Formate as electron carrier in the gut acetogen Blautia luti: a model for electron transfer in the gut microbiome
Raphael Trischler, Volker Müller

TL;DR
This study explores how Blautia luti, a gut bacterium, uses formate to transfer electrons during metabolism, shedding light on gut microbiome functions.
Contribution
The study reveals a novel role of formate as an intraspecies electron carrier in gut acetogens.
Findings
Blautia luti produces acetate, succinate, lactate, formate, and hydrogen from sugar fermentation.
Formate is generated via pyruvate-formate lyase and used as an electron acceptor in CO2 fixation.
Formate connects oxidative and reductive branches of glucose metabolism in gut acetogens.
Abstract
Species of the genus Blautia are commonly found in the human gut and are known to be beneficial for the human well-being. However, only little is known about the physiology and the specific role of Blautia species in the human gut. In this study, we investigated the heterotrophic metabolism of the formate dehydrogenase lacking gut acetogen Blautia luti. We identified acetate, succinate, lactate, formate, and hydrogen as end products of sugar fermentation. Interestingly, formate is produced by the pyruvate-formate lyase reaction and used as electron acceptor in the Wood–Ljungdahl pathway of CO2 fixation. Thus, formate connects the oxidative branch of glucose metabolism with the reductive branch. The use of formate as an intraspecies electron carrier seems to be common in gut acetogens. This study highlights the role of formate as electron carrier in the gut microbiome and improves our…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
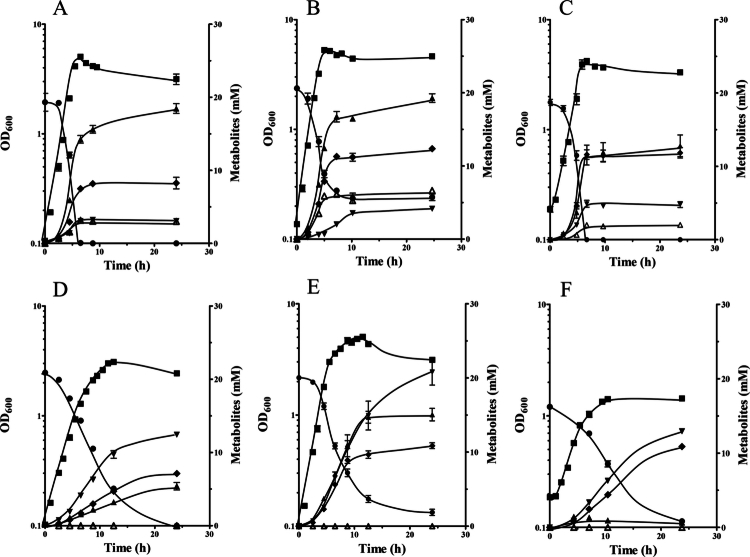 Figure 1
Figure 1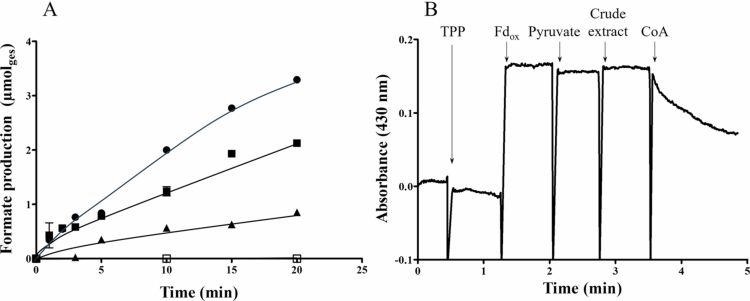 Figure 2
Figure 2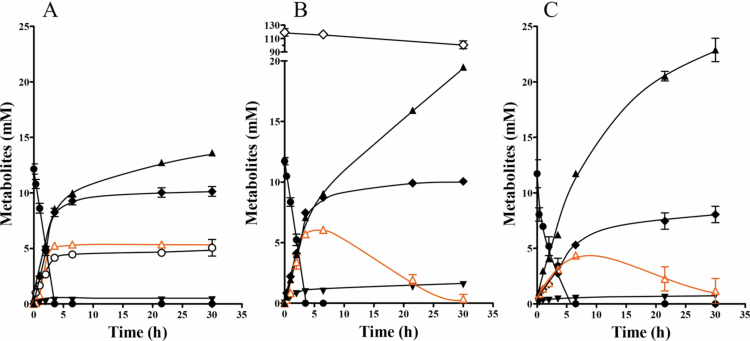 Figure 3
Figure 3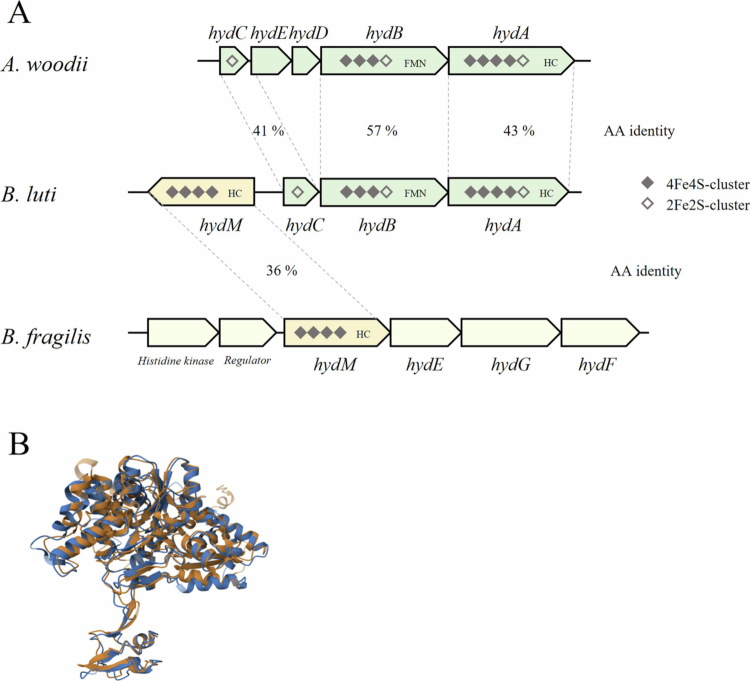 Figure 4
Figure 4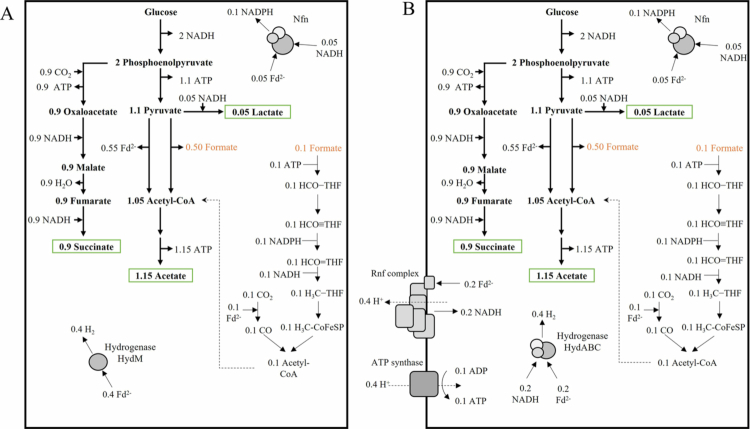 Figure 5
Figure 5| Xylose C5H10O5 | Arabinose C5H10O5 | Glucose C6H12O6 | Sorbitol C6H14O6 | Sucrose C12H22O11 | Trehalose C12H22O11 | Maltose C12H22O11 | Raffinose C18H32O16 | ||
|---|---|---|---|---|---|---|---|---|---|
| Fermentation balance (metabolite/carbohydrate) | |||||||||
| Acetate | 1.39 ± 0.09 | 1.09 ± 0.03 | 0.97 ± 0.20 | 0.67 ± 0.18 | 1.50 ± 0.09 | 1.57 ± 0.21 | 1.29 ± 0.04 | 2.08 ± 0.3 | |
| Succinate | 0.26 ± 0.01 | 0.29 ± 0.01 | 0.42 ± 0.04 | 0.62 ± 0.10 | 0.79 ± 0.02 | 0.96 ± 0.16 | 0.83 ± 0.01 | 1.11 ± 0.12 | |
| Lactate | 0.05 ± 0.01 | 0.02 ± 0.01 | 0.16 ± 0.01 | 0.25 ± 0.04 | 0.32 ± 0.01 | 0.02 ± 0.01 | 0.28 ± 0.01 | 0.49 ± 0.07 | |
| Formate | 0.09 ± 0.02 | 0.28 ± 0.01 | 0.15 ± 0.01 | 0.10 ± 0.02 | 0.31 ± 0.02 | 0.72 ± 0.16 | 0.45 ± 0.01 | 0.38 ± 0.05 | |
| H2 | 0.19 ± 0.04 | 0.26 ± 0.04 | 0.17 ± 0.03 | 0.29 ± 0.19 | 0.95 ± 0.20 | 0.68 ± 0.18 | 0.63 ± 0.08 | 1.29 ± 0.14 | |
| Carbon balance | |||||||||
| CAcetate (%) | 55.4 ± 3.5 | 43.4 ± 1.4 | 32.5 ± 6.8 | 22.2 ± 5.9 | 25.1 ± 1.5 | 26.2 ± 3.4 | 21.6 ± 0.7 | 23.1 ± 3.3 | |
| CSuccinate (%) | 21.1 ± 0.8 | 23.2 ± 0.6 | 28.3 ± 2.7 | 41.5 ± 7.0 | 26.3 ± 0.5 | 32.0 ± 5.5 | 27.7 ± 0.4 | 24.6 ± 2.6 | |
| CLactate (%) | 2.7 ± 0.8 | 1.5 ± 0.1 | 7.9 ± 0.3 | 12.5 ± 2.2 | 8.1 ± 0.2 | 0.4 ± 0.1 | 7.0 ± 0.4 | 8.2 ± 1.2 | |
| CFormate (%) | 1.9 ± 0.4 | 5.6 ± 0.1 | 2.5 ± 0.2 | 1.7 ± 0.4 | 2.6 ± 0.1 | 6.0 ± 1.4 | 3.7 ± 0.1 | 2.1 ± 0.3 | |
| Growth parameters | |||||||||
| Growth rate (h−1) | 0.64 ± 0.01 | 0.65 ± 0.01 | 0.67 ± 0.01 | 0.59 ± 0.05 | 0.64 ± 0.01 | 0.58 ± 0.02 | 0.77 ± 0.03 | 0.77 ± 0.03 | |
| Final OD600 | 3.59 ± 0.69 | 3.34 ± 0.47 | 5.05 ± 0.13 | 4.37 ± 0.36 | 7.33 ± 0.17 | 2.99 ± 0.04 | 5.29 ± 0.21 | 6.52 ± 0.70 | |
| Enzyme activity | Substrates | Specific activity (mU mg−1) | |||||
|---|---|---|---|---|---|---|---|
| Glucose | Sorbitol | Maltose | Glucose + Phosphinate | Sorbitol + Phosphinate | Maltose + Phosphinate | ||
| PFOR | Fdox + Pyruvate + CoA + TPP | 1162 ± 129 | 3648 ± 382 | 1452 ± 268 | 2300 ± 182 | 5882 ± 605 | 2106 ± 122 |
| PFL | Pyruvate + CoA | 292 ± 69 | 230 ± 13 | 575 ± 49 | 0.0 | 0.0 | 0.0 |
| Lactate dehydrogenase | Pyruvate + NADH | 209 ± 16 | 289 ± 54 | 324 ± 90 | 246 ± 18 | 235 ± 27 | 295 ± 28 |
| Fumarate reductase | Fumarate + MVred | 504 ± 53 | 880 ± 44 | 852 ± 22 | 682 ± 54 | 989 ± 15 | 695 ± 110 |
| Malate dehydrogenase | Oxaloacetate + NADH | 1070 ± 256 | 923 ± 141 | 668 ± 67 | 690 ± 193 | 2839 ± 246 | 496 ± 90 |
| Hydrogenase | H2 + MV | 290 ± 33 | 644 ± 37 | 19 ± 0.3 | 131 ± 9 | 492 ± 39 | 14 ± 3 |
- —European Research Council (ERC) under the European Union's Horizon 2020 research and innovation program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial metabolism and enzyme function · Anaerobic Digestion and Biogas Production · Gut microbiota and health
