporting the existence of a PIF4‐independent mechanism promoting stomatal opening.
PIF activity is regulated by the red and far‐red light‐absorbing phytochrome photoreceptors (Cheng et al., 2021), with phyB performing a major role. In high R : FR, activated phyB binds to PIFs, leading to their ubiquitination and degradation via the 26S proteasome (Halliday & Whitelam, 2003; Monte et al., 2003; Franklin et al., 2003b). The stomatal response of phyB mutants to low R : FR was therefore explored (Fig. 2e). These plants displayed significantly smaller stomatal apertures than wild‐type controls in WL, consistent with a role for phyB in promoting stomatal opening (Wang et al., 2010). The Ws ecotype contains a natural deleterious mutation at its PHYD locus (Aukerman et al., 1997). PhyD and PhyE perform minor redundant roles with phyB in the regulation of multiple light responses (Devlin et al., 1999; Franklin et al., 2003a). It is possible that phyD also contributes to the regulation of seedling stomatal aperture, but further mutant analysis would be required to confirm this. Apertures of Ws phyB mutants were not further decreased in low R : FR, suggesting that phyE is not involved in this response.
To exclude the possibility that light treatments were indirectly affecting stomatal apertures through leaf temperature changes, thermal imaging was used to track cotyledon temperature over a 6‐h time course (Fig. S6a,b). At 2 h post dawn, there was no significant difference in cotyledon temperature between WL and WL + FR treatments. However, treatments involving the addition of UV‐B showed cotyledons to be c. 0.5°C warmer than treatments without UV‐B (Fig. S6c). Temperatures greater than 35°C have been shown to elicit increased stomatal opening (Devireddy et al., 2020; Kostaki et al., 2020; Korte et al., 2023). To assess whether slightly elevated cotyledon temperature may account for the increased stomatal opening observed in UV‐B supplemented conditions, seedlings were treated ± UV‐B at 20 and 28°C (Fig. S6d). Incubation at 28°C did not promote stomatal opening, and UV‐B responses were observed at both temperatures. The stomatal densities of uvr8‐6 and pif4 mutant cotyledons were additionally analysed and found to show no significant differences from Col‐0 (Fig. S7). It can therefore be concluded that differences in UV‐B and FR responses observed in these genotypes are not due to differences in stomatal density.
R : FR and UV‐B control ABA levels
ABA performs a major role in regulating stomatal responses to drought stress. It is a potent trigger of stomatal closure and functions to keep stomata closed by inhibiting stomatal opening (Hsu et al., 2021; Pei et al., 2022). Multiple studies have reported links between the biosynthesis and signalling of ABA and phytochrome signalling (González et al., 2012; Holalu et al., 2020; Liang et al., 2020; Qi et al., 2020; Li et al., 2022). PIF and ABA accumulation dynamics have been observed to perform a role in the regulation of stomatal apertures over day : night cycles (Rovira et al., 2024). ABA signalling has also been linked to UV‐B responses, as in Zea mays, high‐dose UV‐B treatment stimulates ABA production (Tossi et al., 2009).
To investigate the impact of FR and/or UV‐B supplementation on seedling ABA metabolism, two pre‐existing RNA‐seq datasets from 7‐d‐old Arabidopsis seedlings treated with WL ± FR ± UV‐B for 3–4 h were re‐analysed (Tavridou et al., 2020b; Sharma et al., 2023). Genes tagged with both ABA biosynthesis (GO:0009688) and ABA catabolism (GO:0046345) show clear patterns of regulation, with UV‐B up‐ and downregulated subsets. The effect of FR supplementation alone is less clear, with a smaller number of differentially expressed genes under these conditions (Fig. S8). A similar response was observed for genes tagged with positive and negative ABA signalling GO terms (GO:0009789 and GO:0009788, respectively; Fig. S9).
