Cis-regulatory fragments from the dissatisfaction gene identify novel mating behavior neurons in female Drosophila
Julia A Diamandi, Kara E Miller, Troy R Shirangi

TL;DR
This study identifies specific neurons in female fruit flies that control mating behaviors, using genetic tools to isolate and activate these neurons.
Contribution
The paper introduces novel cis-regulatory enhancers from the dissatisfaction gene to target and study specific female mating behavior neurons in Drosophila.
Findings
Cis-regulatory enhancers within the dsf gene regulate expression in subsets of dsf-expressing neurons.
Optogenetic activation of a specific DDAG neuron subtype triggers vaginal plate opening in female Drosophila.
The study provides new genetic tools to investigate neural circuits underlying female mating decisions.
Abstract
During Drosophila courtship, males chase and sing to females, while females perform abdominal behaviors to indicate their willingness to mate. The nerve cord circuits in females that produce their abdominal behaviors are poorly characterized. We recently identified an anatomically diverse population of abdominal interneurons called the dissatisfaction (dsf)- and doublesex-expressing abdominal ganglion (DDAG) neurons that influence several female mating behaviors. Here, we searched the dsf locus for cis-regulatory enhancer fragments that regulate its spatial expression in the adult and larval central nervous system. We found several enhancers, most located within 2 introns, that drove reporter expression in subsets of dsf-expressing neurons throughout the brain and nerve cord. Using one of these enhancers, we genetically isolated a single subtype of female-specific DDAG local…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
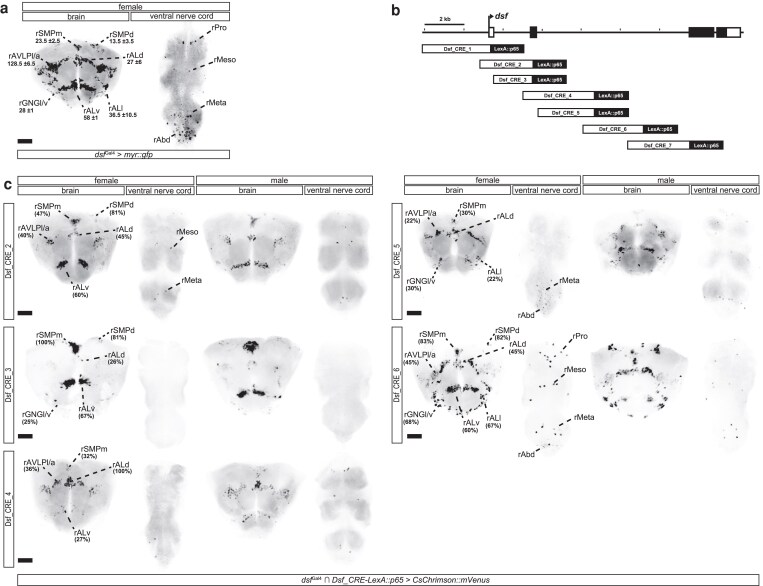 Fig. 1
Fig. 1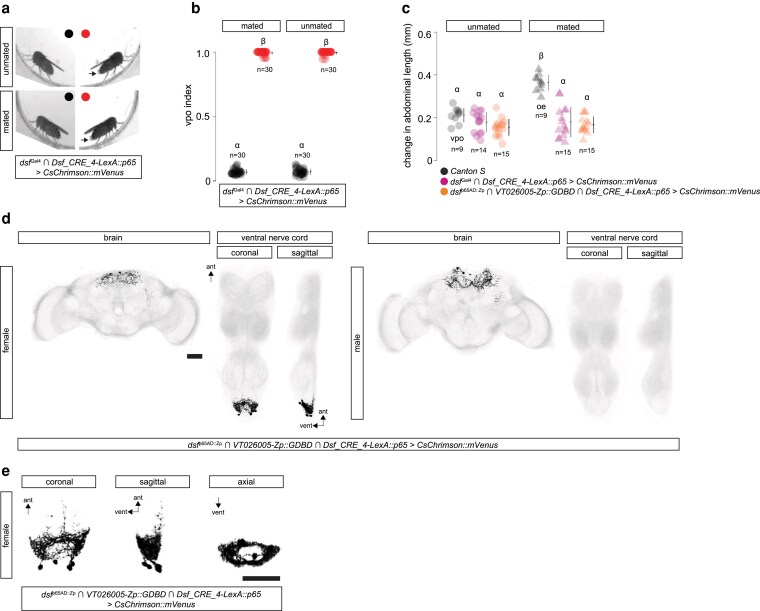 Fig. 2
Fig. 2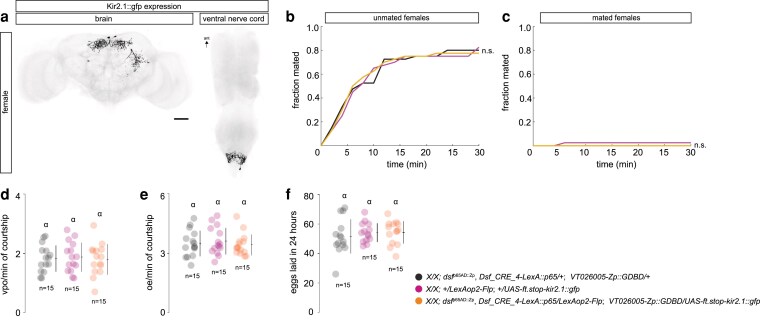 Fig. 3
Fig. 3- —National Science Foundation10.13039/501100008982
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurobiology and Insect Physiology Research · Developmental Biology and Gene Regulation · Animal Behavior and Reproduction
Introduction
Females across animal clades often perform intricate motor behaviors to signal their mating decisions to courting males. In Drosophila, females respond to a male's courtship song with subtle abdominal behaviors that convey either acceptance or rejection, depending on their mating status (Aranha and Vasconcelos 2018). Unmated females are generally receptive and indicate their willingness to mate by slowing down, spreading their vaginal plates, exposing their ovipositor and permitting copulation (Mezzera et al. 2020 ; Wang et al. 2021). After mating, Drosophila females become unreceptive for several days, until they run out of sperm. During this period, mated females still open their vaginal plates in response to male courtship, but subsequently fully extrude their ovipositor, a behavior that signals rejection (Mezzera et al. 2020; Wang et al. 2020).