Introduction
The human gut is a highly complex ecosystem that consists of trillions of microorganisms.1 This community is made of mainly bacteria, but also archaea, viruses, and eukaryotic cells.2 The composition of the microbiome and the presence of specific microbes in the gut is not only important for the breakdown of organic matter, it also impacts human health as well as the development of diseases. Since many different factors such as diet, pharmaceuticals, geography, stress, lifestyle, and many more shape the gut microbiome, each human being is the architect of its own gut microbiome.3-6 One taxon of gut bacteria that typically correlates to human well-being is the genus Blautia, especially due to the production of short-chained fatty acids and succinate, its biotransformation and antibacterial activity.7^,^8
Species of the genus Blautia typically belong to the nonphylogenetic group of acetogenic bacteria, which are defined as strictly anaerobic bacteria that can reduce two mol of CO_2_ by the Wood–Ljungdahl pathway (WLP) to acetyl-CoA. Most acetogens produce acetate from acetyl-CoA via acetyl phosphate, since this reaction results in the production of ATP.9-12 The WLP is divided into two branches: The methyl-branch and the carbonyl-branch. In the methyl-branch, CO_2_ is first reduced to formate which is then subsequently further reduced to a methyl group. In the carbonyl-branch, CO_2_ is reduced to enzyme-bound CO, which is then condensed with the methyl group and CoA to acetyl-CoA. This reaction and the production of CO are catalyzed by the bifunctional CO dehydrogenase/acetyl-CoA synthase complex (CODH/ACS).13-15 The CODH/ACS functions as a key enzyme of the WLP, since it combines the two WLP branches and catalyzes the reaction in the WLP with the highest thermodynamic barrier, the reduction of CO_2_ to CO.16 This reaction can only be catalyzed with reduced ferredoxin as an electron donor due to the negative redox potential of CO_2_. In general, acetogens can grow autotrophically on H_2_ and CO_2_ by utilization of the WLP, which results in the production of acetate as end product.17^,^18 Under this condition H_2_ functions as electron donor for the reduction of electron carriers used by the WLP. However, the redox potential of H_2_ (E_0_′ = −414 mV)19 is too low for reduction of ferredoxin (E_0_′ = −400 to −500 mV).16^,^20 To overcome this barrier, acetogens utilize the mechanism of electron bifurcation.21^,^22 The electron-bifurcating hydrogenase utilizes H_2_ as electron donor for the exergonic reduction of NADH. The energy of this reaction is used to drive the endergonic reduction of ferredoxin.23^,^24 The electron carriers used in the methyl-branch can differ between acetogens. Therefore, many acetogens harbor an electron-bifurcating Nfn or Stn complex to convert NADH and ferredoxin to NADPH.25^,^26 The reduction of CO_2_ to acetate by the WLP results in a net ATP formation of zero, since one ATP is consumed in the methyl-branch and one ATP is produced by the acetate kinase reaction.10 An additional energy-conserving mechanism has to be used for acetogenesis from H_2_ + CO_2_. To date, two respiratory enzyme complexes have been found in acetogens, which produce an ion/proton gradient across the membrane as fuel for ATP synthase for ATP production, the ferredoxin:NAD^+^ oxidoreductase (Rnf) and the ferredoxin:H^+^ oxidoreductase (Ech) complex.16
Acetogens are not limited to the utilization of H_2_ + CO_2_ as growth substrates, they can use various substrates for organoheterotrophic growth such as different sugars, alcohols, carboxylic acids, aldehydes, amino acids or methylated compounds.9^,^16^,^27-37 The flexibility in the usage of different growth substrates gives acetogens an enormous ecological advantage, which can be of great importance for interspecies interactions in the human gut. In addition, the oxidation of carbohydrates is directly linked to the WLP as electron sink. During this process, the respiratory enzymes (Rnf or Ech) lead to additional ATP production, resulting in an energetic advantage of acetogens in contrast to other fermenters.10^,^16
