Acclimation lags in alpine grasslands reveal early warning signs of climate change
Karl Andraczek

Abstract
This article is a Commentary on Bektaş et al. (2026), 249: 1173–1187.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
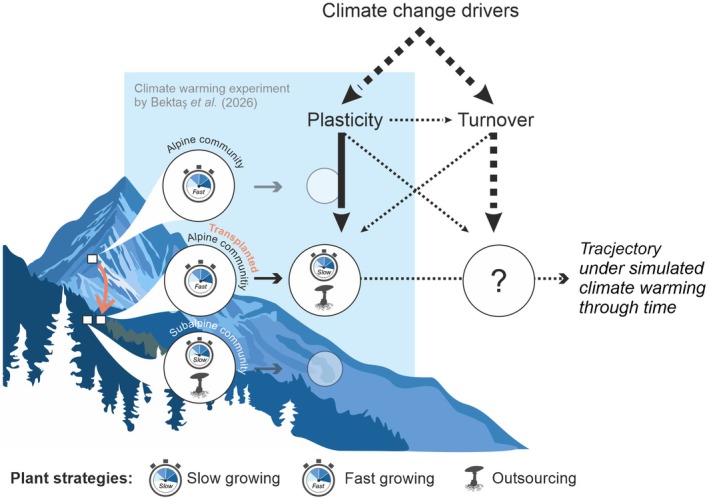 Figure 1
Figure 1- —German Research Foundation (DFG)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTree-ring climate responses · Geology and Paleoclimatology Research · Climate change and permafrost
Climate is a strong evolutionary driver of ecosystem structure and function – yet, it is changing at an unprecedented pace. This trend is especially pronounced in alpine ecosystems (Pepin et al., 2022), triggering a cascade of plant responses. In the short term, plants adjust to novel conditions through phenotypic plasticity, for example by investing in more drought‐resistant tissue. However, in the long term, insufficient adaptation may lead to species turnover. The time lag between these two states – an acclimation lag – is crucial for understanding how efficiently communities adjust to novel climates (Bektaş et al., 2021). Despite the rapid pace of warming in alpine regions (Myers‐Smith et al., 2019), acclimation lags are frequently observed. Yet, the extent to which they are driven by early shifts in plant functional strategies, and how these shifts cascade to ecosystem functioning, remains poorly understood. This is particularly concerning, as changes in ecosystem functioning can feed back onto the plant community, potentially accelerating alpine community change (Bardgett & Wardle, 2010). …these findings reveal short‐term acclimation lags in both traits and ecosystem functioning – an early signal of potential maladaptation.
Plant functional traits help identify ecological trade‐offs above‐ and belowground (Weigelt et al., 2021) while providing mechanistic insights into how shifts in plant strategies scale up to ecosystem functions. In an article published in this issue of New Phytologist, Bektaş et al. (2026; pp. 1173–1187) built on this framework to investigate how short‐term changes in plant strategies mediated early vegetation responses in herbaceous alpine communities to climate change, and how these responses affected ecosystem productivity and litter decomposition. To do so, the authors conducted a 5‐yr transplantation experiment (500‐m downslope) simulating climate warming (corresponding to an increase of 3°C and a 30% longer growing season compared with alpine conditions). In an exceptionally holistic functional approach, they measured key aboveground traits (leaf mass per area, leaf nitrogen and plant height) and belowground traits (root nitrogen, root tissue density, root diameter and specific root length) at the community level, and assessed early vegetation shifts under experimental warming. This allowed the authors to capture key plant strategies above‐ and belowground, such as the fast–slow gradient of resource conservation and the fungal collaboration gradient (outsourcing vs do‐it‐yourself). In doing so, Bektaş et al. (2026) show that alpine communities can rapidly shift their functional strategies, yet some essential traits reveal persistent acclimation lags (Fig. 1). If some traits and processes adjust while others do not, what does this mean for the short‐term trajectories of alpine plant communities under climate change?