We next quantified the transcript abundances of key ABA biosynthesis and catabolism genes (Figs 3a–c, S10) in seedlings treated with FR and/or UV‐B supplementation for 6 h. We observed a significant effect of light treatment on the transcript accumulation of key ABA biosynthesis enzymes NCED3 and NCED5 (Fig. 3b,c). Post hoc tests showed that NCED3 is significantly upregulated in low R : FR, whereas differences between individual groups were not observed for NCED5. We also analysed beta glucosidase enzyme (BG1) (which encodes an enzyme that rapidly generates active ABA from a pool of inactive ABA glucosyl‐ester, also known as BGLU18) and observed no significant effects of light treatment on transcript abundance (Fig. S10). Interestingly, a significant decrease in BG1 transcript levels was observed following UV‐B treatment in the RNAseq analysis, potentially reflecting differences in treatment length between studies (Fig. S9). The genes CYP707A1–3 encode ABA catabolism enzymes involved in the breakdown of ABA (Saito et al., 2004). Similar to the RNAseq data, significant effects of light treatment were observed on the transcript abundance of these genes. CYP707A2 showed significant downregulation in low R : FR and higher transcript levels in UV‐B across multiple biological repeats. Conversely, CYP707A1 and A3 showed opposite responses, with decreased transcript abundance in UV‐B (Fig. S10).
Low R : FR and UV‐B supplementation regulate the transcript abundance of key abscisic acid (ABA) metabolism enzymes and seedling ABA content. (a) CYP707A2, (b) NCED3 and (c) NCED5 transcript abundance was measured in the aerial tissue of 7‐d‐old Arabidopsis seedlings treated with white light (WL) ± far red (FR) ± UV‐B light for 6 h. ABA levels in the aerial tissue of 7‐d‐old (d) Col‐0 and uvr8‐6, and (e) Ws seedlings treated for 6 h with WL ± FR ± UV‐B light and quantified using LC‐MS. All data are presented as small semi‐transparent points and the mean as a larger non‐transparent point. Error bars represent SE. (a–c) n = 3 pooled seedling RNA samples, (d) n = 5 pooled seedling samples over 5 independent experiments, and (e) n = 3 samples over three independent experiments. All data were analysed using a 2‐way ANOVA, followed by Tukey multiple comparison tests, except for (e), where a one‐way ANOVA was used.
To assess how the ABA content of aerial seedling tissue was affected by different light treatments, we assayed the ABA content of Col‐0, uvr8‐6, and Ws seedlings treated with UV‐B, FR or a combination of the two light treatments for 6 h. Analysis of Col‐0 and uvr8‐6 data in Fig. 3(d) using a two‐way ANOVA showed a significant effect of both genotype (F 1,32 = 8.15, P = 0.008) and light treatment (F 3,32 = 11.45, P < 0.001), although no significant interaction was observed (F 1,3 = 1.90, P = 0.150). ABA content in Col‐0 and uvr8‐6 seedlings was quite variable, and post hoc tests showed no significant differences between individual groups. Much less variability was, however, observed in Ws seedlings. Here, analysis using a one‐way ANOVA showed a similar significant effect of light treatment (F 3,8 = 14.78, P = 0.001). Furthermore, post hoc multiple comparison tests showed a significant increase in seedling ABA content in low R : FR, which was abolished when FR was combined with UV‐B (Fig. 3e). In both Col‐0 and Ws accessions, increased ABA content was repeatedly observed in low R : FR conditions, whereas decreased ABA was observed in the presence of UV‐B. When low R : FR and UV‐B treatments were combined, an intermediate ABA content was observed, resembling levels observed in WL. Together, these data suggest that UV‐B perceived by UVR8 can counteract low R : FR‐induced increases in seedling ABA (Fig. 3d,e).