Recently, Blautia strains with an unusual WLP were identified.38 The genome of these Blautia strains does encode for all WLP enzymes except for a formate dehydrogenase, which catalyzes the reduction of CO_2_ to formate as the first reaction of the methyl-branch in the WLP. In addition, it was experimentally proven that cells of the formate dehydrogenase lacking Blautia group were able to perform acetogenesis from CO and formate, demonstrating an unusual but active WLP even in the absence of a formate dehydrogenase.38 One of these strains is the gut acetogen Blautia luti. However, little is known about the physiology of this bacterium and how the WLP interacts with the oxidative branch of its metabolism. This study revealed that B. luti produces acetate, succinate, lactate, formate, and H_2_ as end products during fermentation of various carbohydrates. Formate is produced during heterotrophic metabolism by pyruvate-formate lyase and can subsequently be used as an electron acceptor in the methyl-branch of the WLP instead of CO_2_. This feature of gut acetogens helps to further understand the interactions of different gut bacteria and their role in human well-being. In addition, this study is of interest for biotechnological applications since it also introduces B. luti as a bacterium producing the industrial desired compound succinate.
Material and methods
Organism and cultivation
B. luti DSM14534 was cultivated at 37 °C in CO_2_/KHCO_3_-buffered complex medium.39 For cultivation under CO_2_/KHCO_3_-limited conditions medium was used as described.40 As carbon and energy source 20 mM of glucose, xylose, arabinose, sorbitol, sucrose, trehalose, maltose, or raffinose were used. In addition, some cultures grew in presence of 75 mM Na^+^-phosphinate. Growth was monitored by measuring the optical density at 600 nm (OD_600_).
Preparation of resting cells
B. luti DSM14534 was grown in CO_2_/KHCO_3_-buffered complex medium with 20 mM glucose as carbon and energy source to late exponential growth phase. Resting cells were prepared as described.38 The protein concentration was measured as described.41
Cell suspension experiments
Cell suspension experiments were carried out at 37 °C in 115-ml serum flasks, which contained 10 ml imidazole buffer (50 mM imidazole, 20 mM KCl, 20 mM NaCl, 20 mM MgSO_4_, 2 mM DTE, 4.4 µM resazurin, pH 7.0) with 60 mM KHCO_3_ under a N_2_/CO_2_ [80:20 (v:v)] atmosphere or in buffer without KHCO_3_ under a N_2_ atmosphere. Resting cells were resuspended to a total protein concentration of 1 mg/ml. In each experiment, glucose (10 mM) was used as carbon source. For the mixotrophic fermentation experiments 1 bar 100% H_2_ + CO_2_ [80:20 (v/v)] or 20% of CO were added to the atmosphere.
Determination of metabolites
The concentrations of the metabolites acetate, formate, lactate, and succinate as well as the concentrations of different sugars (glucose, xylose, arabinose, sucrose, trehalose, maltose, raffinose, and sorbitol) were determined by high-performance liquid chromatography as described.34^,^40 The concentrations of H_2_42 and CO43 were measured by gas chromatography as described.
Preparation of cell-free extract
For preparation of cell-free extract B. luti was cultivated with 20 mM glucose, sorbitol or maltose to mid exponential growth phase (OD_600_ = 2). Cell-free extract was prepared as described.38 The protein concentration was measured according to Bradford.44
Enzyme assays
The determination of enzyme activities in cell-free extract was performed under strictly anoxic conditions at 37 °C in glass cuvettes (d = 0.2 cm; Glasgerätebau Ochs, Germany) in Tris-buffer (100 mM Tris-HCl, 2 mM DTE, 4.4 µM resazurin, pH 7.5). For measurement of lactate dehydrogenase activity 0.25 mM NADH were used as electron donor and 20 mM pyruvate as electron acceptor. The oxidation of NADH was determined at 340 nm. The determination of malate dehydrogenase activity was performed similar to the lactate dehydrogenase assay, but with 5 mM oxaloacetate as electron acceptor. For fumarate reductase activity 10 mM methylviologen, previously reduced by addition of sodium dithionite, were used for reduction of 10 mM fumarate. The oxidation of methylviologen was measured at 604 nm. To determine pyruvate formate lyase activity in Tris-buffer (100 mM Tris-HCl, 20 mM MgSO_4_, pH 7.5, 2 mM DTE, 4.4 µM resazurin, pH 7.5) formate production from 10 mM pyruvate in presence of 1.75 mM CoA was measured over time using the “formic acid assay kit” from R-Biopharm AG (Darmstadt, Germany). Pyruvate ferredoxin oxidoreductase was measured in the same buffer in presence of 100 µM thiaminpyrophosphate and 200 µM CoA by the reduction of 3 mM ferredoxin with 10 mM pyruvate as electron donor. The reaction was monitored at 430 nm.
Analysis of transcript abundance by semiquantitative PCR
B. luti was cultivated with 20 mM glucose as a carbon and energy source to mid exponential growth phase. The isolation of RNA, digestion of chromosomal DNA and synthesis of cDNA was performed as described.29 The cDNA was used as template for determination of transcript abundance by semiquantitative PCR as described.45 The primers used in this study are listed in Supplementary Table S1.