Lifting alpine plant communities into novel climates
Climate warming increases temperature and growing‐season length – two major constraints on the growth and survival of alpine plant communities – thus driving shifts in both community composition and ecosystem functioning (Stemkovski et al., 2025). Two contrasting short‐term responses are often expected, largely reflecting changes in species abundance and plasticity. First, in response to climate warming and longer growing seasons, plants may adopt more acquisitive, fast‐growth strategies, typically linked to higher productivity. Alternatively, if warming intensifies water limitation, strategies may shift toward more conservative, stress‐tolerant traits that enhance drought resistance (Myers‐Smith et al., 2019). Bektaş et al. (2026) show that these trajectories are not mutually exclusive. Warmed alpine communities increased both productivity and litter decomposition, while their strategies shifted from fast to slow and outsourcing. Although this may appear counterintuitive – since slow and outsourcing strategies are usually associated with lower productivity – the authors attribute this to contrasting downslope environmental factors: greater energy input boosted productivity, while increased drought induced plastic shifts toward drought resistance. Thus, warmed alpine communities acclimated to novel climates relative to resident subalpine communities. Interestingly, Bektaş et al. (2026) found two key exceptions, which may be pivotal to understanding the short‐term trajectories of alpine communities under climate warming. First, while most traits acclimated to subalpine conditions, one key trait – vegetative height – did not. This suggests that plant species within alpine communities may fail to fully adjust to the novel climates, increasing the risk of species turnover via immigration of taller, more competitive subalpine species. Second, warmed alpine communities showed increased belowground productivity, exceeding that of subalpine communities. Together, these findings reveal short‐term acclimation lags in both traits and ecosystem functioning – an early signal of potential maladaptation.
The next frontiers in understanding acclimation lags
To better understand these acclimation lags, Bektaş et al. (2026) advocate for future research on their key underlying mechanisms – plasticity and species turnover. Plants can adjust their phenotype to novel conditions, but such plasticity has evolutionary limits. If these limits are exceeded, species turnover may become inevitable (Stemkovski et al., 2025). Hence, the transition from early vegetation responses due to plastic responses, up to potential tipping points of species turnover, warrants deeper investigation. Inspired by the article by Bektaş et al. (2026), I propose key future directions that may lie at the frontier of advancing this goal.
Distinguishing plasticity from changes in species abundance and turnover
One challenge is to tease apart plastic responses from changes in species abundance and turnover (Fig. 1). To address this challenge, hyperspectral and high‐resolution 3D imagery may help the scientific community to overcome logistical constraints of quantifying aboveground traits for long‐term experiments. However, these methods are less precise in capturing short‐term plastic responses, which may be relatively subtle. An alternative could be phytometer plants (e.g. see De Giorgi et al., 2025), which allow the capture of plastic responses of standardized plant species. Species turnover in the warmed alpine communities studied in Bektaş et al. (2026) was a minor factor, possibly because of the relatively short duration of the experiment (5 yr). Yet, while one might predict that turnover simply requires more time, another interesting possibility that warrants deeper investigation is whether plasticity has delayed turnover, for example by enhancing the performance of dominant species (Valladares et al., 2014; Stemkovski et al., 2025).
Identifying which traits best predict community trajectories
Another key direction is to determine which functional traits best predict the short‐ and long‐term dynamics of alpine communities under climate change (Fig. 1). While plant traits are essential for capturing ecological strategies, selecting the most relevant ones remains challenging (Weigelt et al., 2023). Bektaş et al. (2026) offer a valuable template for future studies embracing the complexity of whole‐plant strategies, inviting the scientific community to go beyond the traditionally used traits (Matthus et al., 2025). For example, long‐term growth and regeneration may depend strongly on clonal traits (Dolezal et al., 2020), while rooting depth could be key for capturing impacts of receding permafrost (Blume‐Werry et al., 2019). Such traits may also determine the likelihood of turnover as well as vegetation–climate feedbacks – for instance, traits linked to leaf albedo can influence surface temperature and snow cover, potentially buffering climate change impacts.