To further investigate the potential role of ABA signalling in light‐regulated stomatal movement, mutants defective in ABA biosynthesis (nced3/5 – defective in de novo ABA biosynthesis (Frey et al., 2012), ABA activation (bg1; Lee et al., 2006), ABA signalling (q1124 – a quadruple ABA receptor mutant; Park et al., 2009) and stomatal opening (ost1‐3 – a mutant in a key downstream kinase; Yoshida et al., 2002)) were assayed in response to UV‐B and low R : FR treatments. Here, the ABA biosynthesis mutants, nced3/5 and bg1, and the quadruple receptor mutant q1124 displayed wild‐type responses to UV‐B supplementation, whereas the ost1‐3 mutant showed no significant increase in stomatal opening (Fig. 4a–c). The nced3/5 ABA biosynthesis and q1124 ABA receptor mutants displayed increased apertures in WL, supporting previous observations (Merilo et al., 2013; Pridgeon & Hetherington, 2021). In all ABA biosynthesis and receptor mutants, no significant differences were observed between WL and low R : FR conditions, supporting the involvement of ABA in the low R : FR‐mediated inhibition of stomatal opening (Fig. 4d). Interestingly, this phenotype was most pronounced in the bg1 ABA biosynthesis mutant, suggesting that deconjugation of ABA from ABA‐GE is central to elevating ABA levels in low R : FR conditions (Fig. 4d). Similarly to bg1, the ost1‐3 signalling mutant showed no stomatal aperture response to low R : FR conditions, confirming the importance of OST1 in stomatal aperture regulation (Yoshida et al., 2002) (Fig. 4e). Together, these data suggest that ABA biosynthesis and signalling are not essential for UV‐B‐induced stomatal opening but are required for low R : FR‐induced inhibition of stomatal opening.
Abscisic acid (ABA) signalling and metabolism mutants respond to UV‐B but display defective responses to low R : FR. Stomatal apertures of 7‐d‐old Arabidopsis (a) ABA biosynthesis mutants nced3/5 and (b) bg1, and (c) ABA signalling mutants q1124 and ost1‐3, were measured predawn and following a 6 h white light (WL) ± UV‐B treatment. The stomatal aperture response of ABA biosynthesis and signalling mutants (d) bg1, q1124, nced3/5 and (e) ost1‐3 was measured predawn and in response to 2 h WL ± far red (FR) treatment. Data are presented as boxplots showing the median and interquartile range (IQR) of each group. The upper and lower whiskers represent data within 1.5× IQR. Each individual plant's mean stomatal aperture is represented as a point on the plot. For all genotype and treatment combinations, n = 9 seedlings over three independent experiments. Each mean seedling stomatal aperture was calculated from 10 stomatal measurements. All data were analysed using a 2‐way ANOVA, followed by Tukey multiple comparison tests, except for (d) where t tests were used to make specific comparisons and adjusted for multiple comparisons using a Holm correction. (e) ost1‐3 was grown in parallel with pif7‐2 (Fig. 2c) each plot uses the same Col‐0 control data.
Discussion
This study shows that the stomatal movements of seedlings are modulated by integrated light quality signals. In both Arabidopsis and the Brassica crop Chinese Kale, low R : FR acts to limit stomatal aperture in seedling cotyledons (Fig. 2). However, when cotyledons are additionally exposed to low‐dose UV‐B, the low R : FR restriction of stomatal aperture is lifted, and stomatal opening is promoted (Figs 1, 2a). Our data suggest that low R : FR inhibits stomatal opening through promoting ABA accumulation, and that UV‐B light can antagonise this, in part, through suppression of ABA levels (Figs 3, 4).