In both female and male flies, courtship behaviors are largely innate and emerge during metamorphosis through the actions of 2 key sexual differentiation genes, doublesex (dsx) and fruitless (fru) (Oren-Suissa and Shirangi 2025). These genes have been instrumental in uncovering the neural circuits governing courtship behaviors in both sexes. The circuitry in the ventral nerve cord that controls male courtship song, for example, has been elucidated in remarkable detail (Lillvis et al. 2024). In contrast, the neural circuits driving the abdominal motor behaviors of females during courtship remain less well understood.
We recently identified a small, sexually dimorphic population of interneurons in the adult abdominal ganglion—termed the DDAG neurons—that co-express dsx and the Tlx/tailless-like nuclear receptor dissatisfaction (dsf) (Duckhorn et al. 2022). These neurons are anatomically diverse, comprising at least 5 subtypes that influence various female mating behaviors, including vaginal plate opening in unmated females and ovipositor extrusion and egg-laying in mated females (Duckhorn et al. 2022; Diamandi et al. 2024). In previous work, we found that 2 of these subtypes, DDAG_C and DDAG_D, are specifically involved in ovipositor extrusion (Diamandi et al. 2024). The behavioral contributions of the remaining DDAG neurons, however, remain unclear.
In this study, we searched the dsf locus for enhancer elements that drive reporter expression in subsets of dsf-expressing neurons in both the larval and adult central nervous system (CNS). We identified 1 enhancer fragment that enabled genetic access specifically to the DDAG_B neurons. Optogenetic activation of these neurons triggered vaginal plate opening in females regardless of their mating status, suggesting that DDAG_B neurons are part of the motor circuitry controlling this behavior. However, silencing DDAG_B neurons had no detectable effect on female courtship behaviors, implying that other circuit elements may compensate for their loss. Our findings provide new genetic tools for accessing dsf-expressing neurons and offer novel insights into the abdominal circuits that regulate female mating behaviors.
Methods and materials
Fly stocks
Drosophila melanogaster stocks were maintained on standard cornmeal and molasses food at 25 °C and 50% humidity in a 12-h light/dark cycle unless otherwise noted. Fly stocks used in this study were as follows: Canton S, dsf^Gal4^/CyO (Duckhorn et al. 2022), dsf^p65AD::Zp^/CyO (Diamandi et al. 2024), pJFRC79-8XLexAop2-FlpL (attP40), pJFRC41-10XUAS-FRT-STOP-FRT-myr::gfp (su(Hw)attP1), pJFRC56-10XUAS-FRT-STOP-FRT-kir2.1::gfp (attP2), 20XUAS-FRT-STOP-FRT-CsChrimson::mVenus (VK5), and VT026005-Zp::GDBD (attP2).