Results
Sugar fermentation by B. luti
B. luti is known to grow heterotrophically on many different sugars;46 however, growth and fermentation profiles have never been reported in the literature. When grown in CO_2_/KHCO_3_-buffered medium with 20 mM glucose as carbon and energy source, B. luti grew with a growth rate of 0.69 ± 0.01 h^−1^ to a maximal optical density of 5.05 ± 0.13 (Figure 1A). Glucose was completely consumed at the end of fermentation, the major products were acetate (18.46 ± 1.35 mM), succinate (8.26 ± 1.58 mM), lactate (3.05 ± 0.47 mM), and formate (3.01 ± 0.21 mM). Furthermore, small amounts of H_2_ were measured at the end of fermentation (Table 1). The fermentation balance was as follows:
Growth of B. luti with glucose (A, D), maltose (B, E), and sorbitol (C, F) as carbon and energy source in absence (A-C) or presence (D-F) of Na+-phosphinate. B. luti was grown at 37 °C in 115 ml serum flasks containing 50 ml CO2/KHCO3-buffered medium with 20 mM of glucose (A), maltose (B) or sorbitol (C) as substrate. In addition, B. luti was also grown in presence of 75 mM Na+-phosphinate with 20 mM of glucose (D), maltose (E), and sorbitol (F) as substrate. For inoculation (5%) a preculture without Na+-phosphinate was used that was previously adapted to the corresponding sugar. The optical densities at 600 mM (■), as well as the concentrations of each sugar (●), acetate (▲), succinate (♦), formate (Δ), and lactate (▼) were determined. Each data point are mean ± SEM; N = 3 independent experiments.
Noteworthy, B. luti was not able to grow in the absence of CO_2_/KHCO_3_ (data not shown). When grown with different carbon sources such as xylose, arabinose, sucrose, trehalose, maltose or raffinose B. luti always produced acetate as major end product, followed by succinate, lactate, and formate (Figure 1B and C; Supplementary Figure S1; Table 1). In addition, H_2_ was produced as fermentation end product from every sugar tested. Interestingly, growth on the sugar alcohol sorbitol led to production of acetate and succinate in almost equal amounts, resulting in the highest succinate production per hexose molecule (Table 1). The fermentation balance for growth of B. luti on sorbitol as carbon and energy source was as follows:
Obviously, succinate was formed instead of acetate.
Formate as a central metabolite in Blautia species lacking formate dehydrogenases
The genome of B. luti does not harbor a formate dehydrogenase-encoding gene,38 but formate can be fed into the WLP directly. Therefore, the question arises of how formate is produced during growth on all the sugars tested. Analysis of the genome of B. luti revealed the presence of two copies of a pyruvate-formate lyase (PFL)-encoding gene (Supplementary Table S2), which we suspected to catalyze formate formation. To test this hypothesis, growth on glucose in the presence of the PFL inhibitor Na^+^-phosphinate was investigated (Figure 1D). The growth rate was reduced to 0.38 ± 0.01 h^−1,^ and the fermentation profile changed dramatically. Formate was no longer produced. Furthermore, acetate (5.39 ± 0.88 mM) was produced in reduced amounts, and lactate was the major fermentation end product (12.43 ± 0.47 mM). Succinate production was only slightly decreased. The fermentation balance of glucose in the presence of Na^+^-phosphinate was as follows:
The same effect of Na^+^-phosphinate was also observed during fermentation of the disaccharide maltose (Figure 1E). In the absence of Na^+^-phosphinate, acetate (19.28 ± 1.01 mM) was the major fermentation end product, followed by succinate (12.34 ± 0.29 mM), H_2_ (9.45 ± 1.30 mM), formate (6.65 ± 0.38 mM), and lactate (4.15 ± 0.10 mM). However, in the presence of Na^+^-phosphinate, lactate was again the major fermentation product (20.80 ± 3.05 mM), followed by acetate (15.06 ± 1.45 mM), H_2_ (11.7 ± 0.7 mM), and succinate (10.86 ± 0.68 mM). Moreover, formate was not produced in the presence of Na^+^-phosphinate.
To test whether carbon flow can also be redirected to succinate production as an electron sink instead of the WLP or lactate, we tested growth of B. luti with sorbitol in the presence of Na^+^-phosphinate (Figure 1F), since this sugar led to the highest succinate production (see above). As expected, growth rate (0.38 ± 0.04 h^−1^), and final OD (1.48 ± 0.07) were highly reduced in comparison to growth without Na^+^-phosphinate (0.50 ± 0.05 h^−1^; final OD 3.60 ± 0.19). Surprisingly, while acetate was now only produced in trace amounts (0.67 ± 0.12 mM), succinate (10.87 ± 0.11), and lactate (12.94 ± 0.40 mM) were produced in almost equal amounts as the major fermentation end products. Unexpectedly, the amount of H_2_ was slightly reduced at the end of fermentation to 1.9 ± 0.1 mM. Additionally, the production of succinate was similar to Na^+^-phosphinate-free conditions, revealing that lactate production is the preferred electron sink in the absence of the WLP.
These results are in line with the hypothesis that PFL is used during heterotrophic growth for pyruvate oxidation. The most important observation is the absence of formate production during heterotrophic fermentation in presence of Na^+^-phosphinate, but also the increased lactate and decreased acetate production is in line with the use of PFL. We postulate, that formate, which is produced by PFL, is used by B. luti as electron acceptor in the WLP to produce additional acetate. As compensation for the loss of formate by inhibition of PFL, pyruvate is further metabolized to lactate as electron sink alternative to the WLP. However, since acetate production is not completely abolished, additional acetyl-CoA forming enzymes such as the pyruvate-ferredoxin-oxidoreductase (PFOR) must be active.
Enzyme activities in cells grown under different conditions
To verify, whether PFL or PFOR are both active during heterotrophic metabolism, we measured both activities in crude extract of B. luti grown on glucose (Figure 2, Table 2). As expected, crude extract of B. luti was able to produce formate form pyruvate in the presence of CoA with an activity of 292 ± 69 mU mg^−1^, confirming an active PFL (Figure 2A). Without addition of CoA, formate was not produced. Additionally, we measured PFOR activity. Indeed, crude extract of B. luti was also able to reduce ferredoxin with pyruvate as substrate in presence of CoA with an activity of 1019 ± 235 mU mg^−1^ (Figure 2B). Ferredoxin could not be reduced in the absence of CoA. Furthermore, we were able to measure a NADH-dependent lactate dehydrogenase activity (NADH:pyruvate oxidoreductase activity) of 209 ± 16 mU mg^−1^. These results reveal that pyruvate is metabolized in B. luti via three routes: First, it can be used as substrate by PFL for production of acetyl-CoA and formate. The latter can than be used by the WLP as electron acceptor but also for cross-feeding in the gut. Next, pyruvate can also be used by PFOR for production of acetyl-CoA and reduction of ferredoxin. Reduced ferredoxin can be used in the WLP but also as source for molecular hydrogen. And last, pyruvate can be reduced to lactate.