Disentangling the multiple drivers of vegetation change
A third challenge is to evaluate the relative contribution of different climate drivers, such as increased growing‐season length vs drought, as causal factors to vegetation change (Fig. 1). The experiment by Bektaş et al. (2026) is crucial for generating new hypotheses about alpine community trajectories under climate change. Future experimental studies will help us to disentangle these climate change drivers, while long‐term observations offer power to explore additional drivers and their attribution (Dudney et al., 2025) on the plasticity and turnover of alpine communities, and potential cascading effects on ecosystem functioning. In both cases, the scientific community should embrace robust statistical models (e.g. fixed‐effects panel regressions from fields outside ecology) to account for confounding factors, such as human impacts, when explaining community trajectories and their effects on ecosystem functioning (Schrodt et al., 2025; Siegel & Dee, 2025).
Conclusion
Bektaş et al. (2026) embraced the difficult but essential challenge of elucidating the transient dynamics of alpine plant communities under climate change. Their work sparks numerous interesting questions: how do short‐term acclimatization mechanisms interact with long‐term processes in shaping community trajectories? Can the observed decoupling between traits and ecosystem functioning be generalized across other alpine regions and beyond? Which traits beyond those currently studied are most critical for predicting ecosystem responses under climate change? Joint experimental networks (Bektaş et al., 2024), combined with mechanistic modeling of climate drivers (Dudney et al., 2025) and their impacts on ecosystem functioning via plasticity and turnover, will be among the frontiers of this research (Myers‐Smith et al., 2019).
Disclaimer
The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bardgett RD , Wardle DA . 2010. Aboveground‐belowground linkages: biotic interactions, ecosystem processes, and global change. Oxford, UK: Oxford University Press.
- 2Bektaş B , Chisholm C , Egelkraut D , Lynn J , Block S , Deola T , Dommanget F , Enquist BJ , Goldberg DE , Haider S et al. 2024. Colonization and extinction lags drive non‐linear responses to warming in mountain plant communities across the Northern Hemisphere. Ecography: e 07378. doi: 10.1111/ecog.07378. · doi ↗
- 3Bektaş B , Rutten G , Saillard A , Friaud R , Arnoldi C , Renaud J , Guéguen M , Foulquier A , Poulenard J , Lyautey E et al. 2026. Traits and functions of alpine plant communities respond strongly but not always sufficiently to in situ climate change. New Phytologist 249: 1173–1187.10.1111/nph.70503 PMC 1278031640878748 · doi ↗ · pubmed ↗
- 4Bektaş B , Thuiller W , Saillard A , Choler P , Renaud J , Colace M‐P , Della Vedova R , Münkemüller T . 2021. Lags in phenological acclimation of mountain grasslands after recent warming. Journal of Ecology 109: 3396–3410.
- 5Blume‐Werry G , Milbau A , Teuber LM , Johansson M , Dorrepaal E . 2019. Dwelling in the deep – strongly increased root growth and rooting depth enhance plant interactions with thawing permafrost soil. New Phytologist 223: 1328–1339.31074867 10.1111/nph.15903 · doi ↗ · pubmed ↗
- 6De Giorgi F , Durka W , Huang Y , Schmid B , Roscher C . 2025. Selection and phenotypic plasticity shape plant performance in a grassland biodiversity experiment. Ecology and Evolution 15: e 71117.40083731 10.1002/ece 3.71117 PMC 11904805 · doi ↗ · pubmed ↗
- 7Dolezal J , Kurnotova M , Stastna P , Klimesova J . 2020. Alpine plant growth and reproduction dynamics in a warmer world. New Phytologist 228: 1295–1305.32632948 10.1111/nph.16790 · doi ↗ · pubmed ↗
- 8Dudney J , Dee LE , Heilmayr R , Byrnes J , Siegel K . 2025. A causal inference framework for climate change attribution in ecology. Ecology Letters 28: e 70192.40808550 10.1111/ele.70192 · doi ↗ · pubmed ↗