Construction of Dsf_CRE transgenes
All regulatory fragments from dsf were cloned into pENTR/D and then Gateway cloned into pBPnlsLexA::p65Uw (Addgene Plasmid number: 26230). All Dsf_CRE-LexA::p65 transgenes were inserted into the attP40 landing site. Forward and reverse primers used to amplify each fragment were the following:
Dsf_CRE_1:
5′-CACCTAACTCAATTCCCCAATTCTATCCAAGG-3′
5′-TGACGGACCAACTGCGAATGAAAC-3′
Dsf_CRE_2:
5′-CACCTCATTTGCTGTAATCGCATCAGGCC-3′
5′-CCTTGCAGGGAATGTCCAACAGGC-3′
Dsf_CRE_3:
5′-CACCAGCGAATCGCGTCGGCATCTGAAC-3′
5′-CCTTGCAGGGAATGTCCAACAGGC-3′
Dsf_CRE_4:
5′-CACCAACACAGGGGAATGCTCTATTCAAC-3′
5′-TTGACAAGTGCCGAGTGTTGAACTTA-3′
Dsf_CRE_5:
5′-CACCAACACAGGGGAATGCTCTATTCAAC-3′
5′-TTCCAGTCGGCCATGAACAAGGATG-3′
Dsf_CRE_6:
5′-CACCTCTCGCCGTGGGAACTTGCCAGCTG-3′
5′-ATCAGCATGCCACCGTACTTTAGAGC-3′
Dsf_CRE_7:
5′-CACCTGCAACTTGGGTTTCCTAGGACCGC-3′
5′-AGGAGTCAAGCAGCAAATCGATGG-3′
In situ hybridization chain reaction
Nervous systems were dissected in 1× phosphate-buffered saline (PBS) and fixed in 4% paraformaldehyde in PBS for 35 min at room temperature, then rinsed 3 times in PBS with 1% Triton X-100 (PBT) and washed in PBT for 20 min. The HCR™ Gold RNA-FISH Kit (Molecular Instruments) was used to perform in situ hybridization chain reaction (HCR) as previously described (Duckhorn et al. 2022). HiFi Probe Hybridization Buffer was pre-warmed to 37 °C for 15 to 20 min before use. Tissues were rinsed 1 to 2 times in pre-warmed HiFi Probe Hybridization Buffer, then incubated for 30 min at 37 °C in fresh buffer. 1 μL of HCR™ HiFi Probe (GFP X1) was added to 250 μL Probe Hybridization Buffer, and tissues were incubated overnight (∼16 h) at 37 °C. The following day, tissues were washed 4 times for 15 min at 37 °C in pre-warmed 1× HiFi Probe Wash Buffer and 3 times for 5 min in 5× SSCT (UltraPure 20× SSC Buffer, Invitrogen #15557044, diluted in water) at room temperature. Samples were then pre-amplified in Gold Amplifier Buffer for 30 min at room temperature. 4 μL of hairpins (X2 647 HCR™ Gold Amplifier) were snap cooled for 30 min in the dark before being adding 100 μL of Amplifier Buffer to make the final Hairpin solution. Tissues were incubated in the Hairpin solution overnight at room temperature, protected from light. On the third day, tissues were washed twice for 5 min, twice for 30 min, and once for 5 min in 5× SSCT at room temperature, then rinsed 3 times in PBT and washed for 20 min. Samples were mounted in VectaShield (Vector Laboratories, Cat. #H-1000-10) and imaged on a Leica TCS SP8 Confocal Microscope at 40× magnification.