Measurement of PFL (A) and PFOR activity (B) in crude extract of B. luti grown on 20 mM glucose. (A) PFL activity was determined by formate production from 10 mM pyruvate and 1.75 mM CoA with 0.75 (●), 0.5 (■), and 0.25 mg ml−1 (▲) crude extract. In addition, formate production from pyruvate with 0.5 mg ml−1 (□) crude extract in absence of CoA was determined. Each data point are mean ± SEM; N = 2 independent experiments. (B) For measurement of PFOR activity the reduction of 3 mM ferredoxin with 10 mM pyruvate, 200 µM CoA, and 100 µM thiamine pyrophosphate (TPP) as substrates was monitored at 430 nm. Both reactions were performed in Tris-buffer (100 mM Tris-HCl, 20 mM MgSO4, pH 7.5, 2 mM DTE, 4.4 µM resazurin, pH 7.5) at 37 °C.
Interestingly, addition of Na^+^-phosphinate had stronger effects on the production of acetate during growth on sorbitol than on acetate production from glucose or maltose (see above), indicating that PFL or PFOR might be differentially produced during growth on these sugars. Unexpectedly, PFL activity was almost similar in crude extract of B. luti grown on all of these sugars (Table 2). However, PFOR activity was approximately three times increased in crude extract of cells previously grown on sorbitol (3648 ± 382 mU mg^−1^) in comparison to crude extract of glucose (1019 ± 235 mU mg^−1^) and maltose (1452 ± 268 mU mg^−1^) grown cells (Table 2). The NADH:pyruvate oxidoreductase activity was comparable in crude extract of glucose, maltose, and sorbitol grown cells (192 ± 22 mU mg^−1^; 323 ± 90 mU mg^−1^; 289 ± 54 mU mg^−1^). Due to inhibition of PFL by Na^+^-phosphinate pyruvate can only be metabolized via the production of lactate or the PFOR reaction. Since the oxidation of the sugar alcohol sorbitol already leads to production of more reducing equivalents than the oxidation of glucose, the abolishment of acetate production and switch to lactate production might be a protective mechanism against accumulation of reducing power.
In addition, crude extract of cells cultivated with glucose, sorbitol or maltose in presence of Na^+^-phosphinate showed no PFL activity, but PFOR activity was highly increased by approximately 98%, 61%, and 45%, respectively, in comparison to crude extract of cells cultivated in absence of Na^+^-phosphinate. Additionally, the cultivation in presence of Na^+^-phosphinate had no effect on lactate dehydrogenase activity. A comparable effect was observed by cultivation of the gut bacterium Roseburia intestinalis, which uses PFOR and PFL simultaneously during heterotrophic fermentation at high iron concentration, but switches from PFOR usage to lactate dehydrogenase utilization at low iron concentrations.47 Therefore, a switch between PFL, PFOR, and lactate dehydrogenase at different growth conditions might also be a common feature for gut acetogens.
To further reconstruct the pathway for the heterotrophic metabolism of B. luti, the route for succinate production was investigated next. The generation of succinate is possible via three metabolic routes: The ATP-generating carboxylation of phosphoenolpyruvate (PEP) to oxaloacetate by PEP carboxykinase, the carboxylation of pyruvate to oxaloacetate by pyruvate carboxykinase or the carboxylation of pyruvate to malate catalyzed by the decarboxylating malate dehydrogenase. Of these three, only the PEP carboxykinase and the decarboxylating malate dehydrogenase encoding genes are present in the genome of B. luti (Supplementary Table S2). We were able to measure a NADH-dependent reduction of oxaloacetate to malate in crude extract of B. luti grown on glucose with an activity of 1070 ± 256 mU mg^−1^, which is in line with the hypothesis that oxaloacetate is an intermediate of glucose fermentation in B. luti. The MDH activity was comparable in crude extract of maltose- (668 ± 67 mU mg^−1^) and sorbitol- (923 ± 141 mU mg^−1^) grown cells. Malate is then further metabolized to fumarate and finally reduced to succinate. Crude extract reduced fumarate with reduced methylviologen as electron donor with an activity of 499 ± 79 mU mg^−1^. Interestingly, crude extract of B. luti grown on maltose or sorbitol showed an almost two times higher fumarate reductase activity (Table 2). Na^+^-phosphinate did not affect fumarate reductase activity. Interestingly, addition of Na^+^-phosphinate to cells cultivated with glucose led to a decrease in malate dehydrogenase activity while the activity was approximately three times higher in sorbitol-grown cells. This might indicate that in absence of a functional WLP, succinate production might be used as alternative electron sink during fermentation of sorbitol, second to lactate production. The H_2_:MV-oxidoreductase activity measured in all crude extracts tested, was relatively low (approximately 640 to 20 mU mg^−1^) in comparison to other acetogens like Blautia schinkii with approximately 23,000 mU mg^−1^.38 In crude extract of B. luti cultivated in presence of Na^+^-phosphinate, hydrogenase activity was reduced by approximately 30%–50%. These results are in line with the reduced H_2_ production of growing cells of B. luti in the presence of Na^+^-phosphinate (see above). The absence of formate production by PFL and increased PFOR activity due to the addition of Na^+^-phosphinate suggests increased production of reduced ferredoxin under these conditions. However, without formate the WLP cannot be used as electron sink. Therefore, an alternative electron sink is needed. Unexpectedly, lactate production seems to be the favored as alternative electron sink to the WLP instead of H_2_ production, which is commonly found in acetogens. The production of lactate might even be favored in terms of energetics, since the interconversion of reduced ferredoxin to NADH is performed via the energy-conserving Rnf reaction. In sum, the data demonstrates that PFL plays a central role in the heterotrophic metabolism of Blautia species lacking formate dehydrogenases such as B. luti.
Formate as a transient intermediate during mixotrophic fermentation
To further investigate carbon flow during glucose fermentation in the absence of biomass production, resting cells experiments were performed. B. luti was grown in CO_2_/KHCO_3_-buffered medium with 20 mM glucose as carbon- and energy source. Cells were harvested, washed, and resuspended in imidazole buffer. In buffer containing CO_2_/KHCO_3_ cells fermented glucose (12.15 ± 0.66 mM) to 13.62 ± 0.33 mM acetate, 10.12 ± 0.63 mM succinate, 5.23 ± 0.24 mM formate and 5.05 ± 1.06 mM H_2_. Only trace amounts of lactate (0.36 ± 0.05 mM) were produced (Figure 3A). A carbon balance of 101,6% ± 0.6% and an electron balance of 94.5% ± 0.1% were calculated. The fermentation balance was as follows:
Fermentation of glucose in the presence of hydrogen or carbon dioxide by the resting cells of B. luti. B. luti was cultivated at 37 °C in CO2/KHCO3-buffered medium with 20 mM glucose as substrate to late exponential growth phase, harvested, washed, and resuspended in 10 ml imidazole buffer (50 mM imidazole, 60 mM KHCO3, 20 mM NaCl, 20 mM MgSO4, 2 mM DET, 4.4 µM resazurin, pH 7.0) to a final protein concentration of 1 mg ml−1 within 115 ml serum flasks. The resting cells were incubated under a N2/CO2 atmosphere [80%/20% (v/v)] with 10 mM glucose (A), with 10 mM glucose in the presence of 20% CO in the gas phase (B) and with 10 mM glucose and an atmosphere of 1 bar H2 + CO2 (C). At each timepoint, the concentrations of glucose (●), acetate (▲), succinate (♦), formate (Δ), lactate (▼), and H2 (○) were determined. Each data point is the mean ± SEM; N = 2 independent experiments.