Low‐dose UV‐B promotes stomatal opening in seedling cotyledons
In contrast to our observations, most studies focusing on stomatal responses to UV‐B have shown UV‐B irradiation to induce stomatal closure. A number of components underlying this response have been identified, including the UVR8 photoreceptor, COP1, HY5 and HYH (Tossi et al., 2014; Ge et al., 2020); a G alpha protein (He et al., 2013); MAP KINASES (MAPKs) (Li et al., 2017); ethylene (He et al., 2011; Ge et al., 2020); hydrogen peroxide (H_2_O_2_); nitric oxide (NO) (He et al., 2005, 2013); and the LIPOXYGENASE 1 (LOX1) enzyme (Liu et al., 2025). The latter are stress signalling components, most likely induced by the high doses of UV‐B applied. Here, we found that lower dose UV‐B (1 μmol m^−2^ s^−1^) applied in a background of WL increases stomatal apertures of the cotyledon abaxial epidermis (Figs 1, 2a). This occurs in both Col‐0 and Ws Arabidopsis accessions and Chinese Kale (Figs 1, S3, S4a), but not in mature plant leaves (Fig. S2a–c). However, caution must be applied when comparing responses of cotyledons and mature rosette leaves. Many physiological factors differ between the two, including the UV‐B penetrance of cotyledon and mature plant leaf tissue. When stomata are isolated from the rest of the leaf (epidermal strips) or leaf tissue is excised from the plant (leaf discs), a small amount of opening is observed (Fig. S2d,e), suggesting that although isolated guard cells can respond to low‐dose UV‐B supplementation, other cells are required for a full opening response. It is unlikely that the light treatments used in this study were damaging, as F v/F m measurements of Chl fluorescence (commonly used as an indicator of photosystem health (Murchie & Lawson, 2013)) varied little in response to low R : FR or UV‐B. Unsurprisingly, a mild reduction in F v/F m was observed in UV‐B‐treated uvr8‐6 mutants, which are unable to initiate photoprotective responses (Fig. S11). Some studies have reported stomatal opening in response to UV‐B supplementation in Vicia faba (Jansen & Van Den Noort, 2000), cucumber (Teramura, 1983) and certain Ericaceae species (Musil & Wand, 1993), and it has been suggested that the effect of UV‐B light on stomata is dependent on the plant metabolic state (Nogués et al., 1999; Jansen & Van Den Noort, 2000).
We show that the opening of seedling abaxial stomata following UV‐B treatment requires the presence of light within the photosynthetically active range (400–700 nm), functional phototropin and UVR8 photoreceptors (Fig. 1), together with the downstream phototropin signalling component BLUS1 (Fig. 1e). The requirement for phototropin signalling differs from observations showing that low doses of UV‐B alone could stimulate stomatal opening in both Arabidopsis and Vicia faba (Eisinger et al., 2003). However, there are numerous differences between plant growth conditions, ages and experimental procedures that may explain the contrasting observations. In this study, we have focused on the stomatal apertures of intact seedling cotyledon tissue and tracked apertures over a longer period through the course of the day.
Our data suggest that UV‐B‐mediated promotion of stomatal opening does not require the UV‐B signalling components HY5 or HYH (Fig. S3a). This is in contrast to the UVR8‐mediated promotion of stomatal closure (Tossi et al., 2014; Ge et al., 2020). Upon assaying mutants deficient in the UV‐B signalling component, COP1, we observed a constitutively open response before dawn, and no difference between WL and WL + UV‐B conditions (Fig. S3b). These mutants have previously been shown to present a constitutively open stomata phenotype in the dark (Mao et al., 2005; Wang et al., 2010; Khanna et al., 2014; An et al., 2022), confounding interpretation of the role of COP1 in UV‐B promotion of stomatal opening.
Low R : FR inhibits cotyledon stomatal opening in a PIF4‐dependent manner
We further show that seedlings exposed to low R : FR ratio display reduced stomatal apertures following 1–3 h of treatment in a response requiring phyB and PIF4 (Figs 2, 3, S2). Previous studies have shown low R : FR to inhibit stomatal opening in Commelina communis and orchids (Roth‐Bejerano et al., 1990; Talbott et al., 2002), with no effect observed in Arabidopsis (Talbott et al., 2003). The differences in our observations may be due to using intact cotyledon tissue, as opposed to epidermal peels from mature leaves. Monochromatic FR treatment has been shown to reduce stomatal apertures in rice in a mechanism requiring the PIF‐homologue, OsPIL15 (Li et al., 2022). PIFs have also been shown to play a role in regulating the daily rhythmic opening and closing of stomata (Rovira et al., 2024). Here, the authors show PIFs to act in opposition to ABA, promoting the accumulation of K^+^ import channel POTASSIUM CHANNEL IN ARABIDOPSIS THALIANA (KAT1) and ultimately stomatal opening at dawn in 3‐d‐old Arabidopsis seedlings. This positive role in stomatal opening contrasts with the negative role identified in this study and may reflect differences in plant age or growth conditions. Here, seedlings were soil‐grown, whereas Rovira et al. used ½ Murashige & Skoog (½ MS) plates, where humidity would be considerably higher. Growth under high humidity conditions is known to affect stomatal responses to several signals, including ABA (Aliniaeifard & Van Meeteren, 2013). KAT1 activity depends on its cellular location. ABA can trigger endocytosis of KAT1 from guard cell plasma membranes to endosomal compartments, thus preventing it from contributing to stomatal opening (Sutter et al., 2007). We observed a (non‐significant) increase in KAT1 transcript abundance in low R : FR (Fig. S10), which may involve PIFs (Rovira et al., 2024). The functional relevance of increased KAT1 abundance in low R : FR is, however, unclear, as the increased ABA levels in these conditions would likely reduce KAT1 activity. Further work is therefore required to explore the potential role of KAT1 in this response.