Immunohistochemistry
Immunohistochemistry was performed as previously described (Duckhorn et al. 2022). Briefly, nervous systems were dissected in 1× PBS, fixed in 4% paraformaldehyde in PBS for 35 min, rinsed and washed in PBT. We did not block. Tissues were incubated with primary antibodies diluted in PBT at 4 °C. Three washes were performed the next day over several hours before nervous systems were incubated overnight at 4 °C in secondary antibodies also diluted in PBT. Tissues were then washed 3 times over the course of several hours and placed on cover slips coated in poly-lysine, dehydrated in an increasing ethanol concentration series, and cleared in a xylene series. Nervous systems were mounted onto slides using DPX mounting medium and imaged on a Leica TCS SP8 Confocal Microscope at 40 × magnification. The following primary antibodies were used: rabbit anti-GFP (Invitrogen #A11122; 1:1000), and rat anti-DN-cadherin (DN-Ex#8, Developmental Studies Hybridoma Bank; 1:50). The following secondary antibodies were used: Fluorescein (FITC) conjugated donkey anti-rabbit (Jackson ImmunoResearch #711-095-152; 1:500), and AF-647 goat anti-rat (Invitrogen #A21247; 1:500).
Optogenetics and behavioral assays
Optogenetics and behavioral assays was performed as previously described (Duckhorn et al. 2022). Briefly, unmated females used in optogenetic assays were raised in darkness and on food containing 0.2 mM all-trans-retinal (sigma-Aldrich #R2500) and were incubated at 25 °C and 50% humidity. Females were grouped in vials consisting of 15 to 20 flies for 8 to 12 d before testing. Flies were anesthetized on ice for 2 min, decapitated under low-intensity light, and given 15 to 20 min to recover before being transferred to individual behavioral chambers (diameter: 10 mm, height: 3 mm). A FLIR Blackfly S USB3, BFS-U3-31S4M-C camera with a 800 nm long-pass filter (Thorlabs, FEL0800) was used to record optogenetic videos in SpinView. Chambers were placed on top of an LED panel with continuous infrared (850 nm) light and recurring photoactivating red (635 nm) light using an Arduino script. To measure change in abdominal length before and during vaginal plate opening or ovipositor extrusion, a ruler (cm/mm) was included in the frame to set the scale. The change in abdominal length was calculated as the difference in the abdominal length from the base of the scutellum to the tip of the abdomen before and during photoactivation. Behavior indices were measured by calculating the average fraction of time spent performing the behavior during the first three 15-s lights-on periods and the first three 45-s light-off periods. For courtship assays, unmated females and males were collected under CO_2_ and aged for 7 to 10 d in a 12-h light/dark cycle and incubated at 25 °C and 50% humidity. Unmated females were group-housed in vials consisting of 15 to 20 flies, and Canton S males were individually housed. Courtship assays were done within the first 2 h of the subjective day. Unmated females and Canton S males were transferred to individual behavioral chambers (diameter: 10 mm, height: 3 mm) and recorded for 30 min using a Sony Vixia HFR700 video camera at 25 °C under white light. For experiments using mated females, unmated females were housed with males for 24 h, anesthetized on ice for 2 min, and mated females were collected into a new vial and given 30 min to recover. Mated females and Canton S males were loaded to chambers and recorded as described above. Vaginal plate opening and ovipositor extrusion frequency was measured as the total number of times a female performed the behavior in a 6-min period of active male courtship. Egg laying was measured by allowing females to mate with males before transferring them to individual vials for 24 h. The total number of eggs laid in 24 h by each female was then counted.
Results and discussion
The dsf gene is expressed in ∼0.5% of neurons in each hemisphere of the adult CNS, organized in ∼11 sex-shared groups of cells in the brain and nerve cord (Fig. 1a) (Duckhorn et al. 2022). One of these groups, rAbd, contain the DDAG neurons, consisting of eleven cells in females that contribute to several reproductive behaviors (Duckhorn et al. 2022). We previously showed that the DDAG neurons fall into 5 anatomical subtypes, A to E, and that the DDAG_C and DDAG_D subtypes regulate a specific mating behavior in mated females, i.e. ovipositor extrusion (Diamandi et al. 2024). To determine how other DDAG subtypes and other dsf+ neuronal groups contribute to behavior, we sought to develop reagents that provide genetic access to subsets of dsf-expressing neurons.