Interestingly, resting cells of B. luti were not able to ferment glucose in the absence of CO_2_/KHCO_3_ (Supplementary Figure S2), similar to growing cells. CO_2_ seems to play an essential role during sugar fermentation by B. luti. To be more specific, CO_2_ might not only be essential for the use of the WLP but might also be required to produce succinate from phosphoenolpyruvate since that pathway requires a carboxylation reaction.
To further study the role of the WLP as well as the role of formate during heterotrophic metabolism, fermentation of glucose by resting cells of B. luti in the presence of external electron donors was investigated, which might be a more accurate reflection of the gut environment. Since B. luti can produce acetate from formate and CO via the WLP,38 glucose fermentation in the presence of similar CO concentrations (20% CO) used by Trischler et al.38 was investigated first (Figure 3B). As expected, at the end of fermentation, the acetate level increased to 19.49 ± 0.40 mM. In addition, during the first seven hours of fermentation, approximately 5 mM formate was produced, but afterwards, it was completely consumed. The addition of CO did not affect the production of succinate; however, lactate levels slightly increased to 1.54 ± 0.08 mM. In total, a consumption of 18.5 ± 0.5 mM CO was measured. H_2_ was not produced. These products contained 78.2% ± 2.3% of the carbon and 99.0% ± 3.3% of the electrons from the utilized glucose and CO. The fermentation balance was as follows:
When the cells were incubated with glucose under an atmosphere of 1 bar overpressure of H_2_ + CO_2_ (Figure 3C), the fermentation profile was similar to that of fermentation with additional CO. Not only was the production of acetate increased to 22.88 ± 1.48 mM, but formate was also first produced and afterwards almost completely consumed. The level of succinate and lactate were only slightly affected. The fermentation balance was as follows:
In the gut colon, bacteria most likely encounter mixotrophic conditions. We speculate that in FDH-lacking gut acetogens, formate serves as an intraspecies as well as interspecies electron carrier and that formate reduction via the WLP can be commonly used as an electron sink.
Hydrogenases of B. luti
Since the resting cells of B. luti were able to utilize H_2_ as electron donor and growing cells produced H_2_ as fermentation end product, we further investigated the types of hydrogenases present in B. luti. Inspection of the genome sequence of B. luti revealed two hydrogenase-encoding genes hydA and hydM, which are located in close proximity to each other in the genome (Figure 4A). The gene hydA is part of a gene cluster with the genes hydB and hydC, similar to the electron-bifurcating hydrogenase-encoding gene cluster of Acetobacterium woodii. Comparison of the corresponding hydrogenase subunits revealed amino acid identities between 41% and 57%. In addition, the binding motifs for all cofactors of the A. woodii electron-binding hydrogenase complexes are conserved in the corresponding gene products of B. luti (Supplementary Figures S3, S4, S5).
Comparison of the genetic organization and postulated cofactor composition of the hydrogenases of B. luti. (A) The genome of B. luti harbors a gene cluster consisting of genes encoding the electron-binding hydrogenase (hydABC) and group B [FeFe]-hydrogenase (hydM). The hydrogenase-encoding genes of B. luti were compared with the electron-bifurcating hydrogenase complex encoding genes (hydABC) of A. woodii as well as the postulated ferredoxin-dependent hydrogenase-encoding gene (hydM) of B. fragilis. (B) Comparison of the predicted structure of the HydM of B. luti (orange) and B. fragilis (blue). Structure predictions were performed by RCSB protein Data Bank.48
In sharp contrast, hydM does not form a complex with other genes (Figure 4A). Further analysis of this gene revealed sequence motifs for four 4Fe-4S cluster and one H-cluster. The presence of a single hydrogenase subunit and composition of 4Fe-4S cluster led to the speculation that hydM encodes a group B [FeFe]-hydrogenase. Comparison of HydM to the group B [FeFe]-hydrogenase of B. fragilis49 lead to an amino acid identity of 36% (Figure 4A). However, on structural level both hydrogenases were almost identical (Figure 4B). Especially the typical formation of two distinct globular domains as well as the sequence motive for the H-cluster and four [4Fe4S] clusters are conserved. While the H-cluster and two [4Fe4S] clusters are present in the catalytic domain, the other two [4Fe4S] clusters are located in the smaller ferredoxin-like domain, which is typical for group B [FeFe]-hydrogenases. These hydrogenases are postulated to catalyze the production of H_2_ with reduced ferredoxin as an electron donor.49 Furthermore, we were able to identify the presence of HydM in several Blautia species, with amino acid identities between 71.9% and 99.8% (Supplementary Figures S6, S7).
Reconstruction of the heterotrophic glucose fermentation pathway of B. luti
Glucose is first metabolized by the Embden–Meyerhof–Parnas pathway (glycolysis). The intermediate phosphoenolpyruvate is further metabolized via oxaloacetate and malate to succinate. On the other hand, phosphoenolpyruvate is further metabolized to pyruvate. The latter can then be further metabolized to lactate or acetate by PFL or PFOR reactions as fermentation end products, as already discussed in detail. The WLP of B. luti uses a NADH-dependent methylene-THF reductase and an NADPH-dependent methylene-THF dehydrogenase.38 For NADP^+^ reduction and redox balancing, B. luti harbors genes encoding an electron-bifurcating Nfn complex. Furthermore, reduced ferredoxin, which is gained during glucose fermentation by PFOR reaction, is converted to NADH by the energy-conserving Rnf complex. This reaction leads to the establishment of a proton gradient across the membrane, which is used by ATP synthase for ATP production. To produce H_2_, two different hydrogenases can be used (see above). The postulated Fd-dependent HydM is commonly found in gut bacteria (Bacteroidetes and Firmicutes). However, the use of reduced ferredoxin as a direct electron donor for H_2_ production is a waste of energy for B. luti, since the use of ferredoxin by Rnf leads to energy conservation. A theoretical glucose fermentation pathway of B. luti that uses the Fd-dependent hydrogenase HydM and a NADH-dependent fumarate reductase would work independently of Rnf (Figure 5A). The calculated net ATP production would be 3.05 ATP/glucose. By use of the electron-bifurcating hydrogenase HydABC, only half the amount of ferredoxin would be used for hydrogen production. As a result, energy conversion by the Rnf would be possible (Figure 5B). Under these conditions, a net ATP production of 3.15 ATP/glucose can be calculated.
Biochemistry and bioenergetics of glucose fermentation by B. luti. (A) Glucose fermentation in the presence of HydM. (B) Glucose fermentation in the presence of the electron-bifurcating hydrogenase HydABC. For explanation, see text. Assumed stoichiometries: H+/ATP = 3.6 (ATP Synthase) and 2 H+/2e− (Rnf).