Several studies have identified connections between ABA signalling, phytochrome signalling and red/far‐red light responses. PIFs have been shown to bind to the promoter regions of ABA biosynthesis and signalling genes (Liang et al., 2020), as well as interact with the ABA receptor proteins PYL8 and PYL9 (Qi et al., 2020). Mutants deficient in phyB show increased ABA levels but reduced ABA sensitivity in well‐watered conditions (González et al., 2012). ABA content has been shown to decrease in plants treated with red light (Zhu et al., 2020), and conversely, increase in low R : FR (Cagnola et al., 2012; González‐Grandío et al., 2013; Holalu & Finlayson, 2017; Ortiz‐Alcaide et al., 2019; Holalu et al., 2020; Michaud et al., 2023). Additionally, genes tagged with the GO term ‘response to ABA’ are upregulated in low R : FR‐treated Arabidopsis seedlings (Kohnen et al., 2016) and FR‐treated leaf tips (Küpers et al., 2023).
Analysis of RNAseq data from experiments using similar treatments to this study (Tavridou et al., 2020b; Sharma et al., 2023) showed that UV‐B (and to a smaller extent FR) supplementation altered the transcript abundance of genes tagged with ABA metabolism and signalling GO terms (Figs S8, S9). In our conditions, transcripts of the ABA biosynthesis enzyme NCED3 were upregulated, while transcripts of the ABA degradation enzyme CYP707A2 were downregulated by FR, but not by FR + UV‐B supplementation (Fig. 3). We observed multiple ABA signalling and metabolism mutants to show reduced responses to low R : FR (Fig. 4). These data support a mechanism whereby, in low R : FR, expression of the ABA biosynthesis enzyme NCED3 is increased and BG1 functions to generate active ABA from a pool of inactive ABA‐GE, which together increase ABA signalling, OST1 activation and inhibition of stomatal opening. The parallel decrease in ABA degradation enzyme CYP707A2 transcripts in these conditions may further contribute to increased seedling ABA content.
UV‐B antagonises ABA accumulation and stomatal responses to low R : FR
When seedlings were exposed to both FR and UV‐B supplementation simultaneously, UV‐B antagonised low R : FR‐mediated inhibition of stomatal opening (Fig. 2a). A parallel response is observed in hypocotyl elongation, where low R : FR‐induced elongation is inhibited in the presence of UV‐B. This results, in part, from UVR8‐mediated sequestration of COP1, destabilising PIF proteins, independently from HY5/HYH‐mediated signalling pathways (Hayes et al., 2014; Sharma et al., 2019). Addition of UV‐B to low R : FR reversed elevations in ABA (Fig. 3d,e), likely through the upregulation of CYP707A2 and downregulation of NCED3 (Fig. 3a,b). These observations, coupled with analyses of ABA biosynthesis and signalling mutants (Fig. 4d,e), suggest that an increase in ABA content is required for low R : FR‐mediated inhibition of stomatal opening. A hypothetical model outlining the mechanism behind this response is proposed in Fig. 5. In vegetational shade, low R : FR inactivates phyB, stabilising and promoting PIF4 activity. ABA accumulates via de novo synthesis and de‐conjugation of ABA‐GE. When plants reach a gap in the canopy, UV‐B perceived by UVR8 suppresses PIF4 activity (Hayes et al., 2014; Sharma et al., 2019) and ABA accumulation, removing repression of stomatal opening. In non‐shaded (high R : FR) conditions, UV‐B supplementation decreases ABA content in a UVR8‐dependent manner and promotes stomatal opening through a mechanism requiring phototropins (Fig. 3e).