cis-regulatory fragments from the dsf gene label subsets of dsf-expressing neurons in the adult CNS. a) Confocal images of the adult brain and ventral nerve cord from a dsfGal4 > UAS-myr::gfp female showing expression of GFP mRNA (black) labeled by in situ Hybridization Chain Reaction. DsfGal4 is expressed in ∼11 groups of neurons in the adult brain and ventral nerve cord. Dsf-expressing neuronal groups were categorized according to standardized nomenclatures (Duckhorn et al. 2022). The average number ( standard deviation) of neurons within each cluster is shown. b) An illustration showing the location of regulatory fragments from the dsf gene that were analyzed in this study. c) The intersection of dsfGal4 and most Dsf_CRE-LexA::p65 transgene targets various subsets of dsf-expressing neurons in the brain and ventral nerve cord of adult females and males. Fragments 1 and 7 failed to generate any clear HCR signal and were thus not included here. Fragment 1 did, however, label a single rALv neuron by immunohistochemistry (Supplementary Fig. 1). The approximate percentage of neurons labeled within each cluster is shown in females. GFP mRNA is shown in black. Scale bar = 50 μm.
The noncoding regions of pleiotropic genes in metazoans often contain multiple, modular transcriptional cis-regulatory elements (CREs), i.e. enhancers, that direct gene expression in distinct tissues or cells. We systematically surveyed the dsf gene for enhancers that drive reporter expression in subsets of dsf-expressing cells in the CNS. We analyzed dsf using 7 partially overlapping 2 to 3-kb fragments (designated as Dsf_CRE_1–7) that cover upstream and intronic regions of the locus (Fig. 1b). Transgenic fly stocks were generated carrying each of the 7 fragments attached to a heterologous promoter and the coding sequence for the transcriptional activator, LexA::p65.
To determine whether these fragments drive LexA::p65 expression in dsf-expressing neurons, we genetically intersected each Dsf_CRE-LexA::p65 transgene with our dsf^Gal4^ allele, which faithfully recapitulates the endogenous expression of dsf (Duckhorn et al. 2022). In this system, LexA::p65, expressed in cells targeted by the regulatory fragment, drives the expression of a LexAop-regulated Flp recombinase, which excises a transcriptional stop cassette from a UAS-regulated myr::gfp transgene. Expression of GFP is then driven by dsf^Gal4^. We probed GFP mRNA expression in the adult CNS by in situ HCR (Fig. 1c), and protein expression in the adult and larval CNS by immunohistochemistry (Supplementary Figs. 1, 2). Much of dsf's transcriptional cis-regulatory content maps to the 5′ portion of its large third intron. Most regulatory fragments in our analysis were active in subsets of dsf-expressing neurons in the CNS. All eleven groups of dsf-expressing neurons in the brain and nerve cord were labeled by 1 or more enhancers, and some enhancers targeted subsets of neuronal groups or subsets of cells within specific neuronal groups or both. Of the 7 fragments, 4 (Dsf_CRE_2, 4, 5, and 6) had activity in subsets of female-specific DDAG neurons (Supplementary Fig. 1). We focused our attention on Dsf_CRE_4, as this fragment labeled 3 to 4 female-specific DDAG neurons in each hemisphere of the abdominal ganglion and an otherwise sparse number of dsf-expressing neurons in the nerve cord.
Unmated Drosophila females indicate their receptivity to courting males with a behavior called “vaginal plate opening,” or VPO, in which they open their vaginal plates and partially expose their ovipositor (Wang et al. 2021). Mated females, however, reject courting males by fully extruding their ovipositor, a behavior called “ovipositor extrusion,” or OE, which may block copulation or male courtship drive (Wang et al. 2020). The length of the abdomen increases during both behaviors, but the change in abdominal length is greater during an OE than during a VPO (Wang et al. 2020; Duckhorn et al. 2022). We previously showed that optogenetic activation of all DDAG neurons using the mVenus-tagged, red-light-gated cation channelrhodopsin, CsChrimson::mVenus (Klapoetke et al. 2014), caused unmated females to open their vaginal plates and mated females to extrude their ovipositors (Duckhorn et al. 2022). Upon photoactivation, dsf^Gal4^ Dsf_CRE_4-LexA::p65 > CsChrimson::mVenus females opened their vaginal plates regardless of their mating status (Fig. 2a to c; Supplementary Video 1, 2). VPO was penetrant and occurred largely during the photoactivation period (Fig. 2b).