To tackle the question which hydrogenase is used by B. luti during growth on glucose, we analyzed transcript abundance of hydA and hydM with semiquantitative polymerase chain reaction (PCR) (Supplementary Figure S8). As a control, frdAB, encoding the fumarate reductase was amplified. frdAB was highly transcribed, which is in line with the production of succinate during growth on glucose and the enzyme measurements and reflects the high fumarate reductase activity measured in crude extract (see above). In case of the hydrogenase encoding genes, hydA was not expressed, but hydM, although expression levels were comparably low, which reflects the relatively low H_2_:MV oxidoreductase activity of crude extract of B. luti (Table 2).
Discussion
B. luti is an acetogenic bacterium38 and found to be one of the most abundant of all Blautia species in the human gut.38^,^50 Acetogens are characterized by the WLP,9^,^51^,^52 a pathway that can convert C1-compounds at different redox levels. Reductants used are molecular hydrogen or carbon monoxide. Formate is an intermediate of the WLP and therefore, many acetogens can grow on external formate. In the absence of external reductants, three quarters of the formate have to be oxidized to CO_2_ to gain the electrons to reduce the other 25% to methyl-THF.53^,^54 In the presence of an external electron donor formate is exclusively reduced to acetate. This is what we observed in our studies.
The WLP also brings an ecological advantage to acetogens: disposal of electrons from sugar oxidation with CO_2_ or formate or a methyl-group (plus CO_2_) as electron acceptor. This allows homoacetogens the complete oxidation of the hexose (by glycolysis and pyruvate oxidation) to acetate which gives the highest ATP yield possible: 4 mol of ATP per mol of hexose by substrate-level phosphorylation.10^,^55 Acetogens have an oxidative and a reductive branch in their metabolism that are connect by various electron carriers, including H_2_. Sugars are oxidized and the reduced electron carriers are used in anaerobic respiration to reduce CO_2_, formate (+CO_2_) or methyl-groups (+CO_2_) via the WLP to acetate.56 What is described here for a single organism is also found between two different organism and a classical limb in an anaerobic food web. A strict fermenter oxidizes a hexose to 2 mol of acetate, 2 mol of CO_2_, and 4 mol of H_2_, which functions as electron carrier between two organisms. The oxidation of hydrogen by methanogens, sulfate reducers or acetogens lowers the hydrogen partial pressure in the environment and allows thermodynamically unfavorable hydrogen production by the fermenter and allows the hydrogen oxidizer to make a living as well.57 However, the free energy change decreases in the order of sulfidogenesis, methanogenesis and acetogenesis, and thus, methanogens and sulfate reducers outcompete acetogens on H_2_ utilization.19^,^58 Nevertheless, acetogens such as species of the genus Blautia can utilize an enormous range of carbohydrates and many other substrates (see above). Therefore, acetogens do not necessarily have to compete with methanogens or sulfate reducers for hydrogen to thrive in the gut. As shown here, the gut representative B. luti can grow on a number of different sugars that are oxidized via pyruvate as an intermediate, and electrons are channeled to the WLP but also to the production of reduced compounds such as succinate, lactate or H_2_. Pyruvate is oxidized by two different enzymes: PFL and PFOR. Whereas the former produces acetyl-CoA and formate, the latter produces acetyl-CoA and reduces ferredoxin and CO_2_. This makes a huge difference for the bacterium: formate can be directly used in the methyl-branch of the WLP as an electron acceptor, and NADH (from glycolysis) can be used directly or indirectly (converted to NADPH or reduced to ferredoxin by transhydrogenases) as an electron donor. From the data presented herein, it is clear that PFL is the major source of formate in B. luti and not formate dehydrogenase, which is lacking in B. luti. In the presence of CO or hydrogen, formate is only transiently produced from sugars and then completely reduced via the WLP to acetate. Like hydrogen, formate can also be used as an electron carrier between different species. The gut acetogen Clostridium bovifaecis also lacks a formate dehydrogenase but uses solely PFOR for pyruvate oxidation instead of PFL,59 in contrast to B. luti. Thus, formate must be provided by an external source, such as another bacterium, otherwise C. bovifaecis can no longer grow on sugars. The same effect of formate-dependent growth was observed by the genetic deletion of formate dehydrogenase/hydrogen-dependent CO_2_ reductase in the homoacetogens T. kivui or A. woodii.60^,^61 In sharp contrast, the production of formate by PFL enables B. luti to use the WLP as a reductive branch independent of formate cross-feeding. Interestingly, PFOR in addition to PFL is also present in B. luti, to produce reduced ferredoxin. This electron carrier could be reoxidized by the Rnf complex to reduce NAD^+^ and conserve additional energy or could be reoxidized by a hydrogenase (either by HydM or in combination with NADH by the electron bifurcating hydrogenase HydABC) to produce molecular hydrogen. The use of the postulated Fd-dependent hydrogenase HydM by B. luti, as our data suggest, is unexpected, since the use of the electron-bifurcating hydrogenase HydABC would provide an energetic advantage. However, the use of HydM is in line with the study of Welsh et al.49 which proposed that HydM is generally responsible for most H_2_ production in the human gut. Nevertheless, hydrogen production is only possible at low partial hydrogen concentrations since the redox potentials of ferredoxin (E_0_′ = −400 to −500 mV) and H_2_/H^+^ (E_0_′ = −414 mV) are very close.16^,^19 In addition, HydM could work as an emergency valve when the cytoplasm is too reduced. When the PFL was inhibited by phosphinate, PFOR did not step in. Instead, electrons were redirected from pyruvate to yield lactate as a major fermentation end product. Lactate formation under high electron loading was also observed in A. woodii and when the methylene-THF reductase was genetically deleted, A. woodii switched from acetogenesis to mixed acid fermentation.61^,^62
The fact that B. luti produces only minor amounts of hydrogen but can also use hydrogen as an electron donor might contribute to the finding that Blautia species are generally indicators of a healthy gut. Hydrogen is a central fermentation end product of carbohydrate breakdown in the gut microbiome. Its presence in the gut has positive and negative effects on human well-being. High hydrogen concentrations can impair the metabolism of gut bacteria,19 as already mentioned above and is connected to diseases such as carbohydrate malabsorption and inflammatory bowel disease.63-65 On the other hand, hydrogen can reduce oxidative stress and has positive effects on diseases such as obesity, inflammation, cancer or even Parkinson's disease.66-68 The finding that gut acetogens like B. luti can produce hydrogen and formate during carbohydrate fermentation but are also able to consume these compounds make gut acetogens appear in a new light. The primary role for acetogens might be the breakdown of carbohydrates to produce short-chain fatty acids, such as acetate, propionate, and butyrate, as well as lactate, succinate, formate, hydrogen and other compounds. These products might not only be directly useful for the gut epithel cells, but the fermentation end products can also be used for cross-feeding by other gut bacteria. In addition, acetogens can step in as hydrogen consumer to prevent accumulation of hydrogen in the gut, at times when methanogens or sulfate-reducing bacteria are not able to do so. In the rumen of sheep, for example, over 90% of the hydrogen is consumed by methanogens, the H_2_-utilization of acetogens is almost irrelevant. However, in the gut of methanogen-free sheep, acetogens were able to capture up to 25% of the hydrogen.69 Moreover, our study suggests that species of the genus Blautia might not only be able to prevent accumulation of H_2_ in the gut but also the accumulation of toxic amounts CO. Interestingly, CO is naturally produced by the human body in low concentrations (0.5 to 4.5 ppm CO each hour) mainly as result of heme degradation via heme oxygenase reaction.70^,^71 In low concentrations CO functions as anti-inflammatory agent,72 neurotransmitter,73^,^74 regulator of the mucosal immune response75 and has in general beneficial effects on the human well-being.71 However, due to external exposure to small amounts of CO over a longer period or due to exposure of higher concentrations in general (approximately >100 ppm), CO can also become a toxic compound due to binding and inhibition of haemoproteins.76^,^77 Interestingly, a CO concentration of 10 to 13 ppm could be measured in the flatulence of heathy people, while the CO concentration of people with gastrointestinal disorders was highly increased to 258 ppm,78 showing that the human gut microbiome contributes to CO homeostasis in the gut. We postulate that gut acetogens, especially species of the genus Blautia play a crucial role in CO uptake. In a recently published study,38 we were able to demonstrate CO utilization by the gut acetogens B. wexlerae and B. luti, that was further characterized in this study. Species of the genus Blautia devoid of a FDH can utilize CO in combination with formate to produce the short chained fatty acid acetate. These findings are further supported by the study of Katayama et al.79 which revealed that most CO dehydrogenase-encoding transcripts of the human gut microbiome belonged to species of the genus Blautia.