Proposed model for seedling stomatal aperture control by low R : FR and UV‐B. A proposed mechanism for the action of low‐red : far‐red (R : FR) and UV‐B on stomatal opening. (1) Low R : FR promotes the conversion of the phytochrome B photoreceptor to its inactive Pr state, which, in turn, stabilises the phytochrome interacting factors (PIFs). In this study, we observe PHYTOCHROME INTERACTING FACTOR 4 (PIF4) is required for seedling stomatal responses to FR supplementation. (2) Abscisic acid (ABA) biosynthesis and signalling components are required for inhibition of stomatal opening, likely downstream of PIF activity. The beta glucosidase enzyme (BG1) is involved in cleaving the glucose ester off an inactive pool of ABA‐GE to rapidly generate active ABA, and the NCED3 and NCED5 enzymes catalyse a key step in de novo ABA biosynthesis. Here, we suggest that low R : FR‐induced increases in seedling ABA levels drive inhibition of stomatal opening. UV‐B supplementation abolishes the inhibition of stomatal opening in low R : FR. (3) UV‐B, perceived by UV RESISTANCE LOCUS 8 (UVR8) targets PIFs for inactivation and/or degradation. Inhibition of PIF function likely prevents the accumulation of ABA, relieving inhibition of stomatal opening. (4) UVR8 may additionally function to directly inhibit ABA biosynthesis and/or signalling. (5) UVR8 also promotes further stomatal opening through additional mechanisms unrelated to ABA signalling, involving phototropin photoreceptors and BLUE LIGHT SIGNALLING 1 (BLUS1). Solid and dashed lines represent known and unknown connections respectively.
Conclusion: seedlings integrate light quality signals to coordinate growth and stomatal aperture in dynamic light environments, via regulation of hormone abundance
Following germination, seedlings must balance growth and water use to transition to photoautotrophic development without depleting available resources. Light quality provides key information concerning the prevailing levels of vegetational shade and sunlight availability (Fernández‐Milmanda & Ballaré, 2021). Plants perceive light quality using multiple photoreceptors, and integrate this information to regulate levels of multiple hormones, altering their physiology and development to optimise survival (Brini et al., 2022). Low R : FR and UV‐B control auxin and gibberellin abundance and signalling to adjust plant architecture for maximum light capture. Here, we show that low R : FR and UV‐B also regulate ABA content in seedling aerial tissue to control stomatal movements.
While the data presented here were gathered from plants grown in controlled environment conditions and have direct relevance to protected crops, it is pertinent to ask whether they have wider relevance to the field situation. In this context, it is worth considering that when a seedling emerges under a vegetative canopy, low R : FR‐mediated inhibition of stomatal opening may allow seedlings to restrict water loss without greatly impacting photosynthetic rates. While testing this possibility is beyond the scope of the current work, it seems likely that the low R : FR‐mediated inhibition of opening would be overridden in the presence of low amounts of UV‐B light. This would allow seedling stomata to rapidly respond to sunflecks (gaps in the canopy known to contain quantities of UV‐B light) (Moriconi et al., 2018). When seedlings emerge from a canopy, they are exposed to increased amounts of photosynthetic light. R : FR increases, and the inhibition of stomatal opening is likely attenuated. Prolonged exposure to ambient UV‐B would then further open stomata, ensuring maximum photosynthetic productivity in sunlight. Additional experiments carried out in the field are required to test this possibility directly. However, it should be noted that the relevance of our experiments to other species, at least in the Brassicaceae, is strengthened by our findings in two members of this family.
Competing interests
None declared.
Author contributions
MG, KAF and AJP designed experiments. MG, LH and AJP performed experiments and analysed data. MG, KAF and AJP wrote the manuscript.
Disclaimer
The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.