Optogenetic activation of the DDAB_B neurons opens the vaginal plates of adult females. a) Still-frames of decapitated dsfGal4 ∩ Dsf_CRE_4-LexA::p65 > CsChrimson::mVenus unmated (top) and mated (bottom) females before (left) and during (right) a bout of red light (10.5 mW/mm2). Arrows point to the exposed ovipositor during a VPO. b) Average fraction of time females open their vaginal plates during (red) and between (black) three 15-s photoactivation bouts in a row, i.e., VPO index. Lights are off for 45 s between bouts. It takes a few seconds for females to retract the ovipositor once the lights turn off, so indices are slightly above zero during lights off. n = number of females. c) Photoactivation of DDAG_B neurons induces VPO in unmated and mated females. The change in abdominal length when an unmated (black circles) or mated (black triangles) Canton S female performs VPO or OE, respectively, during courtship is shown. Photoactivation of the DDAG_B neurons in dsfGal4 ∩ Dsf_CRE_4-LexA::p65 > CsChrimson::mVenus (magenta) and Dsf_CRE_4-LexA::p65 ∩ dsfp65AD::Zp ∩ VT026005-Zp::GDBD > CsChrimson::mVenus (orange) unmated (circles) and mated (triangles) females induces a similar change in abdominal length to Canton S females performing a VPO. n = number of females. d) Confocal images of a Dsf_CRE_4-LexA::p65 ∩ dsfp65AD::Zp ∩ VT026005-Zp::GDBD > CsChrimson::mVenus female and male CNS. GFP-expressing neurons and DNCad (neuropil) are shown in black and light gray, respectively. Scale bar = 50 μm. e) Anatomy of the DDAG_B neurons from a Dsf_CRE_4-LexA::p65 ∩ dsfp65AD::Zp ∩ VT026005-Zp::GDBD > CsChrimson::mVenus female. Scale bar = 50 μm.
The female-specific neurons labeled by dsf^Gal4^ Dsf_CRE_4-LexA::p65 appeared to correspond to a single anatomical cholinergic subtype we previously identified called the DDAG_B neurons (Diamandi et al. 2024). We previously developed a split-gal4 driver, dsf^p65AD::Zp^ VT026005-Zp::GDBD, that labeled the DDAG_C, DDAG_D, and DDAG_B neurons in adult females (Diamandi et al. 2024). To confirm that the DDAG neurons labeled by Dsf_CRE_4 are the DDAG_B neurons, we genetically intersected Dsf_CRE_4-LexA::p65 with dsf^p65AD::Zp^ VT026005-Zp::GDBD. Indeed, this three-way intersection specifically labeled 3 to 4 female-specific DDAG_B neurons and no other neurons in the nerve cord (Fig. 2d). The DDAG_B neurons are female-specific interneurons that arborize locally in the abdominal neuropil, forming a hollow circle in an axial view (Fig. 2e). Photoactivation of Dsf_CRE_4-LexA::p65 dsf^p65AD::Zp^ VT026005-Zp::GDBD > CsChrimson::mVenus females evoked VPO behaviors (Fig. 2b, c; Supplementary Video 3) that were qualitatively and quantitatively similar to what we observed when we photoactivated dsf^Gal4^ Dsf_CRE_4-LexA::p65 > CsChrimson::mVenus females.
To determine whether the activity of the DDAG_B neurons labeled by Dsf_CRE_4-LexA::p65 dsf^p65AD::Zp^ VT026005-Zp::GDBD is required for VPO behavior in unmated females during courtship, we used the three-way intersection to drive the expression of a GFP-tagged inwardly rectifying K+ channel, Kir2.1::gfp (Baines et al. 2001) (Fig. 3a). We observed no effect on female courtship behaviors from the expression of Kir2.1::gfp in the DDAG_B neurons. Unmated and mated females expressing Kir2.1::gfp in the DDAG_B neurons copulated with males at a rate that was similar to control females (Fig. 3b, c); they opened their vaginal plates and extruded their ovipositors during courtship at a frequency comparable to controls (Fig. 3d, e); and mated Dsf_CRE_4-LexA::p65 dsf^p65AD::Zp^ VT026005-Zp::GDBD > Kir2.1::gfp females laid a number of eggs similar to controls (Fig. 3f). These data suggest the involvement of additional neural circuit elements for VPO in the abdominal ganglion that may compensate for the absence of DDAG_B activity. Alternatively, the DDAG_B neurons may not have been fully silenced, or additional DDAG_B neurons may exist that were not targeted by our labeling strategy.