Another important feature of B. luti is the production of succinate as fermentation end product. In the human gut, succinate plays a major role in the maintenance of the gut homeostasis.80 On one site, the presence of succinate has many beneficial effects for human health. Succinate can help to prevent diseases such as obesity but also stimulates the host mucosal immune cells and the function of other gut microbes which leads to a healthy balance between gut bacteria and their host.81 Additionally, it can be directly used as energy source for gut epithelial cells, but can also be used by other gut microbes to produce healthy short-chain fatty acids.80^,^82 In a healthy gut, the succinate concentration is approximately 0.5 to 5 mM.83^,^84 However, gut dysbiosis is often accompanied by drastically increased succinate concentrations that can reach up to 25 mM.85 Many studies revealed that an abnormal increase of succinate levels in the gut is often accompanied with immune disorders and diseases such as inflammation and even cancer.85-87 Therefore, a balance between succinate producing bacteria such as B. luti and succinate fermenting bacteria is essential for a healthy gut.
Furthermore, succinate is a highly desirable product for biotechnology. In fact, succinate and other four carbon 1,4-diacids (fumarate and malate) are placed as number one of the 12 most valuable chemicals derived from sugars and synthesis gas.88 The biotechnologically relevant organisms such as Actinocacillus succinogenes, Anaerobiospirillium succiniproducens, Mannheimia succiniproducens, Corynebacterium glutamicum or Basfia succiniproducens can produce up to 105.8 g/l (89.5 mM) succinate mostly by sugar fermentation.89^,^90 B. succiniproducens can even produce succinate from waste glycerol.91^,^92 One of the highest succinate-yields form glucose (for wildtype organism) was reported for A. succiniciproducens with 0.97 succinate/glucose at optimal conditions.90^,^93 In this study, growing cells of B. luti achieved a yield of 0.42 succinate/glucose, which is similar to the succinate yield of A. succiniciproducens (0.41 succinate/glucose) at comparable pH.93 However, since the aim of this study was not to optimize succinate production, we do not know the true limit for succinate production by B. luti. By changing growth conditions such as pH or CO_2_ concentrations, higher succinate yields might be achieved.
In this study, we were able to elucidate novel features in the physiology of the genus Blautia, specifically for Blautia species without formate dehydrogenase. These bacteria seem to play in general a major role as a cornerstone in a healthy gut by the fermentation of carbohydrates and production of the short-chain fatty acid acetate, lactate, and succinate, but also by the production of formate and H_2_ in moderate amounts, which are central metabolites for cross-feeding in the gut. In addition, species of the genus Blautia might be able to prevent accumulation of toxic amounts of formate, CO or H_2_ in the gut. Therefore, it does not surprise that the genus Blautia is commonly associated with the human well-being.
Supplementary Material
Trischler and Mueller Supplementary Data clean version.docxTrischler and Mueller Supplementary Data clean version.docx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Turnbaugh PJ, Ley RE, Hamady M, Fraser-Liggett CM, Knight R, Gordon JI. The human microbiome project. Nature. 2007;449:804–810. doi: 10.1038/nature 06244.17943116 PMC 3709439 · doi ↗ · pubmed ↗
- 2Ursell LK, Metcalf JL, Parfrey LW, Knight R. Defining the human microbiome. Nutr Rev. 2012;70:38–44. doi: 10.1111/j.1753-4887.2012.00493.x.PMC 342629322861806 · doi ↗ · pubmed ↗
- 3Cresci GA, Bawden E. Gut microbiome: what we do and don't know. Nutr Clin Pract. 2015;30:734–746. doi: 10.1177/0884533615609899.26449893 PMC 4838018 · doi ↗ · pubmed ↗
- 4Hou K, Wu ZX, Chen XY, Wang JQ, Zhang D, Xiao C, Zhu D, Koya JB, Wei L, Li J. Microbiota in health and diseases. Signal Transduct Target Ther. 2022;7:135. doi: 10.1038/s 41392-022-00974-4.35461318 PMC 9034083 · doi ↗ · pubmed ↗
- 5Parizadeh M, Arrieta MC. The global human gut microbiome: genes, lifestyles, and diet. Trends Mol Med. 2023;29:789–801. doi: 10.1016/j.molmed.2023.07.002.37516570 · doi ↗ · pubmed ↗
- 6Van Hul M, Cani PD, Petitfils C, De Vos WM, Tilg H, El-Omar EM. What defines a healthy gut microbiome?. Gut. 2024;73:1893–1908. doi: 10.1136/gutjnl-2024-333378.39322314 PMC 11503168 · doi ↗ · pubmed ↗
- 7Holmberg SM, Feeney RH, Vishnu Prasoodanan PK, Puertolas-Balint F, Singh DK, Wongkuna S, Zandbergen L, Hauner H, Brandl B, Nieminen AI, Skurk T, et al. The gut commensal Blautia maintains colonic mucus function under low-fiber consumption through secretion of short-chain fatty acids. Nat Commun. 2024;15:3502. doi: 10.1038/s 41467-024-47594-w.38664378 PMC 11045866 · doi ↗ · pubmed ↗
- 8Liu X, Mao B, Gu J, Wu J, Cui S, Wang G, Zhao J, Zhang H, Chen W. Blautia-a new functional genus with potential probiotic properties?. Gut Microbes. 2021;13:1–21. doi: 10.1080/19490976.2021.1875796.PMC 787207733525961 · doi ↗ · pubmed ↗