Supporting information
Fig. S1 Light spectra of the different treatments described in this study. Fig. S2 Cotyledons and rosette leaves differ in stomatal responses to light quality. Fig. S3 *UV‐B‐*mediated stomatal opening likely requires COP1, but not HY5/HYH. Fig. S4 Low dose UV‐B treatment overrides far‐red inhibition of opening in Chinese kale cotyledons. Fig. S5 UV‐B regulates PIF4 transcript abundance and induces stomatal opening in pif4/7 mutants but not uvr8‐6. Fig. S6 UV‐B‐mediated increases in stomatal aperture do not result from small elevations in cotyledon temperature. Fig. S7 uvr8 and pif4 mutants show no significant differences in cotyledon stomatal density. Fig. S8 FR and UV‐B supplementation affect the transcript levels of genes involved in ABA metabolism. Fig. S9 FR and UV‐B supplementation affect the transcript levels of genes involved in ABA signalling. Fig. S10 QPCR of candidate low R : FR and low dose UV‐B response genes. Fig. S11 Low dose UV‐B supplementation has only minor effects on maximum photosystem II efficiency.
Table S1 A list of Arabidopsis lines used in this study.
Table S2 A list of qPCR primers used in this study.Please note: Wiley is not responsible for the content or functionality of any Supporting Information supplied by the authors. Any queries (other than missing material) should be directed to the New Phytologist Central Office.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aliniaeifard S , Van Meeteren U . 2013. Can prolonged exposure to low VPD disturb the ABA signalling in stomatal guard cells? Journal of Experimental Botany 64: 3551–3566.23956410 10.1093/jxb/ert 192PMC 3745724 · doi ↗ · pubmed ↗
- 2An Y‐Y , Li J , Feng Y‐X , Sun Z‐M , Li Z‐Q , Wang X‐T , Zhang M‐X , He J‐M . 2022. COP 1 mediates dark‐induced stomatal closure by suppressing FT, TSF and SOC 1 expression to promote NO accumulation in Arabidopsis guard cells. International Journal of Molecular Sciences 23: 15037.36499365 10.3390/ijms 232315037 PMC 9736015 · doi ↗ · pubmed ↗
- 3Aukerman MJ , Hirschfeld M , Wester L , Weaver M , Clack T , Amasino RM , Sharrock RA . 1997. A deletion in the PHYD gene of the Arabidopsis Wassilewskija ecotype defines a role for phytochrome D in red/far‐red light sensing. Plant Cell 9: 1317–1326.9286109 10.1105/tpc.9.8.1317 PMC 157000 · doi ↗ · pubmed ↗
- 4Ballare CL , Scopel AL , Sanchez RA . 1990. Far‐red radiation reflected from adjacent leaves: an early signal of competition in plant canopies. Science 247: 329–332.17735851 10.1126/science.247.4940.329 · doi ↗ · pubmed ↗
- 5Brini F , Mseddi K , Brestic M , Landi M . 2022. Hormone‐mediated plant responses to light quality and quantity. Environmental and Experimental Botany 202: 105026.
- 6Brown BA , Cloix C , Jiang GH , Kaiserli E , Herzyk P , Kliebenstein DJ , Jenkins GI . 2005. A UV‐B‐specific signaling component orchestrates plant UV protection. Proceedings of the National Academy of Sciences, USA 102: 18225–18230.10.1073/pnas.0507187102 PMC 131239716330762 · doi ↗ · pubmed ↗
- 7Brown BA , Jenkins GI . 2008. UV‐B signaling pathways with different fluence‐rate response profiles are distinguished in mature Arabidopsis leaf tissue by requirement for UVR 8, HY 5, and HYH. Plant Physiology 146: 323–324.10.1104/pp.107.108456 PMC 224585018055587 · doi ↗ · pubmed ↗
- 8Burko Y , Willige BC , Seluzicki A , Novák O , Ljung K , Chory J . 2022. PIF 7 is a master regulator of thermomorphogenesis in shade. Nature Communications 13: 4942.10.1038/s 41467-022-32585-6PMC 942423836038577 · doi ↗ · pubmed ↗