Expression of Kir2.1::GFP in the DDAG_B neurons has no effect on female courtship behaviors. a) Confocal image of a Dsf_CRE_4-LexA::p65 ∩ dsfp65AD::Zp ∩ VT026005-Zp::GDBD > kir2.1::gfp female brain and ventral nerve cord. Kir2.1::GFP-expressing neurons are in black. b, c Fraction of unmated (b) and mated (c) females that mated with naive Canton S males in 30 min. Log rank test for significance. n.s. = not significant. n = 40 females for each genotype. d) Number of VPOs an unmated female performed per minute of active courtship. e) Number of OEs a mated female performed per minute of active courtship. f) Number of eggs laid 24-h post-mating. d to f) Individual points, mean, and SD. n = number of females. A one-way ANOVA Tukey-Kramer multiple comparison test measured significance (P < 0.05). Same letter indicates no significant difference.
The DDAG neurons are an anatomically diverse population consisting of at least 5 subtypes that collectively influence multiple female mating behaviors (Duckhorn et al. 2022; Diamandi et al. 2024). We previously showed that the DDAG_C and DDAG_D neurons regulate OE in mated females (Diamandi et al. 2024). Here, we found that the DDAG_B neurons likely contribute to VPO in unmated females. Our results suggest that the motor circuits for VPO and OE in the abdominal ganglion of females may include neurons with functions specific to each behavior. The EM connectome data of the female and male ventral nerve cord (Marin et al. 2024; Stürner et al. 2025) will provide important insights into the position of these neurons in the abdominal circuits that modulate female abdominal behaviors during courtship.
The DDAG neurons are all recycled from a population of dsf-expressing interneurons in the larval abdominal ganglion (Diamandi et al. 2024). The DDAG_C and DDAG_D neurons are segmental homologs of a single, sex-shared larval interneuron type called A26 g (Diamandi et al. 2024), whose function is currently unknown. During early metamorphosis, A26 g neurons at abdominal segments A5 and A6 acquire dsx expression, undergo programmed cell death in males, and get repurposed in females for functions in ovipositor extrusion (Diamandi et al. 2024). What about the DDAG_B neurons? In future studies, it will be fascinating to identify the larval counterparts of the DDAG_B neurons and determine how they contribute to larval life.
Supplementary Material
jkaf249_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aranha MM, Vasconcelos ML. 2018. Deciphering Drosophila female innate behaviors. Curr Opin Neurobiol. 52:139–148. 10.1016/j.conb.2018.06.005.29940518 · doi ↗ · pubmed ↗
- 2Baines RA, Uhler JP, Thompson A, Sweeney ST, Bate M. 2001. Altered electrical properties in Drosophila neurons developing without synaptic transmission. J Neurosci. 21:1523–1531. 10.1523/JNEUROSCI.21-05-01523.2001.11222642 PMC 6762927 · doi ↗ · pubmed ↗
- 3Diamandi JA et al 2024. Developmental remodeling repurposes larval neurons for sexual behaviors in adult Drosophila. Curr Biol. 34:1183–1193.e 3. 10.1016/j.cub.2024.01.065.38377996 · doi ↗ · pubmed ↗
- 4Duckhorn JC et al 2022. Regulation of Drosophila courtship behavior by the Tlx/tailless-like nuclear receptor, dissatisfaction. Curr Biol. 32:1703–1714.e 3. 10.1016/j.cub.2022.02.031.35245457 · doi ↗ · pubmed ↗
- 5Klapoetke NC et al 2014. Independent optical excitation of distinct neural populations. Nat Methods. 11:338–346. 10.1038/nmeth.2836.24509633 PMC 3943671 · doi ↗ · pubmed ↗
- 6Lillvis JL et al 2024. Nested neural circuits generate distinct acoustic signals during Drosophila courtship. Curr Biol. 34:808–824.e 6. 10.1016/j.cub.2024.01.015.38295797 · doi ↗ · pubmed ↗
- 7Marin EC et al 2024. Systematic annotation of a complete adult male Drosophila nerve cord connectome reveals principles of functional organisation. e Life. 13:1–466. 10.7554/e Life.97766.1. https://elifesciences.org/reviewed-preprints/97766. Accessed 15 August 2025.. · doi ↗
- 8Mezzera C et al 2020. Ovipositor extrusion promotes the transition from courtship to copulation and signals female acceptance in Drosophila melanogaster. Curr Biol. 30:3736–3748.e 5. 10.1016/j.cub.2020.06.071.32795437 · doi ↗ · pubmed ↗